")
Back to Journals » Infection and Drug Resistance » Volume 16
Epidemic Trends and Biofilm Formation Mechanisms of Haemophilus influenzae: Insights into Clinical Implications and Prevention Strategies
Authors Xiao J, Su L, Huang S , Liu L , Ali K , Chen Z
Received 5 June 2023
Accepted for publication 10 August 2023
Published 16 August 2023 Volume 2023:16 Pages 5359—5373
DOI https://doi.org/10.2147/IDR.S424468
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor Suresh Antony
Jiying Xiao,1 Lin Su,2,3 Shumin Huang,2,3 Lingyue Liu,1 Kamran Ali,4 Zhimin Chen2,3
1Department of Pulmonology, Hangzhou Children’s Hospital, Hangzhou, Zhejiang, 310015, People’s Republic of China; 2Department of Pulmonology, Children’s Hospital, Zhejiang University School of Medicine, Hangzhou, Zhejiang, 310052, People’s Republic of China; 3National Clinical Research Center for Child Health, National Children’s Regional Medical Center, Hangzhou, Zhejiang, 310052, People’s Republic of China; 4Department of Oncology, The Fourth Affiliated Hospital, International Institutes of Medicine, Zhejiang University School of Medicine, Yiwu, Zhejiang, 322000, People’s Republic of China
Correspondence: Zhimin Chen, Department of Pulmonology, Children’s Hospital, Zhejiang University School of Medicine, 3333 Bingsheng Road, Hangzhou, Zhejiang, 310052, People’s Republic of China, Email [email protected]
Abstract: Haemophilus influenzae (H. influenzae) is a significant pathogen responsible for causing respiratory tract infections and invasive diseases, leading to a considerable disease burden. The Haemophilus influenzae type b (Hib) conjugate vaccine has notably decreased the incidence of severe infections caused by Hib strains, and other non-typable H. influenzae (NTHi) serotypes have emerged as epidemic strains worldwide. As a result, the global epidemic trends and antibiotic resistance characteristics of H. influenzae have been altered. Researches on the virulence factors of H. influenzae, particularly the mechanisms underlying biofilm formation, and the development of anti-biofilm strategies hold significant clinical value. This article provides a summary of the epidemic trends, typing methods, virulence factors, biofilm formation mechanisms, and prevention strategies of H. influenzae. The increasing prevalence of NTHi strains and antibiotic resistance among H. influenzae, especially the high β-lactamase positivity and the emergence of BLNAR strains have increased clinical difficulties. Understanding its virulence factors, especially the formation mechanism of biofilm, and formulating effective anti-biofilm strategies may help to reduce the clinical impact. Therefore, future research efforts should focus on developing new approaches to prevent and control H. influenzae infections.
Keywords: Haemophilus influenzae, epidemiology, virulence factors, biofilms, prevention strategies
Introduction
H. influenzae is an opportunistic pathogen that commonly colonizes the human nasopharynx and causes a variety of infections, such as acute sinusitis, otitis media, conjunctivitis, community-acquired pneumonia, bacterial vaginitis, and invasive diseases, including septic meningitis, septicemia, peritonitis, and arthritis, among others.1 Invasive diseases are becoming increasingly prevalent with each passing year. These conditions are marked by high mortality rates and have the potential to cause severe sequelae, posing a significant threat to the health of children.2 Based on the presence or absence of cell wall capsular polysaccharide components, H. influenzae can be classified into six encapsulated types (Hia-Hif) and non-encapsulated NTHi. Of these, Hib is the most virulent and is associated with invasive diseases. The introduction of the Hib conjugate vaccine in some countries and regions has led to a change in the epidemiological characteristics of H. influenzae, resulting in a significant decrease in severe infections associated with Hib.3 The public’s waning attention to H. influenzae has resulted in the serious underestimation of other serotypes, particularly NTHi-related infections. The escalating rate of antibiotic resistance, as well as the evolution of virulence factors, specifically the formation of biofilms, have presented significant challenges to clinical treatment. As a result, anti-biofilm strategies and preventative methods for H. influenzae have become prominent areas of research.
Epidemiology
The introduction of the Hib conjugate vaccine in Asia significantly trailed its implementation in developed Western countries. This lag, particularly in developing nations, led to considerable heterogeneity in H. influenzae epidemiology globally. Although Japan introduced the Hib vaccine in November 2008, the vaccination rate was initially low due to the voluntary nature of the program. Notably, it was not until December 2010, when the national policy to lessen the burden of self-pay vaccination was instituted, that the rate of vaccination increased significantly to over 90.0%.4 In China, the overall coverage of the Hib conjugate vaccine was 54.9%. The vaccine coverage was found to be lower among foreign populations as compared to local residents. Furthermore, the eastern region of China had a higher coverage as compared to the central and western regions.5 In Nepal, H. influenzae had been identified as the primary etiological agent of meningitis in pediatric patients.6 In Vietnam, before the nationwide introduction of Hib conjugate vaccine, a cross-sectional Hib carriage study among 1000 children < 5 years of age found that the carriage of H. influenzae was detected in 37.0% of children and the Hib carriage rate was 3.0%.7 The infection caused by H. influenzae remains a pertinent public health issue, particularly in emerging economies. As a result, it is advised to incorporate the Hib vaccine into national immunization schemes to address disparities in vaccination coverage.
As the incidence of Hib infection gradually decreased, the incidence of other serotypes and NTHi strains increased in relative terms. In North America, current epidemiological data suggested that invasive Hia disease predominantly affected indigenous communities, with Sequence Type (ST) 23 being the predominant clone described in most invasive diseases and MLST sites.8 The incidence rate of invasive Hia cases in the United States increased by 11.1% annually on average from 2008 to 2017, and the total mortality rate was 7.8% (30/386), 42.7% of Hia cases occurred in children under 5 years old.9 In Canada, H. influenzae diseases were predominantly caused by NTHi (74.2%, 950/1281), far exceeding serotype Hia (8.9%, 114/1281) and Hif (10.2%, 130/1281).10 In Portugal, 79.2% (206/260) of the invasive H. influenzae disease isolates were NTHi, and ST103 and ST57 were the major STs.11 The total carrying rate of H. influenzae among children aged 0–6 in daycare was 84.1% (1282/1524), of which 96.7% (1240/1282) were NTHi.12 A study in Shanghai, China found that all 51 isolates collected were NTHi, of which 5.9% (3/51) caused invasive infections, with ST103 (7.84%, 4/51) being the most common.13 A multicenter study in Zhejiang, China, found that H. influenzae infections accounted for 18.5% (280/1514) and NTHi accounted for 93.6% (262/280).14 NTHi can lead to severe invasive diseases, especially in the elderly, pregnant/postpartum women, and newborns. In the United States from 2008 to 2019, 92.0% (207/225) of newborns with invasive NTHi infections occurred within the first week of birth, and 72.4% (163/225) were born prematurely. Compared to full-term newborns, the risk of NTHi in premature infants was 23 times higher.15 In Belgium, the carrying rate of NTHi in children with acute otitis media and invasive diseases was 98.2% (214/218) and 68.1% (64/94) respectively.16 95.0% (190/200) of Japanese adults with invasive H. influenzae diseases were NTHi strains.17 NTHi has become a major subtype of H. influenzae infections worldwide and has the ability to cause invasive disease. Due to the collective global morbidity and mortality, it has been recognized by the World Health Organization as one of the 12 priority pathogens.18 With the introduction of Hib conjugate vaccine, the epidemiology of H. influenzae infection has changed, and NTHi has become a major subtype of lower respiratory tract infection and invasive disease, which requires great attention and focus. It is particularly important to carry out epidemiological investigations of H. influenzae in large samples and multicenter, which can provide an epidemiological basis for clinical diagnosis and treatment and national vaccine strategy.
H. influenzae Typing
There are currently multiple typing methods for H. influenzae, each with different focuses and advantages (Table 1). Choosing appropriate, convenient, and economical typing methods can provide scientific basis for clinical research and epidemiological investigations.
 |
Table 1 Characteristics of Different H. influenzae Typing Methods |
Capsule Typing
Slide Agglutination Serotyping (SAST)
Traditional serologic capsule typing was performed using a standard slide agglutination method. However, this traditional method’s accuracy is limited and some encapsulated strains may end up being misclassified as NTHi due to inaccuracies in performing and interpreting the slide agglutination test, or altered ability to produce capsule due to partial deletion of the capsule gene bexA.19
PCR Typing
The capsular polysaccharide gene loci has three functional regions I~III. Regions I and III are functional genes, encoded by bexDCBA and hcsAB respectively. They are highly conserved in all capsular types and are related to cell transport and post-translational processing. Region II genes encode capsular type a to f specific proteins, and there are differences in genes between different serotypes.20 The conventional typing method relies on the bexA gene and is not suitable for the study of a large number of isolates, which may lead to the misclassification of bexA deficient mutant strains. Some researchers described a molecular strategy that targeted the bexB gene, which provided a simple and reliable method for differentiating strains that lack the entire capsule locus from those containing a partial or complete capsule locus.21
Whole-Genome Sequencing (WGS) Typing
Due to the considerable accuracy provided by WGS, public health and clinical laboratories are currently beginning to incorporate WGS technology into diagnostic and disease surveillance projects.22 Recently, two in silico approaches using WGS, “hinflfluenzae_capsule_characterization”23 and “hicap”24 have provided a more robust and reliable method for H. influenzae capsule typing. The consistency between the two typing methods and the actual phenotype serum typing method is 99–100%, respectively,25 which are expected to replace the laborious traditional phenotype methods in clinical settings.
