Back to Journals » Clinical, Cosmetic and Investigational Dermatology » Volume 16
Exogenous Hydrogen Sulfide Induces A375 Melanoma Cell Apoptosis Through Overactivation of the Unfolded Protein Response
Authors Ma W ![]() , Zhang X, Zhuang L
, Zhang X, Zhuang L
Received 16 April 2023
Accepted for publication 10 June 2023
Published 27 June 2023 Volume 2023:16 Pages 1641—1651
DOI https://doi.org/10.2147/CCID.S412588
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Jeffrey Weinberg
Weiyuan Ma,1 Xiuwen Zhang,2 Le Zhuang3
1Department of Dermatology, Affiliated Hospital of Weifang Medical University, Weifang, Shandong Province, People’s Republic of China; 2Department of Dermatology, Weihai Municipal Hospital, Weihai, Shandong Province, People’s Republic of China; 3Department of Dermatology, Central Hospital Affiliated to Shandong First Medical University, Jinan, Shandong Province, People’s Republic of China
Correspondence: Le Zhuang, Department of Dermatology, Central Hospital Affiliated to Shandong First Medical University, No.105, Jiefang Road, Lixia District, Jinan, Shandong Province, 250013, People’s Republic of China, Tel +8615966301378, Fax +86053185695114, Email [email protected]
Purpose: Melanomas are highly malignant and rapidly develop drug resistance due to dysregulated apoptosis. Therefore, pro-apoptotic agents could be effective for the management of melanoma. Hydrogen sulfide is ubiquitous in the body, and exogenous hydrogen sulfide has been reported to show inhibitory and pro-apoptotic effects on cancer cells. However, whether high concentrations of exogenous hydrogen sulfide have pro-apoptotic effects on melanoma and its mechanisms remain unknown. Hence, this study aimed to explore the pro-apoptotic effects and mechanisms of exogenous hydrogen sulfide on the A375 melanoma cell line treated with a hydrogen sulfide donor (NaHS).
Methods: The cell proliferation test, flow cytometric analysis, Hoechst 33258 staining, and Western blotting of B-cell lymphoma 2 and cleaved caspase-3 were used to explore the pro-apoptotic effects of hydrogen sulfide on A375 cells. The transcriptional profile of NaHS-treated A375 cells was further explored via high-throughput sequencing. Western blotting of phosphorylated inositol-requiring enzyme 1α (p‐IRE1α), phosphorylated protein kinase R-like ER kinase (p-PERK), phosphorylated eukaryotic translation initiation factor 2α (p-eIF2α), C/EBP homologous protein, glucose-regulating protein 78, IRE1α, PERK, and eIF2α was performed to verify the changes in the transcriptional profile.
Results: NaHS inhibited A375 melanoma cell proliferation and induced apoptosis. The endoplasmic reticulum stress unfolded protein response and apoptosis-associated gene expression was upregulated in NaHS-treated A375 melanoma cells. The overactivation of the unfolded protein response and increase in endoplasmic reticulum stress was verified at the protein level.
Conclusion: Treatment with NaHS increased endoplasmic reticulum stress, which triggered the overactivation of the unfolded protein response and ultimately lead to melanoma cell apoptosis. The pro-apoptotic effect of NaHS suggests that it can be explored as a potential therapeutic agent in melanoma.
Keywords: hydrogen sulfide, melanoma, apoptosis, unfolded proteins response, endoplasmic reticulum stress
Introduction
Melanoma is caused by the malignant transformation of melanocytes and is a fatal type of skin cancer that is resistant to chemo- and radiotherapy.1 Each year, approximately 232,100 individuals are diagnosed with cutaneous melanomas worldwide.2 About half of the patients diagnosed with melanomas have BRAF mutations.3 Combined treatment with BRAF inhibitors (such as dabrafenib and vemurafenib) and mitogen-activated protein kinase (MAPK) inhibitors (such as trametinib and cobimetinib) improves the treatment response and overall survival in patients with cutaneous melanoma.4 Recently, the development and application of immune checkpoint inhibitors, such as anti-cytotoxic T lymphocyte antigen-4 and anti-programmed cell death-1 antibodies, have been shown to prolong the overall survival of patients with melanoma.5 Despite the initial response, genetic mutations in multiple pathway molecules and intrinsic tumor factors or microenvironment features lead to a high proportion of melanoma patients developing resistance to targeted drugs and immune checkpoint inhibitors after a few months of treatment.6–9 Due to the high rate of disease recurrence and treatment resistance, melanoma patients succumb to the disease within a few months, resulting in approximately 55,500 melanoma-attributed deaths worldwide each year.2,10 The dysregulation of apoptosis is a major factor contributing to drug resistance in melanoma. A significant decrease in the rate of apoptosis was reported in melanoma cells resistant to chemotherapy and targeted drugs.11,12 The frequent anti-apoptotic phenomenon observed in melanoma patients implies that exploring pro-apoptotic agents might be a promising anti-cancer strategy.13–16 Therefore, further improvements, especially the development of highly effective drugs to induce a pro-apoptotic effect in melanoma, are warranted to improve the overall survival of patients with cutaneous melanoma.
Hydrogen sulfide (H2S) is the third most common gasotransmitter in the body apart from carbon monoxide (CO) and nitric oxide (NO).17 The three enzymes that can catalyze the generation of H2S in human cells are: 3-mercaptopyruvate sulfotransferase, cystathionine beta-synthase, and cystathionine gamma-lyase.18 H2S possesses functions in an assortment of physiological and pathological processes in human cells. It can relax the blood vessels and protect the neurons and myocardium from oxidative stress.19,20 Different doses of H2S have various effects on cancer. Low concentrations of H2S promotes cancer progression by increasing the expression of vascular endothelial growth factor and promoting angiopoiesis; preventing tumor cell apoptosis by decreasing the intracellular reactive oxygen species (ROS) levels; reducing DNA damage caused by mitochondrial disruption; and accelerating the cell cycle of cancer cells by regulating the phosphorylation level of the protein kinase B/Akt.21,22 In contrast, higher concentrations of exogenous H2S exhibited an inhibitory effect on cancer cells — it promoted uncontrolled intracellular acidification and accelerated apoptosis by inducing cell cycle arrest in human colon cancer cells.23 The H2S donor, ATB-346, significantly downregulated the expression of anti-apoptotic genes (eg, Fas-associated death domain-like interleukin-1β-converting enzyme inhibitory protein and B-cell lymphoma 2 [Bcl-2]), which lead to a pro-apoptotic effect in the cancer cells.24,25 Exogenous H2S increments the capacity of the anion and sodium exchangers to promote the generation of metabolic acids, which can lead to cancer cell death.23 The potential pro-apoptotic activity of H2S donors leads to the question of whether high concentrations of exogenous H2S exerts a pro-apoptotic effect on melanoma, and by which mechanism(s) is this accomplished.
