")
Back to Journals » Infection and Drug Resistance » Volume 17
Epidemiology and Azole Resistance of Clinical Isolates of Aspergillus fumigatus from a Large Tertiary Hospital in Ningxia, China
Authors Kang Y, Li Q, Yao Y, Xu C, Qiu Z, Jia W, Li G, Wang P
Received 6 October 2023
Accepted for publication 18 January 2024
Published 2 February 2024 Volume 2024:17 Pages 427—439
DOI https://doi.org/10.2147/IDR.S440363
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Zhi Ruan
Yuting Kang,1,* Qiujie Li,2,* Yao Yao,3 Chao Xu,2 Zhuoran Qiu,2 Wei Jia,1,3 Gang Li,1,3 Pengtao Wang1
1Ningxia Key Laboratory of Clinical and Pathogenic Microbiology, Institute of Medical Sciences, General Hospital of Ningxia Medical University, Yinchuan, Ningxia, 750004, People’s Republic of China; 2College of Clinical Medicine, Ningxia Medical University, Yinchuan, Ningxia, 750004, People’s Republic of China; 3Center of Medical Laboratory, General Hospital of Ningxia Medical University, Yinchuan, Ningxia, 750004, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Gang Li; Pengtao Wang, General Hospital of Ningxia Medical University, Technology Building 601, 804 Shengli Road, Yinchuan, People’s Republic of China, Tel +86-0951-6744462 ; +86-0951-6743330, Email [email protected]; [email protected]
Purpose: The objective of this study was to determine the clinical distribution, in vitro antifungal susceptibility and underlying resistance mechanisms of Aspergillus fumigatus (A. fumigatus) isolates from the General Hospital of Ningxia Medical University between November 2021 and May 2023.
Methods: Antifungal susceptibility testing was performed using the Sensititre YeastOne YO10, and isolates with high minimal inhibitory concentrations (MICs) were further confirmed using the standard broth microdilution assays established by the Clinical and Laboratory Standards Institute (CLSI) M38-third edition. Whole-Genome Resequencing and RT-qPCR in azole-resistant A. fumigatus strains were performed to investigate the underlying resistance mechanisms.
Results: Overall, a total of 276 A. fumigatus isolates were identified from various clinical departments, showing an increasing trend in the number of isolates over the past 3 years. Two azole-resistant A. fumigatus strains (0.72%) were observed, one of which showed overexpression of cyp51A, cyp51B, cdr1B, MDR1/2, artR, srbA, erg24A, and erg4B, but no cyp51A mutation. However, the other strain harbored two alterations in the cyp51A sequences (L98H/S297T). Therefore, we first described two azole-resistant clinical A. fumigatus strains in Ningxia, China, and reported one azole-resistant strain that has the L98H/S297T mutations in the cyp51A gene without any tandem repeat (TR) sequences in the promoter region.
Conclusions: This study emphasizes the importance of enhancing attention and surveillance of azole-resistant A. fumigatus, particularly those with non-TR point mutations of cyp51A or non-cyp51A mutations, in order to gain a better understanding of their prevalence and spread in the region.
Keywords: Aspergillus fumigatus, azole resistance, cyp51A mutation, tandem repeat sequences
Introduction
Aspergillus fumigatus (A. fumigatus) is the most common Aspergillus species that can cause various serious diseases such as life-threatening invasive aspergillosis (IA) in immunocompromised individuals with hematopoietic stem cells and solid organ transplantation, acute leukemia, and receiving immunotherapy with/without corticosteroids.1–3 Non-invasive infections in immunocompetent patients with underlying pulmonary conditions can lead to chronic pulmonary aspergillosis (CPA), and others can suffer from allergic bronchopulmonary aspergillosis (ABPA), fungal asthma, and Aspergillus bronchitis.4–6 A. fumigatus has been identified as a critical priority fungal pathogen in the first-ever fungal priority pathogen list released by the World Health Organization (WHO) (https://www.who.int/publications/i/item/9789240060241). Recently, there has been a rise in reported cases of IA in patients with influenza7 and coronavirus disease 2019 (COVID-19).8,9 According to existing reports, the prevalence of aspergillosis varies across different provinces in China. A study conducted by Linna Huang et al found that 29.3% of patients with viral pneumonia in Beijing from December 2022 to February 2023 also had IA.10 A retrospective cohort study revealed that between August 2016 and December 2019, a total of 617 adult viral pneumonia patients were admitted to six hospitals in China, with 14.7% patients being diagnosed with IPA.11 In Nanjing, among clinically suspected patients, 26.6% of patients were ultimately diagnosed with invasive pulmonary aspergillosis (IPA),12 which is lower compared to Shanghai, where 58.5% of patients were diagnosed with IPA.13 However, there is currently a lack of epidemiological statistics regarding A. fumigatus infection in Ningxia and its surrounding areas.
Treatment for A. fumigatus has heavily relied on three classes of antifungal drugs: azoles, polyenes, and echinocandins.3 However, the extensive use of azoles such as itraconazole (ITC), voriconazole (VRC), posaconazole (POS), and isavuconazole (ISV) over the past decades has led to the worldwide emergence of azole resistance in A. fumigatus. This increase in resistance, both environmentally and clinically, may be responsible for the failure of aspergillosis treatment and the consequent high mortality rates.14,15
Azole resistance in A. fumigatus is primarily caused by alterations to the sterol biosynthesis pathway, resulting from point mutations of cyp51A and the insertion of tandem repeats (TR) in the promoter region of cyp51A (such as TR34/L98H and TR46/Y121F/T289A).16–18 However, there are increasing reports of A. fumigatus strains that are resistant to clinical azoles despite lacking cyp51A modifications.19,20 Extensive research has been conducted on strains with multiple mutations in hmg1 and hmg2, which encode the 3-hydroxy-3-methylglutaryl-coenzyme A (HMG-CoA) reductase. These mutations have provided valuable insights into the mechanism of azole drug resistance in A. fumigatus.20–25
Several attempts have been made to correlate the mutations in hapE, which encodes one of the three subunits of the CCAAT box binding complex (CBC), with azole resistance. Interestingly, a new study investigated the combination of hapEP88L and hmg1F262del mutations as potential contributors to azole resistance.26–28 In addition to these mutations, other candidate genes such as crd1B, MDR1, MDR2, and erg6 have also been extensively studied and are believed to play a role in non-cyp51A azole resistance.1 The growing number of resistant strains and the continuous emergence of new resistance mechanisms have aroused great concern worldwide.14,29
The epidemiology of aspergillus-disease in Ningxia has not been previously reported, according to an extensive literature search available on PubMed. A. fumigatus infection in China exhibits regional differences.30 General Hospital of Ningxia Medical University, the largest tertiary hospital in Ningxia with approximately 3500 beds, serves both local patients and those from surrounding areas. We believe that our hospital may be regionally representative to some extent. Furthermore, Ningxia, located in northwest China, has a unique geographical climate and lacks research on A. fumigatus infection and drug resistance. Therefore, our study focused on assessing clinical A. fumigatus isolates collected from the General Hospital of Ningxia Medical University from November 2021 to May 2023. We aim to evaluate the susceptibility of these isolates to different antifungal agents and to investigate potential resistance mechanisms, including cyp51A mutations and expression of genes related to azole-resistance.
