")
Back to Journals » OncoTargets and Therapy » Volume 13
Exosomal Transfer of Macrophage-Derived miR-223 Confers Doxorubicin Resistance in Gastric Cancer
Authors Gao H, Ma J, Cheng Y, Zheng P
Received 23 September 2020
Accepted for publication 5 November 2020
Published 25 November 2020 Volume 2020:13 Pages 12169—12179
DOI https://doi.org/10.2147/OTT.S283542
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr William C. Cho
Huijie Gao,1 Jincheng Ma,1 Yanhui Cheng,1 Peiming Zheng2
1Department of Oncology, The First Affiliated Hospital of Henan University, Kaifeng, Henan 475000, People’s Republic of China; 2Department of Clinical Laboratory, Henan Provincial People’s Hospital, People’s Hospital of Zhengzhou University, People’s Hospital of Henan University, Zhengzhou, Henan 450003, People’s Republic of China
Correspondence: Peiming Zheng Email [email protected]
Purpose: Macrophages are a major component of the tumour microenvironment and play an important role in chemoresistance of cancer. However, how exosomal microRNAs (miRNAs) derived from macrophages contribute to the development of doxorubicin resistance in gastric cancer (GC) are not clearly defined. The aim of this study was to investigate whether macrophage-derived exosomes mediate doxorubicin resistance in GC.
Methods: Exosomes isolated from macrophage culture medium were characterized and co-cultured with GC cells and the miR-223 level was detected using real-time quantitative PCR (RT-qPCR). The internalization of exosomes and transfer of miR-223 were observed via immunofluorescence. Macrophages were transfected with an miR-223 inhibitor or negative control. Cell Counting Kit-8 and flow cytometry were employed to explore the effect of macrophage-derived exosomes on the doxorubicin resistance of GC cells. Western blot and RT-qPCR assay were also performed to explore the regulation of GC chemotherapy resistance by exosomal miR-223.
Results: Here, the macrophages and macrophage-derived exosomes promoted doxorubicin resistance in GC cells. MiR-223 was enriched in macrophage-derived exosomes and they could be transferred to co-cultivated GC cells. The miR-223 knockdown in macrophages could reduce the effects of exosomes on GC cells. Functional studies revealed that exosomal miR-223 derived from macrophages promoted doxorubicin resistance in GC cells by inhibiting F-box and WD repeat domain-containing 7 (FBXW7). Clinically, the expression of miR-223 significantly increased in GC tissues and high expression of plasma exosomal miR-223 was highly linked with doxorubicin resistance in GC patients.
Conclusion: The exosomal transfer of macrophage-derived miR-223 conferred doxorubicin resistance in GC and targeting exosome communication may be a promising new therapeutic strategy for GC patients.
Keywords: exosome, miR-223, macrophage, doxorubicin resistance, gastric cancer
Introduction
Gastric cancer (GC) is currently the fifth most common malignancy and the third leading cause of cancer-related deaths worldwide.1 The incidence and mortality of GC are the highest in East Asia (particularly in Korea, Mongolia, Japan and China) and GC has become the second most lethal cancer in China.2 It is commonly diagnosed at the advanced stage and the 5-year overall survival rate is approximately 20–30% due to a lack of sensitive and specific early diagnosis.3 At present, traditional chemotherapy still represents the primary option for advanced GC even though several targeted therapies have achieved some benefits.4 Doxorubicin (DOX), a member of the anthracycline family, has been used to treat numerous malignancies, including GC, in combination with fluorouracil, cisplatin, paclitaxel and mitomycin.5 However, resistance to doxorubicin often occurs with the mechanisms remaining unclear, which results in relapse of cancer and poor survival.6
A plethora of elegant studies have shown that the cellular interaction between cancer cells and surrounding stroma cells in tumour microenvironment (TME) play a pivotal role in regulating cancer progression and therapy response.7 As one of the major populations of the TME, macrophages interact with cancer cells to promote tumorigenesis, progression and therapy resistance. The resident macrophages of the TME exhibiting protumour functions are often termed as tumour-associated macrophages (TAMs), which typically display M2-like phenotype.8,9 The positive correlation and ongoing dynamic interactions between TAMs and tumour cells have been proven in several types of cancer, including GC.10 Increasing evidence has shown that TAMs regulate the therapeutic responses of cancer cell and they are now being recognized as potential targets for anti-cancer therapy.11,12 However, the detailed interaction between anticancer therapies with TAMs remains unclear.
Exosomes, one of the extracellular vesicles with a size in the range of 50–150 nm, are secreted by many cells and involved in intercellular communication by transmitting intracellular cargos, such as proteins, lipids, mRNA and miRNA.13 Several studies have shown that exosomal transmission amongst TME cells were implicated in tumour metastasis and chemotherapy resistance.14,15 MicroRNAs are a class of 18–22 nucleotides small single-stranded non-coding RNA molecules that control the expression of multiple target genes at the posttranscriptional level. The lipid membranes of exosomes protect the internal miRNAs from being digested by RNases.13,16 Accumulating evidence has shown that the exosomal transfer of miRNA contributes to the development of chemoresistance in multiple tumour types.17,18 Shurtleff et al confirmed that one exosome-specific microRNA, miR-223, was selectively packed into exosomes and much more abundant in exosomes than in cells.19 Moreover, miR-223 has been proven highly expressed in the human macrophage cell line THP-1 and primary macrophages.20 It has emerged as a putative predictor of tumour invasiveness, metastasis and recurrence in many tumour types.21
F-box and WD repeat domain-containing 7 (FBXW7) is the substrate recognition component of an evolutionarily conserved SCF (complex of SKP1, CUL1, and F-box protein)-type ubiquitin ligase complex. It is considered as a p53-dependent tumor suppressor protein and leads to ubiquitination-mediated suppression of several oncoproteins including cyclin E, c-Myc, Notch, c-Jun and MCL1.22,23 Previous studies have shown that FBXW7 was a functional target of miR-223, and it has been reported to be involved in drug resistance in several types of cancer, including GC.24
In the present study, TAMs-like M2 polarized macrophages activated by IL4 with IL13 were constructed and the results showed that the macrophages promoted doxorubicin resistance in GC cells by exosomal transmission. We speculated that macrophages may exert their effects on GC cells by exosomal transfer of miR-223. This study aimed to determine whether exosomal miR-223 was involved and through which molecular mechanisms it elicited this function. The clinical relevance between the expression of exosomal miR-223 in plasma and chemotherapy efficiency in GC patients was also explored.