Hence, this study aimed to investigate the pro-apoptotic effect of exogenous H2S on melanoma cells and its mechanism of action to provide new insights regarding alternative strategies for melanoma treatment. The effects of H2S on the proliferation and apoptosis of melanoma cells and the alterations in the transcriptome and corresponding biological processes in melanoma cell lines were examined after administering exogenous H2S. Furthermore, changes in the transcriptome of cells treated with H2S were verified at the protein level.
Materials and Methods
Cell Culture and Treatment
The human melanoma cell line, A375, was acquired from the Institute of Basic Medicine, Chinese Academy of Medical Sciences (Beijing, China). Cells were cultivated in Dulbecco’s Modified Eagle Medium (DMEM; 11995040, Gibco, NY, USA) supplemented with 10% fetal bovine serum (10099-141, Gibco, NY, USA) at 37°C in a humidified incubator with 5% CO2 and 95% air conditions. Salt of sodium hydrosulfide (NaHS; S106641, Aladdin, Shanghai, China) was dissolved in the cell culture medium (DMEM; 50 mL) to make the stock solution (concentration of 400 mM). The temperature and time at which the solution was stored was 4°C and one week. Immediately before experiments, the NaHS stock solution was diluted with DMEM to concentrations of 0, 0.5, 5, 10, 20, and 50 mM, and the A375 cells were treated for 24 h.
Cell Proliferation Analysis
A cell counting kit-8 (CCK-8, CK04, Dojindo Laboratory, Kyushu, Japan) was used according to the manufacturer’s instructions to evaluate cell proliferation at the indicated time points. NaHS-treated A375 cells and control cells were seeded at a density of 5×103 cells/well in 96-well plates in quintuplicate. The optical absorbance value of each well was determined at 450 nm on a microplate reader (iMark Reader, Bio-Rad, USA) after incubating with CCK-8 for 1 h at 37°C. The formula used to calculate the cell proliferation rate was: relative cell proliferation rate% = (450 nm sample – 450 nm background) / (450 nm control – 450 nm background) ×100%.
Hoechst 33258 Staining
Paraformaldehyde (4%) was used to fix 20 mM NaHS-treated cells and control cells for 10 min. The cells were then stained with 5 mg/L Hoechst 33258 for 10 min, after which they were observed under an Olympus fluorescent microscope (Tokyo, Japan). Hoechst 33258 (H266330, Aladdin, Shanghai, China) staining revealed changes in chromatin morphology. The apoptotic cell nuclei showed condensed, fractured fluorescence representing chromosomal condensation and fragmentation, while normal cells exhibited uniform fluorescence and a completely rounded nucleus.
Cell Apoptosis Analysis
NaHS-treated (20 mM) cells and control cells were harvested via trypsinization after treatment with NaHS for 24 h. The cells were resuspended in the binding buffer after washing twice with phosphate-buffered saline (PBS). Annexin V-FITC and propidium iodide solutions (C1067S, Beyotime Institute of Biotechnology, Shanghai, China) were added to the cell suspension. The cells were examined using flow cytometry (BD, Becton, USA) after incubating for 20 min at room temperature.
RNA Isolation and Sequencing
Total RNA from the NaHS-treated and control cells was isolated using the Qiagen RNeasy Plus Mini kit (74134, Hilden, Germany) according to the manufacturer’s instructions. The quality of the RNA was assessed using an Agilent 2100 Bioanalyzer (Agilent Technologies, USA). The Illumina® (NEB, USA) sequencing libraries were generated using a NEBNext® UltraTM RNA Library Prep Kit following the manufacturer’s instructions. The constructed libraries were loaded and sequenced on a Hiseq 4000 platform (Illumina, USA). RNA isolation and sequencing were conducted by the Majorbio Institute, Shanghai, China. Finally, paired-end reads of length approximately 150 bp were generated.
Assembly of Transcriptome and Gene Function Annotation
Raw reads generated in FASTQ format were processed in several steps, including removing adapter sequences, poly-N, and low-quality reads to obtain high-quality clean reads. TopHat2 was used to align the data to the human reference genome (hg38). The quality of the sequencing data was evaluated by FastQC. Gene mapping of the reads was conducted by using FeatureCounts v1.5.0-p3. The transcripts per million values were used to estimate gene expression levels.
Differential Gene Expression and Functional Enrichment Analyses
The expression pattern of each sample was analyzed using a principal component analysis (PCA). The differential gene expression (DGE) analysis was performed using the R DESeq2 package (version 3.2.3). The p-value and fold change (FC) of the comparison between samples and controls were calculated. The differentially expressed genes (DEGs) were filtrated based on a FC of <0.5 or >2 and a p-value of <0.05. Volcano plots of the DEGs were generated using the R ggplot2 package (version 3.3.3). The Kyoto Encyclopedia of Genes and Genomes (KEGG) and Gene Ontology (GO) databases (https://david.ncifcrf.gov/) were utilized to conduct biological function and pathway enrichment analysis of the DEGs. Inter-cluster similarities and intra-cluster redundancies were evaluated using the Metascape online database (http://metascape.org).