Materials and Methods
A. fumigatus Strains and Molecular Identification
A. fumigatus isolates were obtained from individual patients hospitalized in the General Hospital of Ningxia Medical University between November 2021 and May 2023. Af293 was kindly donated by Professor Wei Liu of Peking University First Hospital. The fungal isolates were subcultured on sabouraud dextrose agar (SDA) at 25°C for 1 week. Morphological identification was performed based on macro and microscopic features, followed by matrix-assisted laser desorption ionization-time-of-flight mass spectrometry (MALDI-TOF MS, bioMerieux, France). Molecular identification was performed by amplifying the β-tubulin gene (benA) and calmodulin gene (CaM) as previously described,31,32 and the resulting PCR products were sequenced at Tsingke Biotechnology (Beijing, China). The obtained sequences were compared to reference sequences in GenBank. This study was approved by Medical Science Research Ethics Committee IRB of the General Hospital of Ningxia Medical University (2020989 approved on 09 October 2020). Written informed consent was obtained from all study participants (consent for research).
Antifungal Susceptibility Testing
The Sensititre YeastOne YO10 (Thermo Scientific, Cleveland, OH, United States) was used for antifungal susceptibility testing (AFST) with nine antifungals: anidulafungin (AND), micafungin (MFG), caspofungin (CAS), 5-flucytosine (5-FC), posaconazole (POS), voriconazole (VRC), itraconazole (ITC), fluconazole (FLC), amphotericin B (AMB). The values of minimal inhibitory concentration (MIC) and minimum effective concentration (MEC) were determined according to the manufacturer’s instructions. Briefly, conidia were collected from the SDA plates and adjusted to a McFarland standard of 0.5. A 100 µL conidial suspension was added to RPMI-1640 medium to achieve a final working concentration of 0.4–5×104 CFU/mL. 96-well plates with 100 µL working solution in each well were incubated at 35°C for 24–48 h. The minimum inhibitory concentration (MIC) values of POS, VCZ, ITC, and AMB were determined to be the lowest concentrations that produced 100% inhibition, indicated by the first blue or purple well after 48 h of incubation. The minimum effective concentration (MEC) values were determined as the lowest concentrations of AND, MFG, and CAS that produced a morphological change to small, rounded, compact hyphal forms compared to the growth control well after 24 h of incubation. The standard broth microdilution assays were performed in accordance with CLSI M38 third edition33 to determine the isolates with MIC value(s) of VRC≥2 µg/mL or ITC>1 µg/mL base on YeastOne results. For CLSI methodology, the concentration ranges of antifungal agents were as follows: VRC, ITC, POS (0.031–16 µg/mL), CAS, and MFG (0.015–8 µg/mL). The antifungal drugs were purchased from Meilunbio, China. Candida parapsilosis ATCC 22019 and Candida krusei ATCC 6258 were used as quality controls. The epidemiological cut-offs (ECVs) of AMB (2 µg/mL), CAS (0.5 µg/mL), ITC (2 µg/mL), and the clinical breakpoint of VRC (≥2 µg/mL) referred to CLSI-M59 were used to define the isolates as wild type (WT)/Susceptible (S) or non-WT/Resistant (R).34 MIC50 and MIC90 values were determined based on concentrations that inhibited 50% and 90% of the isolates, respectively. The calculation method used in this study can be referenced from the literature guide “Reference Method for Broth Dilution Antifungal Susceptibility Testing of Filamentous Fungi; Approved Standard Second Edition, M38-A2”.
Whole-Genome Resequencing for Azole-Resistant A. fumigatus
In this study, a total of 276 strains of A. fumigatus from Ningxia were labeled as NYDZY 1 to NYDZY 276. Among these strains, NYDZY 162 and NYDZY 247 were identified as being resistant to azole. For the genome resequencing of NYDZY 162 and NYDZY 247, high-purity genomic DNA was extracted using QIAamp DNA Mini Kit (Qiagen). DNA qualification, library construction, and resequencing were executed at DNBSEQ platform.35 High-throughput DNA sequencing (pair-end sequencing) was performed on Pacbio platform, with a read length of 300 bp at each end and an average of 3 Gb sequencing data for each library.
Determination of Azole-Resistance Related Genes Expression Levels by RT-qPCR
The expression levels of cyp51A, cyp51B, cdr1b, atrR, atrF, srbA, erg4A/4B/24A/25A, and MDR1/2/3/4 were determined by real-time qPCR. Briefly, conidia (1.0×106 cells/mL) were collected from SDA plates and incubated at 35°C and 200 rpm for 48 h in 5 mL Sabouraud liquid medium. After centrifugation, the culture supernatant was discarded, mycelia were harvested, and they were washed twice with sterile distilled water. RNA extraction was performed using the RNAsimple Total RNA Kit (TIANGEN, Biotech, China) following the manufacturer’s instructions. The concentration of RNA was determined using NanoDrop spectrophotometer (ThermoFisher), and cDNA was synthesized using a PrimeScript RT Master Mix kit (TaKaRa Biotechnology, China) according to the manufacturer’s instructions. qPCR was conducted with specific primers (Table 1) using LightCycler 480 SYBR green I master mix (Roche). Af293 was used as susceptible control. The difference in gene expression was calculated using the 2-ΔΔCT method. Each sample was performed in triplicate.
 |
Table 1 Sequence Information of the qPCR Primers Used in This Study |
Statistical Analysis
Statistical differences between multiple groups were performed by a one-way analysis of variance (ANOVA) followed by Tukey’s multiple comparisons tests, using GraphPad Prism 8.0 software. Data were presented as medians or means ± standard deviation. P < 0.05 was considered statistically significant.
Results
Fungal Isolates
A total of 276 strains identified as A. fumigatus using morphological and molecular methods and MALDI-TOF MS were collected in this study. The number of A. fumigatus isolates during the first 5 months of 2023 was 192, which is 3.4 times higher than the number during the same period of 2022. Notably, there was a significant increase in January and March of 2023 (Figure 1A). The A. fumigatus were collected from patients mainly distributed in department of respiratory (n=119, 43.12%), emergency department (n=49, 17.75%), intensive care medicine (ICU) (n=22, 7.97%), cardiology (n=12, 4.35%), geriatric (n=9, 3.26%), nephrology (n=7, 2.54%), hematology (n=7, 2.54%), and infection (n=6, 2.17%) (Figure 1B). In terms of specimen types, over 50% of the A. fumigatus isolates (n=246, 89.13%) were recovered from sputum, 9.78% (n=27) were from bronchoalveolar lavage fluid (BALF), 0.72% (n = 2) were from wounds, 0.37% (n = 1) were from puncture fluid (Figure 1C).
