Back to Journals » Drug Design, Development and Therapy » Volume 10
Establishment of a novel therapeutic vector targeting the trigeminal ganglion in rats
Authors Xu K, Pan S, Song J, Liu X, An N, Zheng X
Received 20 September 2015
Accepted for publication 15 October 2015
Published 4 February 2016 Volume 2016:10 Pages 585—592
DOI https://doi.org/10.2147/DDDT.S96730
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Prof. Dr. Wei Duan
Kun Xu,1,2 Shi-Yin Pan,1,2 Jin-Xin Song,1 Xian-Ning Liu,1,2 Na An,1,2 Xuan Zheng1,2
1Department of Ophthalmology, The No 1 Hospital of Xi’an, 2Shaanxi Provincial Key Laboratory of Ophthalmology, Ophthalmological Institute of Shaanxi Province, Xi’an, Shaanxi Province, People’s Republic of China
Background: In the pathogenesis of herpes simplex keratitis, herpes simplex virus type 1 (HSV-1) infection begins in corneal epithelium cells and then progresses through the sensory nerve endings and finally travels up forward to the trigeminal ganglion (TG), where it remains as latent virus. The available anti-HSV therapies do not completely suppress the recurrence of active HSV-1 infection. The aim of this study was to establish a novel replication-defective (rd) HSV-1 (rdHSV) vector (rdHSV-interferon gamma [IFNγ]) that could effectively target the TG.
Methods: Recombinant HSV-1 virus was inserted into a shuttle plasmid carrying IFNγ to establish the rdHSV-IFNγ vector. Safety was evaluated in vitro by 50% cellular cytotoxicity in transfected SH-SY5Y neuroblastoma cells and in vivo by Kaplan–Meier survival estimate and infection rate. Wistar rats were immunized with rdHSV-IFNγ to evaluate the TG targeting efficiency. Real-time polymerase chain reaction and Western blot assays were used to evaluate IFNγ mRNA and protein expression and rdHSV-IFNγ localization.
Results: The rdHSV-IFNγ vector was successfully constructed and showed high in vitro safety and overall survival and a corneal infection rate similar to that of control rats immunized with saline (control group; P>0.05). Real-time polymerase chain reaction and immunohistochemistry assays confirmed IFNγ expression and effective TG targeting on days 14 and 21, which increased with postimmunization time. Moreover, IFNγ was expressed sufficiently in the TG tissues.
Conclusion: The rdHSV-IFNγ can act as an effective gene transporting vector that carries the therapeutic genes to the TG and triggers its expression.
Keywords: replication-defective HSV-1, interferon gamma, trigeminal ganglion, therapeutic gene
Introduction
Herpes simplex virus (HSV) infection in the corneas is the main kind of keratitis, which always develops to a chronic or an acute corneal inflammation.1,2 Among several HSVs, the herpes simplex virus type 1 (HSV-1) is the most common cause of the herpes simplex keratitis (HSK). The HSK is a prevalent reason for corneal blindness in the modern clinical ophthalmology.3 Regarding the pathology of the HSK, the HSV-1 first infects the corneal epithelium cells, then invades the sensory nerve endings, and finally travels to the trigeminal ganglion (TG).4 When the HSV-1 passes the acute stage, it becomes latent in the TG.4 It is well known that HSK is one of the most frequently recurrent corneal diseases based on the reactivation of the latent HSV-1 in the trigeminal sensory neurons. The recurrence rate of HSK within 2 years is ~23%–33%. Approximately 20%–25% of the recurrent HSK could develop to the T-cell-mediated HSK, which is the prevalent reason for the unilateral blindness.5
Although some anti-HSV or immunoregulatory treatments are available, they do not eliminate the latent virus and therefore cannot completely suppress the recurrence of the HSV-1 infection.6 Recently, DNA vaccines or vectors have been shown to trigger a cell-mediated immune response in the HSK rat models. However, HSV-1 infection was blocked only in the acute stage of HSK and there was no effect against the latent HSV-1 infection of the TG. Hu et al7 developed a DNA vaccine that triggered a better immune protection against the HSV-1 infection of the ocular surface but did not inhibit the recurrence of corneal HSV-1 infection. Over the past 20 years, investigations to develop therapies for HSV-1 infection have been focused on developing effective treatment on the removal of latent infection of the TG and on the elimination of the latent virus in the TG. In this study, we attempted to establish an effective therapeutic system targeting the TG and cleaning up the latent virus in the TG. We took advantage of the neurotrophic properties of HSV-1 to make a recombinant, replication-defective (rd) HSV-1 vector that could be efficiently transported along neurons and could carry therapeutic IFN to the TG without any toxicity.
Materials and methods
Animals, cell lines, and viruses
Male specific pathogen-free Wistar rats aged 5–6 weeks were purchased from the Experimental Animal Center of Xi’an Jiaotong University (Xi’an, People’s Republic of China). The rats were raised under pathogen-free conditions in containers supplied with filtered air. The Ethics Committee of the No 1 Hospital of Xi’an, Xi’an, People’s Republic of China, approved all the animal experiments.