Materials and Methods
Cell Culture and Treatment
GC cell line AGS and human monocyte THP-1 were purchased from the Chinese Academy of Sciences Cell Bank of Type Culture Collection. All cell lines were cultured at 37°C with 5% CO2 in 1640/DMEM (HyClone) containing 10% exosome-depleted FBS supplemented with 100 U/mL penicillin and 100 μg/mL streptomycin (Gibco). For in-vitro differentiation of macrophages, THP-1 cells were first stimulated with 320 nM PMA for 6 h and then incubated for 48 h with 20 ng/mL of IL-4 plus 20 ng/mL of IL-13 (PeproTech, USA) to achieve M2-polarized macrophages. For co-culture experiment, the macrophages were grown on Transwell insert with 0.4 µm pore size (Corning) and the GC cells were grown in the bottom well of the Transwell chamber. For inhibition of exosome generation, macrophages were pre-treated with culture media containing 10 μM GW4869 (Sigma).
Exosome Isolation and Identification
In brief, the polarized macrophages were plated in 60-mm dishes at a density of 4×106 cells per well, then incubated for 48 h in complete PRMI1640 medium with 10% FBS that was previously depleted of contaminating vesicles by overnight centrifugation at 100,000×g. For each experiment, about 50mL conditioned medium was collected. The conditioned medium or plasma was centrifuged at 800 g for 10 min, followed by a centrifugation step of 3000 g for 30 min to remove cell debris. Next, the supernatant was filtered using a 0.22 µm filter (Millipore). The exosomes were pelleted by ultracentrifugation at 100,000 g for 90 min, washed in PBS, pelleted again and resuspended in PBS for further analysis. The morphology and size distribution of the isolated exosomes were determined using transmission electron microscopy (TEM) and nanoparticle tracking analysis (NTA) following our previous description.25,26
Cell Viability and Apoptosis Assay
GC cells were seeded in a 96-well plate at 1500–3000 cells per well and incubated with doxorubicin for 48 h. Cell viability was detected with Cell Counting Kit-8 (Dojindo Laboratories, Japan). The optical density at 450 nm was measured on a multiwall plate reader (FLX800, Bio-TEK). Apoptosis was measured using the FITC Annexin V Apoptosis Detection Kit I (BD Pharmagen, USA) following the manufacturer’s protocol. In brief, the cells were washed twice with cold PBS and then resuspended in 100 μL of 1X binding buffer. Then, 5 μL of FITC Annexin V and 5 μL of propidium iodide were added for 15 min at room temperature in the dark. After incubation, 400 μL of 1X binding buffer was added to each tube and analyzed by FACS Canto II flow cytometry (BD Biosciences).
In vitro Detection of miR-223 Transfer
The exosomes prepared from M2 macrophages transfected with Cy3-labelled miR-223 or without transfection (ctrl) were added to GC cell cultures to further observe the transfer of miRNA. The GC cells were fixed in 4% PFA, treated with 0.3% Triton X-100 and blocked with 3% BSA at 37°C. After being washed with PBS, cellular F-actin was visualized by staining with Alexa 488 phalloidin (LifeTechnologies, USA) in accordance with the manufacturer’s guidelines. The cells were mounted with ProLong Gold Antifade Reagent with DAPI (LifeTechnologies, USA). Images were captured using a Leica SP5 laser scanning confocal microscope.
EGFP Reporter Assay
The GC cells were seeded into 48-well plates at a density of 3 × 104 cells/well before transfection. When the cells reached 80% confluence, they were co-transfected with miR-223 mimics and pcDNA3/EGFP-FBXW7 3′UTR or mutant 3ʹUTR or control construct by using Lipofectamine 3000 in accordance with the manufacturer’s instructions. The cells were harvested at 48 h after transfection and lysed with RIPA buffer. Then, fluorescence intensity was measured by using F-4500 Fluorescence Spectrophotometer (Hitachi, Tokyo, Japan).
RNA Extraction and Real-Time Quantitative PCR
The total RNA from tissues/cells was extracted using TRIzol reagent (Invitrogen) and exosomal RNA was extracted using a miRNeasy Micro Kit (Qiagen) in accordance with the respective manufacturer’s instructions. RNA quantity and quality were evaluated using a NanoDrop spectrophotometer (Thermo Fisher Scientific). For mRNA expression analysis, reverse transcription was performed using PrimeScript RT master mix (TaKaRa, Japan). For miRNA expression analysis, total RNA was first reverse transcribed using Mir-X miRNA First-Strand Synthesis Kit (TaKaRa, Japan). Real-time quantitative PCR analysis was performed in triplicate on 7900 HT Real-Time PCR System (Applied Biosystems, USA) using SYBR Premix Ex Taq (TaKaRa, Japan) and the expression levels of GAPDH or U6 were used as endogenous control. The results were analyzed using the 2–ΔΔct calculation method.
