Back to Journals » OncoTargets and Therapy » Volume 13
Exosomal miR-638 Inhibits Hepatocellular Carcinoma Progression by Targeting SP1
Authors Yang J, Li B, Zhao S, Du H, Du Y ![]()
Received 9 March 2020
Accepted for publication 15 June 2020
Published 7 July 2020 Volume 2020:13 Pages 6709—6720
DOI https://doi.org/10.2147/OTT.S253151
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Leo Jen-Liang Su
This paper has been retracted.
Jing Yang, 1 Bo Li, 1 Shuo Zhao, 2 Hongyu Du, 1 Yaming Du 3
1Department of Pathology, Jinzhou Medical University, Jinzhou, Liaoning, People’s Republic of China; 2Department of Nursing, The First Affiliated Hospital of Jinzhou Medical University, Jinzhou, Liaoning, People’s Republic of China; 3Department of Vascular Surgery, The First Affiliated Hospital of Jinzhou Medical University, Jinzhou, Liaoning, People’s Republic of China
Correspondence: Yaming Du Email [email protected]
Purpose: Exosomal microRNAs (miRNAs) play essential roles in the development of hepatocellular carcinoma (HCC). Nevertheless, the role and mechanism of exosomal miR-638 in HCC development remain largely unknown.
Methods: Exosomes were isolated and confirmed via transmission electron microscopy and western blot. The abundances of miR-638 and specificity protein 1 (SP1) were measured via quantitative reverse transcription polymerase chain reaction or western blot. Cell proliferation was investigated by Cell Counting Kit-8, colony formation assay, apoptosis, cell cycle distribution and related protein expression. Cell migration and invasion were detected via transwell assay and western blot. Co-culture experiment was performed to assess exosome transfer from HCC cells to endothelial cells. The target correlation between miR-638 and SP1 was analyzed via dual-luciferase reporter and RNA immunoprecipitation assays. The subcutaneous xenograft experiment was conducted to test the function of miR-638 in vivo.
Results: The miR-638 level declined in exosomes from serum or HCC cell medium. miR-638 overexpression repressed HCC cell proliferation by decreasing viability and colony formation and inducing apoptosis and cell cycle arrest at G1 phase, and decreased abilities of migration and invasion. Exosomal miR-638 from HCC cells could transfer to human umbilical vein endothelial cells (HUVECs) and suppress HUVEC proliferation, migration and invasion. SP1 was a target of miR-638 and overexpression of SP1 reversed the effect of miR-638 on HCC cells. Overexpression of miR-638 reduced xenograft tumor growth via decreasing SP1.
Conclusion: Exosomal miR-638 inhibited HCC tumorigenesis by targeting SP1. This study indicated the potential clinical implications of miR-638 in HCC.
Keywords: hepatocellular carcinoma, exosome, miR-638, SP1
Introduction
Liver cancer is a public malignancy with the second leading cause of cancer-related deaths, and hepatocellular carcinoma (HCC) represents more than 90% of cases.1 In recent years, new hope has been brought for HCC patients with the progress of treatment options.2 Nevertheless, the outcome of patients remains unsatisfactory. Therefore, it is urgent to find a new strategy for the treatment of HCC.
Exosomes are a class of extracellular vesicles that take part in intercellular communication in HCC.3 Exosomes play important roles in the tumorigenesis, diagnosis and treatment of HCC by transmitting nucleic acids, proteins or lipids.4 Noncoding RNAs are enriched in exosomes and exosomal noncoding RNAs have essential roles in HCC development.5 MicroRNAs (miRNAs) are small noncoding RNAs which could be transferred by exosomes to participate in the progression and therapeutics of HCC.6 Previous studies suggest that miR-638 has an important clinical significance in HCC development.7,8 Moreover, knockdown of miR-638 could promote HCC development by increasing cell growth, angiogenesis, migration and invasion.9,10 In addition, miR-638 could be enriched in exosomes, and exosomal miR-638 plays a key role in human cancers.11,12 More importantly, serum exosomal miR-638 has an important prognostic role in HCC.13 However, the mechanism of exosomal miR-638 in HCC development remains largely unclear.
miRNAs exhibit their roles in HCC by regulating gene expression and translation.14 Specificity protein 1 (SP1) is overexpressed and plays an oncogenic role in many cancers by regulating cell proliferation, differentiation, apoptosis and angiogenesis.15 Accruing evidence indicates that SP1 is associated with cell proliferation, migration, invasion and angiogenesis in HCC.16–18 Moreover, starBase online predicts that SP1 might act as a target of miR-638. Hence, we hypothesized that exosomal miR-638 might target SP1 to mediate HCC progression. In the present research, we measured the exosomal miR-638 level, and investigated the effect of miR-638 on HCC development. Moreover, we analyzed the target association between miR-638 and SP1.
Patients and Methods
Patient Tissues and Serum
Forty-two HCC patients and 20 normal volunteers were recruited from Jinzhou Medical University. The peripheral blood samples were harvested and centrifuged for serum collection. The cancer and para-tumor tissues were obtained from HCC patients. The patients did not receive any other therapy prior to sample collection. Written informed consent was obtained from all subjects. The patients’ features are displayed in Table 1. This study was carried out in accordance with the Guidelines “Declaration of Helsinki” and under the approval of the Ethics Committee of Jinzhou Medical University.