The human neuroblastoma cell line SH-SY5Y and Vero cells were obtained from the Chinese Center for Disease Control and Prevention (Beijing, People’s Republic of China), grown in a 5% CO2 atmosphere at 37°C in Dulbecco’s Modified Eagle’s Medium (DMEM) supplemented with 10% fetal bovine serum, 100 μg/mL streptomycin sulfate, and 100 units/mL penicillin G sodium.
The HSV-1 strain SM44 was obtained from Professor Xiuping Zhu at the Ophthalmological Institute of Shaanxi Province (Xi’an, People’s Republic of China) and was propagated, activated, and cultured in Vero cells. No ethics statement was required from the institutional review board for the use of these cell lines.
Recombination of rdHSV-1
To control the replication of the established vector, we removed the immediate-early, ICP27, ICP4, and ICP34.5 genes, which are essential for replication, to form the rd viruses.8 In order to inhibit the cytotoxicity of the rdHSV-1, the pathogenic gene, VP16, was inactivated and mutated.9 To allow prolonged expression of the exogenous genes, we inserted a strong heterologous promoter at the location of the 1.4 kb downstream under the latency-associated promoter 1 TATA box (Figure 1A).
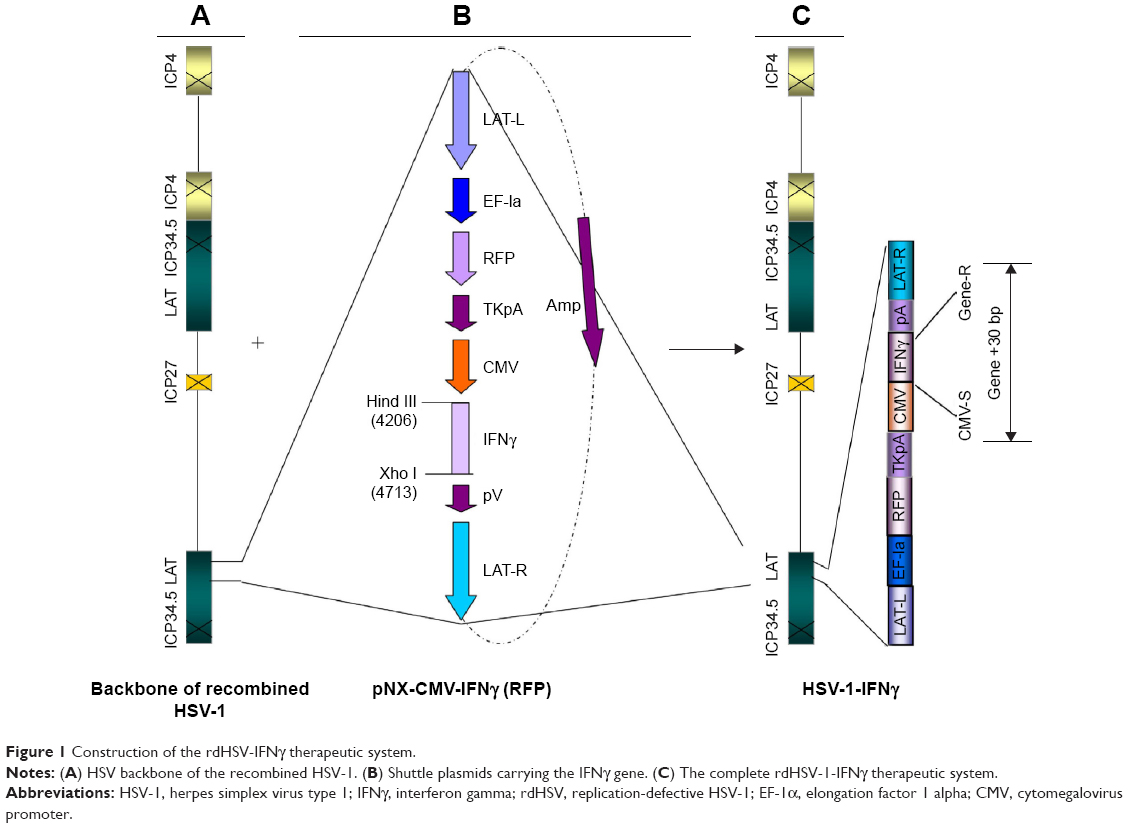 | Figure 1 Construction of the rdHSV-IFNγ therapeutic system. |
IFNγ clone and establishment of rdHSV-1 therapeutic system
Lymphocytes were isolated from the blood of a healthy volunteer and were grown in primary culture in the DMEM solution supplemented with 10% fetal bovine serum. The HSV-1 strain SM44 virus was used to activate the lymphocytes, and the total RNA was extracted from the activated lymphocytes. The total RNA was employed to synthesize the cDNA. IFNγ was amplified from the cDNA by PCR using 5′-ACGAAGCTTATGAAATATACAAGTTATATCTTG-3′ as the forward primer and 5′-ATCCTCGAGTTACTGGGATGCTCTTCGAC-3′ as the reverse primer.