Matrix-Associated Laser Desorption Ionization-Time of Flight Mass Spectrometry (MALDI-TOF MS) Typing
Several studies have found that MALDI-TOF MS may be a simple and rapid alternative method for capsule serotyping of H. influenzae.26 It is also the first report that MALDI-TOF MS can distinguish between Hie and Hif. MALDI-TOF MS can also be used as a surveillance tool in vaccinated populations to monitor the ongoing effectiveness of Hib conjugate vaccines and as a preliminary screening method for invasive H. influenzae strains.
Biotyping
Biotypic analysis used internal medium for urease reaction and ornithine decarboxylase reaction. H. influenzae can be classified into eight different biological types (I–VIII), depending on the biochemical reactions to urease fermentation, indole tests, and ornithine decarboxylase fermentation. There may be some correlation between the biotype and the source of the disease and strain. Different studies have found that among all biotypes, biotype II is usually the most common.16,25 A survey found that biotype II was the most common in all groups, followed by type III found in the acute otitis media group and type I found in the invasive disease group.16 Type I may be related to invasive diseases.27
Multilocus Sequence Typing (MLST)
Pathogenic strains were identified by sequencing the internal fragments of seven housekeeping genes (adk, atpG, frdB, fucK, mdh, pgi, and recA).28 The typing results of isolates from different laboratories are comparable and can be stored, and the allele number, sequence type and corresponding clonal complexes (CC) of the strains are available in an Internet database. Through software, phylogenetic analysis can be conducted on different isolates to evaluate their phylogenetic relationships and diversity. Numerous studies have reported significant differences in MLST types among NTHi strains from different countries and regions. NTHi has multiple MLST types, indicating phylogenetic diversity among H. influenzae strains.29,30 The differences in molecular epidemiology and genetic evolution and the correlation between MLST clustering types and antibiotic resistance, are still controversial and require further research.
H. influenzae Virulence Factors
The virulence factors of H. influenzae include capsule, pili, adhesins, IgA proteases and so on, which mediate bacterial adhesion, colonization, invasion, and biofilm formation (Table 2), and have important clinical significance. Some virulence factors have become potential targets for new drug development and vaccine research.
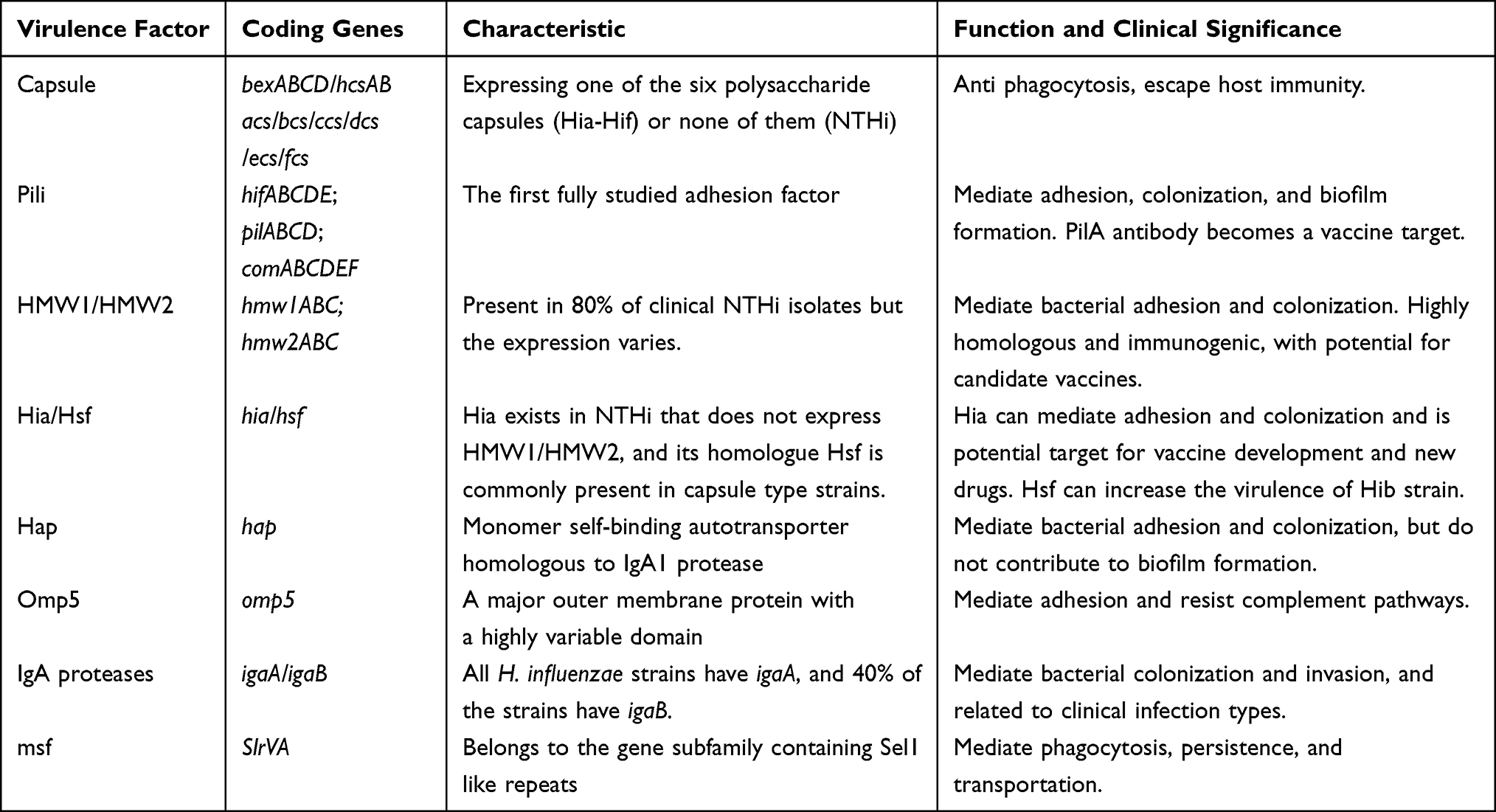 |
Table 2 Genes and Functions of Common Virulence Factors in H. influenzae |
Capsule
The capsule is a polymer composed of ribose and polyribosylribol phosphate (PRP), the amount of which is related to complement resistance.31 The capsule is encoded by the capsule gene in two identical copies of the chromosome. Amplification of the capsule gene can be seen in clinical isolates, which may be a mechanism for the bacteria to escape host defense. Bacteria containing 4 copies are significantly more resistant to antibody-dependent, classical complement pathway-directed phage cleavage compared to bacteria containing 2 copies.32 Bacterial capsules are composed of negatively charged polysaccharides that limit interactions with similarly negatively charged host cells, such as leukocyte phagocytosis.33
Pili
The pili is a polymeric spiral protein structure on the surface of H. influenzae, another important virulence factor, and the first fully studied adhesion factor. Type IV pili (T4P) plays an important role in bacterial colonization. T4P is a filamentous polymer that spreads across the surface of bacterial cells. It is crucial for bacterial adhesion and colonization in respiratory epithelial cells, twitching movements, and biofilm formation.34 The adhesion of NTHi expressing T4P is enhanced, and its receptor is intercellular adhesion molecule 1 (ICAM1) on the surface of epithelial cells35. The product of PilA gene in the coding gene pilABCD and comABCDEF gene cluster is the main fimbrial protein subunit.36 In vitro experiments, antibodies targeting pilA could prevent the formation of NTHi biofilms and damage the formed biofilms.37 Due to the importance of T4P in the colonization and pathogenesis, and the high conservation of the pilA subunit in different H. influenzae strains, pilA is being clinically studied as a candidate vaccine target for the prevention of NTHi disease.38
Adhesins
Adhesins are a large group of proteins including HWM1/HWM2, Hia/Hsf, Hap, and outer membrane proteins. The attachment of various adhesin proteins to host tissues is a critical step in the process of bacterial infection. In 80% of NTHi strains, bacterial attachment to the nasopharynx for colonization is mediated by two major groups of adhesion proteins, HMW1/HMW2.39 They can bind to different specific receptors in host cells and act at different stages of H. influenzae invasion, and their expression correlates with H. influenzae aggressiveness and virulence. Some researchers have identified the promoting effects of HMW1 and HMW2 adhesins on respiratory tract colonization through a monkey model, which may be related to stimulating immune responses.40 In patients with acute otitis media, HMW1 and HMW2 were the main targets of serum antibody response after infection, with high immunogenicity.41 The high homology, high immunogenicity, and important roles of adhesion and colonization of HMW1 and HMW2 have made these adhesins promising as vaccine candidates.41
In approximately 25% of NTHi strains that do not express HMW1/HMW2, the Hia protein can be found, a key factor in the initial colonization of the nasopharynx. Hia is the main non fimbriae adhesin of NTHi. Recent studies suggest that Hia expression may vary in stages to evade immune responses to NTHi, and variants with lower Hia levels may help the bacteria avoid being killed by anti-Hia antisera.42 Hia has dual binding activity, mimicking the combined activity of HMW1 and HMW2 adhesion proteins in function. Hia plays an important role in the biofilm formation and is expected to become a potential target for NTHi vaccine development and new drugs43.