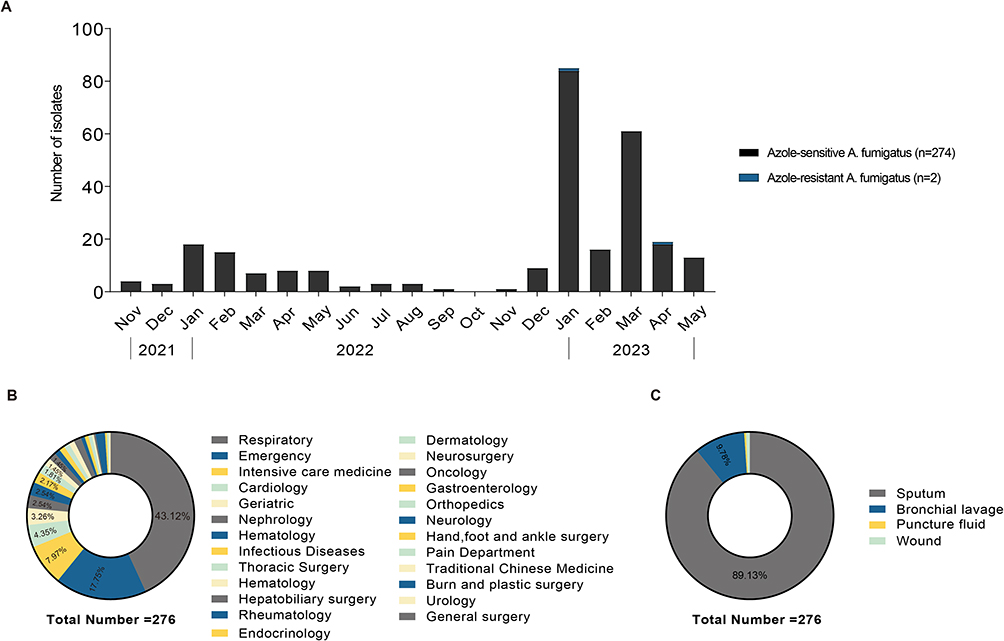 |
Figure 1 Distribution of the 276 A. fumigatus isolates from November 2021-May 2023: (A) Number of A. fumigatus isolates by month of detection; (B) Distribution by department type; (C) Distribution by specimen type. |
Antifungal Susceptibility Testing and Detection of cyp51A
The MIC50, MIC90, range, ECVs, and categories are summarized in Table 2. Among the A. fumigatus isolates, 49.28% (n=136) exhibited high MIC values of 5-FC (>64 µg/mL), while more than 50% of isolates (n=232, 84.06%) demonstrated higher MIC values for FLC (≥256 µg/mL). These findings suggest the inherent resistance of A. fumigatus to these antifungal agents. All strains were wild type (WT) to AMB, with MIC values ranging from 0.25 to 2 µg/mL, and both MIC50 and MIC90 = 1 µg/mL. The MIC values of CAS for three strains (NYDZY 39, NYDZY 65, NYDZY 200) were 8 μg/mL, 8 μg/mL, 4 μg/mL, respectively, which are categorized as Non-WT. However, all three strains showed low MIC values for VRC, ITC, and POS. Two isolates of A. fumigatus (NYDZY 162, NYDZY 247) exhibited higher MIC values of >16 μg/mL for ITC and 2 μg/mL for POS. In the case of VRC, NYDZY 162 exhibited high MIC values of 8 μg/mL, while NYDZY 247 showed values of 4 μg/mL, indicating azole resistance. The distribution of VRC, ITZ, and POS MICs for clinical A. fumigatus isolates is shown in Figure 2. The MIC/MEC values of VRC, ITC, POS, CAS, MFG, and AMB against two azole-resistant isolates, determined by YeastOne YO10 and CLSI methods, are summarized in Table 3. Both methods showed high categorical agreement. Compared to the CLSI method, NYDZY 162 exhibited one-dilution higher MIC values of VRC and one-dilution lower MIC values of AMB. Sequencing results revealed no mutations in the cyp51A coding and promoter region in NYDZY 162, but amid acid substitution was observed in NYDZY 247.
 |
Table 2 Susceptibility Profiles of 276 A. Fumigatus Isolates |
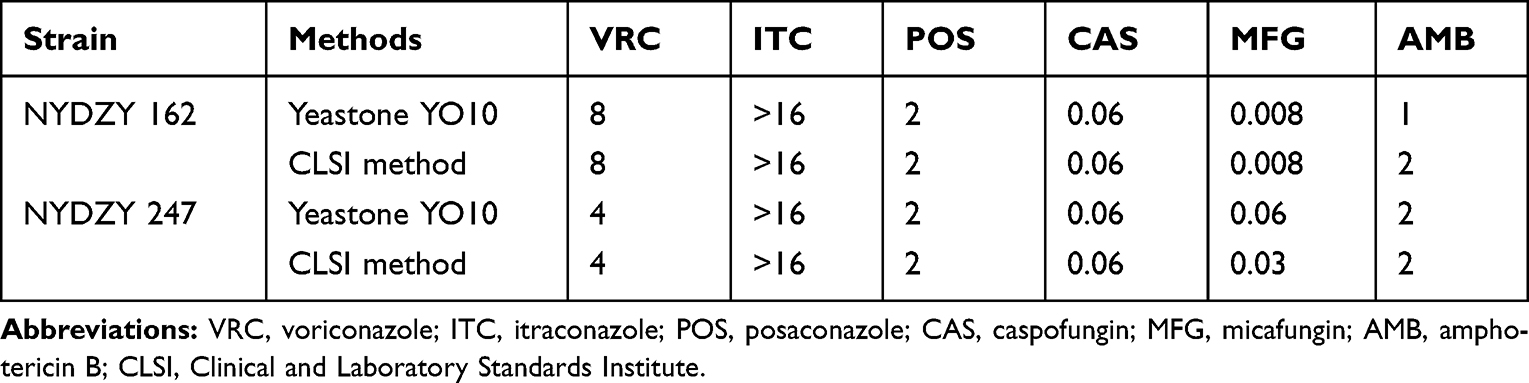 |
Table 3 The MIC/MEC Values (µg/mL) of Different Antifungals Against Two A. Fumigatus Isolates as Determined by the YeastOne YO10 and CLSI Methods |
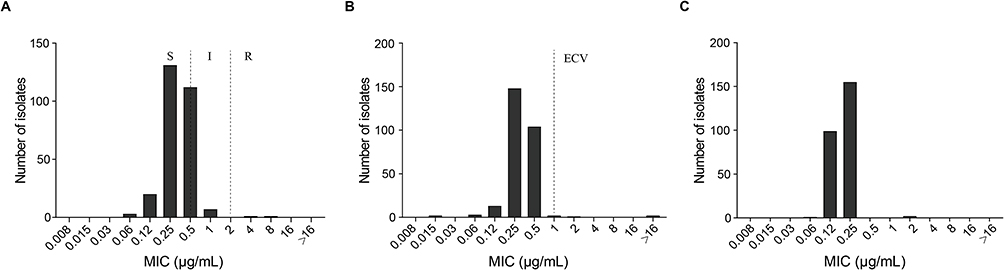 |
Figure 2 MICs’ distributions of antifungals tested for 276 clinical A. fumigatus isolates with YeastOne YO10. (A) voriconazole; (B) itraconazole; (C) posaconazole. Abbreviations: S, susceptible; I, intermediate; R, resistant; ECV, epidemiological cut-offs. |
The Expression Levels of Azole-Resistance Related Genes
RT-PCR results revealed that A. fumigatus clinical isolates (NYDZY 162) resistant to triazoles showed increased expression of cyp51A (P < 0.05), cdr1B (P < 0.0001), MDR1 (P < 0.0001), MDR2 (P < 0.01), artR (P < 0.01), srbA (P < 0.001), erg24A (P < 0.01), and erg4B (P < 0.01) compared to the wild-type Af293. However, there was no significant difference in the expression of these genes in NYDZY 247 (P > 0.05). On the contrary, NYDZY 247 exhibited significantly lower levels of erg24A (P < 0.001) and erg4B (P < 0.05) expression compared to both Af293 and NYDZY 162. Additionally, the expression levels of genes MDR3 (P < 0.0001, P < 0.0001), erg25A (P < 0.01, P < 0.001), and erg4A (P < 0.0001, P < 0.0001) significantly decreased in both NYDZY 162 and NYDZY 247 (Figure 3).