The amplified IFNγ gene was cloned and inserted into a shuttle plasmid (Figure 1B). The rdHSV-1 vector was generated by calcium phosphate cotransfection of complementing cells with the shuttle plasmid and HSV-1 backbone as previously described.9,10 The rdHSV-1 system (rdHSV-IFNγ) was established successfully (Figure 1C), and titers were evaluated by using tenfold serial dilution as previously reported.11 The plaque forming units (PFUs) typically ranged from 3×108 PFU/mL to 2×109 PFU/mL.
XTT cytotoxicity assay
SH-SY5Y cells cultured in 96- or six-well plates were infected with the rdHSV-IFNγ vector (at final concentrations of 25 TCID50, 50 TCID50, 100 TCID50, and 200 TCID50, respectively). After 48 hours, antiviral activity and cytotoxicity were determined by XTT assay. The dimethyl sulfoxide solubilization solution was used as a negative control, and the assay was performed as previously described.12 The results of XTT assay were reported as the percentage inhibition of cytotoxicity calculated as follows: inhibition% = [100- (At/As) ×100]%, where At/As is the ratio of the absorbance of the formazan-containing assay solution to the absorbance of the XTT solubilization solution at 48 hours. The vector concentration resulting in 50% cellular cytotoxicity (CC50) was determined as described by Chiang et al.13
Immunization with rdHSV-IFNγ
Wistar rats were anesthetized, and their corneas were scarified with a syringe needle (1 mL type) for at least ten times. The rats were then randomly assigned to three groups (n=20 per group), including blank rdHSV vector group (blank rdHSV-1), rdHSV-IFNγ group, and control (CN) group (treated with saline), that is, to the topical treatment of the cornea with 10 μL of blank rdHSV (2×106 PFU), rdHSV-IFNγ (2×106 PFU/mL), and a CN group given normal saline solution.
Immunohistochemistry analysis
Trigeminal ganglia were isolated from the rats, fixed in 4% paraformaldehyde, embedded in commercial tissue freezing medium (TIANGEN Biotech Co., Ltd., Beijing, People’s Republic of China), and immediately frozen in isopentane. Cryosections (4 μm) were cut, fixed in 70% ethanol, and incubated with mouse anti-HSV-1 monoclonal antibody (1:1,000; sc-57862; Santa Cruz Biotechnology Inc., Dallas, TX, USA) for at least 1 hour. The sections were also stained with hematoxylin and eosin, and all the procedures were performed according to the manufacturer’s instructions.
IFNγ detection by Western blot assay
Trigeminal ganglia were isolated for the detection of IFNγ expression by Western blot assay. The treatment of trigeminal ganglia and the Western blot processes were performed according to the previous study.14 The primary antibodies were mouse anti-IFNγ monoclonal antibody (1:2,000) and mouse anti-β-actin monoclonal antibody (1:2,000). The secondary antibody was horseradish-peroxidase-conjugated rabbit anti-mouse antibody (1:1,000; all from Santa Cruz Biotechnology Inc.). The Western blot bands were scanned, and the pixel count and intensity of each band were semiquantified. The signals were normalized against β-actin, and the data were expressed as the percentage of the negative control signals.
IFNγ assay by real-time polymerase chain reaction
The expression of IFNγ mRNA in trigeminal ganglia was assayed by real-time polymerase chain reaction (RT-PCR). The forward primer was 5′-ACGAAGCTTATGAAATATACAAGTTATATCTTG-3′, and the reverse primer was 5′-ATCCTCGAGTTACTGGGATGCTCTTCGAC-3′. β-actin was used as the internal control; the forward primer was 5′-GGACTTC GAGCAGGAGATGG-3′, and the reverse primer was 5′-GCACCGTG TTGGCGTAGAGG-3′. The isolation of total RNA and cDNA syntheses was carried out using a commercial kit (TIANGEN Biotech Co., Ltd.). The procedure and conditions of the RT-PCR were followed according to the requirement or the instruction of the PCR reaction kit (Takara Biotechnology Co., Ltd., Dalian, People’s Republic of China), using the ABI7500 RT-PCR System (Thermo Fisher Scientific, Waltham, MA, USA). The amplified DNA was run on 1.5% agarose gels; images were then digitally captured with a charge-coupled device camera and analyzed with an imaging analysis system.
Statistical analysis
The analyses were performed using SPSS 20.0 statistical software (IBM Corporation, Armonk, NY, USA). Student’s t-test was used to evaluate the significance of differences between groups. P-values <0.05 were statistically significant.