Hsf is commonly found in capsular H. influenzae strains and was originally reported in Hib strains.44 Hsf is a unique twisted hairpin shaped trimer autologous transporter that is twice the size of Hia.45 Hib strain mainly obtains multifunctional glycoprotein vitronectin (Vn) through Hsf and inhibits the formation of membrane attack complex (MAC) to protect bacteria from serum-mediated killing. The fine-tuned protein-protein interaction between Hsf and Vn can help inhibiting MAC formation and lung epithelial cell invasion, which may contribute to increasing the toxicity of Hib.46
Hap is a monomeric self-binding autotransporter homologous to serine-type IgA1 protease that promotes interaction with respiratory epithelial cells.47 Hap is a widely expressed adhesion protein that binds to basement membrane components such as fibronectin, laminin and type IV collagen.48 The C-terminal adhesion region and N-terminal serine protease structure region of Hap can promote adhesion and regulate Hap self-protein hydrolysis, respectively.49
The P5 protein, encoded by the ompP5 gene, is a major outer membrane protein and is predicted to have a highly variable domain.50 P5 can mediate the binding of bacteria to respiratory epithelial cells and mucin, leading to increased complement resistance.49 P5 is a necessary condition for NTHi to resist classical and alternative complement pathways.51 C4b binding protein (C4BP) plays an important role in complement escape of NTHi, and the P5 protein of NTHi is a new ligand of C4BP. The expression of P5 is positively correlated with the binding of C4BP.52 However, the P5 protein does not seem to contribute to biofilm production.49
IgA Proteases
IgA proteases contribute to host mucosal colonization and tissue invasion and can cleave the heavy chain of the hinge region of human IgA1, disrupting IgA protection of the mucosal barrier. The H. influenzae genome has two alleles (igaA and igaB) encoding IgA protease, with all strains having igaA and 40% of strains having igaB.53 The expression of IgA protease varies among different NTHi strains. A study evaluated the relationship between IgA protease genotypes and clinical infection types and found that igaB was prevalent in 46% of respiratory tract infections, while it was relatively rare in invasive infections.54 The activity level of IgA1 protease isolated from symptomatic NTHi infected individuals was significantly higher than that of asymptomatic individuals, which determined that IgA1 protease was a virulence factor.55 Some researchers have discovered the first small molecule, non-peptide IgA1 protease inhibitor, which may be a potential treatment for resistant H. influenzae strains.56
Macrophage Survival Factor
Macrophage survival factor (msf) is a new virulence factor for phagocytosis, persistence, and trafficking to non-mucosal sites.57 Statistical genetic analyses found a significant association between possession of the SlrVA subfamily (that is msf) and the disease isolates. Msf-containing isolates were significantly higher in disease isolates than in carrier isolates, and they may play a role in disease chronicity as well as in invasive disease by providing an adaptive advantage in biofilms and increasing the survival of macrophages.
H. influenzae Biofilm
Biofilm is a microbial community with a certain spatial configuration that is attached to the surface of an object and consists of coated bacteria, lipopolysaccharide, matrix proteins, nucleic acids, and other components. Its formation is influenced by the factors of bacterial pathogens themselves, such as pili, liposaccharides, proteins, nucleic acids, quorum sensing system (QS) and two-component signaling system.58,59 Biofilms protect bacteria from environmental, host, and chemical stressors, and can enhance bacterial virulence and resistance. Biofilm formation is associated with recurrent respiratory diseases and asymptomatic colonization.60 More than 80% of human infectious diseases may be mediated by biofilms.61 H. influenzae can form biofilms in vivo and in vitro. A variety of clinical diseases and H. influenzae biofilms are closely related, such as chronic rhinosinusitis, secretory otitis media, adenoid hypertrophy, chronic obstructive pulmonary disease, cystic pulmonary fibrosis, and prolonged bacterial bronchitis. The in-depth study of H. influenzae biofilm can provide new treatment ideas for recurrent, chronic, and refractory infections.
Biofilm Classification
The formation of H. influenzae biofilms can be evaluated by modified microtitration plate method stained with crystal violet.62 However, there is currently no recognized classification standard for determining the ability of biofilm formation. Referring to some relevant literatures,63,64 classification is based on the calculated cut-off optical density (ODc). ODc is defined as the average optical density value of the blank control group plus three times the standard deviation. OD≤ODc indicates no biofilm formation, ODc<OD≤4ODc indicates weak biofilm formation, and OD>4ODc indicates strong biofilm formation.
Structure and Components
The process of biofilm formation can be divided into five stages: initial adhesion, proliferation, maturation, depolymerization and recolonization.65 In the adhesion stage, H. influenzae is irreversibly attached to respiratory epithelial cells through pili and adhesins, including binding ICAM-135 receptor and carcinoembryonic antigen-related cell adhesion molecules (CEACAMs).66 During the proliferation and maturation stages, bacteria can produce extracellular polymeric substances (EPS) and it mediates signal transmission, metabolic changes, gene expression changes, nutrition and oxygen gradient changes through the QS system, and finally forms a mature biofilm with mushroom like or tower like structure.67 When the internal environment of mature biofilm reaches saturation, bacteria can be depolymerized to become planktonic again and colonize other positions.
Biofilm formation is influenced by numerous autochthonous factors, such as T4P and adhesin proteins, as previously described. During NTHi biofilm formation in vitro, the relatively low average temperature of the human nasopharynx (34°C) significantly enhances T4P expression and motility compared to in vivo temperature (37°C).68 NTHI biofilms contain variants expressing lipooligosaccharide (LOS) that contain phosphorylcholine (PCho) and sialic acid. The expression of PCho is significantly correlated with an increase in biofilm thickness, surface coverage, total biomass, and a decrease in biofilm roughness.69 LOS containing sialic acid can promote NTHi to form biofilm in vitro and promote the persistence of bacteria in the middle ear or lung in vivo.70 However, some studies have found incorporation of phosphorylcholine into the LOS of NTHi does not correlate with the level of biofilm formation in vitro,71 so further in-depth research is needed.
QS is a density-dependent coordination system of gene expression among bacteria in a community, which is essential for the formation, development, and diffusion of biofilms. Most biofilms achieve QS through the Auto-Inducer 2 (AI-2) pathway controlled by the luxS manipulator.72 These signals are detected by the ABC translocator protein RbsB, leading to a downstream transcriptional cascade response that drives biofilm formation.73 The expression of AI-2 can promote the formation of NTHi biofilm and the persistence of bacteria in vivo.74,75 A study used NTHi mutant experiments to confirm that activation and interruption of the luxS operon in the AI-2 pathway can affect the maturation and diffusion of biofilms, respectively.76 Induction of luxS synthesis can lead to increased transcription of glycosyltransferase. Glycosyltransferase is a key enzyme involved in the production of extracellular matrix in bacterial biofilms formed by streptococci found in the lungs of COPD patients.77 NTHi also expresses a QSeB/C two-component secondary system independent of the AI-2 system, which may support the coordination of biofilm gene expression when the luxS-RbsB system is damaged.58
Extracellular DNA (eDNA) and DNABII proteins are important components of NTHi biofilm EPS. Immunofluorescence techniques have shown that eDNA provides structural stability, maintenance, and extension of the biofilm to secondary sites.78 In the chinchilla otitis media model, eDNA can reduce the biological activity of an important effector of innate immunity, namely Human β-Defensin-3 (hBD-3).79 The degradation of eDNA can destabilize the biofilm of NTHi and also reduce surface adhesion and biofilm formation of bacteria in the planktonic state.80 The integration host factor (IHF) and histone-like protein HU in the DNABII protein family, located in different regions of the extracellular matrix of biofilm, have independent functions and play a central role in the structural integrity of NTHi biofilms.81 In the chinchilla model, targeted immunization with these proteins can lead to rapid removal of biofilms.81 NTHi T4P can transport eDNA and DNABII from bacterial cytoplasm to periplasm through the expression of ComE pore, which suggests a novel form of DNA release from viable NTHi.82 Monoclonal antibodies targeting DNABII protein can significantly damage NTHi biofilms.83
Biofilms and Antibiotic Resistance