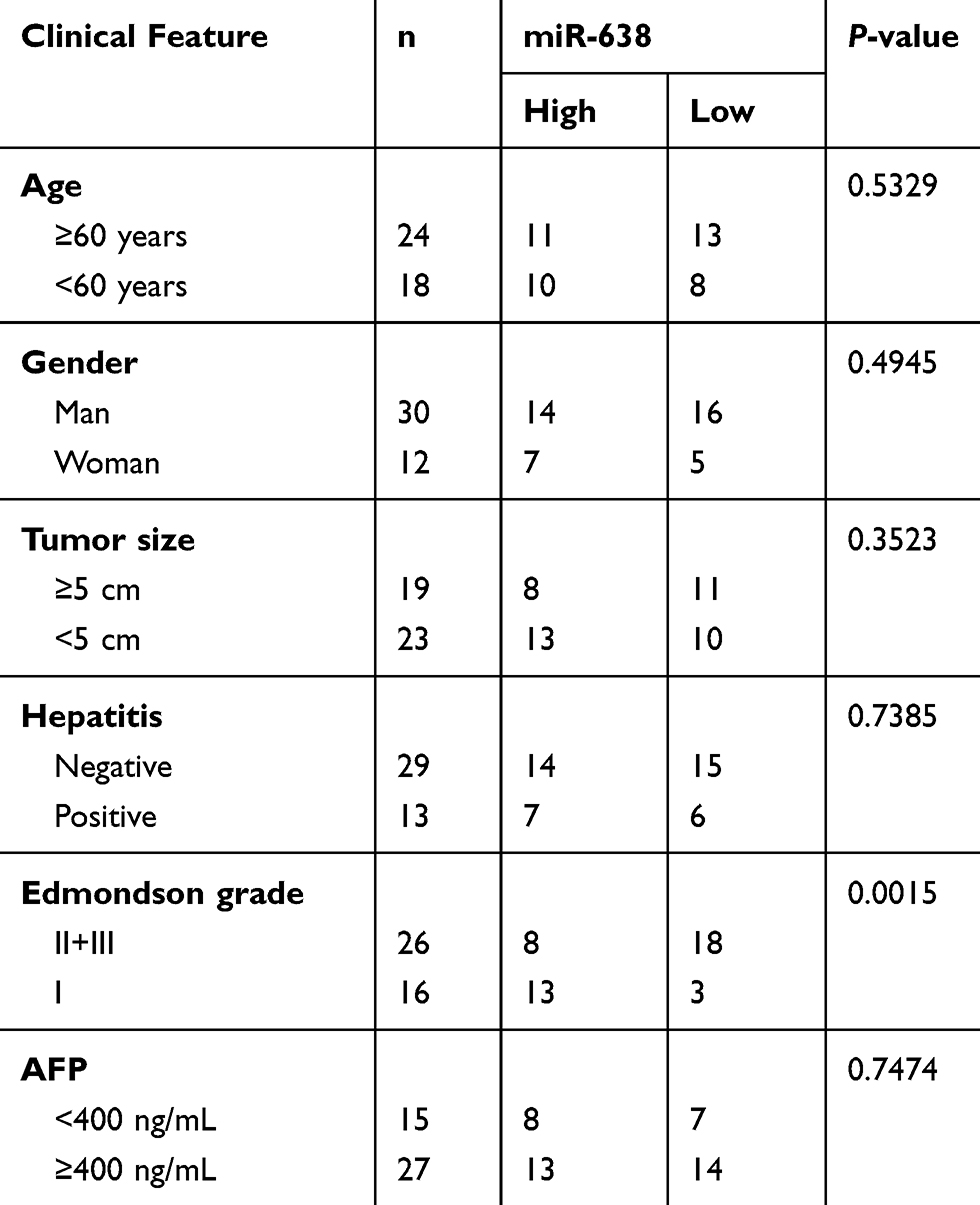 |
Table 1 The Relationship Between miR-638 Expression and Clinicopathological Features in HCC (n=42) |
Cell Culture and Treatment
Human umbilical vein endothelial cells (HUVECs), HCC cell lines (MHCC97-H, HCCLM3 and Huh7) and normal human liver cell line THLE-2 were provided via Bena Culture Collection (Beijing, China). DMEM (Sigma, St. Louis, MO, USA) with 10% fetal bovine serum (Biosun, Shanghai, China) as well as 1% penicillin–streptomycin (Sigma) was applied to cell culture. The cells were maintained at 37 °C in 5% CO2, and medium was changed every 3 days. To block the release of exosomes, cells were incubated with 10 μM of SW4869 (Sigma).
Exosome Purification and Validation
The exosomes were purified from serum or cell medium using the Exoquick exosome extraction kit (SBI System Biosciences, Mountain View, CA, USA) according to the instructions.19 The exosomes were validated by transmission electron microscopy (TEM) (Hitachi, Tokyo, Japan) and expression of exosomal markers.20,21
Quantitative Reverse Transcription Polymerase Chain Reaction (qRT-PCR)
Exosomal RNA was isolated using the Exosome Purification and RNA Isolation kit (Amyjet Scientific, Wuhan, China), and RNA from tissues or cells was extracted using TRIzol (Solarbio, Beijing, China). The cDNA was generated using the specific reverse transcription kit (Thermo Fisher, Wilmington, DE, USA). The cDNA was mixed with SYBR Green (Vazyme, Nanjing, China) and specific primers, and then applied to qRT-PCR. The primers were synthesized from Genscript (Nanjing, China) and shown as: SP1 (sense, 5ʹ-CGCCCTCTGACCAAGATCACT-3ʹ; antisense, 5ʹ-GGGAGTTGTTGCTGTTCTCATTGG-3ʹ), miR-638 (sense, 5ʹ-GAGAGGATCCTGCCGCAGATCGCTG-3ʹ; antisense, GAGTAAGCTTCAGGGAGTCCTCTGCC), U6 (sense, 5ʹ-ACGCAAATTCGTGAAGCGTT-3ʹ; antisense, AACGCTTCACGAATTTGCGT) and GAPDH (sense, 5ʹ-GACAGTCAGCCGCATCTTCT-3ʹ; antisense, 5ʹ-GCGCCCAATACGACCAAATC-3ʹ). The relative expression of SP1 or miR-638 was calculated with GAPDH or U6 as a reference according to the delta–delta cycle threshold method.22
Cell Transfection
SP1 overexpression vector was generated using pcDNA3.1 (Thermo Fisher). The pcDNA3.1 vector was exploited as a negative control (vector). miR-638 mimic (5ʹ-AGGGAUCGCGGGCGGGUGGCGGCCU-3ʹ) and mimic negative control (NC, 5ʹ-UUCUCCGAACGUGUCACGUTT-3ʹ) were generated via Ribobio (Guangzhou, China). Huh7 cells were transfected with 50 or 100 nM oligonucleotides or 600 ng vectors via Lipofectamine 3000 reagent (Thermo Fisher) for 24 h. The non-transfected cells were regarded as the control group.
Co-Culture of HUVECs and Huh7 Cells
Huh7 cells were transfected with miR-638 mimic or NC, and then the exosomes were isolated, named as Huh7-exo(miR-638) or Huh7-exo(NC) group, respectively. To validate that miR-638 was transferred by exosome, Huh7 cells transfected with miR-638 mimic were pretreated by SW4869, and then exosomes were isolated, named as Huh7-exo(miR-638)-GW4869 group. The exosomes were co-cultured with HUVECs for 48 h.
Cell Counting Kit-8 (CCK-8)
The viability of Huh7 cells or HUVECs was examined via CCK-8 assay. 5×103 cells were added into 96-well plates and incubated for 24, 48 and 72 h. Then, 10 μL CCK-8 reagent (Apexbio, Houston, TX, USA) was introduced and the cells were maintained for another 4 h. The optical density (OD) value was measured through a microplate reader (Bio-Gene Technology, Guangzhou, China). The detection wavelength was 450 nm.
Colony Formation Assay
Huh7 cells or HUVECs (500 cells/well) were placed into 6-well plates and cultured for 2 weeks. The clones were dyed with 0.5% crystal violet (Beyotime, Shanghai, China), and then observed under a microscope (Agilent, Chengdu, China).
Flow Cytometry
Cell apoptosis and cycle distribution were analyzed via flow cytometry. For cell apoptosis assay, Huh7 cells or HUVECs (2×105 cells/well) were placed into 12-well plates, and cultured for 72 h. Next, cells were incubated in the binding buffer and interacted with Annexin V-FITC and PI (Solarbio) in the dark. The apoptotic cells were analyzed using a flow cytometer (BD Biosciences, San Jose, CA, USA).
For the cell cycle assay, 5×105 cells were placed into 6-well plates, and cultured for 72 h. Next, cells were fixed with 70% ethanol and then treated via 50 μg/mL PI and RNase A for 30 min. The cycle distribution was analyzed with a flow cytometer.
Transwell Analysis
The migrated and invasive abilities of Huh7 cells or HUVECs were analyzed via 24-well transwell chambers (Costar, Corning, NY, USA). The transwell invasion assay was conducted using chambers precoated with Matrigel, while the transwell migration assay was conducted using those without Matrigel. 1×105 cells were resuspended in medium without serum and added into the upper chambers. The medium containing 10% serum was injected into the lower chambers. After 24 h, cells on the low chambers were incubated with 0.5% crystal violet. The number of migrated or invasive cells was counted under a microscope (Agilent) with three random fields.