Results
Safety evaluation for rdHSV-IFNγ
As therapeutic vectors must display high and proven safety, we evaluated the safety of rdHSV-IFNγ both in vivo and in vitro. There was no evidence of HSV-1 or HSV-1 infection in any of the specific pathogen-free rats when the study began (data not shown). First, we also evaluated the safety of the rdHSV-IFNγ in vivo. Rats were immunized by the topical administration of drops containing rdHSV-IFNγ in their scarified corneas (Figure 2A). Second, we examined the survival rate and infection rate of rdHSV-IFNγ-immunized rats. Kaplan–Meier estimate confirmed that cumulative survival in the rdHSV-IFNγ group was equal to that in the CN group (Figure 2B, P>0.05). Furthermore, the side effects of rdHSV-IFNγ treatment were also evaluated from day 1 to day 21 following the immunization and found no significant difference in the infection rate in the rdHSV-IFNγ group compared with that in the CN group (Figure 2C).
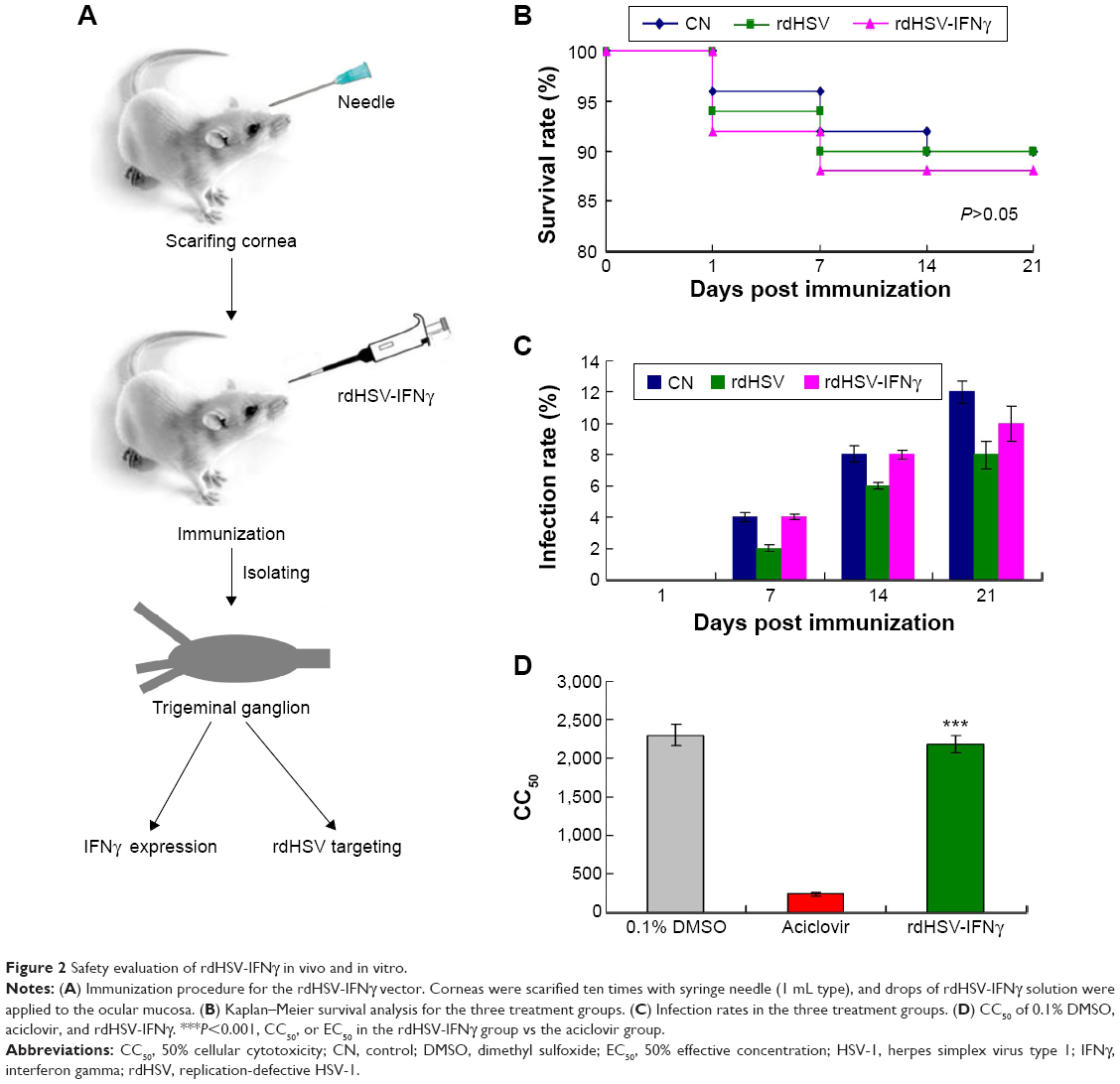 | Figure 2 Safety evaluation of rdHSV-IFNγ in vivo and in vitro. |
In addition, the cytotoxicity of rdHSV-IFNγ in SH-SY5Y cells, as indicated by the CC50 results, was significantly lower than that of aciclovir (P<0.05) and not different from that of 0.1% dimethyl sulfoxide (Figure 2D, P>0.05). Overall, the results support the safety of rdHSV-IFNγ as a therapeutic system for the treatment of latent HSV-1 infection.