Western Blot
Protein lysates extracted from A375 cells were separated through sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to polyvinylidene difluoride (PVDF) membranes (88585, Thermo Fisher Scientific, MA, USA). The PVDF membranes were blocked using 5% bovine serum albumin, and then incubated with the following primary antibodies at 4°C overnight: Bcl-XL (dilution 1:2000, 10783-1-AP, Proteintech, Wuhan, China), CHOP (dilution 1:1000, 15204-1-AP, Proteintech, Wuhan, China), GRP78 (dilution 1:2000, 1587-1-AP, Proteintech, Wuhan, China), anti-β-actin (dilution 1:2000, 20536-1-AP, Proteintech, Wuhan, China), cleaved Caspase-3 (dilution 1:1000, AF7022, Affinity, Changzhou, China), p-IRE1α (dilution 1:2000, AF7150, Affinity, Changzhou, China), IRE1α (dilution 1:2000, DF7709, Affinity, Changzhou, China), p-PERK (dilution 1:2000, DF7576, Affinity, Changzhou, China), PERK (dilution 1:2000, AF5304, Affinity, Changzhou, China), p-eIF2α (dilution 1:2000, AF3087, Affinity, Changzhou, China), and eIF2α (dilution 1:2000, AF6087, Affinity, Changzhou, China), and then incubated with HRP-conjugated secondary antibodies (dilution 1:5000, ZB-5301, ZSBIO, Beijing, China) for 1 h and detected using the ECL System (NEL104001EA, PerkinElmer, Shanghai, China).
Statistical Analysis
All data are presented as the mean ± standard deviation (S.D.). SPSS (version 26.0; NY, USA) was used for the data analysis. Images were drawn using GraphPad Prism 8.0 (San Diego, CA, USA). One-way analysis of variance (ANOVA) and non-parametric tests were performed to analyze the differences between groups. p < 0.05 was considered statistically significant.
Results
NaHS Suppresses Cell Proliferation and Induces Apoptosis of A375 Melanoma Cells
A375 cells were treated with different concentrations of NaHS to explore the effects of NaHS. Treatment with NaHS (0, 20, and 50 mM) resulted in morphological changes in the A375 cells, including cell roundness and detachment from the cell culture plate (Figure 1A). CCK8 assay was performed to determine the cell proliferation rate. NaHS treatment (0, 0.5, 5, 10, 20, and 50 mM) inhibited the proliferation of A375 cells in a dose-dependent manner (Figure 1B). Hochest 33258 is a blue, fluorescent dye that penetrates cell membranes and is used to detect apoptosis. Hoechst 33258 staining showed that the nuclei of A375 cells subjected to NaHS (20 mM) exposure for 5 h were fragmented and densely stained compared with the control (0 mM; Figure 1C). Moreover, NaHS (20 mM) induced apoptosis in A375 cells, as confirmed by flow cytometry (Figure 1D). Caspase-3 and Bcl-XL are the key proteins involved in apoptosis. The decreased Bcl-XL and up-regulated cleaved Caspase-3 expression in 20 mM NaHS-treated A375 cells were further confirmed by Western blotting (Figure 1E) (Supplementary Material 1). Therefore, NaHS suppressed cell proliferation and induced apoptosis by providing exogenous H2S in human A375 melanoma cells.
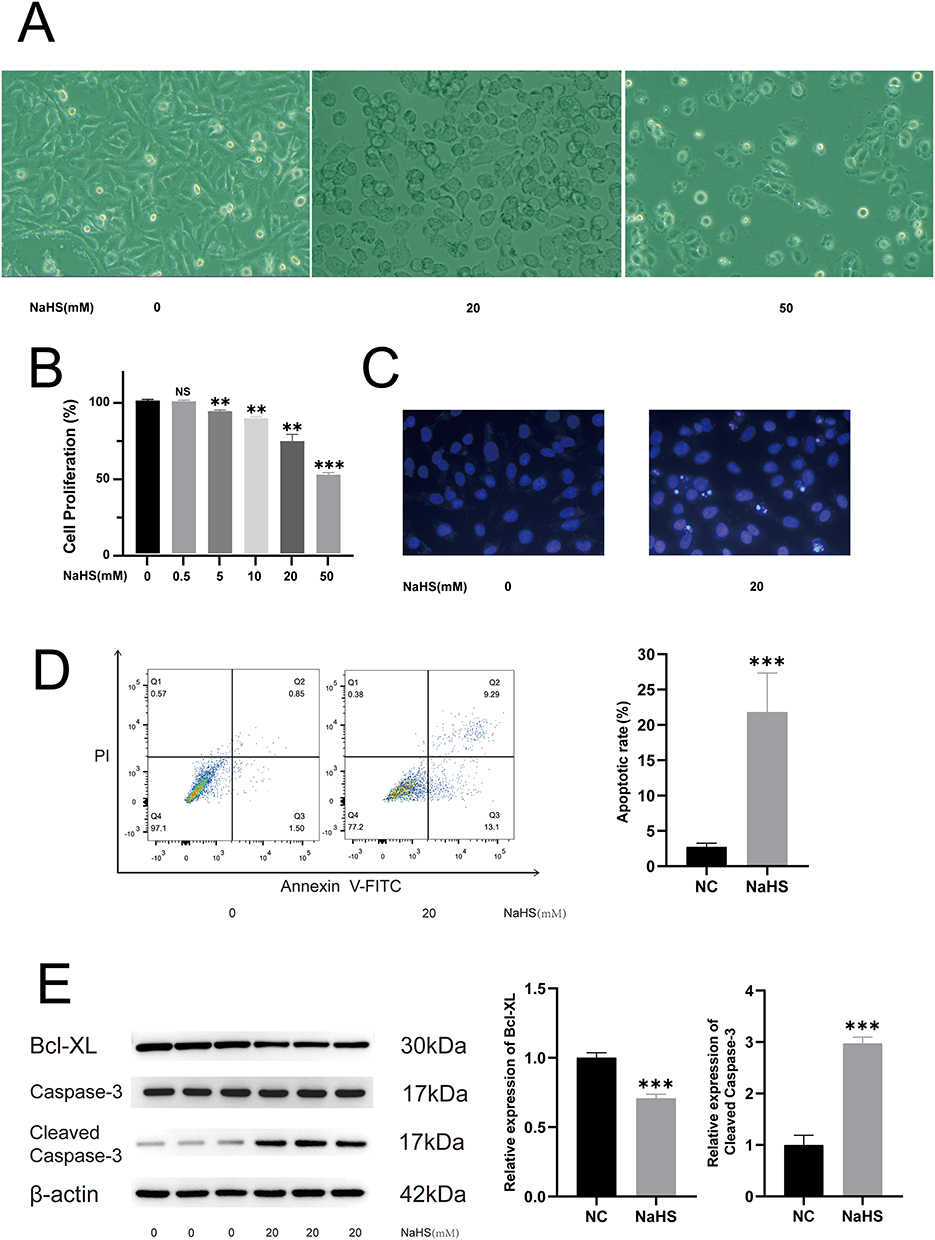 |
Figure 1 NaHS suppressed cell proliferation and induced apoptosis of A375 cells. (A) Morphological changes of A375 cells caused by treatment at different concentrations of NaHS (0, 20, and 50 mM). (B) CCK-8 assay at different concentrations of NaHS-treated A375 cells. (C) Hoechst 33258 staining of A375 cells subjected to NaHS (20 mM) exposure and control cells (0 mM). (D) Cytometric analysis of NaHS (20 mM) treated A375 cells and control cells (0 mM). (E) Western blotting analysis of cleaved Caspase-3 and Bcl-XL in NaHS (20 mM) treated A375 cells and control cells (0 mM). **P < 0.01, ***P < 0.001 compared with negative control (NC). |