NTHi biofilms exhibit a complex multidrug resistance strategy to a variety of widely used antibiotics. Studies have shown that ciprofloxacin, azithromycin and amoxicillin are 100% effective against NTHi planktonic strains, while killing only 68%, 57% and 4% of NTHi strains in biofilms.84 Sub-inhibitory concentrations of β-lactam antibiotic stimulated the biofilm-forming ability of NTHi strains, genes involved in glycogen production and transporter function were up-regulated and down-regulated genes were linked to multiple metabolic processes but not those involved in stress response.85 The present proteomic study indicates that the NTHi biofilm exists in a semi-dormant state with decreased energy metabolism and protein synthesis yet is still capable of managing oxidative stress.86 The eDNA-rich extracellular matrix can impede the penetration of a range of antimicrobials, including ampicillin and ciprofloxacin.80 Fluctuating gene expression and cellular metabolism within biofilms result in a drug-resistant bacterial phenotype. Planktonic H. influenzae has been shown to exhibit a heterogeneous drug-resistant phenotype to imipenem,87 and this resistance can be enhanced in biofilm communities.88 Differences in biofilm formation depend on the type of diseases, and there is a high degree of heterogeneity among biofilms produced by H. influenzae clinical isolates,89 but there does not appear to be a significant correlation between biofilm thickness and resistance to antibiotics.90
Biofilms and the Immune System
Biofilms have important host components, including neutrophil extracellular traps (NETs). However, the formation of NETs is not an important determinant of NTHi clearance, but NTHi communities with biofilm phenotypes can survive in NETs.91 The study found that peroxidase and peroxiredoxin-glutaredoxin (pdgx) are the decisive factors to promote the survival and persistence of bacteria in NETs.92 Biofilms also induce metabolically inactive persistent cells and provide physical and chemical barriers to immune system effectors and antimicrobials. Biofilms can produce IgA proteases that integrate into the extracellular matrix and block the host immune response by cleaving human IgA. IgA proteases are differentially expressed in vivo but play an important role for bacterial survival in respiratory epithelial cells.93 The eDNA of H. influenzae biofilms can also bind to hBD-3, reducing its antimicrobial properties.79 HBD3 is a member of the innate immune system and is essential in the protection of respiratory epithelial cells from bacterial invasion.94
Polymicrobial Biofilms
Biofilms are rarely formed by a single species, and the clinical significance of polymicrobial biofilms has been increasingly recognized. In polymicrobial biofilms, pathogens continuously communicate with each other through direct contact or the release and uptake of QS molecules.95 NTHi and Streptococcus pneumoniae often colonize the respiratory tract of COPD patients together and interact synergistically to promote initial attachment, biofilm formation and survival.96 The mechanism of interaction between these two bacteria has been previously studied. NTHi can provide passive protection for Streptococcus pneumoniae in vivo through two distinct mechanisms: production of β-lactamase and formation of biofilm communities.97 NTHi and Streptococcus pneumoniae can mutually regulate the expression of virulence genes. The lytA and cbpD gene expression, which regulate autolysis and self-cleavage in Streptococcus pneumoniae, were significantly downregulated98 and the expression of T4P structural protein in NTHi was significantly upregulated.99 However, another study claimed that NTHi could coexist with Streptococcus pneumoniae during the first 24 hours, but after 48 hours, the viability of NTHi rapidly decreased to undetectable levels, when moraxella catarrhalis was present in the polymicrobial biofilm NTHi could survive for 48 hours.100
Biofilms and Clinical Correlation
The study on the correlation between biofilm formation ability and clinical characteristics has clinical value. A study of NTHi of lower respiratory tract origin found biofilm formation in 10.2% (10/98) of isolates, with no significant differences in antimicrobial susceptibility patterns and severity of clinical symptoms between the biofilm-forming and non-forming groups, but correlated with ICU length of stay, demonstrating for the first time the clinical impact of NTHi biofilm production.101 Another study found that the biofilm index was higher in the invasive disease group compared to the non-invasive group, but the difference was not statistically significant.102 NTHi isolated from children with fever or fever with earache could produce biofilms more frequently than children without these symptoms.103 The production of biofilms was not significantly associated with treatment failure or recurrence of acute otitis media, but the production of biofilms was significantly lower in conjunctivitis-otitis media syndrome and H. influenzae strains modified with penicillin-binding proteins.62
Anti-Biofilm Strategy
Biofilms have strong drug resistance and immune defense effects and are closely related to clinical severe and chronic infections. The urgent need to develop effective anti-biological strategies will be an important direction for future research. Physical clearance methods such as surgical debridement, ultrasound, and electrical current clearance are not suitable for respiratory mucosa. Biological clearance methods such as bacteriophages have demonstrated anti biofilm activity against various bacteria such as Pseudomonas aeruginosa and Staphylococcus aureus in vitro and in vivo experiments.104,105 However, there are few studies on H. influenzae. Bacteriophages are highly heterogeneous and have potential biosafety risks, so strict in vivo research is still needed. Chemical clearance methods such as antibacterial drugs, antibodies, enzymes, surfactants, and some QS inhibitors are currently the mainstream direction of applied research. In vitro models and animal experiments, relevant research on antibodies has achieved phased results, which will be described in detail in the subsequent chapter on vaccine prevention. Some natural compounds, such as plant essential oils, Chinese herbal extracts, and synthetic chemicals can inhibit and destroy biofilms by inhibiting the QS system, like Oldenlandia diffusa extract,106 plant essential oils,107–109 3-hydroxychalcone (chalcone 8)110 and so on. However, due to the possible cytotoxicity and complex synthesis and extraction processes, in vivo research and even clinical trials still require further in-depth research. In addition, serum proteases have proteolytic activity against bacterial adhesions involved in biofilm formation.111 Antibacterial photodynamic therapy (aPDT) of photosensitizer Chlorin e6 (Ce6) has significant bactericidal activity against NTHi in planktonic and biofilm states, and is an effective and promising adjuvant treatment for acute and recurrent otitis media.112,113
In clinical practice, rational selection of antibiotics or combined use of certain drugs is more clinically valuable. Studies have found that high concentrations of amoxicillin cannot remove NTHi from biofilms, while respiratory quinolones can inhibit biofilm formation and eradicate mature biofilms and can kill NTHi in biofilms even under sub-MIC conditions.114 Simultaneous administration of carbapenem, neoquinolone and macrolide has the effect of anti NTHi mature biofilm.115 The combination of metal chelating agent EDTA and antibiotics can inhibit biofilm formation and damage mature biofilms, and increase the sensitivity of NTHi biofilms to ampicillin and ciprofloxacin.80
Antibiotic Resistance
The resistance rate of H. influenzae to common antibiotics, particularly β-lactam antibiotics, has been continuously increasing. The resistance mechanism of H. influenzae includes the production of β lactamase (encoded by TEM−1 and ROB−1 resistance genes)116 and the decrease of PBP3 affinity caused by mutations in ftsI genes (encoded by PBP3S and PBP3BLN resistance genes).117 The mechanism of the β-lactamase-negative ampicillin resistant (BLNAR) strain is the alteration of the PBP3 protein encoded by the ftsI gene. In addition, strains with both mechanisms are referred to as β-lactamase-positive amoxicillin-clavulanate resistant (BLPACR).118 The β-lactamase positivity and BLNAR strains varied in different countries and regions. In Shanghai, China, 33.3% (17/51) of the strains produced TEM-1-type β-lactamase, and BLNAR strains accounted for 11.8% (6/51),119 while the proportion of BLNAR strains in Guangzhou, China in 2016–2017 was 22.1% (89/402).120 In Portugal, the β-lactamase positivity rate was 7.5% (96/1282).12 In contrast, in France, the rate of β-lactamase positivity was as high as 27.1% (157/580) and 35.0% (208/595) for the BLNAR strains.121 In Belgium, β-lactamase positivity was 23.8% (132/554) and 16.0% (15/94) in the carrier and invasive groups, respectively.16
In addition to β-lactam antibiotics, fluoroquinolones are also commonly used for respiratory infections. Fluoroquinolone-resistant H. influenzae was first reported in 1993, and child fluoroquinolone-resistant H. influenzae was first reported in Hong Kong in 2004.122 Subsequently, fluoroquinolone-resistant H. influenzae increased gradually, with different prevalence trends worldwide. No fluoroquinolone-resistant strains were found in invasive H. influenzae isolates collected in Canada between 2007 and 2014.123 In Spain, 0.4% (28/7267) strains were resistant to fluoroquinolone.124 2.7% (11/402) ciprofloxacin-resistant H. influenzae were detected in Guangzhou, China.120 A study in Taiwan found that 41.7% (20/48) of H. influenzae isolated from nursing home residents were not sensitive to fluoroquinolones, and the reduced sensitivity to fluoroquinolones was associated with a gyrA gene mutation at site 84.125 The drug resistance situation of H. influenzae is very serious. The in-depth investigation of large samples and multi-centers to grasp the mechanism of drug resistance and the changing situation is of great significance for the scientific and rational selection of antibiotics in the clinic.
H. influenzae Vaccine Prevention
Prevention against H. influenzae is mainly based on vaccines. In China, the main vaccine is Hib polysaccharide conjugate vaccine, including single Hib vaccine, DTaP-Hib (quadruple vaccine) and DTap-Hib-IPV (pentavalent vaccine). In addition, in Western countries, there is the DTap-Hepatitis B-Inactivated Polio-Hib (hexavalent vaccine). But there is no specific vaccine for NTHi. Several studies on potential vaccine targets for NTHi have achieved certain results.