Western Blot
The cells were lysed in equal volumes of ice cold lysis buffer and a protease inhibitor cocktail. Cell lyses were separated by SDS-PAGE and then transferred to a 0.2-μm PVDF membrane (Bio-Rad, USA). After blocking with Odyssey Blocking Buffer (Li-COR Biosciences, USA), the membrane was incubated with primary antibody (1:1000) at 4°C overnight, followed by incubation with IRDye 800CW or 680 secondary antibodies (1:5000, LI-COR Biosciences, USA). GAPDH was used as endogenous control. The Odyssey Infrared Imaging System was used to visualize targeted protein bands.
Patients and Samples
Human plasma samples from 30 GC patients and 20 age-matched healthy controls were collected from The First Affiliated Hospital of Henan University. The average age of the patients was 54.21±8.29 (X±SD), with 14 males and 16 females. All enrolled patients were diagnosed by gastroscopy or histopathology and did not undergo any preoperative treatment, including 8 stage I patients, 17 stage II patients and 5 stage IIIA patients. The clinicopathologic parameters were estimated according to the 8th AJCC/TNM staging system. All patients were treated with the standard care of doxorubicin-based therapy after surgery, doxorubicin resistance or doxorubicin sensitivity was defined by relapse or progression within 6 months or 6months after the last doxorubicin-based chemotherapy, respectively. GC tissues were obtained from resection before receiving any other therapy. This study was performed in accordance with the rules of the Declaration of Helsinki of 1975 (revised in 2013) and approved by the Ethics Committee of The First Affiliated Hospital of Henan University. Informed consent forms were obtained from all subjects before they participated in the study. Plasma samples were separated within 2 h after collection following a two-step centrifugation protocol (3000 g for 10 min at 4°C and 12,000 g for 10 min at 4°C) to thoroughly remove cell debris. The separated plasma was stored in RNase-free centrifuge tubes at −80°C until exosome extraction.
Statistical Analysis
Statistical significance between groups was determined by a two-tailed Student’s t-test or a one-way ANOVA test. Differences were considered to be significant when P<0.05. All statistical data were displayed as means ± standard deviation (SD) and analyzed for statistical significance with SPSS 20.0 and GraphPad Prism 7.0 Software.
Results
TAMs-Derived Exosomes Induce the Resistance of GC Cells to Doxorubicin in vitro
A previous study indicated that TAMs infiltrated in GC exhibited an M2-polarized phenotype and played a role in tumour chemotherapy resistance.25 In the present study, M2-polarized macrophages were generated from THP-1 monocytes in vitro and co-cultured with GC cells in a transwell insert to determine whether M2-polarized macrophages conferred doxorubicin resistance in GC cells. Then, the GC cells were treated with doxorubicin for 48 h. CCK8 assay showed that co-culture with macrophages could substantially increase cell viability and enhance drug resistance of GC cells. More importantly, when GW4869 was used to inhibit neutral sphingomyelinase and prevent exosome release in macrophages co-cultured with GC cells, the IC50 values of doxorubicin were significantly reduced (Figure 1A). These results indicated that macrophages may enhance the doxorubicin resistance of GC cells by transmitting exosomes.
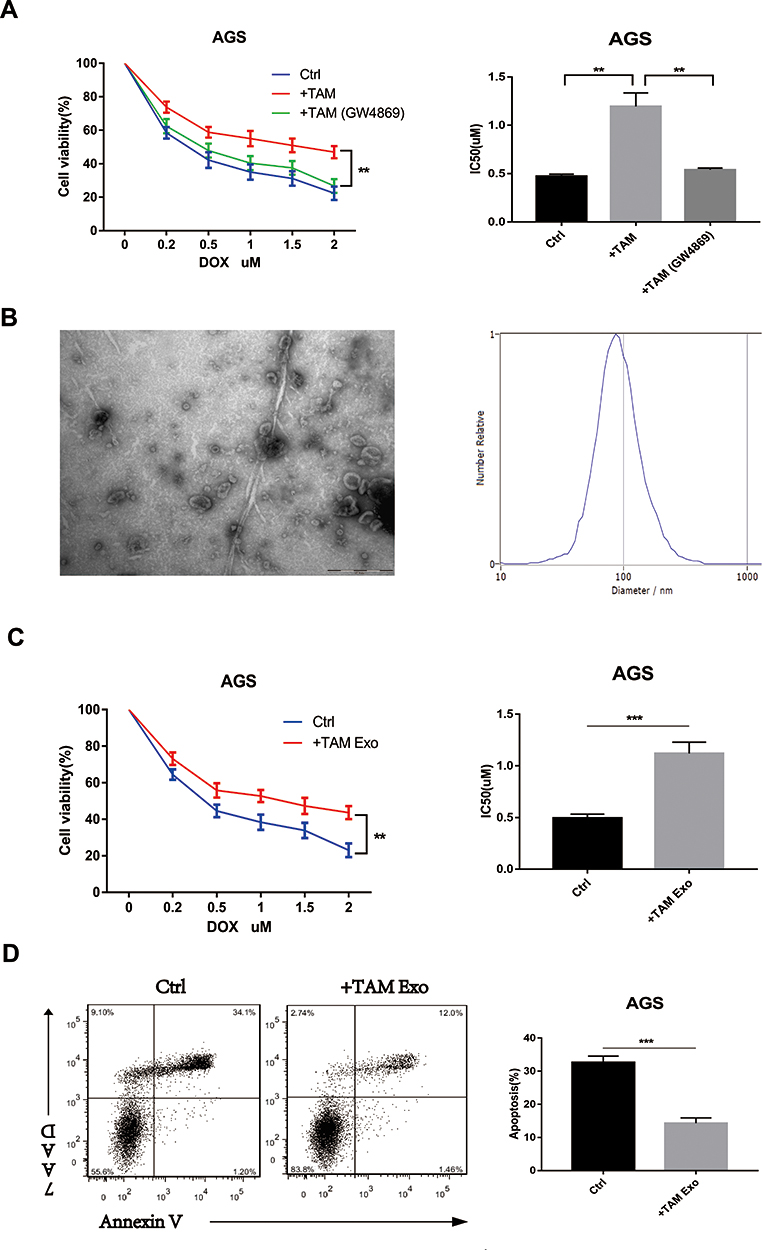 |
Figure 1 TAMs-derived exosomes induce the resistance of GC cells to doxorubicin (DOX) in vitro. (A) The AGS cells were co-cultured with TAMs pre-treated with or without 10 µM GW4869 (blocking exosome generation), and then exposed to DOX for 48h. Cell viability and IC50 of DOX was analyzed respectively. (B) The morphology and size distribution of the isolated exosomes were identified using TEM and NTA. (C and D) AGS cells were pre-treated with PBS (ctrl) or TAMs-derived exosomes (TAM Exo) for 24h, then exposed to DOX for 48h. Cell viability, IC50 of DOX and cell apoptosis was detected respectively. **p<0.01, *** p<0.001. |
Exosomes were isolated and identified from the conditioned medium of M2 macrophages to further determine the effects of macrophage-derived exosomes on the doxorubicin resistance of GC cells. The purified exosomes displayed a typical cuppy structure, with a diameter ranging from 80 nm to 120 nm (Figure 1B). The GC cells were treated with PBS and macrophage-derived exosomes. Then, the cells were treated with doxorubicin for another 48 h. As shown in Figure 1C and D, the macrophage-derived exosomes reduced the apoptosis rate, increased cell viability and enhanced the doxorubicin resistance of GC cells. The IC50 value of doxorubicin in GC cells treated with macrophage-derived exosomes increased. These data confirmed that macrophage-derived exosomes could induce the resistance of GC cells to doxorubicin.