Western Blot
The protein was isolated with RIPA buffer (Beyotime) containing 1 mM PMSF (Beyotime), and quantified using a protein quantification kit (Abbkine, Redlands, CA, USA). 20 μg samples were separated via sodium dodecyl sulfate-polyacrylamide gel electrophoresis and transferred onto polyvinylidene fluoride membranes (Solarbio). Following blocking in 5% nonfat milk, the membranes were interacted with specific primary antibodies and secondary antibody. The antibodies were shown as: anti-TSG101 (ab30871, 1:500 dilution; Abcam, Cambridge, MA, USA), anti-CD63 (ab216130, 1:2000 dilution; Abcam), anti-HSP70 (ab31010, 1:1000 dilution; Abcam), anti-CD81 (ab109201, 1:5000 dilution; Abcam), anti-caspase 3 (total caspase 3 and Cleaved caspase 3) (ab184787, 1:2000 dilution; Abcam), anti-PCNA (ab152112, 1:2000 dilution; Abcam), anti-Cyclin D1 (ab226977, 1:2000 dilution; Abcam), anti-MMP9 (ab73734, 1:1000 dilution; Abcam), anti-SP1 (ab227383, 1:500 dilution; Abcam) and anti-GAPDH (ab9485, 1:2000 dilution; Abcam), as well as horseradish peroxidase-conjugated IgG (ab205718, 1:20,000 dilution; Abcam). GAPDH was used as a reference. Next, the bands were visualized using the BeyoECL Plus kit (Beyotime). The relative protein level was normalized to the control group.
Dual-Luciferase Reporter Analysis and RNA Immunoprecipitation (RIP)
The complementary sequence of miR-638 and SP1 was predicted using starBase (http://starbase.sysu.edu.cn/index.php). The 3ʹUTR sequence of SP1 containing miR-638 binding sites was inserted into the luciferase reporter vector psiCHECK-2 (Promega, Madison, WI, USA) to synthesize the wild-type luciferase constructs SP1-wt. The mutant-type constructs were generated via mutating the binding sites, named as SP1-mut. Huh7 cells were co-transfected with 600 ng constructs and miR-638 mimic or NC for 24 h. Subsequently, luciferase activity was determined with a dual-luciferase assay kit (Promega).
For RIP assay, the RNA-Binding Protein Immunoprecipitation kit (Sigma) was used. Huh7 cells transfected with miR-638 mimic or NC were lysed, and lysates were incubated with magnetic beads coated with Ago2 antibody. IgG antibody was used as negative control. SP1 mRNA was extracted from beads, and examined via qRT-PCR.
Xenograft Experiment
Male BALB/c nude mice (5 weeks old) were provided via Beijing Laboratory Animal Center (Beijing, China) and acclimatized for 1 week. Huh7 cells (2×106 cells/mouse) were subcutaneously inoculated into mice. After 9 days, mice with the formed tumors were divided into two groups (n=5 per group). 5 nM miR-638 agomir (miR40003308-4-5, Ribobio) or NC was injected into the sites of tumors every other day for 4 times. Tumor volume was measured before the injection of agomir. At 3 days after the last injection, animals were killed via cervical dislocation under anesthesia using isoflurane. Tumor tissues were collected and the weight was detected. Furthermore, the levels of miR-638, SP1, PCNA, total caspase 3 and Cleaved caspase 3 were detected in tumor tissues. The animal procedures were performed in accordance with the Guidelines for Care and Use of Laboratory Animals of “National Institutes of Health” and approved via the Institutional Animal Care and Use Committee of Jinzhou Medical University.
Statistical Analysis
All experiments were carried out 3 times. Statistical analysis was conducted using GraphPad Prism 7 (GraphPad, La Jolla, CA, USA). The data were normally distributed and shown as mean±SD. The linear correlation of miR-638 and SP1 expression was analyzed via Pearson correlation analysis. The difference between groups was compared via Student’s t-test or ANOVA followed via Tukey’s test. The difference was statistically significant when P<0.05.
Results
miR-638 Level is Decreased in HCC Exosomes
To identify the exosomal miRNAs in HCC, the exosomes were purified from serum of HCC patients or normal subjects. As shown in Figure 1A and B, the exosomes were secreted without an obvious difference in serum of the two groups, and the diameters of exosomes from HCC serum were mainly between 80 and 120 nm. Meanwhile, the exosomes were also confirmed by the presence of exosome-specific markers (TSG101, CD63, HSP70 and CD81) (Figure 1C). Moreover, the miR-638 level was significantly decreased in HCC serum-derived exosomes in comparison to the normal group (Figure 1D). In addition, the miR-638 level was lower in HCC tissues than para-tumor samples (Figure 1E). Besides, the abundance of miR-638 was also detected in HCC cells or medium-derived exosomes. Results exhibited that the miR-638 level was remarkably reduced in HCC cells or exosomes (Figure 1F and G). These data indicated that a low level of exosomal miR-638 was displayed in HCC serum and cell medium. Furthermore, low expression of miR-638 was associated with Edmondson grade, but not with age, gender, tumor size, hepatitis and AFP level (Table 1).
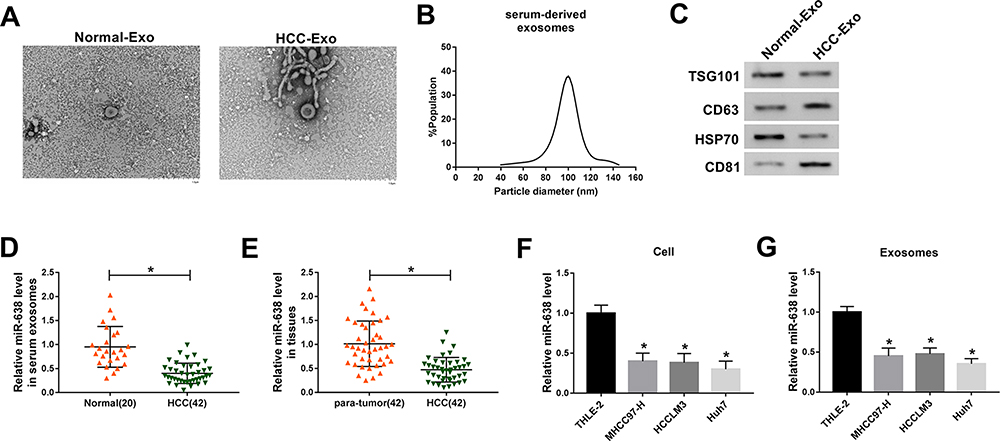 |
Figure 1 The level of miR-638 in HCC. (A) Representative graphs of HCC-Exo and Normal-Exo were provided via TEM. (B) The particle size of HCC-Exo was analyzed. (C) The protein levels of TSG101, CD63, HSP70 and CD81 were detected in HCC-Exo and Normal-Exo by western blot. (D) The level of miR-638 was measured in serum exosomes from HCC patients and normal subjects via qRT-PCR. (E) miR-638 level was detected in HCC and para-tumor tissues. (F and G) miR-638 level was detected in HCC and normal liver cells or exosome. *P<0.05. |
Up-Regulation of miR-638 Suppresses Proliferation, Migration and Invasion of HCC Cells
To explore the role of miR-638 in HCC cells, Huh7 cells with the relative lowest abundance of miR-638 were chosen for further experiments, and cells were transfected with 50 or 100 nM miR-638 mimic or NC. As displayed in Figure 2A and B, cell viability and colony formation were evidently decreased via transfection of miR-638 mimic in a concentration-dependent manner. Furthermore, flow cytometry analysis showed that transfection of different concentrations of miR-638 mimic induced cell apoptosis and cell cycle arrest at G1 phase (Figure 2C and D). In addition, the migrated and invasive abilities of Huh7 cells were remarkably inhibited via different concentrations of miR-638 mimic (Figure 2E and F). Besides, the related protein expression was measured, such as PCNA (promoting proliferation),23 Cyclin D1 (regulating cell cycle from G1 to S phase),24 Cleaved caspase 3 (promoting apoptosis)25 and MMP9 (promoting migration and invasion).26 Results showed that elevation of Cleaved caspase 3 and reduction of PCNA, Cyclin D1 and MMP9 were induced by transfection of miR-638 mimic in Huh7 cells (Figure 2G). These results suggested that miR-638 overexpression using miR-638 mimic repressed HCC cell proliferation, migration and invasion.