Transcriptional Profiles of NaHS-Treated A375 Melanoma Cells Showed Increased Endoplasmic Reticulum Stress and Overactivation of the Unfolded Protein Response
To understand transcriptome changes after NaHS treatment, high-throughput sequencing was performed on 20 mM NaHS-treated A375 cells. The quality of the sequencing data was verified, and the mapping rates were 96.25% to 96.51%. The data were then dimensionally reduced, and the noise was removed by PCA to demonstrate the relationship between the samples, resulting in distinct clustering of both the treated and untreated samples (Figure 2A). Correlation analysis between samples was based on the gene expression matrix. Correlation analysis showed that the treated and untreated samples were positively correlated with biological replicates (Figure 2B).
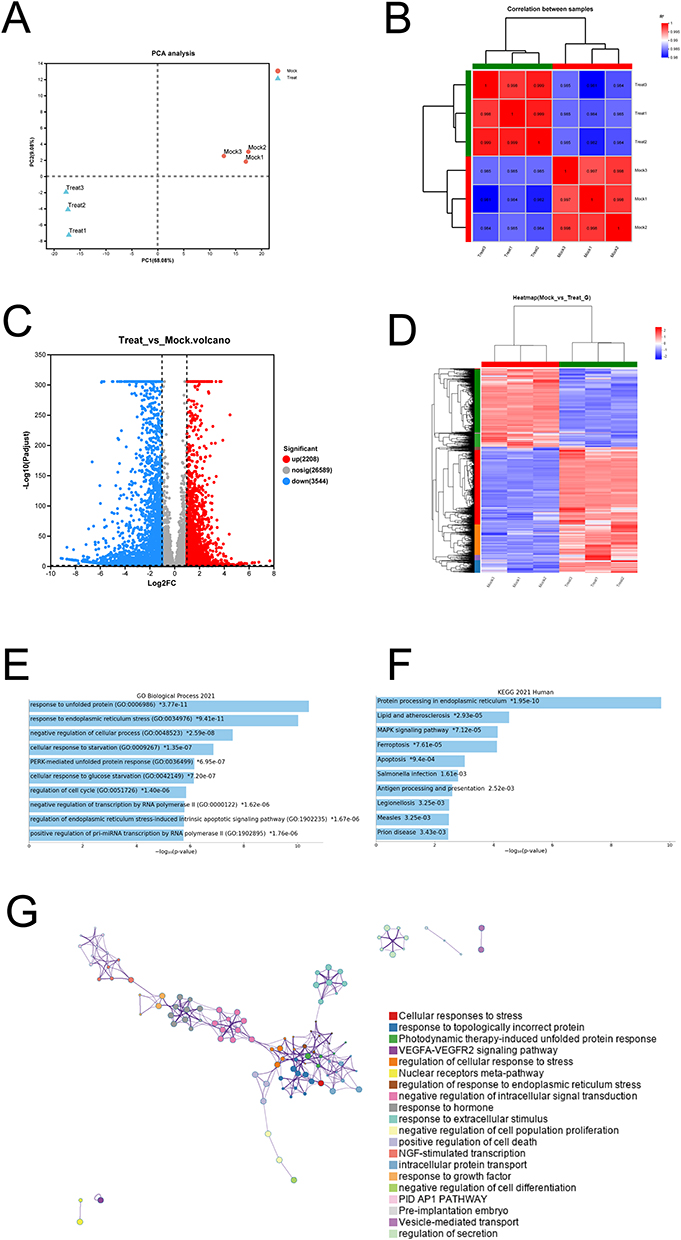 |
Figure 2 Transcriptional profiles showed increased endoplasmic reticulum stress and unfolded protein response in NaHS-treated A375 cells. (A) Principal-component analysis (PCA) of the samples. (B) Correlation analysis of the samples. (C) Volcano plot of the differential expression of genes between NaHS (20 mM) treated A375 cells and control cells (0 mM). (D) Heat map of the differentially expressed genes (DEGs). (E) GO biological process enrichment analysis of the up-regulated DEGs. (F) KEGG pathway analysis of the up-regulated DEGs. (G) Metascape network showed inter-cluster enriched GO terms for the up-regulated DEGs. |
The R DESeq2 package (version 3.2.3) was used to perform DGE analysis. A total of 5752 DEGs were found between the treated and untreated cells. As shown on the volcano plot, 2208 upregulated and 3544 downregulated genes were found in the NaHS-treated cells compared to the untreated cells (Figure 2C). Unsupervised hierarchical clustering based on the similarities in the expression of these DEGs distinguished the treated cells from the untreated cells on the heat map (Figure 2D). Furthermore, biological process enrichment analysis was performed using the GO and KEGG databases to understand the functions of these DEGs. The DEGs were related to the regulation of response to unfolded proteins, response to the endoplasmic reticulum (ER) stress, negative regulation of cellular processes, and regulation of the cell cycle, according to the GO database analysis (Figure 2E). The KEGG pathway analysis revealed that protein processing in the ER, apoptosis, ferroptosis, MAPK signaling pathway, and other cell stress-associated pathways were enriched in the NaHS-treated A375 cells (Figure 2F). The Metascape database was also used to perform a network enrichment analysis. The findings revealed that the unfolded protein response (UPR) and ER stress-associated processes were prominent in the transcriptome and induced a series of related biological processes with a radial distribution (Figure 2G). Our findings indicate that the unfolded protein response, endoplasmic reticulum stress, and apoptosis-associated gene expression signatures were prominent in the NaHS-treated A375 melanoma cells.