Exosomal Transfer of Functional miR-223 Confers Doxorubicin Resistance of GC Cells
Several studies and our previous research proved that miR-223 was enriched in macrophage-derived exosomes and played a vital role in tumour progression. Here, the roles of exosomal miR-223 in the doxorubicin resistance of GC cells were further investigated. The results of RT-qPCR validated that the intracellular miR-223 levels significantly increased in GC cells co-cultured with M2-polarized macrophages or macrophage-derived exosomes. Similarly, when exosomal transfer was blocked by GW4869, the miR-223 levels of GC cells were significantly reduced (Figure 2A). The M2-polarized macrophages were transiently transfected with Cy3-labeled miR-223 to further confirm that the elevated miR-223 was caused by direct exosomal transfer from M2 macrophages to GC cells. Then, the GC cells were incubated with exosomes isolated from the above macrophages. By using confocal microscopy, Cy3-labelled signals were detected in the GC cells incubated with macrophage-derived exosomes transfected with Cy3-labeled miR-223 but not mock-transfected cells (Figure 2B). In summary, these data revealed that macrophage-derived exosomes delivered miR-223 to GC cells.
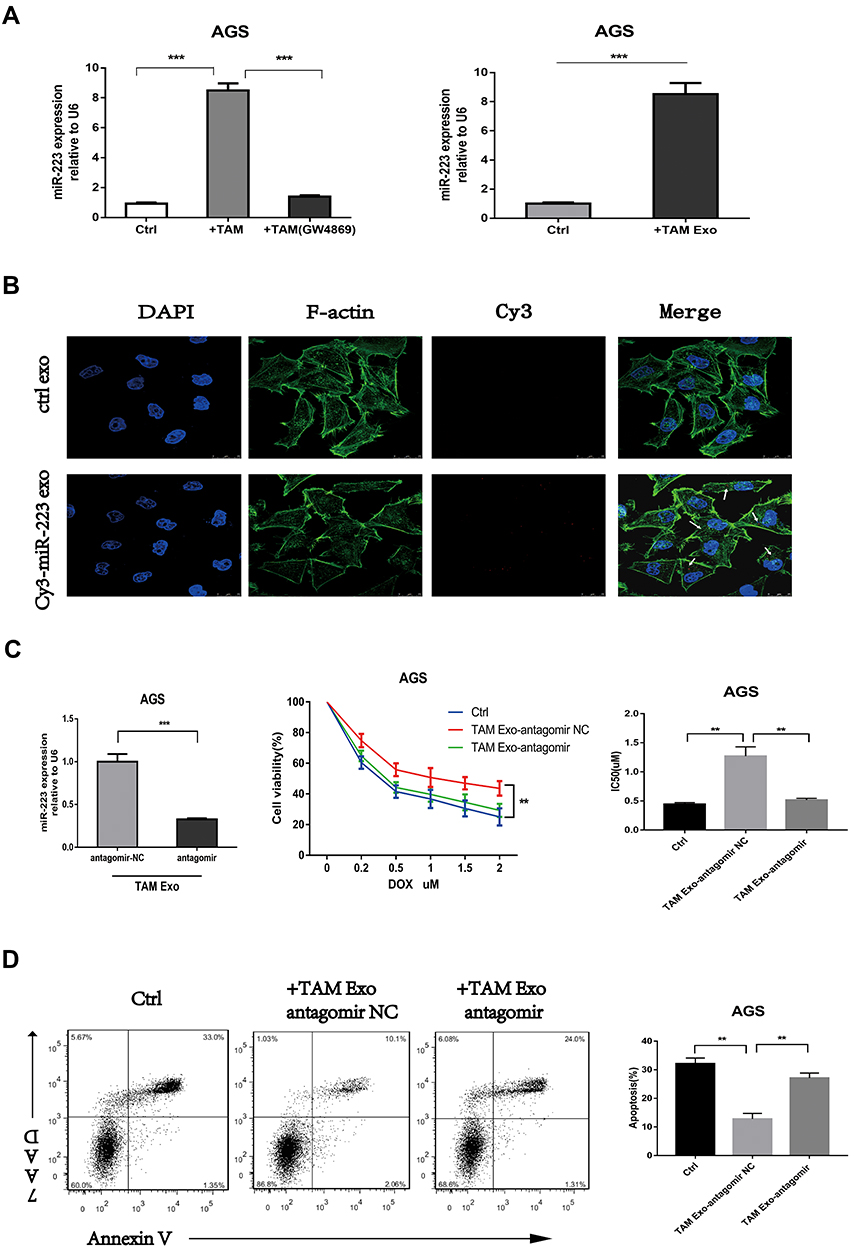 |
Figure 2 Exosomal transfer of functional miR-223 confers doxorubicin resistance of GC cells. (A) The expression level of miR-223 was detected using RT-qPCR in AGS cells co-cultured with TAMs (with or without GW4869) or TAMs-derived exosomes (TAM Exo). (B) The internalization of exosomes and transfer of Cy3-labeled miR-223 were observed via immunofluorescence. (C and D) The AGS cells were pre-treated with exosomes derived from TAMs which were transfected with miR-223 antagomir (TAM Exo antagomir) or NC control, then exposed to DOX for 48h. Cell viability, IC50 of DOX and cell apoptosis was detected respectively. **p<0.01, *** p<0.001. |
MiR-223 was inhibited using antagomir in M2-polarized macrophages to determine the effects of exosomal miR-223 on the chemoresistance of GC cells. Then, the exosomes were isolated from transfected macrophages and incubated with the GC cells. The miR-223 levels of the treated GC cells also decreased when macrophages were transfected with miR-223 antagomir (Figure 2C). Meanwhile, the inhibition of miR-223 increased the apoptosis rate, reduced cell viability and impaired the doxorubicin resistance of GC cells compared with the mock transfectant (Figure 2C and D). Taken together, these results suggested that macrophage-derived exosomes transferred miR-223 to GC cells and promoted drug resistance in GC cells.