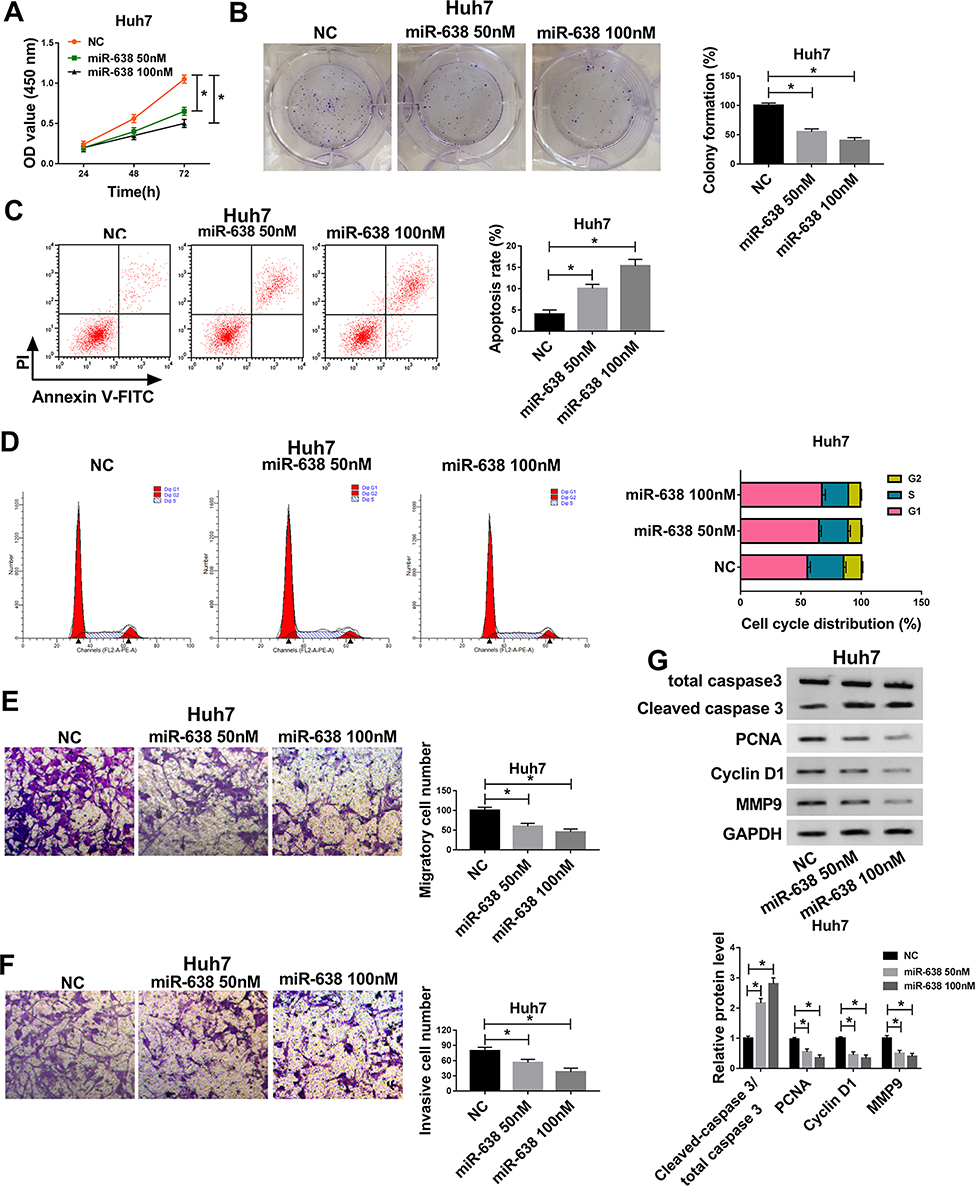 |
Figure 2 The effect of miR-638 on HCC cell proliferation, migration and invasion. Cell viability (A), colony formation (B), apoptotic rate (C), cell cycle distribution (D), migration (E), invasion (F) and related protein levels (G) were measured in Huh7 cells with transfection of 50 or 100 nM miR-638 mimic or NC via CCK-8, colony formation assay, flow cytometry, transwell and western blot. *P<0.05. |
Exosomal miR-638 Inhibits Endothelial Function
Endothelial function is responsible for tumor angiogenesis in HCC. To assess the influence of exosomal miR-638 on endothelial function, Huh7 cells were transfected with miR-638 mimic or NC, and exosomes in medium were collected and used for co-culture with HUVECs. As displayed in Figure 3A–D, Huh7 cell-derived exosomal miR-638 obviously repressed HUVEC proliferation via decreasing viability and colony formation and promoting apoptosis and affecting cell cycle distribution, while these events were abrogated via GW4869, an inhibitor of exosome secretion. After the treatment of GW4869, the appearance of exosome and miR-638 expression were reduced (Supplementary Figure 1A and B). Moreover, Huh7 cell-derived exosomal miR-638 significantly inhibited HUVEC migration and invasion, which was reversed by GW4869 (Figure 3E and F). Besides, Huh7 cell-derived exosomal miR-638 evidently enhanced the level of Cleaved caspase 3 and declined the protein levels of PCNA, Cyclin D1 and MMP9 in HUVECs, and treatment of GW4869 abolished this effect (Figure 3G). These findings indicated that exosomal miR-638 might inhibit HCC angiogenesis by regulating endothelial function.