The pneumococcal NTHi protein D conjugate vaccine (PHiD-CV) had a vaccine protective effect of 35.3% against NTHi-associated otitis media, suggesting for the first time that a vaccine strategy targeting NTHi is possible.126 The vaccine efficacy of PHiD-CV in treating clinical acute otitis media and bacterial otitis media in children<24 months was 24.0% and 48.0%, respectively, and the efficacy in treating moderate and severe clinical otitis media was 17.7% and 32.7%, respectively.127 Researches into the development of vaccine targets in vitro and vivo primarily focused on H. influenzae adhesion proteins, particularly those that are critical in biofilm formation and stabilization. The anti-recombinant soluble form of PilA (rsPilA) antibody could destroy and prevent the formation of NTHi biofilms in vitro, and could also prevent experimental otitis media caused by NTHi in animal models.37 A chimeric immunogen, “chimV4”, against both the P5 and T4P, could provide significant protection against otitis media caused by NTHi in chinchilla models by percutaneous immunization.128 Recombinantly expressed protein D and outer membrane protein 26 could induce a robust antibody response after vaccination as a separate vaccine in a mouse model.129 The fusion protein of E protein and PilA also expressed certain vaccine potential against NTHi biofilm in vitro mouse models.130 The cationic cholesterol pullulan based (cCHP) - P6 nasal vaccine in the mouse model could also induce effective protective effect in the airway mucosa.131 The mixture of anti IHF and amoxicillin clavulanate completely inhibited the formation of NTHi biofilms.132 Australian Aboriginal otitis-prone children had lower serum IgG titres to major NTHi vaccine candidate antigens (rsPilA, ChimV4 and outer membrane protein 26), suggesting that Aboriginal children may benefit from immunisation with vaccines containing these antigens to increase titres of protective antibodies.133 There are also other NTHi surface dominant antigens that can be used as candidate vaccine molecules. However, unlike capsular type H. influenzae, NTHi lacks a single surface antigen and may require a multi component binding targeted vaccine. How to combine candidate vaccine antigen molecules will be a research hotspot in the future.
Discussion and Future Work
H. influenzae is an important pathogen that causes respiratory tract infections and invasive diseases, posing a serious threat to children’s health and remaining a public health issue that requires attention at present. With the introduction of the Hib conjugate vaccine, there has been a significant reduction in severe Hib-related infections. However, the infection situation in developing countries with low conjugate vaccine coverage is still severe, leading to a serious disease burden. It is recommended to incorporate the Hib vaccine into national immunization planning as soon as possible.
With the decline in the Hib incidence rate, other serotypes, especially the NTHi subtype, have gradually become epidemic strains and have led to severe infections. Due to the global incidence rate and mortality, NTHi has been recognized as one of the 12 priority pathogens by the WHO. Additionally, the drug resistance situation of H. influenzae is severe, and the high β-lactamase positivity and the emergence of BLNAR strains have made clinical treatment more difficult. There are significant regional differences in the detection rate of β-lactamase, drug resistance genes, and drug resistance patterns. Timely regional and multicenter epidemiological surveys are necessary to understand the epidemic trend of H. influenzae strains and provide epidemiological evidence for clinical treatment and national vaccine strategies.
Virulence factors, such as the capsule, fimbriae, adhesion proteins, and IgA protease, play an important role in the adhesion, colonization, invasion, and biofilm formation of H. influenzae. In-depth research on these factors provides ideas for clinical treatment and prevention strategies. Some antibodies like PilA antibody,37 HMW1 and HMW2 protein antibodies,41 Outer membrane protein P5128, P6131, IgA1 protease inhibitor56 have shown promising results in vitro experiments and are expected to become candidate vaccine targets. Bacterial biofilm is a complex spatial structure that can protect bacteria from environmental, host, and chemical stress sources, and can enhance bacterial virulence and drug resistance. Its complex regulatory mechanisms, strong drug resistance, and correlation with clinical chronic infections have been research hotspots in recent years. The ultimate goal is to actively develop effective and clinically translatable anti-biofilm strategies. Drugs targeting bacterial biofilm constituent proteins and QS systems may be new research directions and approaches. Fortunately, some protein antibodies targeting biofilm formation and stability have achieved certain results in vitro and in vivo experiments. We hope to see more scientific clinical research results, ultimately benefiting the public.
In summary, the increasing prevalence of NTHi strains and antibiotic resistance among H. influenzae has increased clinical difficulties. H. influenzae infection is still a public health problem that can cause serious disease burden, although it has been underestimated by the public. Future studies should concentrate on the epidemic trends and drug resistance characteristics of H. influenzae in different regions, the mechanisms underlying biofilm formation, and the development of anti-biofilm strategies. These efforts will provide a scientific basis for the clinical diagnosis, treatment, and prevention of H. influenzae infections.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This research received no external funding.
Disclosure
The authors declare no potential conflicts of interest in this work.
References
1. Butler DF, Myers AL. Changing epidemiology of Haemophilus influenzae in children. Infect Dis Clin North Am. 2018;32(1):119–128. doi:10.1016/j.idc.2017.10.005
2. Soeters HM, Blain A, Pondo T, et al. Current epidemiology and trends in invasive Haemophilus influenzae disease-United States, 2009–2015. Clin Infect Dis. 2018;67(6):881–889. doi:10.1093/cid/ciy187
3. Wahl B, O’Brien KL, Greenbaum A, et al. Burden of Streptococcus pneumoniae and Haemophilus influenzae type b disease in children in the era of conjugate vaccines: global, regional, and national estimates for 2000–15. Lancet Glob Health. 2018;6(7):e744–e757. doi:10.1016/S2214-109X(18)30247-X
4. Sakata H, Adachi Y, Morozumi M, Ubukata K. Invasive Haemophilus influenzae infections in children in Kamikawa subprefecture, Hokkaido, Japan, 2006–2015: the effectiveness of H. influenzae type b vaccine. J Infect Chemother. 2017;23(7):459–462. doi:10.1016/j.jiac.2017.03.019
5. Yang Y, Yang Y, Scherpbier RW, et al. Coverage of Haemophilus influenzae type b conjugate vaccine for children in Mainland China: systematic review and meta-analysis. Pediatr Infect Dis J. 2019;38(3):248–252. doi:10.1097/INF.0000000000002132
6. Shrestha RG, Tandukar S, Ansari S, et al. Bacterial meningitis in children under 15 years of age in Nepal. BMC Pediatr. 2015;15:94. doi:10.1186/s12887-015-0416-6
7. Yoshida LM, Nguyen HA, Watanabe K, et al. Incidence of radiologically-confirmed pneumonia and Haemophilus influenzae type b carriage before Haemophilus influenzae type b conjugate vaccine introduction in Central Vietnam. J Pediatr. 2013;163(1 Suppl):S38–S43. doi:10.1016/j.jpeds.2013.03.029
8. Tsang RSW, Ulanova M. The changing epidemiology of invasive Haemophilus influenzae disease: emergence and global presence of serotype a strains that may require a new vaccine for control. Vaccine. 2017;35(33):4270–4275. doi:10.1016/j.vaccine.2017.06.001
9. Soeters HM, Oliver SE, Plumb ID, et al. Epidemiology of invasive Haemophilus influenzae serotype a disease-United States, 2008–2017. Clin Infect Dis. 2021;73(2):e371–e379. doi:10.1093/cid/ciaa875