Exosomal miR-223 Regulates Doxorubicin Sensitivity Through FBXW7 Targeting
Next, whether the exosomal transfer of functional miR-223 regulates doxorubicin sensitivity through FBXW7 targeting and the potential mechanism was investigated. Epithelial–mesenchymal transition (EMT) has been proven to be associated with the development of doxorubicin resistance in several cancer cells.27,28 Here, the results showed that FBXW7 expression was down-regulated in GC cells co-cultured with macrophages or macrophage-derived exosomes, accompanied with a decreased expression of epithelial marker proteins (E-cadherin) and an increased expression of mesenchymal marker proteins (N-cadherin, vimentin). However, these effects were reversed when macrophages were pre-treated with GW4869 or transfected with miR-223 antagomir (Figure 3A and B). These results indicated that miR-223/FBXW7 axis may regulate the doxorubicin sensitivity of GC cells via affecting EMT. Further, EGFP reporter assay was conducted in GC cells to observe the direct targeting of FBXW7 by miR-223. The results revealed that EGFP intensity significantly decreased in cells transfected with the FBXW7 wild 3ʹUTR reporter plus mimic-miR-223, while no difference was found when transfecting with the FBXW7 mutant 3ʹUTR reporter (Figure 3C).
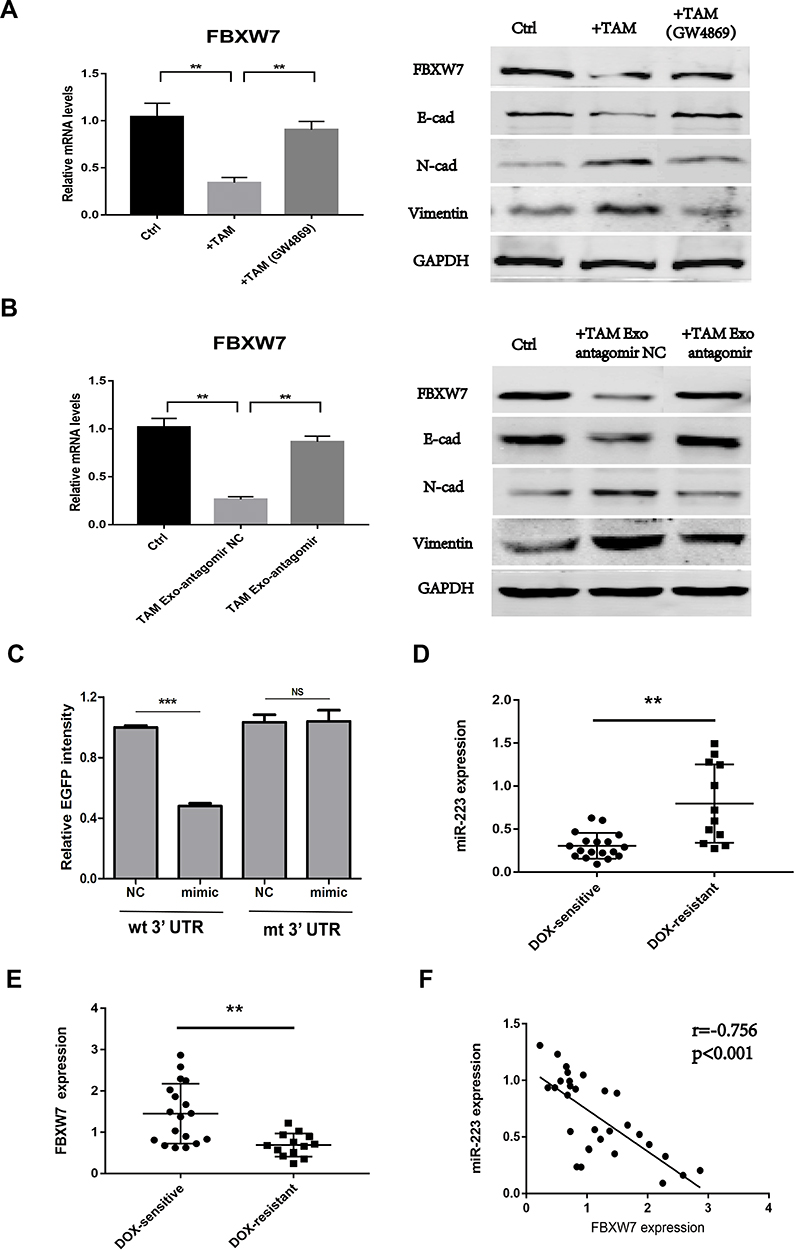 |
Figure 3 Exosomal miR-223 regulates doxorubicin sensitivity through FBXW7 targeting. (A) The expression level of FBXW7 and EMT marker proteins were detected in AGS cells co-cultured with TAMs treated with or without GW4869. (B) The expression level of FBXW7 and EMT marker proteins were detected in AGS cells treated with TAMs-derived exosomes, which were transfected with miR-223 antagomir (TAM Exo antagomir) or NC control. (C) EGFP reporter analysis was performed in GC cells after wild-type or mutant 3ʹUTR reporter plasmids were co-transfected with miR-223 mimic or NC. (D) The expression level of miR-223 was detected using RT-qPCR in GC tissues from DOX-resistant and DOX-sensitive patients. (E) The expression level of FBXW7 was detected using RT-qPCR in GC tissues from DOX-resistant and DOX-sensitive patients. (F) Pearson correlation analysis between miR-223 and FBXW7 expression level was performed in 30 GC patients. NS p>0.05, **p<0.01, *** p<0.001. |
Then, the expression levels of FBXW7 and miR-223 in primary GC tissues derived from 30 GC patients were detected. The results showed that low FBXW7 expression and high miR-223 expression were presented in patients with DOX resistance (Figure 3D and E). FBXW7 was then correlated with miR-223 expression by using Spearman correlation analysis and a significant inverse correlation was observed between miR-223 and FBXW7 (Figure 3F).
Plasma Exosomal miR-223 Level is Elevated in DOX-Resistant GC Patients
The plasma exosomal miR-223 expression levels in different GC patients and paired healthy controls were analysed using RT-qPCR. The plasma exosomal miR-223 levels in GC patients were significantly up-regulated compared with those in healthy controls. Moreover, DOX-resistant patients showed higher exosomal miR-223 expression than those with DOX sensitivity (Figure 4A). The exosomal miR-223 expression in 10 GC patients at the time of their first chemotherapy and relapse was also compared. The exosomal miR-223 levels increased in these patients at recurrent (Figure 4B). Despite the relatively small sample size, these results indicated that circulating exosomal miR-223 may be a useful indicator for DOX resistance in GC.