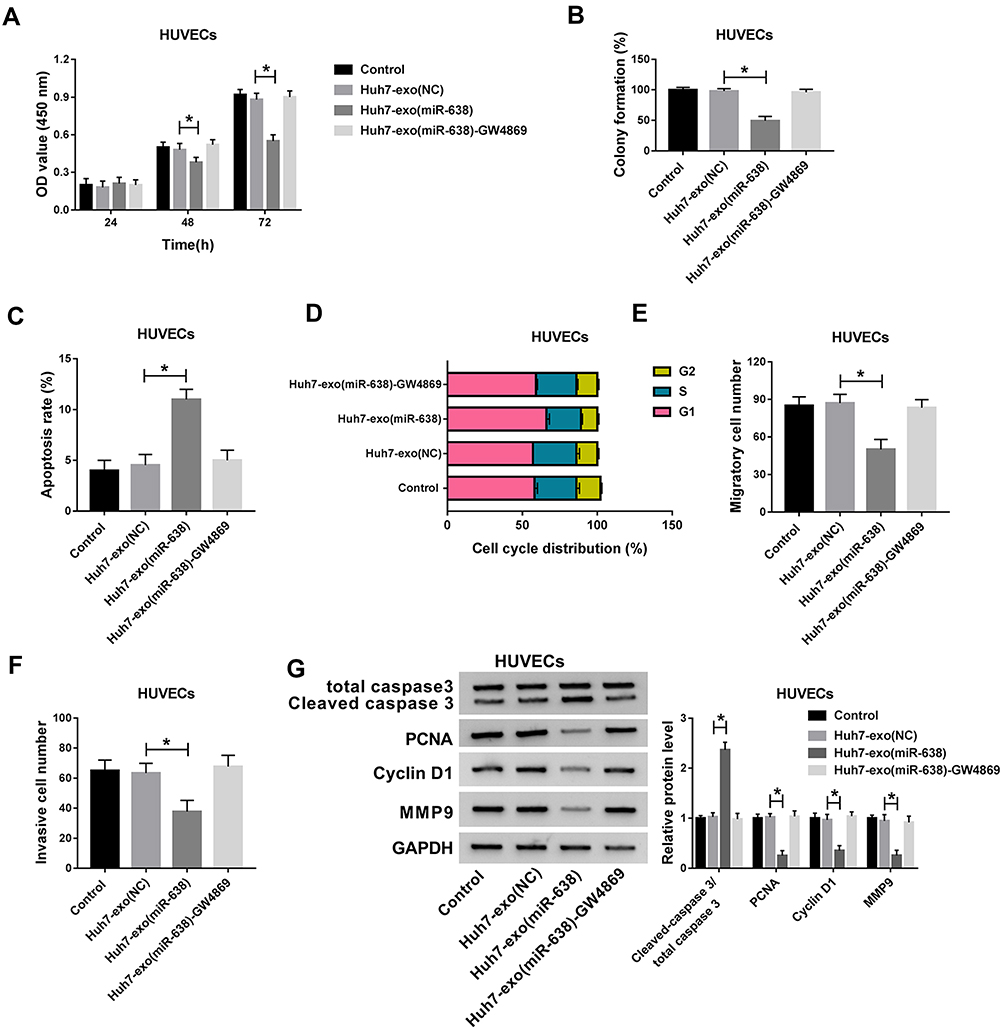 |
Figure 3 The effect of exosomal miR-638 on endothelial function. Cell viability (A), colony formation (B), apoptotic rate (C), cell cycle distribution (D), migration (E), invasion (F) and related protein levels (G) were measured in HUVECs after treatment of Huh7-exo(NC), Huh7-exo(miR-638) or Huh7-exo(NC)-GW4869 via CCK-8, colony formation assay, flow cytometry, transwell and western blot. *P<0.05. |
SP1 is a Target of miR-638 in HCC Cells
To explore the mechanism of miR-638 in HCC, the targets of miR-638 were predicted via starBase. Among these targets, SP1 expression was enhanced at mRNA and protein levels and negatively correlated with miR-638 level in HCC tissues (Figure 4A–C). Moreover, SP1 protein expression was evidently elevated in HCC cells (Figure 4D). To further confirm the target association between miR-638 and SP1, SP1-wt and SP1-mut were constructed (Figure 4E), and used for dual-luciferase reporter analysis. Results displayed that addition of miR-638 induced a 76% reduction in luciferase activity of SP1-wt, but its effect was abolished via mutating the binding sites in SP1-mut group (Figure 4F). Additionally, Ago2 RIP assay showed that SP1 could be bound to miR-638 (Figure 4G). Besides, SP1 protein expression in Huh7 cells was significantly decreased via transfection of miR-638 mimic (Figure 4H). These data suggested that SP1 could be targeted via miR-638 in HCC.
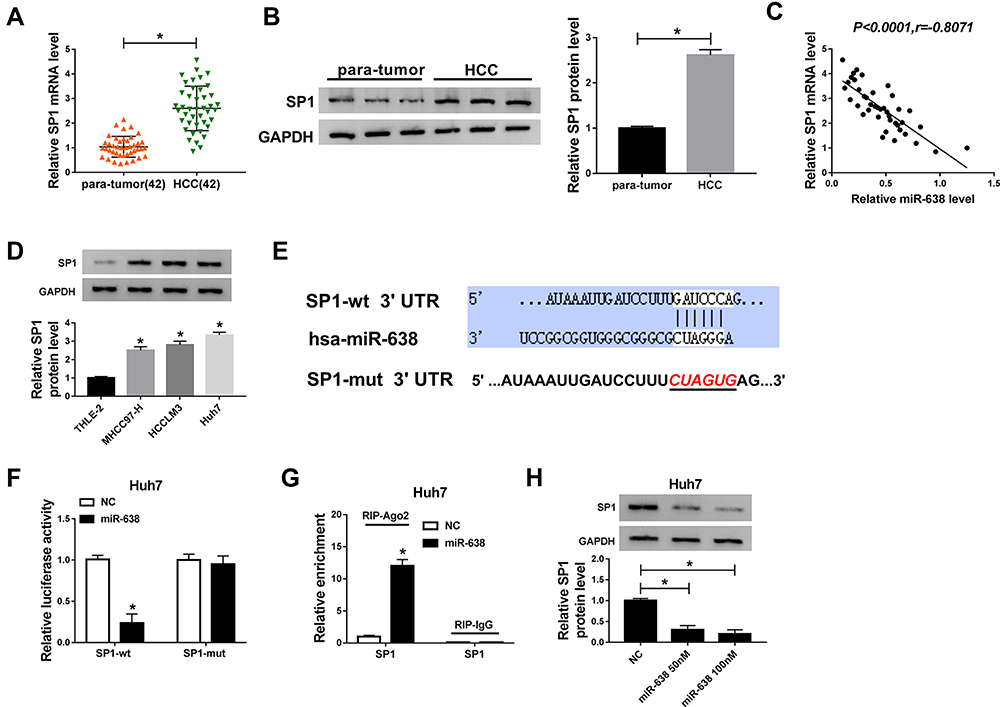 |
Figure 4 The association between miR-638 and SP1 in HCC. (A and B) SP1 mRNA and protein levels in HCC and para-tumor tissues were examined via qRT-PCR and western blot. (C) The linear correlation between expression of miR-638 and SP1 in HCC samples was analyzed. (D) SP1 protein expression was detected in HCC and normal liver cells. (E) The complementary sequence between miR-638 and SP1 was predicted via starBase. (F and G) Dual-luciferase reporter and RIP assays were conducted in Huh7 cells with transfection of miR-638 mimic or NC. (H) SP1 protein expression was detected in Huh7 cells transfected with 50 or 100 nM miR-638 mimic or NC. *P<0.05. |
Addition of SP1 Reverses the Suppressive Effect of miR-638 on Proliferation, Migration and Invasion of HCC Cells
To explore whether SP1 could explain the influence of miR-638 in HCC cells, Huh7 cells were transfected with NC, miR-638 mimic, miR-638 mimic+vector or SP1 overexpression vector. As exhibited in Figure 5A, introduction of SP1 overexpression vector restored the abundance of SP1 protein in Huh7 cells in the presence of miR-638 mimic. Furthermore, up-regulation of SP1 attenuated miR-638-mediated proliferation reduction by affecting cell viability, colony formation, apoptosis and cell cycle distribution (Figure 5B–E). In addition, restoration of SP1 weakened the inhibitive effect of miR-638 on Huh7 cell migration and invasion (Figure 5F and G). Besides, overexpression of SP1 alleviated the effect of miR-638 on protein levels of Cleaved caspase 3, PCNA, Cyclin D1 and MMP9 in Huh7 cells (Figure 5H). These findings showed that miR-638 repressed HCC cell proliferation, migration and invasion via decreasing SP1.