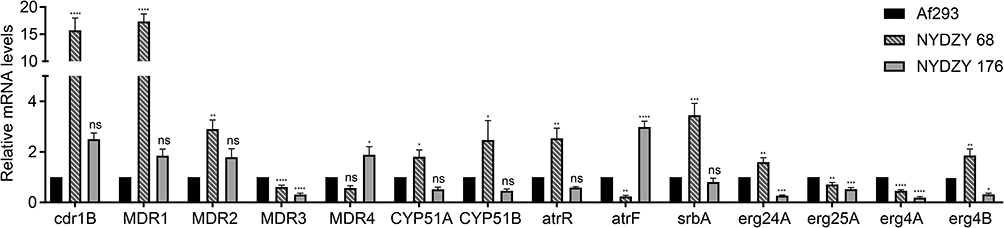 |
Figure 3 RT-qPCR of azole-resistance related genes’ expression (*P < 0.05; **P < 0.01; ***P < 0.001; ****P < 0.0001 in one-way ANOVA with Tukey’s multiple comparison). Abbreviation: ns, non-significant. |
The Clinical Profiles of Patients with Azole-Resistant A. fumigatum
NYDZY 162 was isolated from the sputum of a severe COVID-19 patient (Case 1) who also had non-Hodgkin’s lymphoma, diabetes, hypertension, arrhythmia, and other conditions. Chest CT scans revealed images of lungs with abnormal infiltrates, reticular and linear opacities, pleural thickening, and the accumulation of lung lesions on both sides. After receiving treatment with voriconazole and ceftazidime, the patient’s condition improved significantly, leading to his discharge. NYDZY 247 was isolated from the sputum of a patient (Case 2) with novel coronavirus pneumonia who also had co-infection with bacteria and fungi during the acute exacerbation stage of chronic obstructive pulmonary disease. This patient had a history of old pulmonary tuberculosis, old bronchial tuberculosis, pulmonary hypertension, and other diseases. The chest CT scan results showed the accumulation of lung lesions on both sides, and features like reticular and linear opacities, nodule, pleural thickening, and pleural effusion. The condition of this patient improved after treatment with voriconazole, carbofenozin, imipenem/cilastatin, cefoperazone/sulbactam. The clinical profiles of patients with azole-resistant A. fumigatus was shown in Table 4. It is worth noting that the serum galactomannan (GM) tests were negative in both patients infected with azole-resistant A. fumigatus strains.
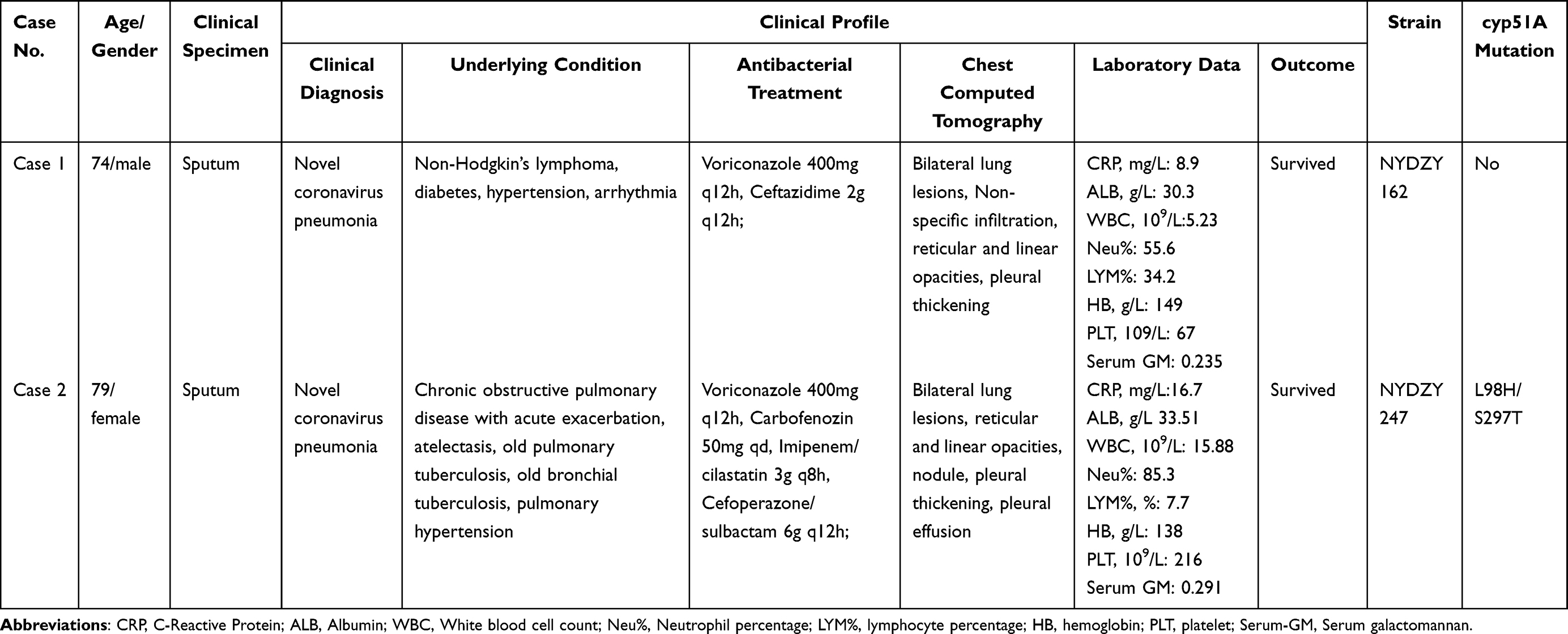 |
Table 4 The Clinical Profile and Examinations of Patients Infected by Resistant A. Fumigatus |
Discussion
A. fumigatus is a globally prevalent environmental mold that can cause a variety of human diseases, leading to notable mortality. Around 3,000,000 cases of chronic pulmonary aspergillosis (CPA) and invasive aspergillosis (IA) are reported worldwide annually.39,40 Moreover, the extensive use of triazole antifungals has resulted in the emergence of azole-resistant A. fumigatus, which poses significant clinical and economic challenges.41–44 Our study conducted in Ningxia province, China, observed an increase in the number of identified A. fumigatus strains from 2021 to 2023. Additionally, we discovered two azole-resistant strains during our research. These findings highlight a concerning trend of A. fumigatus infection in the area.