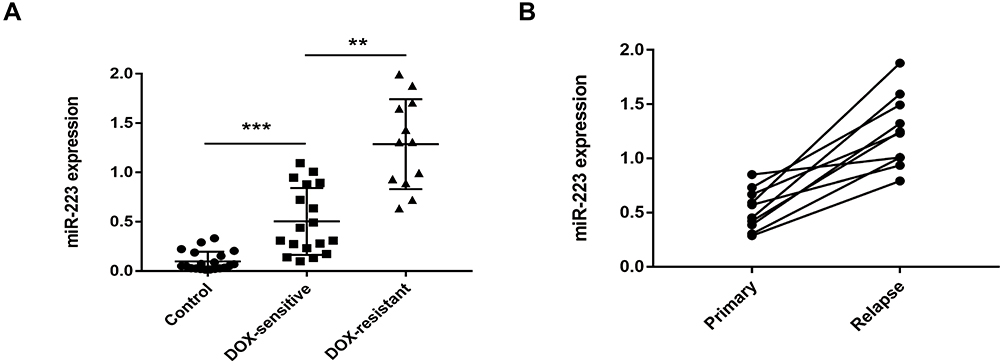 |
Figure 4 Plasma exosomal miR-223 level is elevated in DOX-resistant GC patients. (A) Expression levels of plasma exosomal miR-223 in different GC patients and paired healthy controls. (B) Expression levels of plasma exosomal miR-223 in 10 paired GC patients at the time of primary and recurrent. **p<0.01, *** p<0.001. |
Discussion
Although targeted therapy and immunotherapy have been emerging as important methods and achieved some promising benefits in cancer treatment, traditional chemotherapy is still considered the first-line treatment for most tumours, including advanced GC.3 Drug resistance is one of the most significant obstacles to effective GC therapy.29 The multiple interactions between tumour cells and stroma cells in the TME, including macrophages, CAFs and immune cells, cause tumour progression and therapy resistance.7,8,11 However, whether macrophage-derived exosomes are involved in doxorubicin chemoresistance in GC and the potential mechanisms have not been clearly identified.
We have previously shown that TAMs were enriched in GC and contributed to tumour metastasis and chemotherapy resistance by exosome-mediated cellular communication.14,25 In the present study, macrophage-derived exosomes delivered miR-223 to GC cells to induce doxorubicin resistance. Exosomes are documented to play an important role in cancer progression and chemoresistance. Apart from cancer cells, exosomes from tumour stromal cells contribute to chemotherapy resistance in a wide range of human tumors.17,30,31 Increasing evidence demonstrated that miRNAs were encapsulated in exosomes to avoid degradation and horizontal transfer of exo-miRNAs modulated angiogenesis, metastasis and chemoresistance.16,18,20 Exosomal miR-126a and miR-222 were involved in the doxorubicin resistance of breast cancer.32,33 Exosomal transfer of stroma-derived miR-21 conferred paclitaxel resistance in ovarian cancer cells through targeting APAF1.17 Hu et al demonstrated that CAF-secreted exosomal miR-92a-3p promoted metastasis and chemotherapy resistance by enhancing cell stemness and epithelial–mesenchymal transition in colorectal cancer.31 A recent study reported that TAMs affected the cisplatin resistance of epithelial ovarian cancer cells through the exchange of exosomal miR-223.34
MiR-223 has been known to be up-regulated in many human cancers and proven to be involved in tumour invasiveness, metastasis and chemoresistance.21 In GC, a previous study revealed that miR-223 increased GC cell resistance to cisplatin and trastuzumab.24,35 While broadly expressed in myeloid cells, miR-223 was highly expressed in macrophages and selectively packed into exosomes, which was consistent with the results of our research.20,34 In addition, the present study showed that circulating exosomal miR-223 increased in the plasma of DOX-resistant and recurrent GC patients. These results showed that high levels of miR-223 frequently observed in primary tumours and peripheral circulation may not only result from cancer cells but also the exosomal transfer of miR-223 from the surrounding TAMs.