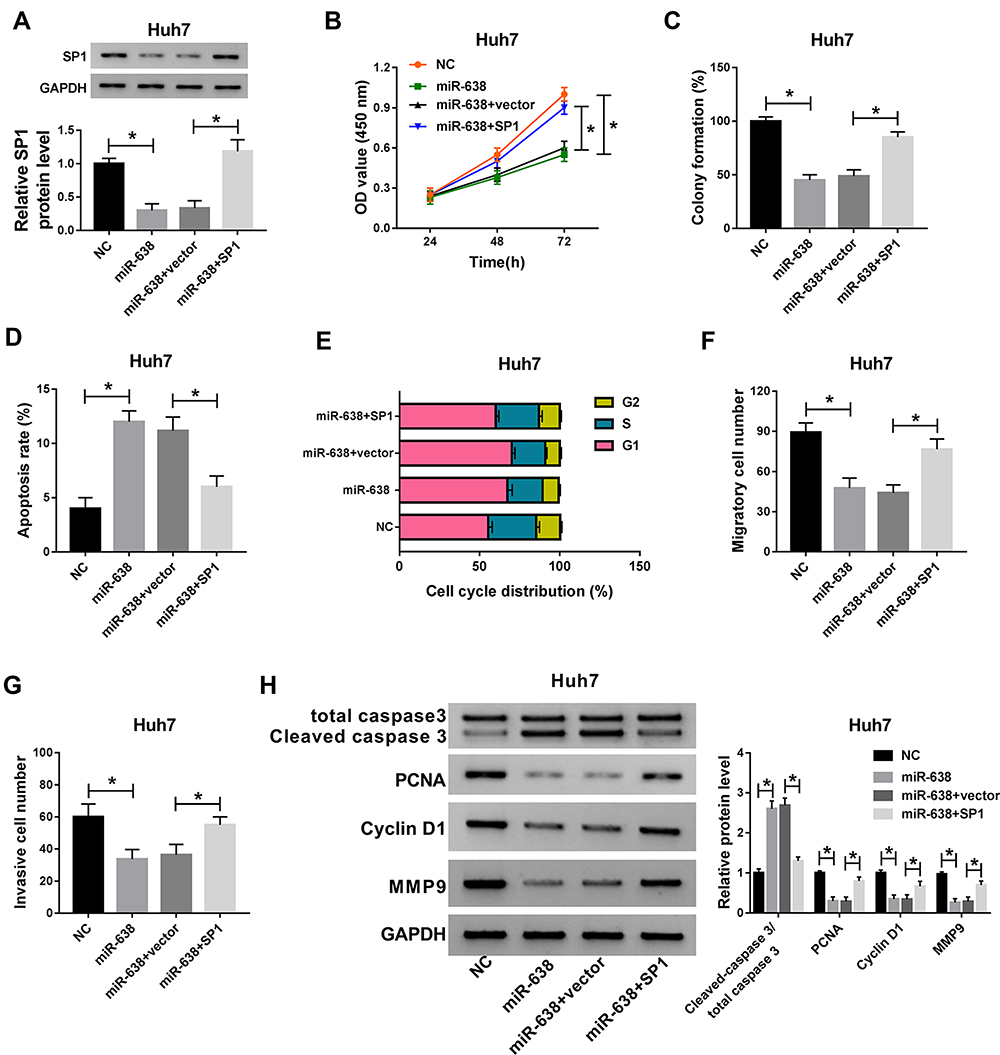 |
Figure 5 The effect of miR-638 and SP1 on HCC cell proliferation, apoptosis, migration and invasion. SP1 protein expression (A), cell viability (B), colony formation (C), apoptotic rate (D), cell cycle distribution (E), migration (F), invasion (G) and related protein abundances (H) were examined in Huh7 cells transfected with NC, 100 nM miR-638 mimic, miR-638 mimic+vector or SP1. *P<0.05. |
miR-638 Decreases Xenograft Tumor Growth
To analyze the role of miR-638 in HCC in vivo, Huh7 cells were used to establish the subcutaneous xenograft model for 9 days, and then tumor sites were treated with miR-638 agomir or NC every other day. As shown in Figure 6A, tumor volume was obviously reduced after the injection of miR-638 agomir for 5, 7 and 10 days. Moreover, the tumor tissues were collected at the ending point. Tumor weight was markedly decreased by treatment of miR-638 agomir (Figure 6B). In addition, miR-638 expression and Cleaved caspase 3 protein level were significantly increased, and protein levels of SP1 and PCNA were remarkably decreased by treatment of miR-638 agomir (Figure 6C and D). These findings suggested that miR-638 overexpression decreased HCC development in vivo. A pathway illustration of miR-638/SP1 in HCC development is exhibited in Figure 7.
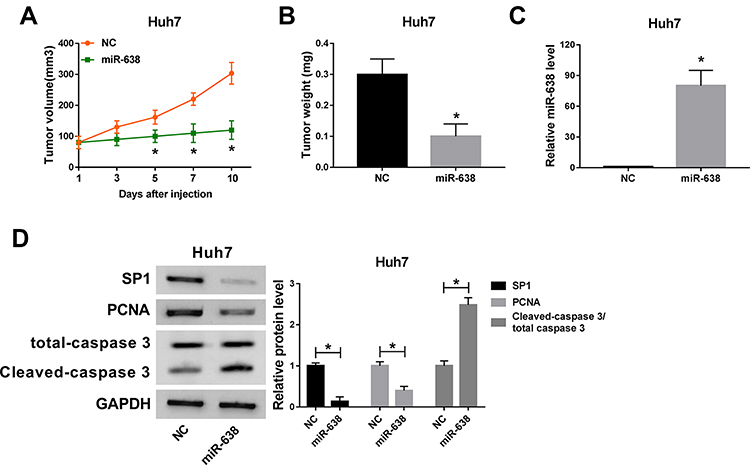 |
Figure 6 The effect of miR-638 on xenograft tumor growth. 2×106 Huh7 cells were subcutaneously injected into nude mice for 9 days, and then mice were treated with miR-638 agomir or NC every other day. n=5 per group. (A and B) Tumor volume and weight were measured in each group. (C and D) miR-638 expression and protein levels of SP1, PCNA, total caspase 3 and Cleaved caspase 3 were detected in each group. *P<0.05. |
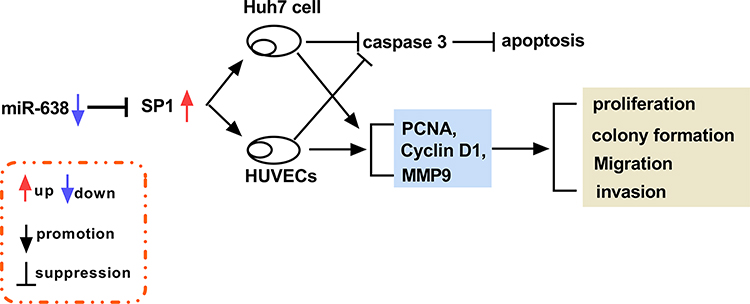 |
Figure 7 A schematic diagram of the miR-638/SP1 axis in HCC development. |
Discussion
HCC is a common cancer with high incidence and mortality, and the effective treatment options for HCC are limited.27 Exosomes play important roles in the pathogenesis, diagnosis, development and treatment of HCC.28 Moreover, exosomal miRNAs are implicated in the anti-cancer treatment of HCC.29 In our research, the exosomal miR-638 level was reduced in HCC, and exosomal miR-638 inhibited HCC cell proliferation, migration and invasion. Furthermore, here we were the first to confirm the target relationship of miR-638 and SP1 (Figure 7).