In our study, we collected a total of 276 strains of A. fumigatus. Among these, 192 strains were collected in 2023, which represents a more than 2.5 times increase compared to the previous year. This significant rise in isolates was strongly correlated with the surge in the number of COVID-19 infected patients following the lifting of epidemic control restrictions. This is similar to studies conducted in other provinces in China.45 It has been reported worldwide that there is a high incidence of COVID-19-Associated Pulmonary Aspergillosis (CAPA) among critically ill COVID-19 patients,46–49 which is believed to be caused by severe immunomodulation and lymphocyte depletion resulting from the virus and the subsequent use of immune-suppressive drugs.50 The most common methods used for diagnosing CAPA include recovering Aspergillus spp. in culture media of bronchoalveolar fluid (BALF) and tracheal aspirate, as well as detecting conventional GM from BALF, tracheal aspirate, and serum specimens.51 However, in our study, diagnosing and confirming CAPA in two patients with isolated azole-resistant A. fumigatus remains challenging due to the patients’ serious underlying conditions, the overlap of clinical manifestations and imaging findings between COVID-19 and CAPA, the limited use of bronchoscopies to avoid the risk of virus transmission, and the imperfect sensitivity of serum GM.1,52,53 Fortunately, both patients recovered and were discharged after receiving treatment with voriconazole and antibacterials, highlighting the importance of combination therapy and appropriate dosage for severe infections. But other researchers have indicated that patients infected with azole-resistant strains have a higher mortality rate compared to those infected with sensitive strains.1,54 A 5-year retrospective cohort study showed a 21% higher day-42 mortality in azole-resistant invasive aspergillosis compared with wild-type infections cases.55 In another multicentre retrospective study, it was also observed that the mortality rate at 6 and 12 weeks was higher in haematology patients with voriconazole-resistant cases compared to those with susceptible cases.56 Therefore, it is an urgent issue to monitor drug resistance in clinical isolates and unravel the molecular mechanisms underlying azole resistance.
In addition to patients with associated diseases, strains with azole-resistance have also been identified in patients with no previous history of antifungal treatment.57,58 Furthermore, environmental isolates have been found to develop azole-resistance due to the use of fungicides. Molecular epidemiological studies have demonstrated that people can acquire infections from their immediate surroundings.59 The emergence of these resistant A. fumigatus in both natural and clinical settings has led to an expansion of the population of patients at risk, which has generated well-founded concerns in the face of limited treatment options.
The prevalence of azole-resistant Aspergillus species varies geographically. Our study found that 2 out of 276 samples (0.72%) were resistant to VRC and non-wild type for ITC, which is lower compared to Beijing (4.3%),60 Shanghai (3.57–7.02%),61,62 and Anhui (5.79%)63 in China. In a separate study conducted in China, A. fumigatus isolates were collected from various regions, revealing a 4.4% frequency of azole resistance.30 However, when compared to several other countries, the rates of azole-non-wild type A. fumigatus were found to be higher in Europe (9.5%) and North America (9.1%) than in Latin America and the Asia-Pacific region (5.3%) over a 5-year surveillance period.42
Azole resistance in A. fumigatus is primarily linked to mutations in cyp51A and overexpression of cyp51A, as well as overexpression of efflux pumps. Point mutations in the coding gene in cyp51A combined with tandem repeats in the promoter region have been frequently observed, such as TR34/L98H, which means that the sequence of tandem repeats 34 times, followed by the presence of the amino acid substitution from leucine (L) to histidine (H) at position 98. Mechanically, tandem repeats within the promoter region of cyp51A is associated with overexpression of cyp51A and the L98H substitution decreases hydrogen bonding between residue 98 and the polar side chains of neighboring residues, thereby preventing the triazole from fitting into the binding pocket.64,65 Additionally, strains carrying a TR46 allele in conjunction with Y121F and T289A exchanges were also currently the most prevalent isolates worldwide.1 The sequencing analysis results in our study revealed that NYDZY 247 carried L98H/S297T mutations in the cyp51A gene without the presence of TR in the promoter region (TR34 and TR46), which was quite different from other previous studies where the occurrence of such amino acid substitution was always accompanied by TR variations.66–69 The mRNA level of cyp51A did not show any increase in NYDZY 247, which could be explained by the presence of non-TR mutations in cyp51A. Other single nucleotide polymorphisms (SNPs), such as G54, M220, and G448, in the cyp51A gene have been linked to drug resistance.29 These polymorphisms are frequently observed in patients with chronic pulmonary aspergillosis, invasive aspergillosis bronchitis, aspergilloma, and chronic cavitary pulmonary aspergillosis (CCPA) who have undergone long-term treatment with azole antifungals and have been associated with several clinical treatment failures.37,70–72 The specific amino acid substitution in the cyp51A sequence without tandem repeats determines the alteration in the interaction between 14α-demethylase and the azole drugs, leading to different patterns of azole cross-resistance.73 Therefore, Non-TR mutation of cyp51A in A. fumigatus can still lead to drug resistance and have significant clinical consequences, which highlights the importance of closely monitoring such mutations.
Findings from several groups have shown that the transcription factors AtrR and SrbA play a significant role in azole resistance in A. fumigatus. These factors co-regulate the expressions of cyp51A and efflux transporter cdr1B,36,74,75 which aligns with our main finding that NYDZY 247 exhibited co-upregulation of these genes. Moreover, several clinical azole-resistant A. fumigatus isolates have shown high levels of cyp51B expression, even in the absence of cyp51A mutations,76,77 similar to what has been observed in NYDZY 162. Recent evidence suggests that cyp51B may act as a backup system when cyp51A expression is compromised.78 Consistent with the results of other studies, our findings also showed higher levels of cyp51A and cdr1B expression in the azole-resistant strain NYDZY 162. However, there were some observed differences: NYDZY 162 exhibited a greater increase in cdr1B expression compared to cyp51A, whereas other studies reported a >500-fold induction of cyp51A and only a modest change in cdr1B expression (>5-fold).79 These results imply that the increased expression of the efflux transporter gene in NYDZY 162 may be the primary cause of drug resistance.
Although antifungal susceptibility testing of A. fumigatus is not routinely performed in most clinical laboratories, regular assessment of it can be helpful in guiding clinical management and understanding changes in drug resistance. Prior to our study, no reports had been made regarding azole-resistant A. fumigatus in Ningxia. Although we only identified two azole-resistant strains, this finding represents the first occurrence in the entire northwest region of China. Given the increasing trend in the number of A. fumigatus isolates from 2022 to 2023, it is likely that more azole-resistant strains will be found in the future. Moreover, we report a novel resistance mechanism in one of the azole-resistant strains, which involves the non-TR L98H/S297T mutations of the cyp51A gene. However, it should be noted that our investigation is not exhaustive, and further research will be conducted to delve into the underlying mechanisms in our future work.
Conclusions
This study, for the first time, revealed a prevalence of azole resistance in A. fumigatus at 0.72% in Ningxia province of China. Among the isolates, NYDZY 162 and NYDZY 247 exhibited the same azole resistance phenotype with different resistance mechanisms. NYDZY 162 exhibited overexpression of cyp51A without any mutations, while NYDZY 247 showed non-TR point mutations (L98H/S297T) in cyp51A that have not been previously reported. Our study emphasized the severity of A. fumigus spread in the region and the emergence of azole resistance. Therefore, it is crucial to consistently enhance the surveillance of antifungal susceptibility in clinical strains and implement effective measures to eliminate transmission and nosocomial infections, which may be helpful to prevent further exacerbation of clinical azole resistance.
Data Sharing Statement
The datasets generated during and/or analyzed during the current study are available from the corresponding author, Pengtao Wang, upon reasonable request.