The data also suggested that macrophage-derived exosomal miR-223 was able to directly target FBXW7, a protein involved in the regulation of chemoresistance of GC and other cancer cells.31,35 Inuzuka et al revealed that FBXW7 regulates the apoptotic pathway by controlling MCL1 destruction.36 FBXW7 and PTEN work together to inhibit breast cancer progression by suppressing mTOR.37 Furthermore, several studies have demonstrated that the miR-223/FBXW7 axis was implicated in the regulation of chemotherapeutic sensitivity of human cancers.24,35,38 In addition, a negative correlation between miR-223 and FBXW7 was observed in GC tissues. The results of WB indicated that miR-223/FBXW7 axis may regulate the doxorubicin sensitivity of GC cells via affecting EMT. Of course, mechanisms leading to the lack of response to antitumor drugs are numerous and rarely act alone, other targets or pathways may also be modulated by exosomal miR-223. The focus of our study was the exosome-mediated reciprocity between TAM and GC cells, which represented a potential target for therapeutic intervention.
Mounting evidence elucidated the potential of exosomal miRNAs as novel diagnostic and prognostic markers for human cancers.39,40 Therefore, the clinical values of exosomal miR-223 in the DOX resistance of GC were preliminarily investigated in the present study. The findings showed that the exosomal miR-223 in the plasma of DOX-resistant patients was highly expressed compared with that in healthy controls and DOX-sensitive patients. The exosomal miR-223 expression in plasma was correlated with the recurrence of GC patients. These results indicated that exosomal miR-223 may act as a biomarker for DOX response in GC.
In conclusion, this study provided evidence that the exosome-mediated transfer of miR-223 derived from macrophages enhanced the chemoresistance of GC to DOX. Blocking the function of exosomal miR-223 secreted by TAMs may be a promising adjuvant therapeutic strategy to overcome GC chemoresistance. Moreover, circulating exosomal miR-223 was up-regulated in the plasma of DOX-resistant GC patients. Future studies on large-scale clinical samples, dynamic monitoring and long-term follow-up are necessary to further confirm these conclusions.
Funding
This study was funded by the National Natural Science Foundation of China (No. 81802094).
Disclosure
The authors declare that they have no conflicts of interest.
References
1. Bray F, Ferlay J, Soerjomataram I, et al. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2018;68(6):394–424. doi:10.3322/caac.21492
2. Chen W, Zheng R, Baade PD, et al. Cancer statistics in China, 2015. CA Cancer J Clin. 2016;66:115–132.
3. Orditura M, Galizia G, Sforza V, et al. Treatment of gastric cancer. World J Gastroenterol. 2014;20:1635–1649.
4. Tirino G, Pompella L, Petrillo A, et al. What’s New in Gastric Cancer: the Therapeutic Implications of Molecular Classifications and Future Perspectives. Int. J Mol Sci. 2018;19:119.
5. Murad A, Santiago F, Petroianu A, et al. Modified therapy with 5-fluorouracil, doxorubicin, and methotrexate in advanced gastric cancer. Cancer. 1993;72:37–41.
6. Ramos P, Bentires-Alj M. Mechanism-based cancer therapy: resistance to therapy, therapy for resistance. Oncogene. 2015;34:3617–3626.
7. Quail DF, Joyce JA. Microenvironmental regulation of tumor progression and metastasis. Nat Med. 2013;19:1423–1437.
8. Noy R, Pollard JW. Tumor-Associated Macrophages: from Mechanisms to Therapy. Immunity. 2014;41:49–61.
9. Qian BZ, Pollard JW. Macrophage diversity enhances tumor progression and metastasis. Cell. 2010;141:39–51.
10. Cardoso AP, Pinto ML, Pinto AT, et al. Macrophages stimulate gastric and colorectal cancer invasion through EGFR Y1086, c-Src, Erk1/2 and Akt phosphorylation and smallGTPase activity. Oncogene. 2014;33(16):2123–2133. doi:10.1038/onc.2013.154
11. Ruffell B, Coussens LM. Macrophages and therapeutic resistance in cancer. Cancer Cell. 2015;27(4):462–472. doi:10.1016/j.ccell.2015.02.015
12. Brown JM, Recht L, Strober S. The Promise of Targeting Macrophages in Cancer Therapy. Clinical Cancer Research. 2017;23(13):3241–3250. doi:10.1158/1078-0432.CCR-16-3122
13. Penfornis P, Vallabhaneni KC, Whitt J, et al. Extracellular vesicles as carriers of microRNA, proteins and lipids in tumor microenvironment. Int J Cancer. 2016;138(1):14–21.