Therapeutic rdHSV-IFNγ effectively targets the TG
The targeting efficiency of rdHSV-IFNγ was assayed by the detection and amplification of the cloned IFNγ protein. In this experiment, we isolated and detected the IFNγ protein in cornea, trigeminal nerve, and TG on days 1, 7, 14, and 21 by RT-PCR. The IFNγ mRNA was found in the cornea on days 1 and 7, but the expression had significantly decreased by day 14 and day 21 compared with day 1 and day 7 (P<0.05, Figure 3). Meanwhile, IFNγ mRNA was not found in the TG on days 1 and 7, but the expression increased significantly from day 7 to day 21 (Figure 3). On day 21, IFNγ mRNAs were primarily distributed in the TG and were very low in the cornea. The pattern of IFNγ mRNA distribution over time from day 1 to day 21 led us to conclude that rdHSV-IFNγ could finally target the TG on day 21, which may indicate that day 21 is optimal for the clearance of latent HSV-1 in the HSK rat models (Figure 3).
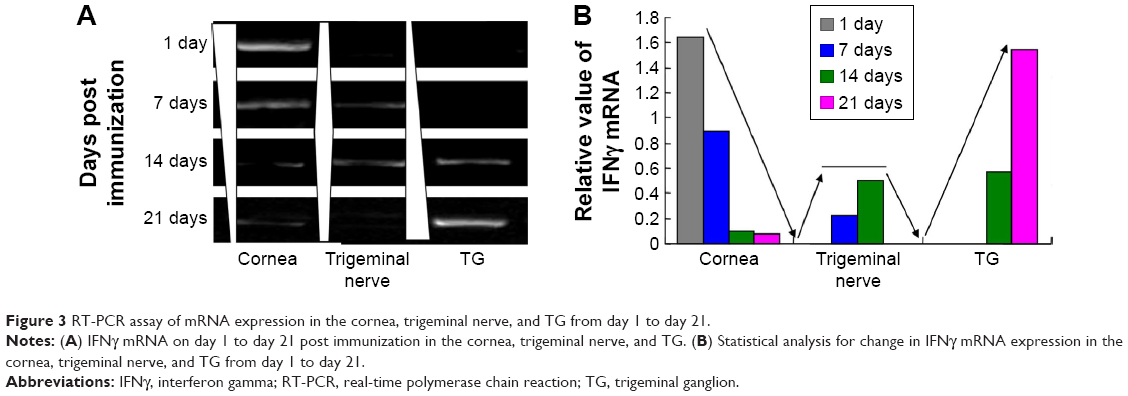 | Figure 3 RT-PCR assay of mRNA expression in the cornea, trigeminal nerve, and TG from day 1 to day 21. |
The rdHSV-IFNγ targeting and expression were also characterized by immunohistochemistry (Figure 4). The results showed that IFNγ protein expression in the cornea decreased significantly from day 1 to day 21 (Figure 4A). The IFNγ protein was located mainly in the cornea on days 1 and 7, but the level was significantly lower on days 14 and 21. IFNγ protein was expressed in the trigeminal nerve on days 7 and 14 but was no longer detected there on day 21 (Figure 4B). Most important, IFNγ expression in the TG began on day 14 and rapidly reached a peak level on day 21 (Figure 4C). IFNγ expression in the TG thus increased with time after immunization.