10. McTaggart LR, Cronin K, Seo CY, Wilson S, Patel SN, Kus JV. Increased incidence of invasive Haemophilus influenzae disease driven by non-type B isolates in Ontario, Canada, 2014 to 2018. Microbiol Spectr. 2021;9(2):e0080321. doi:10.1128/Spectrum.00803-21
11. Heliodoro CIM, Bettencourt CR, Bajanca-Lavado MP; Portuguese Group for the Study of Haemophilus influenzae invasive i. Molecular epidemiology of invasive Haemophilus influenzae disease in Portugal: an update of the post-vaccine period, 2011–2018. Eur J Clin Microbiol Infect Dis. 2020;39(8):1471–1480. doi:10.1007/s10096-020-03865-0
12. Bajanca-Lavado MP, Cavaco L, Fernandes M, et al. Haemophilus influenzae carriage among healthy children in Portugal, 2015–2019. Microorganisms. 2022;10. doi:10.3390/microorganisms10101964
13. Li XX, Xiao SZ, Gu FF, He WP, Ni YX, Han LZ. Molecular epidemiology and antimicrobial resistance of Haemophilus influenzae in adult patients in Shanghai, China. Front Public Health. 2020;8:95. doi:10.3389/fpubh.2020.00095
14. Fan X, Liu X, Ji L, et al. Epidemiological analysis and rapid detection by one-step multiplex PCR assay of Haemophilus influenzae in children with respiratory tract infections in Zhejiang Province, China. BMC Infect Dis. 2018;18(1):414. doi:10.1186/s12879-018-3295-2
15. Oliver SE, Rubis AB, Soeters HM, et al. Epidemiology of invasive nontypeable Haemophilus influenzae disease-United States, 2008–2019. Clin Infect Dis. 2023;76(11):1889–1895. doi:10.1093/cid/ciad054
16. Ekinci E, Willen L, Rodriguez Ruiz JP, et al. Haemophilus influenzae carriage and antibiotic resistance profile in Belgian infants over a three-year period (2016–2018). Front Microbiol. 2023;14:1160073. doi:10.3389/fmicb.2023.1160073
17. Hachisu Y, Tamura K, Murakami K, et al. Invasive Haemophilus influenzae disease among adults in Japan during 2014–2018. Infection. 2023;51(2):355–364. doi:10.1007/s15010-022-01885-w
18. Bakaletz LO, Novotny LA. Nontypeable Haemophilus influenzae (NTHi). Trends Microbiol. 2018;26(8):727–728. doi:10.1016/j.tim.2018.05.001
19. Cerquetti M, Cardines R, Giufre M, et al. Genetic diversity of invasive strains of Haemophilus influenzae type b before and after introduction of the conjugate vaccine in Italy. Clin Infect Dis. 2006;43(3):317–319. doi:10.1086/505499
20. Lam TT, Elias J, Frosch M, Vogel U, Claus H. New diagnostic PCR for Haemophilus influenzae serotype e based on the cap locus of strain ATCC 8142. Int J Med Microbiol. 2011;301(2):176–179. doi:10.1016/j.ijmm.2010.07.004
21. Davis GS, Sandstedt SA, Patel M, Marrs CF, Gilsdorf JR. Use of bexB to detect the capsule locus in Haemophilus influenzae. J Clin Microbiol. 2011;49(7):2594–2601. doi:10.1128/JCM.02509-10
22. Revez J, Espinosa L, Albiger B, et al. Survey on the use of whole-genome sequencing for infectious diseases surveillance: rapid expansion of european national capacities, 2015–2016. Front Public Health. 2017;5:347. doi:10.3389/fpubh.2017.00347
23. Potts CC, Topaz N, Rodriguez-Rivera LD, et al. Genomic characterization of Haemophilus influenzae: a focus on the capsule locus. BMC Genomics. 2019;20(1):733. doi:10.1186/s12864-019-6145-8
24. Watts SC, Holt KE. hicap: in silico serotyping of the Haemophilus influenzae capsule locus. J Clin Microbiol. 2019;57(6). doi:10.1128/jcm.00190-19
25. Slotved HC, Johannesen TB, Stegger M, Fuursted K. Evaluation of molecular typing for national surveillance of invasive clinical Haemophilus influenzae isolates from Denmark. Front Microbiol. 2022;13:1030242. doi:10.3389/fmicb.2022.1030242
26. Takeuchi N, Segawa S, Ishiwada N, et al. Capsular serotyping of Haemophilus influenzae by using matrix-associated laser desorption ionization-time of flight mass spectrometry. J Infect Chemother. 2018;24(7):510–514. doi:10.1016/j.jiac.2018.02.007
27. Klibanov OM, Kehr H, Jeter Z, Ekwonu T. Fatal meningitis and sepsis caused by nontypeable Haemophilus influenzae. J Med Cases. 2022;13(8):396–401. doi:10.14740/jmc3974
28. Meats E, Feil EJ, Stringer S, et al. Characterization of encapsulated and noncapsulated Haemophilus influenzae and determination of phylogenetic relationships by multilocus sequence typing. J Clin Microbiol. 2003;41(4):1623–1636. doi:10.1128/JCM.41.4.1623-1636.2003
29. Wen S, Mai Y, Chen X, et al. Molecular epidemiology and antibiotic resistance analysis of Non-Typeable Haemophilus influenzae (NTHi) in Guangzhou: a representative city of Southern China. Antibiotics. 2023;12(4):656.
30. Dong Q, Shi W, Cheng X, et al. Widespread of non-typeable Haemophilus influenzae with high genetic diversity after two decades use of Hib vaccine in China. J Clin Lab Anal. 2020;34(4):e23145. doi:10.1002/jcla.23145
31. Riesbeck K. Complement evasion by the human respiratory tract pathogens Haemophilus influenzae and Moraxella catarrhalis. FEBS Lett. 2020;594(16):2586–2597. doi:10.1002/1873-3468.13758
32. Noel GJ, Brittingham A, Granato AA, Mosser DM. Effect of amplification of the Cap b locus on complement-mediated bacteriolysis and opsonization of type b Haemophilus influenzae. Infect Immun. 1996;64(11):4769–4775. doi:10.1128/iai.64.11.4769-4775.1996
33. Hauser S, Wegele C, Stump-Guthier C, et al. Capsule and fimbriae modulate the invasion of Haemophilus influenzae in a human blood-cerebrospinal fluid barrier model. Int J Med Microbiol. 2018;308(7):829–839. doi:10.1016/j.ijmm.2018.07.004
34. Jurcisek JA, Bookwalter JE, Baker BD, et al. The PilA protein of non-typeable Haemophilus influenzae plays a role in biofilm formation, adherence to epithelial cells and colonization of the mammalian upper respiratory tract. Mol Microbiol. 2007;65(5):1288–1299. doi:10.1111/j.1365-2958.2007.05864.x
35. Novotny LA, Bakaletz LO. Intercellular adhesion molecule 1 serves as a primary cognate receptor for the Type IV pilus of nontypeable Haemophilus influenzae. Cell Microbiol. 2016;18(8):1043–1055. doi:10.1111/cmi.12575
36. Carruthers MD, Tracy EN, Dickson AC, Ganser KB, Munson RS, Bakaletz LO. Biological roles of nontypeable Haemophilus influenzae type IV pilus proteins encoded by the pil and com operons. J Bacteriol. 2012;194(8):1927–1933. doi:10.1128/JB.06540-11
37. Mokrzan EM, Novotny LA, Brockman KL, Bakaletz LO. Antibodies against the Majority Subunit (PilA) of the type IV pilus of nontypeable Haemophilus influenzae disperse moraxella catarrhalis from a dual-species biofilm. mBio. 2018;9(6):e02423–18.
38. Wilkinson TMA, Schembri S, Brightling C, et al. Non-typeable Haemophilus influenzae protein vaccine in adults with COPD: a Phase 2 clinical trial. Vaccine. 2019;37(41):6102–6111. doi:10.1016/j.vaccine.2019.07.100
39. Buscher AZ, Burmeister K, Barenkamp SJ, St Geme JW. Evolutionary and functional relationships among the nontypeable Haemophilus influenzae HMW family of adhesins. J Bacteriol. 2004;186(13):4209–4217. doi:10.1128/JB.186.13.4209-4217.2004
40. Rempe KA, Porsch EA, Wilson JM, St Geme JW. The HMW1 and HMW2 adhesins enhance the ability of nontypeable Haemophilus influenzae to colonize the upper respiratory tract of rhesus macaques. Infect Immun. 2016;84(10):2771–2778. doi:10.1128/IAI.00153-16
41. Kadry NA, Porsch EA, Shen H, St Geme JW. Immunization with HMW1 and HMW2 adhesins protects against colonization by heterologous strains of nontypeable Haemophilus influenzae. Proc Natl Acad Sci U S A. 2021;118(32):e2019923118.
42. Atack JM, Winter LE, Jurcisek JA, Bakaletz LO, Barenkamp SJ, Jennings MP. Selection and counterselection of hia expression reveals a key role for phase-variable expression of hia in infection caused by nontypeable Haemophilus influenzae. J Infect Dis. 2015;212(4):645–653. doi:10.1093/infdis/jiv103
43. Atack JM, Day CJ, Poole J, et al. The nontypeable Haemophilus influenzae major adhesin hia is a dual-function lectin that binds to human-specific respiratory tract sialic acid glycan receptors. mBio. 2020;11(6):10–128.