14. Zheng PM, Luo Q, Wang WW. Tumor-associated macrophages-derived exosomes promote the migration of gastric cancer cells by transfer of functional Apolipoprotein E[J]. Cell Death Dis. 2018;9(4):434. doi:10.1038/s41419-018-0465-5
15. Qu L, Ding J, Chen C, et al. Exosome-Transmitted lncARSR Promotes Sunitinib Resistance in Renal Cancer by Acting as a Competing Endogenous RNA. Cancer Cell. 2016;29(5):653–668. doi:10.1016/j.ccell.2016.03.004
16. Turchinovich A, Weiz L, Burwinkel B, et al. Extracellular miRNAs: the mystery of their origin and function. Trends Biochem Sci. 2012;37(11):460–465. doi:10.1016/j.tibs.2012.08.003
17. Au Yeung CL, Co -N-N, Tsuruga T, et al. Exosomal transfer of stroma-derived miR21 confers paclitaxel resistance in ovarian cancer cells through targeting APAF1. Nat Commun. 2016;7(1):11150. doi:10.1038/ncomms11150
18. Challagundla KB, Wise PM, Neviani P, et al. Exosome-mediated transfer of microRNAs within the tumor microenvironment and neuroblastoma resistance to chemotherapy. JNCI: Journal of the National Cancer Institute. 2015;107(7):254. doi:10.1093/jnci/djv135
19. Shurtleff MJ, Temoche-Diaz MM, Karfilis KV, et al. Y-box protein 1 is required to sort microRNAs into exosomes in cells and in a cell-free reaction. Elife. 2016;5:e19276. doi:10.7554/eLife.19276
20. Aucher A, Rudnicka D, Davis DM. MicroRNAs transfer from human macrophages to hepato-carcinoma cells and inhibit proliferation. J Immunol. 2013;191(12):6250–6260. doi:10.4049/jimmunol.1301728
21. Li X, Zhang Y, Zhang H, et al. miRNA-223 promotes gastric cancer invasion and metastasis by targeting tumor suppressor EPB41L3. Mol Cancer Res. 2011;9(7):824–833. doi:10.1158/1541-7786.MCR-10-0529
22. Welcker M, Clurman BE. FBW7 ubiquitin ligase: a tumour suppressor at the crossroads of cell division, growth and differentiation. Nat Rev Cancer. 2008;8(2):83–93. doi:10.1038/nrc2290
23. Takeishi S, Nakayama K. Role of Fbxw7 in the maintenance of normal stem cells and cancer-initiating cells. Br J Cancer. 2014;111:1054–1059.
24. Zhou XY, Jin WJ, Jia HY, et al. MiR-223 promotes the cisplatin resistance of human gastric cancer cells via regulating cell cycle by targeting FBXW7. J Exp Clin Cancer Res. 2015;34:28.
25. Zheng PM, Chen L, Yuan XL, et al. Exosomal transfer of tumor-associated macrophage-derived miR-21 confers cisplatin resistance in gastric cancer cells. J Exp Clin Cancer Res. 2017;36:53.
26. Zheng PM, Zhang HL, Gao HJ, et al. Plasma Exosomal Long Noncoding RNA lnc-SLC2A12-10:1 as a Novel Diagnostic Biomarker for Gastric Cancer. Onco Targets Ther. 2020;13:4009–4018.
27. Zhang YB, Lu Y, Zhang W, et al. FSCN‑1 increases doxorubicin resistance in hepatocellular carcinoma through promotion of epithelial-mesenchymal transition. Int J Oncol. 2018;52:1455–1464.
28. Du FY, Yu L, Wu Y, et al. miR-137 alleviates doxorubicin resistance in breast cancer through inhibition of epithelial-mesenchymal transition by targeting DUSP4. Cell Death Dis. 2019;10:922.
29. Riquelme I, Letelier P, Riffo-Campos A, et al. Emerging Role of miRNAs in the Drug Resistance of Gastric Cancer. Int J Mol Sci. 2016;17:424.
30. Boelens MC, Wu TJ, Nabet BY, et al. Exosome Transfer from Stromal to Breast Cancer Cells Regulates Therapy Resistance Pathways. Cell. 2014;159:499–513.
31. Hu JL, Wang W, Lan XL, et al. CAFs secreted exosomes promote metastasis and chemotherapy resistance by enhancing cell stemness and epithelial-mesenchymal transition in colorectal cancer. Mol Cancer. 2019;18:91.
32. Wei F, Ma CY, Zhou T, et al. Exosomes derived from gemcitabine-resistant cells transfer malignant phenotypic traits via delivery of miRNA-222-3p. Mol Cancer. 2017;16:132.
33. Deng Z, Rong Y, Teng Y, et al. Exosomes miR-126a released from MDSC induced by DOX treatment promotes lung metastasis. Oncogene. 2017;36:639–651.
34. Zhu XL, Shen HL, Yin XM, et al. Macrophages derived exosomes deliver miR-223 to epithelial ovarian cancer cells to elicit a chemoresistant phenotype. J Exp Clin Cancer Res. 2019;38:81.
35. Eto K, Iwatsuki M, Watanabe M, et al. The sensitivity of gastric cancer to trastuzumab is regulated by the miR-223/FBXW7 pathway. Int J Cancer. 2015;136:1537–1545.
36. Inuzuka H, Shaik S, Onoyama I, et al. SCF(FBW7) regulates cellular apoptosis by targeting MCL1 for ubiquitylation and destruction. Nature. 2011;471:104–109.
37. Mao JH, Kim IJ, Wu D, et al. FBXW7 targets mTOR for degradation and cooperates with PTEN in tumor suppression. Science. 2008;321:1499–1502.
38. Li RY, Wu SJ, Chen X, et al. miR-223/FBW7 axis regulates doxorubicin sensitivity through epithelial mesenchymal transition in non-small cell lung cancer. Am J Transl Res. 2016;8:2512–2524.
39. Matsumura T, Sugimachi K, Iinuma H, et al. Exosomal microRNA in serum is a novel biomarker of recurrence in human colorectal cancer. Br J Cancer. 2015;113:275–281.
40. Zhang YY, Han T, Feng D, et al. Screening of non-invasive miRNA biomarker candidates for metastasis of gastric cancer by small RNA sequencing of plasma exosomes. Carcinogenesis. 2020;41:582–590.
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.