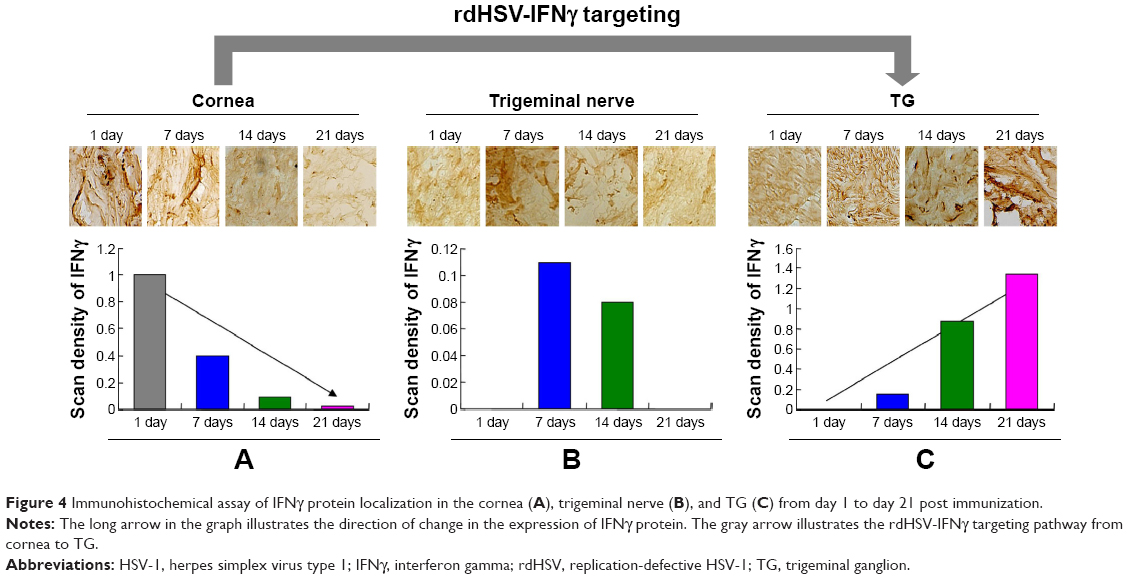 | Figure 4 Immunohistochemical assay of IFNγ protein localization in the cornea (A), trigeminal nerve (B), and TG (C) from day 1 to day 21 post immunization. |
Effective expression of rdHSV-IFNγ in TG tissue
The results described earlier indicated that the rdHSV-IFNγ could effectively localize in TG tissue, where IFNγ mRNA (Figure 5A) and protein expression (Figure 5B) were found to be the strongest on days 14 and 21. The peak expression was coincident with the time that rdHSV-IFNγ was present in the TG. The findings thus suggest that the rdHSV-IFNγ not only targeted the TG tissues but also carried the therapeutic gene to the TG, where it could then clear the latent HSV-1.
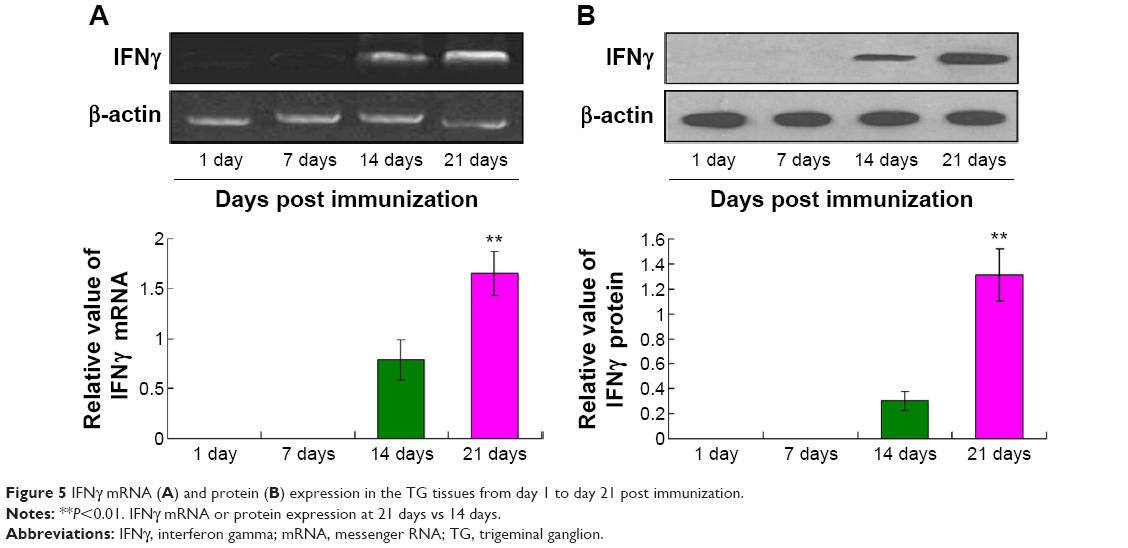 | Figure 5 IFNγ mRNA (A) and protein (B) expression in the TG tissues from day 1 to day 21 post immunization. |
Discussion
Key objectives of drug design are the effective targeting of the diseased region and the induction of protective responses that lead to significantly shortened duration of illness and relief of disease symptoms.7,15 In the present study, we constructed an rdHSV-1 vector with the potential to effectively target the latent HSV-1 in TG tissue. We also cloned a therapeutic antiviral IFNγ gene fragment with the potential of increasing host immunogenicity.
Although there are many methods of delivering genes into mammalian cells in the past few decades, no specific vector targeting the trigeminal nerve or TG has been developed.16–18 Such a vector is especially important for treating HSK, because the available antiviral drugs do not target and eliminate the latent HSV-1 virus in the TG and thus cannot prevent recurrences.19,20 Therefore, we successfully constructed an rdHSV-1 vector system that could be used to effectively deliver therapeutic genes or immune regulators to the TG and clear the latent HSV-1.