Increase in Unfolded Protein Response and Endoplasmic Reticulum Stress in the NaHS-Treated A375 Cells
Treatment with NaHS led to an increase in the number of unfolded proteins inside the ER lumen as well as an increase in ER stress. Considering that UPR and ER stress was prominently observed during the transcriptional phase of 20 mM NaHS-treated A375 cells, Western blotting was conducted to verify these changes. CHOP and GRP78 were considered markers for ER stress. As shown in Figure 3A, the increased expression levels of CHOP and GRP78 in 20 mM NaHS-treated A375 cells indicates a significant increase in ER stress. When the expression levels of unfolded proteins reach the threshold values, the cells initiate UPR to restore homeostasis. If the reaction is mild, cellular homeostasis is restored, and the cells remain alive. When the expression level of unfolded proteins is excessively high and ER stress triggers the apoptotic process, the cell dies. IRE1α, PERK, and eIF2α are critical molecules in the UPR pathway. The UPS was increased in the 20 mM NaHS-treated A375 cells based on the results of Western blotting, and the ratio of p‐PERK to PERK protein, p‐IRE1α to IRE1α protein, and p‐eIF2α to eIF2α protein significantly increased (Figure 3B) (Supplementary Material 1). As a result of high ER stress and overreaction of UPR, the application of 20 mM NaHS induced cancer cell apoptosis (Figure 1D). These findings indicate that NaHS induced the apoptosis of melanoma cells by increasing the ER stress levels and UPR.
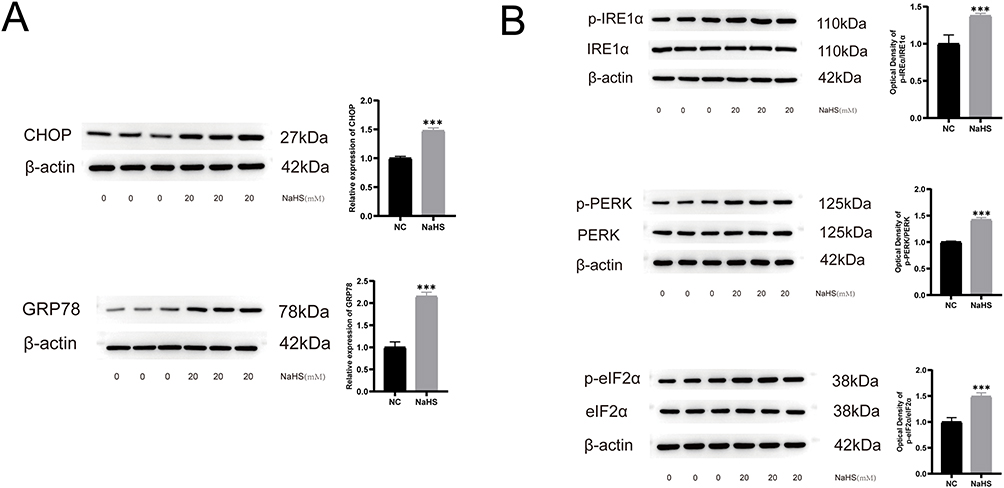 |
Figure 3 The increased protein expression levels of unfolded proteins response and endoplasmic reticulum stress in NaHS-treated A375 cells. (A) Western blot of CHOP and GRP78 in NaHS (20 mM) treated A375 cells and control cells (0 mM). (B) Western blot of p‐IRE1α, IRE1α protein, p‐PERK, PERK protein, p‐eIF2α, and eIF2α in NaHS (20 mM) treated A375 cells and control cells (0 mM). ***P < 0.001 compared with negative control (NC). |
Discussion
The present study showed that exposure of A375 human melanoma cells to NaHS, an exogenous H2S donor, suppressed cell proliferation and increased cell apoptosis. The ER stress and UPS showed remarkable transcriptional changes, identified using high-throughput sequencing. These changes were verified at the protein level. Thus, this study’s findings broadened the understanding of ER stress and UPS mechanisms involved in the antitumor effect of exogenous H2S.
Melanoma is highly malignant, and patients with advanced tumors have a poor prognosis, with a mortality rate of over 90%.26 Systemic chemotherapy has been applied for the treatment of advanced melanoma for a long time.27 Dacarbazine is commonly used as an FDA-approved single agent to treat metastatic melanoma, and clinical trials of dacarbazine in combination with other drugs are currently underway for the treatment of malignant melanoma.28 Carboplatin plus paclitaxel was another widely used first- and second-line combination chemotherapy treatment.29 But unfortunately, in patients with advanced disease, conventional chemotherapy treatments are ineffective in preventing disease recurrence, and several patients with various cancer types develop resistance within a few months of treatment.30 In tumors resistant to chemotherapy, cancer cells resist apoptosis, and cells survive and overproliferate.16 Cancer cell’s evasion of and resistance to apoptosis often emerges in conventional chemotherapeutic treatment, resulting in poor clinical outcomes.31 Therefore, the induction of cancer cell apoptosis is expected to be a promising and effective strategy for the treatment of melanoma.