Ethics Statement
This study was approved by Medical Science Research Ethics Committee IRB of the General Hospital of Ningxia Medical University (2020989, approved 09 October 2020). Written informed consent was obtained from all study participants (consent for research). This study follows the Declaration of Helsinki.
Author Contributions
All authors contributed significantly to the work reported, whether in terms of conception, study design, acquisition of data, analysis and interpretation, or all of these; participated in the drafting, revision, or critical review of the article; provided final approval of the version to be published; agreed on the journal to which the article was to be submitted; and agreed to take responsibility for all aspects of the work.
Funding
This work was supported by Grants from the Key Research and Development Project of Ningxia Hui Autonomous Region (No. 2021BEG03090, Wei Jia), Natural Science Foundation of Ningxia Province (No. 2021AAC03366, Pengtao Wang; No. 2022AAC03470, Yuting Kang).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Arastehfar A, Carvalho A, Houbraken J, et al. Aspergillus fumigatus and aspergillosis: from basics to clinics. Stud Mycol. 2021;100:100115. doi:10.1016/j.simyco.2021.100115
2. Li E, Knight JM, Wu Y, et al. Airway mycosis in allergic airway disease. Adv Immunol. 2019;142:85–140. doi:10.1016/bs.ai.2019.05.002
3. Latgé JP, Chamilos G. Aspergillus fumigatus and Aspergillosis in 2019. Clin Microbiol Rev. 2019;33(1):e00140–00118. doi:10.1128/CMR.00140-18
4. Agarwal R, Muthu V, Sehgal IS. Relationship between Aspergillus and asthma. Allergol Int. 2023;72(4):507–520. doi:10.1016/j.alit.2023.08.004
5. Earle K, Valero C, Conn DP, et al. Pathogenicity and virulence of Aspergillus fumigatus. Virulence. 2023;14(1):2172264. doi:10.1080/21505594.2023.2172264
6. Steels S, Proesmans M, Bossuyt X, Dupont L, Frans G. Laboratory biomarkers in the diagnosis and follow-up of treatment of allergic bronchopulmonary aspergillosis in cystic fibrosis. Crit Rev Clin Lab Sci. 2023;60(1):1–24. doi:10.1080/10408363.2022.2101612
7. Chong WH, Saha BK, Tan CK. Clinical characteristics and outcomes of influenza-associated pulmonary aspergillosis among critically ill patients: a systematic review and meta-analysis. J Hosp Infect. 2022;120:98–109. doi:10.1016/j.jhin.2021.11.016
8. Skóra M, Gajda M, Namysł M, et al. COVID-19-associated pulmonary aspergillosis in intensive care unit patients from Poland. J Fungi. 2023;9(6):666. doi:10.3390/jof9060666
9. Bergmann F, Jorda A, Blaschke A, et al. Pulmonary Aspergillosis in critically ill COVID-19 patients admitted to the intensive care unit: a retrospective cohort study. J Fungi. 2023;9(3):315. doi:10.3390/jof9030315
10. Zhou X, Wu X, Chen Z, et al. Risk factors and the value of microbiological examinations of COVID-19 associated pulmonary aspergillosis in critically ill patients in intensive care unit: the appropriate microbiological examinations are crucial for the timely diagnosis of CAPA. Front Cell Infect Microbiol. 2023;13:1287496. doi:10.3389/fcimb.2023.1287496
11. Li L, Wang C, Ren Y, Liu J, Liu F, Zhang X. Clinical features and risk factors for viral pneumonia complicated with invasive pulmonary aspergillosis in adult patients. Chin Med J. 2023;136(11):1361–1363. doi:10.1097/CM9.0000000000002354
12. Lu Y, Liu L, Li H, et al. The clinical value of Aspergillus-specific IgG antibody test in the diagnosis of nonneutropenic invasive pulmonary aspergillosis. Clin Microbiol Infect. 2023;29(6):797.e1–797.e7. doi:10.1016/j.cmi.2023.02.002
13. Zhu N, Zhou D, Xiong W, Zhang X, Li S. Performance of mNGS in bronchoalveolar lavage fluid for the diagnosis of invasive pulmonary aspergillosis in non-neutropenic patients. Front Cell Infect Microbiol. 2023;13:1271853. doi:10.3389/fcimb.2023.1271853
14. Pimienta DA, Cruz Mosquera FE, Palacios Velasco I, Giraldo Rodas M, Oñate-Garzón J, Liscano Y. Specific focus on antifungal peptides against azole resistant aspergillus fumigatus: current status, challenges, and future perspectives. J Fungi. 2022;9(1):42. doi:10.3390/jof9010042
15. Fisher MC, Alastruey-Izquierdo A, Berman J, et al. Tackling the emerging threat of antifungal resistance to human health. Nat Rev Microbiol. 2022;20(9):557–571. doi:10.1038/s41579-022-00720-1
16. da Fonseca L, Braga VF, Tonani L, et al. Surveillance of Amphotericin B and azole resistance in aspergillus isolated from patients in a tertiary teaching hospital. J Fungi. 2023;9(11):1070. doi:10.3390/jof9111070
17. Simmons BC, Rhodes J, Rogers TR, et al. Genomic epidemiology identifies azole resistance due to TR(34)/L98H in European aspergillus fumigatus causing COVID-19-associated pulmonary aspergillosis. J Fungi. 2023;9(11):1104. doi:10.3390/jof9111104
18. Pérez-Cantero A, López-Fernández L, Guarro J, Capilla J. Azole resistance mechanisms in Aspergillus: update and recent advances. Int J Antimicrob Agents. 2020;55(1):105807. doi:10.1016/j.ijantimicag.2019.09.011
19. Zhu G, Chen S, Zhang Y, Lu L. Mitochondrial membrane-associated protein mba1 confers antifungal resistance by affecting the production of reactive oxygen species in aspergillus fumigatus. Antimicrob Agents Chemother. 2023;67(8):e0022523. doi:10.1128/aac.00225-23
20. Hagiwara D, Arai T, Takahashi H, Kusuya Y, Watanabe A, Kamei K. Non-cyp51A azole-resistant aspergillus fumigatus isolates with mutation in HMG-CoA reductase. Emerg Infect Dis. 2018;24(10):1889–1897. doi:10.3201/eid2410.180730
21. Rybak JM, Ge W, Wiederhold NP, et al. Mutations in hmg1, challenging the paradigm of clinical triazole resistance in aspergillus fumigatus. mBio. 2019;10(2):e00437–00419. doi:10.1128/mBio.00437-19
22. Gonzalez-Jimenez I, Lucio J, Roldan A, Alcazar-Fuoli L, Mellado E. Are point mutations in HMG-CoA reductases (Hmg1 and Hmg2) a step towards azole resistance in aspergillus fumigatus. Molecules. 2021;26(19):5975. doi:10.3390/molecules26195975
23. Arai T, Umeyama T, Majima H, et al. Hmg1 mutations in Aspergillus fumigatus and their contribution to triazole susceptibility. Med Mycol. 2021;59(10):980–984. doi:10.1093/mmy/myab026
24. Wu CJ, Liu WL, Lai CC, et al. Multicenter study of azole-resistant Aspergillus fumigatus Clinical Isolates, Taiwan(1). Emerg Infect Dis. 2020;26(4):804–806. doi:10.3201/eid2604.190840
25. Liang T, Yang X, Li R, et al. Emergence of W272C substitution in hmg1 in a triazole-resistant isolate of aspergillus fumigatus from a Chinese patient with chronic cavitary pulmonary aspergillosis. Antimicrob Agents Chemother. 2021;65(7):e0026321. doi:10.1128/AAC.00263-21
26. Camps SM, Dutilh BE, Arendrup MC, et al. Discovery of a HapE mutation that causes azole resistance in Aspergillus fumigatus through whole genome sequencing and sexual crossing. PLoS One. 2012;7(11):e50034. doi:10.1371/journal.pone.0050034
27. Hortschansky P, Misslinger M, Mörl J, et al. Structural basis of HapE(P88L)-linked antifungal triazole resistance in Aspergillus fumigatus. Life Sci Alliance. 2020;3(7):e202000729. doi:10.26508/lsa.202000729
28. Souza A, Ge W, Wiederhold NP, Rybak JM, Fortwendel JR, Rogers PD. hapE and hmg1 mutations are drivers of cyp51A-independent pan-triazole resistance in an aspergillus fumigatus clinical isolate. Microbiol Spectr. 2023;11(3):e0518822. doi:10.1128/spectrum.05188-22
29. Lockhart SR, Chowdhary A, Gold J. The rapid emergence of antifungal-resistant human-pathogenic fungi. Nat Rev Microbiol. 2023;21(12):818–832. doi:10.1038/s41579-023-00960-9
30. Deng S, Zhang L, Ji Y, et al. Triazole phenotypes and genotypic characterization of clinical Aspergillus fumigatus isolates in China. Emerg Microbes Infect. 2017;6(12):e109. doi:10.1038/emi.2017.97
31. Hong SB, Go SJ, Shin HD, Frisvad JC, Samson RA. Polyphasic taxonomy of Aspergillus fumigatus and related species. Mycologia. 2005;97(6):1316–1329. doi:10.3852/mycologia.97.6.1316
32. Sabino R, Gonçalves P, Martins Melo A, et al. Trends on aspergillus epidemiology-perspectives from a national reference laboratory surveillance program. J Fungi. 2021;7(1):28. doi:10.3390/jof7010028