In this study, we took an advantage of the neurotrophic properties of HSV-1 to reconstruct an rdHSV-1 vector that had lost its pathogenicity and ability to replicate and was effectively transported into the TG. For a clinically useful therapeutic tool, to confirm its safety in vitro, we transfected SH-SY5Y cells with the rdHSV vector. The results showed that rdHSV is very safe, with a CC50 six- to sevenfold higher than that of aciclovir, which is a drug having a wide clinical use.21 Actually, aciclovir has been used in ophthalmology practice for many years and is clinically effective with few side effects and low cytotoxicity for patients.22,23 Furthermore, we have also immunized the rdHSV-IFNγ vector to the rats and examined the overall survival and the infection rate of the rats in every group. The results also indicated that there was no significant difference between the rdHSV-IFNγ vector treatment group and the CN group. The removal of pathogenic gene fragments and genes needed for replication prevented the occurrence of treatment-related side effects and wild-type viral infections previously seen with some recombinant HSV-1 vectors.24–27 The in vivo and in vitro experiments demonstrated the safety of rdHSV-IFNγ.
To evaluate the targeting effectiveness of rdHSV-IFNγ, we isolated trigeminal ganglia from immunized rats and assayed IFNγ expression on days 1, 7, 14, and 21 after immunization. IFNγ protein was expressed following immunization, and the expression level reflected the amount of rdHSV-IFNγ contained in the TG. The IFNγ mRNA and protein expression results indicate that rdHSV-IFNγ targeted the TG, continuously following a pathway from the cornea to the trigeminal nerve and the TG. Therefore, the rdHSV-IFNγ vector could effectively carry a therapeutic gene to HSV-1 virus-infected cells in the HSK rat model or HSK patients. This characteristic of the rdHSV-IFNγ vector could make the elimination of the latent HSV-1 in the TG possible.
In this study, we cloned the human IFNγ gene and incorporated it into the rdHSV-IFNγ vector. Previous studies28,29 have reported that IFNγ always has both antiviral and immunoregulatory roles in virus-infected cells and the intercellular substance. Upstream of the therapeutic gene insertion location in the rdHSV-IFNγ vector, we added a strong promoter, CMV, to ensure sufficient expression of the therapeutic gene. Our results also confirmed that IFNγ was expressed in the TG where it could carry out its antiviral therapy and immunoregulatory functions.
Conclusion
Overall, the results confirm that rdHSV-IFNγ can act as a vector to carry therapeutic genes to regions with the latent HSV-1 infection and to trigger the expression of the therapeutic genes (eg, the TG in the HSK model). The rdHSV-IFNγ vector carrying therapeutic genes may become a promising method to inhibit the reactivation of the latent HSV-1 and to prevent the recurrence of the HSK in clinical practice; however, much more evidence is required before the clinical application of rdHSV-IFNγ becomes possible. We are planning to investigate the antiviral effects of rdHSV-IFNγ against various HSV-1 strains in this rat model system.
Acknowledgment
This research was supported by the Science and Technology Research and Development Program of Shaanxi Province, People’s Republic of China (Grant No 2011K12-55).
Disclosure
The authors report no conflicts of interest in this work.
References
Lin T, Gong L, Sun XH, et al. Effectiveness and safety of 0.5% ganciclovir in situ ophthalmic gel for herpes simplex keratitis, a multicenter, randomized, investigator-masked, parallel group study in Chinese patients. Drug Des Devel Ther. 2013;7:361–368. | ||
Chen D, Zhang D, Xu L, et al. The alterations of matrix metalloproteinase-9 in mouse brainstem during herpes simplex virus type 1-induced facial palsy. J Mol Neurosci. 2013;51(3):703–709. | ||
Alekseev O, Donovan K, Zaizkhan-Clifford J. Inhibition of ataxia telangiectasia mutated (ATM) kinase suppresses herpes simplex virus type 1 (HSV-1) keratitis. Invest Ophthalmol Vis Sci. 2014;55(2):706–715. | ||
Messer HG, Jacobs D, Dhummakupt A, et al. Inhibition of H3K27me3-specific histone demethylases JMJD3 and UTX blocks reactivation of herpes simplex virus 1 in trigeminal ganglion neurons. J Virol. 2015;89(6):3417–3420. | ||
Sheppard JD, Wertheimer ML, Scoper SV. Modalities to decrease stromal herpes simplex keratitis reactivation rates. Arch Ophthalmol. 2009;127(7):852–856. | ||
Arefian E, Bamdad T, Soleimanjahi H, et al. A kinetic study of gamma interferon production in herpes simplex virus-1 DNA prime-protein boost regimen comparing to DNA or subunit vaccination. Mol Biol (Mosk). 2009;43(3):388–393. | ||