Recently, several studies have confirmed the involvement of H2S in cancer development, although these studies have reported conflicting results on the pro- and anticancer effects of H2S. Wu et al treated thyroid cancer cells with NaHS for 24 h; the cells showed reduced ROS levels, the PI3K/Akt/mTOR signaling pathway was activated, and cancer cell proliferation and migration were promoted.22 However, some studies revealed that H2S shows an inhibition effect on the proliferation and viability of cancer cells.32 Lu et al reported that cell cycle arrest was triggered by the downregulation of cell cycle protein D1 in the HepG2 cells within 24 h of treatment with the H2S donor, GYY4137.33 Interestingly, studies proved that H2S has a selective toxic effect on cancer cells while not apparently damaging to normal cells.32,34 Under the same conditions, CA9-22 cancer cells treated with H2S for 48–72 h showed a significant increase in apoptosis, while no pro-apoptotic effect was observed on the normal human gingival keratin-forming cells.35 Therefore, exposure to a high concentration of exogenous H2S selectively suppresses the proliferation of cancer cells and induces apoptosis.36 Here, we demonstrated the ability of NaHS to inhibit melanoma cell proliferation and induce apoptosis in vitro by exposing A375 cells to the H2S donor, NaHS, at higher concentrations for 24 h. A previous study reported that H2S inhibits NF-κB activity in human melanoma cells and decreases the expression level of Bcl-2 to induce spontaneous apoptosis.25 This was confirmed in our study, where Bcl-XL expression was downregulated, while cleaved-caspase-3 was upregulated in NaHS-treated A375 cells. Exogenous H2S exerts anti-cancer effects by enhancing autophagy and promoting apoptosis in human melanoma cells by regulating the PI3K/Akt/mTOR signaling pathway.37 Our study demonstrated the promotion of apoptosis in NaHS-treated cells. Moreover, the unfolded proteins were significantly increased in NaHS-treated cells, thus triggering ER stress and eventually apoptosis, which expanded the antitumor mechanism of high concentration H2S.
Cells broadly alter their transcriptional profile accordingly when facing changes in their environment or specific stimuli to achieve homeostasis.38,39 This is especially true for stress-responsive pathways, where transcription must be consistently altered to adapt to the environment. Our results, using transcriptome sequencing, indicate that treatment of melanoma with high concentrations of H2S significantly upregulated the UPR-related and ER stress-related pathway signals. UPR is an element of the integrative stress response activated under ER stress conditions and aims to achieve cellular and tissue homeostasis.40,41 The ER is an essential site of protein biosynthesis, post-translational modifications, and newly synthesized protein folding and assembly.42 As a membrane compartment, the ER is highly sensitive to changes affecting its structure, integrity, and function, which can disrupt the protein folding process.43 Treatment with high concentrations of H2S may lead to impaired protein synthesis in the ER and increased accumulation of unfolded or misfolded proteins. When the pathological proteins are overloaded, UPR is initiated.44 Three receptor proteins control the classical UPR: PERK, IRE1α, and ATF6.45 Our results showed a significant upregulation of PERK and IRE1α. The UPR, triggered by the activation of stress sensors, aimed to reduce the unfolded protein load, an adaptive response that helps maintain cell viability and function.46 When the UPR fails to adapt to ER stress, the cells are unable to achieve homeostasis, leading to apoptosis.47,48 Our study found that in the presence of high concentrations of exogenous H2S, the UPR of melanoma cells was significantly enhanced, ER stress increased, cellular homeostasis was not achieved, apoptotic processes were activated, and apoptotic pathway signals were significantly upregulated.
Conclusion
This study shows that the exogenous H2S donor, NaHS, enhances the apoptotic process in melanoma cells. This study was the first to report that the protein synthesis process in the ER was affected in treated cells, with a significant increase in unfolded proteins, triggering ER stress and ultimately leading to apoptosis. The present study findings help expand the understanding of the mechanism of the antitumor effect of H2S treatment at high concentrations.
Abbreviations
Bcl-2, B-cell lymphoma 2; Bcl-XL, B-cell lymphoma 2 -extra large; CCK-8, cell counting kit-8; CHOP, C/EBP homologous protein; DMEM, Dulbecco’s modified eagle medium; FITC, fluorescein isothiocyanate; GRP78, glucose-regulating protein 78; mTOR, mammalian target of rapamycin; NaHS, sodium hydrosulfide; p-eIF2α, phosphorylated eukaryotic translation initiation factor 2α; PI, propidium iodide; p‐IRE1α, phosphorylated inositol-requiring enzyme 1α; p-PERK, phosphorylated protein kinase R-like ER kinase; UPR, unfolded protein response.
Data Sharing Statement
Data supporting this study’s findings are available from the corresponding author upon reasonable request.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis, and interpretation, or all these areas; took part in drafting, revising, or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
Research reported in this publication was supported by the Traditional Chinese Medicine Science and Technology Project of Shandong Province (No. 2021Q093), the Doctoral Startup Fund of Affiliated Hospital of Weifang Medical University (No. 2021BKQ02), and Shandong Province Natural Science Foundation (No. ZR2020QH138).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Liu D, Lin JR, Robitschek EJ, et al. Evolution of delayed resistance to immunotherapy in a melanoma responder. Nat Med. 2021;27(6):985–992. doi:10.1038/s41591-021-01331-8
2. Schadendorf D, van Akkooi ACJ, Berking C, et al. Melanoma. Lancet. 2018;392(10151):971–984. doi:10.1016/S0140-6736(18)31559-9
3. Millington GW. Mutations of the BRAF gene in human cancer, by Davies et al. (Nature 2002;417:949–54). Clin Exp Dermatol. 2013;38(2):222–223. doi:10.1111/ced.12015
4. Long GV, Weber JS, Infante JR, et al. Overall survival and durable responses in patients with BRAF V600-mutant metastatic melanoma receiving dabrafenib combined with trametinib. J Clin Oncol. 2016;34(8):871–878. doi:10.1200/JCO.2015.62.9345
5. Hughes T, Klairmont M, Sharfman WH, Kaufman HL. Interleukin-2, Ipilimumab, and Anti-PD-1: clinical management and the evolving role of immunotherapy for the treatment of patients with metastatic melanoma. Cancer Biol Ther. 2021;22(10–12):513–526. doi:10.1080/15384047.2015.1095401
6. Candido S, Salemi R, Piccinin S, et al. The PIK3CA H1047R mutation confers resistance to BRAF and MEK inhibitors in A375 melanoma cells through the cross-activation of MAPK and PI3K-Akt pathways. Pharmaceutics. 2022;14(3):590. doi:10.3390/pharmaceutics14030590
7. Patel M, Eckburg A, Gantiwala S, et al. Resistance to molecularly targeted therapies in melanoma. Cancers. 2021;13(5):1115. doi:10.3390/cancers13051115
8. Stachyra-Strawa P, Ciesielka M, Janiszewski M, Grzybowska-Szatkowska L. The role of immunotherapy and molecular‑targeted therapy in the treatment of melanoma (Review). Oncol Rep. 2021;46(2):158.