33. CLSl. Reference Method for Broth Dilution Antifungal Susceptibility Testing of Filamentous Fungi.
34. CLSI Epidemiological cutoff values for antifungal susceptibility testing. In: CLSI Supplement M59.
35. Li Q, Zhao X, Zhang W, et al. Reliable multiplex sequencing with rare index mis-assignment on DNB-based NGS platform. BMC Genomics. 2019;20(1):215. doi:10.1186/s12864-019-5569-5
36. Hagiwara D, Miura D, Shimizu K, et al. A novel Zn2-Cys6 Transcription Factor AtrR plays a key role in an azole resistance mechanism of aspergillus fumigatus by co-regulating cyp51A and cdr1B expressions. PLoS Pathog. 2017;13(1):e1006096. doi:10.1371/journal.ppat.1006096
37. Chen J, Li H, Li R, Bu D, Wan Z. Mutations in the cyp51A gene and susceptibility to itraconazole in Aspergillus fumigatus serially isolated from a patient with lung aspergilloma. J Antimicrob Chemother. 2005;55(1):31–37. doi:10.1093/jac/dkh507
38. Long N, Xu X, Zeng Q, Sang H, Lu L. Erg4A and Erg4B are required for conidiation and azole resistance via regulation of ergosterol biosynthesis in aspergillus fumigatus. Appl Environ Microbiol. 2017;83(4):e02924–02916. doi:10.1128/AEM.02924-16
39. Bongomin F, Gago S, Oladele RO, Denning DW. Global and multi-national prevalence of fungal diseases-estimate precision. J Fungi. 2017;3(4):57. doi:10.3390/jof3040057
40. Denning DW. The ambitious ‘95-95 by 2025’ roadmap for the diagnosis and management of fungal diseases. Thorax. 2015;70(7):613–614. doi:10.1136/thoraxjnl-2015-207305
41. Chang H, Ashu E, Sharma C, Kathuria S, Chowdhary A, Xu J. Diversity and origins of Indian multi-triazole resistant strains of Aspergillus fumigatus. Mycoses. 2016;59(7):450–466. doi:10.1111/myc.12494
42. Pfaller MA, Carvalhaes CG, Castanheira M. Susceptibility patterns of amphotericin B, itraconazole, posaconazole, voriconazole and caspofungin for isolates causing invasive mould infections from the SENTRY Antifungal Surveillance Program (2018–2021) and application of single-site epidemiological cutoff values to evaluate amphotericin B activity. Mycoses. 2023. doi:10.1111/myc.13620
43. Debergh H, Becker P, Vercammen F, et al. Pulmonary aspergillosis in humboldt penguins-susceptibility patterns and molecular epidemiology of clinical and environmental aspergillus fumigatus isolates from a Belgian zoo, 2017–2022. Antibiotics. 2023;12(3):584. doi:10.3390/antibiotics12030584
44. Huygens S, Dunbar A, Buil JB, et al. Clinical impact of polymerase chain reaction-based aspergillus and azole resistance detection in invasive aspergillosis: a prospective multicenter study. Clin Infect Dis. 2023;77(1):38–45. doi:10.1093/cid/ciad141
45. Steenwyk JL, Mead ME, de Castro PA, et al. Genomic and phenotypic analysis of COVID-19-associated pulmonary aspergillosis isolates of aspergillus fumigatus. Microbiol Spectr. 2021;9(1):e0001021. doi:10.1128/Spectrum.00010-21
46. Bartoletti M, Pascale R, Cricca M, et al. Epidemiology of invasive pulmonary aspergillosis among intubated patients with COVID-19: a prospective study. Clin Infect Dis. 2021;73(11):e3606–e3614. doi:10.1093/cid/ciaa1065
47. Evert K, Dienemann T, Brochhausen C, et al. Autopsy findings after long-term treatment of COVID-19 patients with microbiological correlation. Virchows Arch. 2021;479(1):97–108. doi:10.1007/s00428-020-03014-0
48. Marr KA, Platt A, Tornheim JA, et al. Aspergillosis complicating severe coronavirus disease. Emerg Infect Dis. 2021;27(1):18–25. doi:10.3201/eid2701.202896
49. Janssen N, Nyga R, Vanderbeke L, et al. Multinational observational cohort study of COVID-19-Associated pulmonary aspergillosis(1). Emerg Infect Dis. 2021;27(11):2892–2898. doi:10.3201/eid2711.211174
50. Salmanton-García J, Sprute R, Stemler J, et al. COVID-19-associated pulmonary aspergillosis, March-August 2020. Emerg Infect Dis. 2021;27(4):1077–1086. doi:10.3201/eid2704.204895
51. Arastehfar A, Carvalho A, van de Veerdonk FL, et al. COVID-19 associated pulmonary aspergillosis (CAPA)-from immunology to treatment. J Fungi. 2020;6(2):91. doi:10.3390/jof6020091
52. Alanio A, Dellière S, Fodil S, Bretagne S, Mégarbane B. Prevalence of putative invasive pulmonary aspergillosis in critically ill patients with COVID-19. Lancet Respir Med. 2020;8(6):e48–e49. doi:10.1016/S2213-2600(20)30237-X
53. Koehler P, Cornely OA, Böttiger BW, et al. COVID-19 associated pulmonary aspergillosis. Mycoses. 2020;63(6):528–534. doi:10.1111/myc.13096
54. Rybak JM, Fortwendel JR, Rogers PD. Emerging threat of triazole-resistant Aspergillus fumigatus. J Antimicrob Chemother. 2019;74(4):835–842. doi:10.1093/jac/dky517
55. Lestrade PP, Bentvelsen RG, Schauwvlieghe A, et al. Voriconazole resistance and mortality in invasive aspergillosis: a multicenter retrospective cohort study. Clin Infect Dis. 2019;68(9):1463–1471. doi:10.1093/cid/ciy859
56. Resendiz-Sharpe A, Mercier T, Lestrade P, et al. Prevalence of voriconazole-resistant invasive aspergillosis and its impact on mortality in haematology patients. J Antimicrob Chemother. 2019;74(9):2759–2766. doi:10.1093/jac/dkz258
57. Wang F, Yao S, Cao D, et al. Increased triazole-resistance and cyp51A mutations in Aspergillus fumigatus after selection with a combination of the triazole fungicides difenoconazole and propiconazole. J Hazard Mater. 2020;400:123200. doi:10.1016/j.jhazmat.2020.123200