The tetraspanins (CD63 and CD81), heat shock protein HSP70 and tumor susceptibility gene TSG101 are the important markers for validating the presence of exosomes.21 By detecting these marker levels and TEM, we confirmed that the extracted vesicles were exosomes. Our study displayed that miR-638 abundance was reduced in HCC tissues and cells. We hypothesized the reduced miR-638 might be induced via alteration of the microenvironment. In HCC, the environment might be adverse to miR-638 synthesis or contribute to miR-638 degeneration. A previous study suggested that miR-638 could be released in exosomes from HCC cells.11 In this work, exosomal miR-638 was declined in HCC from serum or cells, which was also in agreement with former work.13 Meanwhile, that report indicated that exosomal miR-638 could inhibit HCC cell proliferation.13 Similarly, we also found that exosomal miR-638 suppressed HCC cell proliferation via restraining cell viability, colony formation, facilitating apoptosis and inducing cell cycle arrest at S1 phase. Moreover, this study showed that miR-638 could repress HCC cell migration and invasion, which was also consistent with former work.10 In addition, a previous study indicated that inhibition of miR-638 facilitated the angiogenesis in HCC.9 Endothelial function is implicated in the angiogenesis, which contributes to tumorigenesis of HCC.30–32 Hence, we further assessed the influence of exosomal miR-638 on endothelial function. Using co-culture of Huh7 and HUVECs, we validated that miR-638 from Huh7 cells could be transmitted to HUVECs by exosomes, and then affected HUVEC function. Collectively, this study showed the anti-cancer role of exosomal miR-638 in HCC by inhibiting cell proliferation, migration, invasion and angiogenesis.
miRNAs act as gene regulators by binding the 3ʹUTR of mRNA to mediate mRNA degradation or translation.33 Increasing evidence demonstrated that miR-638 could participate in the regulation of multiple cancers by targeting numerous mRNAs because of the different binding sites, such as phospholipase D1, tetraspanin 1 and sex-determining region Y-box 2.34–36 To explore a new mechanism mediated via miR-638, here we were the first to identify that SP1 was targeted via miR-638. Previous studies reported that SP1 played an oncogenic role in HCC via promoting cell proliferation, migration and invasion.16,17,37,38 These reports also suggested that miR-31-5p, miR-138, miR-363-3p and miR-548b could target SP1 to participate in HCC development, while the expression changes of these miRNAs in exosomes were less than miR-638 (Supplementary Figure 2). Furthermore, our study also confirmed the carcinogenic role of SP1 in HCC by rescue experiments. These data also indicated that miR-638 could inhibit HCC development by targeting SP1 in vitro. Besides, the xenograft model is a key tool for understanding the pathogenesis of HCC.39 In this research, we also confirmed the anti-cancer role of miR-638 in HCC in vivo using a murine xenograft model. The major function of HUVECs is the ability of vessel formation. Previous studies have reported that SP1 could promote the ability of tube formation of endothelial cells. Thus, we hypothesized that miR-638 might inhibit tube formation of HUVECs via targeting SP1. This would be explored in the future.40,41 Although our study demonstrated the function and mechanism of exosomal miR-638 in HCC using Huh7 cells, more cell lines should be used for the study in the future due to the difference of different cell lines.
In conclusion, exosomal miR-638 played an anti-cancer role in HCC in vitro and in vivo, possibly via targeting SP1. This study elucidated a novel mechanism underlying the pathogenesis of HCC and indicated the potential significance of miR-638 in treatment of HCC.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Llovet JM, Zucman-Rossi J, Pikarsky E, et al. Hepatocellular carcinoma. Nat Rev Dis Primers. 2016;2(1):16018. doi:10.1038/nrdp.2016.18
2. Zhu J, Yin T, Xu Y, et al. Therapeutics for advanced hepatocellular carcinoma: recent advances, current dilemma, and future directions. J Cell Physiol. 2019;234(8):12122–12132. doi:10.1002/jcp.28048
3. Wu Q, Zhou L, Lv D, et al. Exosome-mediated communication in the tumor microenvironment contributes to hepatocellular carcinoma development and progression. J Hematol Oncol. 2019;12(1):53. doi:10.1186/s13045-019-0739-0
4. Wang H, Lu Z, Zhao X. Tumorigenesis, diagnosis, and therapeutic potential of exosomes in liver cancer. J Hematol Oncol. 2019;12:133. doi:10.1186/s13045-019-0806-6
5. Li C, Xu X. Biological functions and clinical applications of exosomal non-coding RNAs in hepatocellular carcinoma. Cell Mol Life Sci. 2019;76(21):4203–4219. doi:10.1007/s00018-019-03215-0
6. Li S, Yao J, Xie M, et al. Exosomal miRNAs in hepatocellular carcinoma development and clinical responses. J Hematol Oncol. 2018;11(1):54. doi:10.1186/s13045-018-0579-3
7. Cheng J, Chen Y, Zhao P, et al. Dysregulation of miR-638 in hepatocellular carcinoma and its clinical significance. Oncol Lett. 2017;13:3859–3865. doi:10.3892/ol.2017.5882
8. Ye W, Li J, Fang G, et al. Expression of microRNA 638 and sex-determining region Y-box 2 in hepatocellular carcinoma: association between clinicopathological features and prognosis. Oncol Lett. 2018;15:7255–7264. doi:10.3892/ol.2018.8208
9. Cheng J, Chen Y, Zhao P, et al. Downregulation of miRNA-638 promotes angiogenesis and growth of hepatocellular carcinoma by targeting VEGF. Oncotarget. 2016;7(21):30702–30711. doi:10.18632/oncotarget.8930
10. Zhang Y, Zhang D, Jiang J, et al. Loss of miR-638 promotes invasion and epithelial-mesenchymal transition by targeting SOX2 in hepatocellular carcinoma. Oncol Rep. 2017;37(1):323–332. doi:10.3892/or.2016.5273