9. Rodriguez-Pascual J, Ayuso-Sacido A, Belda-Iniesta C. Drug resistance in cancer immunotherapy: new strategies to improve checkpoint inhibitor therapies. Cancer Drug Resist. 2019;2(4):980–993. doi:10.20517/cdr.2019.61
10. Chen G, Davies MA. Targeted therapy resistance mechanisms and therapeutic implications in melanoma. Hematol Oncol Clin North Am. 2014;28(3):523–536. doi:10.1016/j.hoc.2014.03.001
11. Chawla-Sarkar M, Bae SI, Reu FJ, Jacobs BS, Lindner DJ, Borden EC. Downregulation of Bcl-2, FLIP or IAPs (XIAP and survivin) by siRNAs sensitizes resistant melanoma cells to Apo2L/TRAIL-induced apoptosis. Cell Death Differ. 2004;11(8):915–923. doi:10.1038/sj.cdd.4401416
12. Zhong J, Yan W, Wang C, et al. BRAF inhibitor resistance in melanoma: mechanisms and alternative therapeutic strategies. Curr Treat Options Oncol. 2022;23(11):1503–1521. doi:10.1007/s11864-022-01006-7
13. Neophytou CM, Trougakos IP, Erin N, Papageorgis P. Apoptosis deregulation and the development of cancer multi-drug resistance. Cancers. 2021;13(17):4363. doi:10.3390/cancers13174363
14. Tsuruo T, Naito M, Tomida A, et al. Molecular targeting therapy of cancer: drug resistance, apoptosis and survival signal. Cancer Sci. 2003;94(1):15–21. doi:10.1111/j.1349-7006.2003.tb01345.x
15. Carneiro BA, El-Deiry WS. Targeting apoptosis in cancer therapy. Nat Rev Clin Oncol. 2020;17(7):395–417. doi:10.1038/s41571-020-0341-y
16. Mohammad RM, Muqbil I, Lowe L, et al. Broad targeting of resistance to apoptosis in cancer. Semin Cancer Biol. 2015;35:S78–S103. doi:10.1016/j.semcancer.2015.03.001
17. Vandiver M, Snyder SH. Hydrogen sulfide: a gasotransmitter of clinical relevance. J Mol Med. 2012;90(3):255–263. doi:10.1007/s00109-012-0873-4
18. Hartle MD, Pluth MD. A practical guide to working with H(2)S at the interface of chemistry and biology. Chem Soc Rev. 2016;45(22):6108–6117. doi:10.1039/C6CS00212A
19. Sun Y, Huang Y, Zhang R, et al. Hydrogen sulfide upregulates KATP channel expression in vascular smooth muscle cells of spontaneously hypertensive rats. J Mol Med. 2015;93(4):439–455. doi:10.1007/s00109-014-1227-1
20. Kimura Y, Kimura H. Hydrogen sulfide protects neurons from oxidative stress. FASEB J. 2004;18(10):1165–1167. doi:10.1096/fj.04-1815fje
21. Ma Z, Bi Q, Wang Y. Hydrogen sulfide accelerates cell cycle progression in oral squamous cell carcinoma cell lines. Oral Dis. 2015;21(2):156–162. doi:10.1111/odi.12223
22. Wu D, Li J, Zhang Q, et al. Exogenous Hydrogen Sulfide Regulates the Growth of Human Thyroid Carcinoma Cells. Oxid Med Cell Longev. 2019;2019:6927298. doi:10.1155/2019/6927298
23. Wu D, Si W, Wang M, Lv S, Ji A, Li Y. Hydrogen sulfide in cancer: friend or foe? Nitric Oxide. 2015;50:38–45. doi:10.1016/j.niox.2015.08.004
24. Panza E, De Cicco P, Armogida C, et al. Role of the cystathionine gamma lyase/hydrogen sulfide pathway in human melanoma progression. Pigment Cell Melanoma Res. 2015;28(1):61–72. doi:10.1111/pcmr.12312
25. De Cicco P, Panza E, Ercolano G, et al. ATB-346, a novel hydrogen sulfide-releasing anti-inflammatory drug, induces apoptosis of human melanoma cells and inhibits melanoma development in vivo. Pharmacol Res. 2016;114:67–73. doi:10.1016/j.phrs.2016.10.019
26. Shain AH, Bastian BC. From melanocytes to melanomas. Nat Rev Cancer. 2016;16(6):345–358. doi:10.1038/nrc.2016.37
27. Falzone L, Bordonaro R, Libra M, et al. SnapShot: cancer chemotherapy. Cell. 2023;186(8):1816–1816.e1. doi:10.1016/j.cell.2023.02.038
28. Karapetyan L, Kirkwood JM. State of melanoma: an historic overview of a field in transition. Hematol Oncol Clin North Am. 2021;35(1):1–27. doi:10.1016/j.hoc.2020.09.003
29. Gupta A, Gomes F, Lorigan P. The role for chemotherapy in the modern management of melanoma. Melanoma Manag. 2017;4(2):125–136. doi:10.2217/mmt-2017-0003
30. Soengas MS, Lowe SW. Apoptosis and melanoma chemoresistance. Oncogene. 2003;22(20):3138–3151. doi:10.1038/sj.onc.1206454
31. Pistritto G, Trisciuoglio D, Ceci C, Garufi A, D’Orazi G. Apoptosis as anticancer mechanism: function and dysfunction of its modulators and targeted therapeutic strategies. Aging. 2016;8(4):603–619. doi:10.18632/aging.100934