58. Cao D, Yao S, Zhang H, et al. Mutation in cyp51A and high expression of efflux pump gene of Aspergillus fumigatus induced by propiconazole in liquid medium and soil. Environ Pollut. 2020;256:113385. doi:10.1016/j.envpol.2019.113385
59. Sewell TR, Zhu J, Rhodes J, et al. Nonrandom distribution of azole resistance across the global population of aspergillus fumigatus. mBio. 2019;10(3):e00392–00319. doi:10.1128/mBio.00392-19
60. Yang X, Chen W, Liang T, et al. A 20-Year Antifungal Susceptibility Surveillance (From 1999 to 2019) for Aspergillus spp. and Proposed Epidemiological Cutoff Values for Aspergillus fumigatus and Aspergillus flavus: a Study in a Tertiary Hospital in China. Front Microbiol. 2021;12:680884. doi:10.3389/fmicb.2021.680884
61. Zhang Y, Wang S, Zhou C, et al. Epidemiology of clinically significant aspergillus species from a large tertiary hospital in shanghai, china, for the period of two years. Infect Drug Resist. 2023;16:4645–4657. doi:10.2147/IDR.S417840
62. Xu Y, Chen M, Zhu J, et al. Aspergillus species in lower respiratory tract of hospitalized patients from shanghai, china: species diversity and emerging azole resistance. Infect Drug Resist. 2020;13:4663–4672. doi:10.2147/IDR.S281288
63. Wang Y, Zhang L, Zhou L, Zhang M, Xu Y. Epidemiology drug susceptibility, and clinical risk factors in patients with invasive aspergillosis. Front Public Health. 2022;10:835092. doi:10.3389/fpubh.2022.835092
64. Nash A, Rhodes J. Simulations of CYP51A from Aspergillus fumigatus in a model bilayer provide insights into triazole drug resistance. Med Mycol. 2018;56(3):361–373. doi:10.1093/mmy/myx056
65. Dudakova A, Spiess B, Tangwattanachuleeporn M, et al. Molecular tools for the detection and deduction of azole antifungal drug resistance phenotypes in aspergillus species. Clin Microbiol Rev. 2017;30(4):1065–1091. doi:10.1128/CMR.00095-16
66. Mortensen KL, Mellado E, Lass-Flörl C, Rodriguez-Tudela JL, Johansen HK, Arendrup MC. Environmental study of azole-resistant Aspergillus fumigatus and other aspergilli in Austria, Denmark, and Spain. Antimicrob Agents Chemother. 2010;54(11):4545–4549. doi:10.1128/AAC.00692-10
67. van der Linden JW, Camps SM, Kampinga GA, et al. Aspergillosis due to voriconazole highly resistant Aspergillus fumigatus and recovery of genetically related resistant isolates from domiciles. Clin Infect Dis. 2013;57(4):513–520. doi:10.1093/cid/cit320
68. Bader O, Tünnermann J, Dudakova A, Tangwattanachuleeporn M, Weig M, Groß U. Environmental isolates of azole-resistant Aspergillus fumigatus in Germany. Antimicrob Agents Chemother. 2015;59(7):4356–4359. doi:10.1128/AAC.00100-15
69. Verweij PE, Snelders E, Kema GH, Mellado E, Melchers WJ. Azole resistance in Aspergillus fumigatus: a side-effect of environmental fungicide use. Lancet Infect Dis. 2009;9(12):789–795. doi:10.1016/S1473-3099(09)70265-8
70. Snelders E, van der Lee HA, Kuijpers J, et al. Emergence of azole resistance in Aspergillus fumigatus and spread of a single resistance mechanism. PLoS Med. 2008;5(11):e219. doi:10.1371/journal.pmed.0050219
71. Montesinos I, Argudín MA, Hites M, et al. Culture-based methods and molecular tools for azole-resistant Aspergillus fumigatus detection in a Belgian University Hospital. J Clin Microbiol. 2017;55(8):2391–2399. doi:10.1128/JCM.00520-17
72. Tsuchido Y, Tanaka M, Nakano S, Yamamoto M, Matsumura Y, Nagao M. Prospective multicenter surveillance of clinically isolated Aspergillus species revealed azole-resistant Aspergillus fumigatus isolates with TR34/L98H mutation in the Kyoto and Shiga regions of Japan. Med Mycol. 2019;57(8):997–1003. doi:10.1093/mmy/myz003
73. Rivelli Zea SM, Toyotome T. Azole-resistant Aspergillus fumigatus as an emerging worldwide pathogen. Microbiol Immunol. 2022;66(3):135–144. doi:10.1111/1348-0421.12957
74. Paul S, Stamnes M, Thomas GH, et al. AtrR is an essential determinant of azole resistance in aspergillus fumigatus. mBio. 2019;10(2):e02563–02518. doi:10.1128/mBio.02563-18
75. Paul S, Ror S, McDonald WH, Moye-Rowley WS. Biochemical identification of a nuclear coactivator protein required for AtrR-dependent gene regulation in aspergillus fumigatus. mSphere. 2022;7(6):e0047622. doi:10.1128/msphere.00476-22
76. Buied A, Moore CB, Denning DW, Bowyer P. High-level expression of cyp51B in azole-resistant clinical Aspergillus fumigatus isolates. J Antimicrob Chemother. 2013;68(3):512–514. doi:10.1093/jac/dks451
77. Gonzalez-Jimenez I, Lucio J, Amich J, et al. A Cyp51B mutation contributes to azole resistance in aspergillus fumigatus. J Fungi. 2020;6(4):315. doi:10.3390/jof6040315
78. Handelman M, Meir Z, Scott J, et al. Point mutation or overexpression of aspergillus fumigatus cyp51B, encoding lanosterol 14α-sterol demethylase, leads to triazole resistance. Antimicrob Agents Chemother. 2021;65(10):e0125221. doi:10.1128/AAC.01252-21
79. Fraczek MG, Bromley M, Buied A, et al. The cdr1B efflux transporter is associated with non-cyp51a-mediated itraconazole resistance in Aspergillus fumigatus. J Antimicrob Chemother. 2013;68(7):1486–1496. doi:10.1093/jac/dkt075
 © 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.