Hu K, Dou J, Yu F, et al. An ocular mucosal administration of nanoparticles containing DNA vaccine Prsc-Gd-IL-21 confers protection against mucosal challenge with herpes simplex virus type 1 in mice. Vaccine. 2011;29(7):1455–1462. | ||
Theopold CP, Gheerardyn R, Bleiziffer O, et al. A novel replication-defective HSV-1 vector for regulatable gene delivery to wounds. J Am Coll Surg. 2004;199(3):57–58. | ||
Palmer JA, Branston RH, Lilley CE, et al. Development and optimization of herpes simplex virus vectors for multiple long-term gene delivery to the peripheral nervous system. J Virol. 2000;74:5604–5618. | ||
Anesti AM, Peeters PJ, Royaux I, et al. Efficient delivery of RNA interference to peripheral neurons in vivo using herpes simplex virus. Nucleic Acids Res. 2008;36(14):e86. | ||
Xu K, Liu XN, Zhang HB, et al. Replication-defective HSV-1 effectively targets targets trigeminal ganglion and inhibits viral pathopoiesis by mediating interferon gamma expression in SH-SY5Y cells. J Mol Neurosci. 2014;53(1):78–86. | ||
Xu K, Wang X, Shi Q, et al. Human prion protein mutants with deleted and inserted octarepeats undergo different pathways to trigger cell apoptosis. J Mol Neurosci. 2011;43(3):225–234. | ||
Chiang LC, Chiang W, Chang MY, et al. Antiviral activity of Plantago major extracts and related compounds in vitro. Antiviral Res. 2002;55(1):53–62. | ||
Wang X, Shi Q, Xu K, et al. Familial CJD associated PrP mutants within transmembrane region induced Ctm-PrP retention in ER and triggered apoptosis by ER stress in SH-SY5Y cells. PLoS One. 2011;6(1):e14602. | ||
Shahidi Bonjar MR, Shahidi Bonjar L. Design of a new therapy for patients with chronic kidney disease: use of microarrays for selective hemoadsorption of uremic wastes and toxins to improve homeostasis. Drug Des Devel Ther. 2015;9:625–629. | ||
Whitehead JL, Ohara PT, Tauscher AN, et al. A procedure to deliver simplex virus to the murine trigeminal ganglion. Brain Res Brain Res Protoc. 2003;12(1):60–66. | ||
Meunier A, Latremoliere A, Mauborgne A, et al. Attenuation of pain-related behavior in a rat model of trigeminal neuropathic pain by viral-driven enkephalin overproduction in trigeminal ganglion neurons. Mol Ther. 2015;11(4):608–616. | ||
Goins WF, Cohen JB, Glorioso JC. Gene therapy for the treatment of chronic peripheral nervous system pain. Neurobiol Dis. 2012;48(2): 255–270. | ||
Farooq AV, Shukla D. Herpes simplex epithelial and stromal keratitis: an epidemiologic update. Surv Ophthalmol. 2012;57(5):448–462. | ||
vanVelzen M, van Loenen FB, Meesters RJ, et al. Latent acyclovir-resistant herpes simplex virus type 1 in trigeminal ganglia of immunocompetent individuals. J Infect Dis. 2012;205(10):1539–1543. | ||
Bieber T, Chosidow O, Bodsworth N, et al. Efficacy and safety of acyclovir mucoadhesive buccal tablet in immunocompetent patients with labial herpes: a double-blind placebo-controlled, self-initiated trial. J Drugs Dermatol. 2014;13(7):791–798. | ||
Colin J, Prisant O, Cochener B, et al. Comparison of the efficacy and safety of valaciclovir and acyclovir for the treatment of herpes zoster ophthalmicus. Ophthalmology. 2000;107(8):1507–1511. | ||
Turner LD, Beckingsale P. Acyclovir-resistant herpetic keratitis in a solid-organ transplant recipient on systemic immunosuppression. Clin Ophthalmol. 2013;7:229–232. | ||
Sia KC, Wang GY, Ho IA, et al. Optimal purification method for herpes-based viral vectors that confers minimal cytotoxicity for systemic route of vector administration. J Virol Methods. 2007;139(2):166–174. | ||
Moriuchi S, Krisky DM, Marconi PC, et al. HSV vector cytotoxicity is inversely correlated with effective TK/GCV suicide gene therapy of rat gliosarcoma. Gene Ther. 2000;7(17):1483–1490. | ||
Goel N, Rong Q, Zimmerman D, et al. ALEAPS heteroconjugate vaccine containing a T cell epitope from HSV-1 glycoprotein D elicits Th1 responses and protection. Vaccine. 2003;21(27–30):4410–4420. | ||
Quenelle DC, Collins DJ, Rice TL, et al. Effect of an immune enhancer, GPI-100, on vaccination with live attenuated herpes simplex virus type 2 or glycoprotein D on genital HSV-2 infections of guinea pigs. Antiviral Res. 2008;80(2):223–224. | ||
Fekete T, Pazmandi K, Szabo A, et al. The antiviral immune response in human conventional dendritic cells is controlled by the mammalian target of rapamycin. J Leukoc Biol. 2014;96(4):579–589. | ||
Wang LC, Cheng SO, Chang SP, et al. Enterovirus 71 proteins 2A and 3D antagonize the antiviral activity of gamma interferon via signaling attenuation. J Virol. 2015;89(14):7028–7037. |
 © 2016 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2016 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.