44. Watson ME, Nelson KL, Nguyen V, et al. Adhesin genes and serum resistance in Haemophilus influenzae type f isolates. J Med Microbiol. 2013;62(Pt 4):514–524. doi:10.1099/jmm.0.052175-0
45. Singh B, Jubair TA, Morgelin M, et al. Haemophilus influenzae surface fibril (Hsf) is a unique twisted hairpin-like trimeric autotransporter. Int J Med Microbiol. 2015;305(1):27–37. doi:10.1016/j.ijmm.2014.10.004
46. Singh B, Su YC, Al-Jubair T, et al. A fine-tuned interaction between trimeric autotransporter haemophilus surface fibrils and vitronectin leads to serum resistance and adherence to respiratory epithelial cells. Infect Immun. 2014;82(6):2378–2389. doi:10.1128/IAI.01636-13
47. Spahich NA, St Geme JW. Structure and function of the Haemophilus influenzae autotransporters. Front Cell Infect Microbiol. 2011;1:5. doi:10.3389/fcimb.2011.00005
48. De Chiara M, Hood D, Muzzi A, et al. Genome sequencing of disease and carriage isolates of nontypeable Haemophilus influenzae identifies discrete population structure. Proc Natl Acad Sci U S A. 2014;111(14):5439–5444. doi:10.1073/pnas.1403353111
49. Euba B, Moleres J, Viadas C, et al. Relative contribution of P5 and Hap surface proteins to nontypable Haemophilus influenzae interplay with the host upper and lower airways. PLoS One. 2015;10(4):e0123154. doi:10.1371/journal.pone.0123154
50. Marti-Lliteras P, Lopez-Gomez A, Mauro S, et al. Nontypable Haemophilus influenzae displays a prevalent surface structure molecular pattern in clinical isolates. PLoS One. 2011;6(6):e21133. doi:10.1371/journal.pone.0021133
51. Rosadini CV, Ram S, Akerley BJ. Outer membrane protein P5 is required for resistance of nontypeable Haemophilus influenzae to both the classical and alternative complement pathways. Infect Immun. 2014;82(2):640–649. doi:10.1128/IAI.01224-13
52. Thofte O, Bettoni S, Su YC, et al. Nontypeable Haemophilus influenzae P5 binds human C4b-binding protein, promoting serum resistance. J Immunol. 2021;207(6):1566–1577. doi:10.4049/jimmunol.2100105
53. Murphy TF, Kirkham C, Jones MM, Sethi S, Kong Y, Pettigrew MM. Expression of IgA proteases by Haemophilus influenzae in the respiratory tract of adults with chronic obstructive pulmonary disease. J Infect Dis. 2015;212(11):1798–1805. doi:10.1093/infdis/jiv299
54. Resman F, Manat G, Lindh V, Murphy TF, Riesbeck K. Differential distribution of IgA-protease genotypes in mucosal and invasive isolates of Haemophilus influenzae in Sweden. BMC Infect Dis. 2018;18(1):592. doi:10.1186/s12879-018-3464-3
55. Vitovski S, Dunkin KT, Howard AJ, Sayers JR. Nontypeable Haemophilus influenzae in carriage and disease: a difference in IgA1 protease activity levels. JAMA. 2002;287(13):1699–1705. doi:10.1001/jama.287.13.1699
56. Shehaj L, Choudary SK, Makwana KM, Gallo MC, Murphy TF, Kritzer JA. Small-molecule inhibitors of Haemophilus influenzae IgA1 protease. ACS Infect Dis. 2019;5(7):1129–1138. doi:10.1021/acsinfecdis.9b00004
57. Kress-Bennett JM, Hiller NL, Eutsey RA, et al. Identification and characterization of msf, a novel virulence factor in Haemophilus influenzae. PLoS One. 2016;11(3):e0149891. doi:10.1371/journal.pone.0149891
58. Unal CM, Singh B, Fleury C, et al. QseC controls biofilm formation of non-typeable Haemophilus influenzae in addition to an AI-2-dependent mechanism. Int J Med Microbiol. 2012;302(6):261–269. doi:10.1016/j.ijmm.2012.07.013
59. Vogel AR, Szelestey BR, Raffel FK, et al. SapF-mediated heme-iron utilization enhances persistence and coordinates biofilm architecture of Haemophilus. Front Cell Infect Microbiol. 2012;2:42. doi:10.3389/fcimb.2012.00042
60. Boisvert AA, Cheng MP, Sheppard DC, Nguyen D. Microbial biofilms in pulmonary and critical care diseases. Ann Am Thorac Soc. 2016;13(9):1615–1623. doi:10.1513/AnnalsATS.201603-194FR
61. Sun F, Qu F, Ling Y, et al. Biofilm-associated infections: antibiotic resistance and novel therapeutic strategies. Future Microbiol. 2013;8(7):877–886. doi:10.2217/fmb.13.58
62. Mizrahi A, Cohen R, Varon E, et al. Non typable-Haemophilus influenzae biofilm formation and acute otitis media. BMC Infect Dis. 2014;14:400. doi:10.1186/1471-2334-14-400
63. Cardines R, Giufre M, Pompilio A, et al. Haemophilus influenzae in children with cystic fibrosis: antimicrobial susceptibility, molecular epidemiology, distribution of adhesins and biofilm formation. Int J Med Microbiol. 2012;302(1):45–52. doi:10.1016/j.ijmm.2011.08.003
64. Hassan A, Usman J, Kaleem F, Omair M, Khalid A, Iqbal M. Evaluation of different detection methods of biofilm formation in the clinical isolates. Braz J Infect Dis. 2011;15(4):305–311.
65. Renner LD, Weibel DB. Physicochemical regulation of biofilm formation. MRS Bull. 2011;36(5):347–355. doi:10.1557/mrs.2011.65
66. Tchoupa AK, Lichtenegger S, Reidl J, Hauck CR. Outer membrane protein P1 is the CEACAM-binding adhesin of Haemophilus influenzae. Mol Microbiol. 2015;98(3):440–455. doi:10.1111/mmi.13134
67. Weeks JR, Staples KJ, Spalluto CM, Watson A, Wilkinson TMA. the role of non-typeable Haemophilus influenzae biofilms in chronic obstructive pulmonary disease. Front Cell Infect Microbiol. 2021;11:720742. doi:10.3389/fcimb.2021.720742
68. Mokrzan EM, Ward MO, Bakaletz LO. Type IV pilus expression is upregulated in nontypeable Haemophilus influenzae biofilms formed at the temperature of the human nasopharynx. J Bacteriol. 2016;198(19):2619–2630. doi:10.1128/JB.01022-15
69. Hong W, Pang B, West-Barnette S, Swords WE. Phosphorylcholine expression by nontypeable Haemophilus influenzae correlates with maturation of biofilm communities in vitro and in vivo. J Bacteriol. 2007;189(22):8300–8307. doi:10.1128/JB.00532-07
70. Swords WE, Moore ML, Godzicki L, Bukofzer G, Mitten MJ, VonCannon J. Sialylation of lipooligosaccharides promotes biofilm formation by nontypeable Haemophilus influenzae. Infect Immun. 2004;72(1):106–113. doi:10.1128/IAI.72.1.106-113.2004
71. Puig C, Marti S, Hermans PW, et al. Incorporation of phosphorylcholine into the lipooligosaccharide of nontypeable Haemophilus influenzae does not correlate with the level of biofilm formation in vitro. Infect Immun. 2014;82(4):1591–1599. doi:10.1128/IAI.01445-13
72. Langereis JD, Hermans PW. Novel concepts in nontypeable Haemophilus influenzae biofilm formation. FEMS Microbiol Lett. 2013;346(2):81–89. doi:10.1111/1574-6968.12203
73. Armbruster CE, Pang B, Murrah K, et al. RbsB (NTHI_0632) mediates quorum signal uptake in nontypeable Haemophilus influenzae strain 86-028NP. Mol Microbiol. 2011;82(4):836–850. doi:10.1111/j.1365-2958.2011.07831.x
74. Armbruster CE, Hong W, Pang B, et al. LuxS promotes biofilm maturation and persistence of nontypeable haemophilus influenzae in vivo via modulation of lipooligosaccharides on the bacterial surface. Infect Immun. 2009;77(9):4081–4091. doi:10.1128/IAI.00320-09
75. Hong W, Juneau RA, Pang B, Swords WE. Survival of bacterial biofilms within neutrophil extracellular traps promotes nontypeable Haemophilus influenzae persistence in the chinchilla model for otitis media. J Innate Immun. 2009;1(3):215–224. doi:10.1159/000205937
76. Pang B, Armbruster CE, Foster G, Learman BS, Gandhi U, Swords WE. Autoinducer 2 (AI-2) production by nontypeable Haemophilus influenzae 86-028NP promotes expression of a predicted glycosyltransferase that is a determinant of biofilm maturation, prevention of dispersal, and persistence in vivo. Infect Immun. 2018;86(12):10–128.