32. Lee ZW, Teo XY, Tay EY, et al. Utilizing hydrogen sulfide as a novel anti-cancer agent by targeting cancer glycolysis and pH imbalance. Br J Pharmacol. 2014;171(18):4322–4336. doi:10.1111/bph.12773
33. Lu S, Gao Y, Huang X, Wang X. GYY4137, a hydrogen sulfide (H(2)S) donor, shows potent anti-hepatocellular carcinoma activity through blocking the STAT3 pathway. Int J Oncol. 2014;44(4):1259–1267. doi:10.3892/ijo.2014.2305
34. Yang CT, Zhao Y, Xian M, et al. A novel controllable hydrogen sulfide-releasing molecule protects human skin keratinocytes against methylglyoxal-induced injury and dysfunction. Cell Physiol Biochem. 2014;34(4):1304–1317. doi:10.1159/000366339
35. Murata T, Sato T, Kamoda T, Moriyama H, Kumazawa Y, Hanada N. Differential susceptibility to hydrogen sulfide-induced apoptosis between PHLDA1-overexpressing oral cancer cell lines and oral keratinocytes: role of PHLDA1 as an apoptosis suppressor. Exp Cell Res. 2014;320(2):247–257. doi:10.1016/j.yexcr.2013.10.023
36. Cao X, Ding L, Xie ZZ, et al. A review of hydrogen sulfide synthesis, metabolism, and measurement: is modulation of hydrogen sulfide a novel therapeutic for cancer? Antioxid Redox Signal. 2019;31(1):1–38. doi:10.1089/ars.2017.7058
37. Xiao Q, Ying J, Qiao Z, et al. Exogenous hydrogen sulfide inhibits human melanoma cell development via suppression of the PI3K/AKT/ mTOR pathway. J Dermatol Sci. 2020;98(1):26–34. doi:10.1016/j.jdermsci.2020.02.004
38. Lempradl A, Pospisilik JA, Penninger JM. Exploring the emerging complexity in transcriptional regulation of energy homeostasis. Nat Rev Genet. 2015;16(11):665–681. doi:10.1038/nrg3941
39. Shi W, Ma W, Xiong L, Zhang M, Tang C. Adaptation with transcriptional regulation. Sci Rep. 2017;7(1):42648. doi:10.1038/srep42648
40. Zhu G, Lee AS. Role of the unfolded protein response, GRP78 and GRP94 in organ homeostasis. J Cell Physiol. 2015;230(7):1413–1420. doi:10.1002/jcp.24923
41. Doyle KM, Kennedy D, Gorman AM, Gupta S, Healy SJ, Samali A. Unfolded proteins and endoplasmic reticulum stress in neurodegenerative disorders. J Cell Mol Med. 2011;15(10):2025–2039. doi:10.1111/j.1582-4934.2011.01374.x
42. Ellgaard L, McCaul N, Chatsisvili A, Braakman I. Co- and Post-Translational Protein Folding in the ER. Traffic. 2016;17(6):615–638. doi:10.1111/tra.12392
43. Bravo R, Parra V, Gatica D, et al. Endoplasmic reticulum and the unfolded protein response: dynamics and metabolic integration. Int Rev Cell Mol Biol. 2013;301:215–290. doi:10.1016/B978-0-12-407704-1.00005-1
44. van der Voorn JP, van Kollenburg B, Bertrand G, et al. The unfolded protein response in vanishing white matter disease. J Neuropathol Exp Neurol. 2005;64(9):770–775. doi:10.1097/01.jnen.0000178446.41595.3a
45. Adams CJ, Kopp MC, Larburu N, Nowak PR, Ali MMU. Structure and molecular mechanism of ER stress signaling by the unfolded protein response signal activator IRE1. Front Mol Biosci. 2019;6:11.
46. Hetz C. The unfolded protein response: controlling cell fate decisions under ER stress and beyond. Nat Rev Mol Cell Biol. 2012;13(2):89–102. doi:10.1038/nrm3270
47. Hetz C, Zhang K, Kaufman RJ. Mechanisms, regulation and functions of the unfolded protein response. Nat Rev Mol Cell Biol. 2020;21(8):421–438. doi:10.1038/s41580-020-0250-z
48. Soltanmohammadi E, Farmaki E, Zhang Y, et al. Coordination in the unfolded protein response during aging in outbred deer mice. Exp Gerontol. 2021;144:111191. doi:10.1016/j.exger.2020.111191
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Zinc Oxide Particles Can Cause Ovarian Toxicity by Oxidative Stress in Female Mice Model

Xu Y, Zhao Y, Liu S, Lv S, Chen L, Wang W, Feng Y, Fu F, Xu H
International Journal of Nanomedicine 2022, 17:4947-4960
Published Date: 19 October 2022
RET Inhibitor SPP86 Triggers Apoptosis and Activates the DNA Damage Response Through the Suppression of Autophagy and the PI3K/AKT Signaling Pathway in Melanoma Cells
Zhang Y, Liu H, Wang K, Zheng J, Luan H, Xin M
Drug Design, Development and Therapy 2025, 19:67-82
Published Date: 7 January 2025
Saikosaponin A Mediates the Anti-Acute Myeloid Leukemia Effect via the P-JNK Signaling Pathway Induced by Endoplasmic Reticulum Stress
Sun XH, Chai YH, Bai XT, Li HX, Yang PP, Xi YM
Drug Design, Development and Therapy 2025, 19:1983-2001
Published Date: 17 March 2025