11. Kubota S, Chiba M, Watanabe M, et al. Secretion of small/microRNAs including miR-638 into extracellular spaces by sphingomyelin phosphodiesterase 3. Oncol Rep. 2015;33(1):67–73. doi:10.3892/or.2014.3605
12. Yan S, Dang G, Zhang X, et al. Downregulation of circulating exosomal miR-638 predicts poor prognosis in colon cancer patients. Oncotarget. 2017;8(42):72220–72226. doi:10.18632/oncotarget.19689
13. Shi M, Jiang Y, Yang L, et al. Decreased levels of serum exosomal miR-638 predict poor prognosis in hepatocellular carcinoma. J Cell Biochem. 2018;119(6):4711–4716. doi:10.1002/jcb.26650
14. Giordano S, Columbano A. MicroRNAs: new tools for diagnosis, prognosis, and therapy in hepatocellular carcinoma? Hepatology. 2013;57(2):840–847. doi:10.1002/hep.26095
15. Beishline K, Azizkhan-Clifford J. Sp1 and the ‘hallmarks of cancer’. FEBS J. 2015;282:224–258. doi:10.1111/febs.13148
16. Zhao G, Han C, Zhang Z, et al. Increased expression of microRNA-31-5p inhibits cell proliferation, migration, and invasion via regulating Sp1 transcription factor in HepG2 hepatocellular carcinoma cell line. Biochem Biophys Res Commun. 2017;490(2):371–377. doi:10.1016/j.bbrc.2017.06.050
17. Liu C, Zhu J, Liu F, et al. MicroRNA-138 targets SP1 to inhibit the proliferation, migration and invasion of hepatocellular carcinoma cells. Oncol Lett. 2018;15(1):1279–1286. doi:10.3892/ol.2017.7357
18. Zou Y, Xiong H, Xiong H, et al. A polysaccharide from mushroom huaier retards human hepatocellular carcinoma growth, angiogenesis, and metastasis in nude mice. Tumour Biol. 2015;36(4):2929–2936. doi:10.1007/s13277-014-2923-8
19. Yu B, Du Q, Li H, et al. Diagnostic potential of serum exosomal colorectal neoplasia differentially expressed long non-coding RNA (CRNDE-p) and microRNA-217 expression in colorectal carcinoma. Oncotarget. 2017;8(48):83745–83753. doi:10.18632/oncotarget.19407
20. Zheng R, Du M, Wang X, et al. Exosome–transmitted long non-coding RNA PTENP1 suppresses bladder cancer progression. Mol Cancer. 2018;17(1):143. doi:10.1186/s12943-018-0880-3
21. Ekstrom K, Omar O, Graneli C, et al. Monocyte exosomes stimulate the osteogenic gene expression of mesenchymal stem cells. PLoS One. 2013;8(9):e75227. doi:10.1371/journal.pone.0075227
22. Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods. 2001;25(4):402–408. doi:10.1006/meth.2001.1262
23. Wang SC. PCNA: a silent housekeeper or a potential therapeutic target? Trends Pharmacol Sci. 2014;35:178–186. doi:10.1016/j.tips.2014.02.004
24. Qie S, Diehl JA. Cyclin D1, cancer progression, and opportunities in cancer treatment. J Mol Med (Berl). 2016;94(12):1313–1326. doi:10.1007/s00109-016-1475-3
25. Nagata S. Apoptosis and clearance of apoptotic cells. Annu Rev Immunol. 2018;36(1):489–517. doi:10.1146/annurev-immunol-042617-053010
26. Huang H. Matrix metalloproteinase-9 (MMP-9) as a cancer biomarker and MMP-9 biosensors: recent advances. Sensors (Basel). 2018;18(10):3249. doi:10.3390/s18103249
27. Bupathi M, Kaseb A, Meric-Bernstam F, et al. Hepatocellular carcinoma: where there is unmet need. Mol Oncol. 2015;9(8):1501–1509. doi:10.1016/j.molonc.2015.06.005
28. Moris D, Beal EW, Chakedis J, et al. Role of exosomes in treatment of hepatocellular carcinoma. Surg Oncol. 2017;26(3):219–228. doi:10.1016/j.suronc.2017.04.005
29. Pan J-H, Zhou H, Zhao -X-X, et al. Role of exosomes and exosomal microRNAs in hepatocellular carcinoma: potential in diagnosis and antitumour treatments (Review). Int J Mol Med. 2018;41(4):1809–1816. doi:10.3892/ijmm.2018.3383
30. Liu P, Atkinson SJ, Akbareian SE, et al. Sulforaphane exerts anti-angiogenesis effects against hepatocellular carcinoma through inhibition of STAT3/HIF-1alpha/VEGF signalling. Sci Rep. 2017;7:12651. doi:10.1038/s41598-017-12855-w
31. Morse MA, Sun W, Kim R, et al. The role of angiogenesis in hepatocellular carcinoma. Clin Cancer Res. 2019;25(3):912–920. doi:10.1158/1078-0432.CCR-18-1254
32. Zhao S, Li J, Zhang G, et al. Exosomal miR-451a functions as a tumor suppressor in hepatocellular carcinoma by targeting LPIN1. Cell Physiol Biochem. 2019;53:19–35. doi:10.33594/000000118
33. Li D, Zhang J, Li J. Role of miRNA sponges in hepatocellular carcinoma. Clin Chim Acta. 2020;500:10–19. doi:10.1016/j.cca.2019.09.013
34. Tang KL, Tang HY, Du Y, et al. MiR-638 suppresses the progression of oral squamous cell carcinoma through wnt/beta-catenin pathway by targeting phospholipase D1. Artif Cells Nanomed Biotechnol. 2019;47:3278–3285. doi:10.1080/21691401.2019.1647222
35. Zhang J, Fei B, Wang Q, et al. MicroRNA-638 inhibits cell proliferation, invasion and regulates cell cycle by targeting tetraspanin 1 in human colorectal carcinoma. Oncotarget. 2014;5(23):12083–12096. doi:10.18632/oncotarget.2499
36. Xia Y, Wu Y, Liu B, et al. Downregulation of miR-638 promotes invasion and proliferation by regulating SOX2 and induces EMT in NSCLC. FEBS Lett. 2014;588(14):2238–2245. doi:10.1016/j.febslet.2014.05.002
37. Ying J, Yu X, Ma C, et al. MicroRNA-363-3p is downregulated in hepatocellular carcinoma and inhibits tumorigenesis by directly targeting specificity protein 1. Mol Med Rep. 2017;16(2):1603–1611. doi:10.3892/mmr.2017.6759
38. Qiu H, Zhang G, Song B, et al. MicroRNA-548b inhibits proliferation and invasion of hepatocellular carcinoma cells by directly targeting specificity protein 1. Exp Ther Med. 2019;18:2332–2340. doi:10.3892/etm.2019.7812
39. Fornari F, Gramantieri L, Callegari E, et al. MicroRNAs in animal models of HCC. Cancers (Basel). 2019;11. doi:10.3390/cancers11121906
40. Xie J, Gong Q, Liu X, et al. Transcription factor SP1 mediates hyperglycemia-induced upregulation of roundabout4 in retinal microvascular endothelial cells. Gene. 2017;616:31–40. doi:10.1016/j.gene.2017.03.027
41. Jiang Y, Liu H, Liu WJ, et al. Endothelial Aquaporin-1 (AQP1) expression is regulated by transcription factor Mef2c. Mol Cells. 2016;39(4):292–298. doi:10.14348/molcells.2016.2223
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.