77. Souza JGS, Bertolini M, Thompson A, et al. Role of glucosyltransferase R in biofilm interactions between Streptococcus oralis and Candida albicans. ISME J. 2020;14(5):1207–1222. doi:10.1038/s41396-020-0608-4
78. Domenech M, Pedrero-Vega E, Prieto A, Garcia E. Evidence of the presence of nucleic acids and beta-glucan in the matrix of non-typeable Haemophilus influenzae in vitro biofilms. Sci Rep. 2016;6:36424. doi:10.1038/srep36424
79. Jones EA, McGillivary G, Bakaletz LO. Extracellular DNA within a nontypeable Haemophilus influenzae-induced biofilm binds human beta defensin-3 and reduces its antimicrobial activity. J Innate Immun. 2013;5(1):24–38. doi:10.1159/000339961
80. Cavaliere R, Ball JL, Turnbull L, Whitchurch CB. The biofilm matrix destabilizers, EDTA and DNaseI, enhance the susceptibility of nontypeable Hemophilus influenzae biofilms to treatment with ampicillin and ciprofloxacin. Microbiologyopen. 2014;3(4):557–567. doi:10.1002/mbo3.187
81. Devaraj A, Buzzo J, Rocco CJ, Bakaletz LO, Goodman SD. The DNABII family of proteins is comprised of the only nucleoid associated proteins required for nontypeable Haemophilus influenzae biofilm structure. Microbiologyopen. 2018;7(3):e00563. doi:10.1002/mbo3.563
82. Jurcisek JA, Brockman KL, Novotny LA, Goodman SD, Bakaletz LO. Nontypeable Haemophilus influenzae releases DNA and DNABII proteins via a T4SS-like complex and ComE of the type IV pilus machinery. Proc Natl Acad Sci U S A. 2017;114(32):E6632–E6641. doi:10.1073/pnas.1705508114
83. Jurcisek JA, Hofer LK, Goodman SD, Bakaletz LO. Monoclonal antibodies that target extracellular DNABII proteins or the type IV pilus of nontypeable Haemophilus influenzae (NTHI) worked additively to disrupt 2-genera biofilms. Biofilm. 2022;4:100096. doi:10.1016/j.bioflm.2022.100096
84. Slinger R, Chan F, Ferris W, et al. Multiple combination antibiotic susceptibility testing of nontypeable Haemophilus influenzae biofilms. Diagn Microbiol Infect Dis. 2006;56(3):247–253. doi:10.1016/j.diagmicrobio.2006.04.012
85. Wu S, Li X, Gunawardana M, et al. Beta- lactam antibiotics stimulate biofilm formation in non-typeable haemophilus influenzae by up-regulating carbohydrate metabolism. PLoS One. 2014;9(7):e99204. doi:10.1371/journal.pone.0099204
86. Post DM, Held JM, Ketterer MR, et al. Comparative analyses of proteins from Haemophilus influenzae biofilm and planktonic populations using metabolic labeling and mass spectrometry. BMC Microbiol. 2014;14:329. doi:10.1186/s12866-014-0329-9
87. Cherkaoui A, Diene SM, Renzoni A, et al. Imipenem heteroresistance in nontypeable Haemophilus influenzae is linked to a combination of altered PBP3, slow drug influx and direct efflux regulation. Clin Microbiol Infect. 2017;23(2):118 e9–118 e19. doi:10.1016/j.cmi.2016.10.009
88. Flemming HC, Wingender J, Szewzyk U, Steinberg P, Rice SA, Kjelleberg S. Biofilms: an emergent form of bacterial life. Nat Rev Microbiol. 2016;14(9):563–575. doi:10.1038/nrmicro.2016.94
89. Puig C, Domenech A, Garmendia J, et al. Increased biofilm formation by nontypeable Haemophilus influenzae isolates from patients with invasive disease or otitis media versus strains recovered from cases of respiratory infections. Appl Environ Microbiol. 2014;80(22):7088–7095. doi:10.1128/AEM.02544-14
90. Reimche JL, Kirse DJ, Whigham AS, Swords WE. Resistance of non-typeable Haemophilus influenzae biofilms is independent of biofilm size. Pathog Dis. 2017;75(1):ftw112.
91. Juneau RA, Pang B, Weimer KE, Armbruster CE, Swords WE. Nontypeable Haemophilus influenzae initiates formation of neutrophil extracellular traps. Infect Immun. 2011;79(1):431–438. doi:10.1128/IAI.00660-10
92. Juneau RA, Pang B, Armbruster CE, Murrah KA, Perez AC, Swords WE. Peroxiredoxin-glutaredoxin and catalase promote resistance of nontypeable Haemophilus influenzae 86-028NP to oxidants and survival within neutrophil extracellular traps. Infect Immun. 2015;83(1):239–246. doi:10.1128/IAI.02390-14
93. Murphy TF, Kirkham C, Gallo MC, Yang Y, Wilding GE, Pettigrew MM. Immunoglobulin a protease variants facilitate intracellular survival in epithelial cells by nontypeable haemophilus influenzae that persist in the human respiratory tract in chronic obstructive pulmonary disease. J Infect Dis. 2017;216(10):1295–1302. doi:10.1093/infdis/jix471
94. Andresen E, Gunther G, Bullwinkel J, Lange C, Heine H. Increased expression of beta-defensin 1 (DEFB1) in chronic obstructive pulmonary disease. PLoS One. 2011;6(7):e21898. doi:10.1371/journal.pone.0021898
95. Peters BM, Jabra-Rizk MA, O’May GA, Costerton JW, Shirtliff ME. Polymicrobial interactions: impact on pathogenesis and human disease. Clin Microbiol Rev. 2012;25(1):193–213. doi:10.1128/CMR.00013-11
96. Jacobs DM, Ochs-Balcom HM, Zhao J, Murphy TF, Sethi S. Lower airway bacterial colonization patterns and species-specific interactions in chronic obstructive pulmonary disease. J Clin Microbiol. 2018;56(10):10–128.
97. Weimer KE, Juneau RA, Murrah KA, et al. Divergent mechanisms for passive pneumococcal resistance to beta-lactam antibiotics in the presence of Haemophilus influenzae. J Infect Dis. 2011;203(4):549–555. doi:10.1093/infdis/jiq087
98. Hong W, Khampang P, Erbe C, Kumar S, Taylor SR, Kerschner JE. Nontypeable Haemophilus influenzae inhibits autolysis and fratricide of Streptococcus pneumoniae in vitro. Microbes Infect. 2014;16(3):203–213. doi:10.1016/j.micinf.2013.11.006
99. Cope EK, Goldstein-Daruech N, Kofonow JM, et al. Regulation of virulence gene expression resulting from Streptococcus pneumoniae and nontypeable Haemophilus influenzae interactions in chronic disease. PLoS One. 2011;6(12):e28523. doi:10.1371/journal.pone.0028523
100. Bair KL, Campagnari AA. Moraxella catarrhalis promotes stable polymicrobial biofilms with the major otopathogens. Front Microbiol. 2020;10:3006. doi:10.3389/fmicb.2019.03006
101. Martinez-Resendez MF, Gonzalez-Chavez JM, Garza-Gonzalez E, et al. Non-typeable Haemophilus influenzae biofilm production and severity in lower respiratory tract infections in a tertiary hospital in Mexico. J Med Microbiol. 2016;65(12):1385–1391. doi:10.1099/jmm.0.000369
102. Naito S, Takeuchi N, Ohkusu M, et al. Clinical and bacteriologic analysis of Nontypeable Haemophilus influenzae strains isolated from children with invasive diseases in Japan from 2008 to 2015. J Clin Microbiol. 2018;56(7). doi:10.1128/JCM.00141-18
103. Vermee Q, Cohen R, Hays C, et al. Biofilm production by Haemophilus influenzae and Streptococcus pneumoniae isolated from the nasopharynx of children with acute otitis media. BMC Infect Dis. 2019;19(1):44. doi:10.1186/s12879-018-3657-9
104. Tagliaferri TL, Jansen M, Horz H-P. Fighting pathogenic bacteria on two fronts: phages and antibiotics as combined strategy. Front Cell Infect Microbiol. 2019;9:22. doi:10.3389/fcimb.2019.00022
105. Chaudhry WN, Concepcion-Acevedo J, Park T, Andleeb S, Bull JJ, Levin BR. Synergy and order effects of antibiotics and phages in killing Pseudomonas aeruginosa biofilms. PLoS One. 2017;12(1):e0168615. doi:10.1371/journal.pone.0168615
106. Wajima T, Anzai Y, Yamada T, Ikoshi H, Noguchi N, Omri A. Oldenlandia diffusa extract inhibits biofilm formation by Haemophilus influenzae clinical isolates. PLoS One. 2016;11(11):e0167335. doi:10.1371/journal.pone.0167335
107. Balazs VL, Horvath B, Kerekes E, et al. Anti-haemophilus activity of selected essential oils detected by TLC-direct bioautography and biofilm inhibition. Molecules. 2019;24(18):3301.
108. Balazs VL, Filep R, Repas F, et al. Immortelle (Helichrysum italicum (Roth) G. Don) essential oil showed antibacterial and biofilm inhibitory activity against respiratory tract pathogens. Molecules. 2022;27(17):5518.
109. Piasecki B, Balazs VL, Kieltyka-Dadasiewicz A, et al. Microbiological studies on the influence of essential oils from several origanum species on respiratory pathogens. Molecules. 2023;28(7):3044.
110. Kunthalert D, Baothong S, Khetkam P, Chokchaisiri S, Suksamrarn A. A chalcone with potent inhibiting activity against biofilm formation by nontypeable Haemophilus influenzae. Microbiol Immunol. 2014;58(10):581–589. doi:10.1111/1348-0421.12194
111. Arenas J, Szabo Z, van der Wal J, et al. Serum proteases prevent bacterial biofilm formation: role of kallikrein and plasmin. Virulence. 2021;12(1):2902–2917. doi:10.1080/21505594.2021.2003115
112. Luke-Marshall NR, Hansen LA, Shafirstein G, Campagnari AA. Antimicrobial photodynamic therapy with chlorin e6 is bactericidal against biofilms of the primary human otopathogens. mSphere. 2020;5(4):10–128.
113. Bair KL, Shafirstein G, Campagnari AA. In vitro photodynamic therapy of polymicrobial biofilms commonly associated with otitis media. Front Microbiol. 2020;11:558482. doi:10.3389/fmicb.2020.558482
114. Umar NK, Kono M, Sakatani H, et al. Respiratory quinolones can eradicate amoxicillin-induced mature biofilms and nontypeable Haemophilus influenzae in biofilms. J Infect Chemother. 2022;28(12):1595–1604. doi:10.1016/j.jiac.2022.07.019
115. Uemura Y, Qin L, Gotoh K, Ohta K, Nakamura K, Watanabe H. Comparison study of single and concurrent administrations of carbapenem, new quinolone, and macrolide against in vitro nontypeable Haemophilus influenzae mature biofilms. J Infect Chemother. 2013;19(5):902–908. doi:10.1007/s10156-013-0598-5
116. Farrell DJ, Morrissey I, Bakker S, Buckridge S, Felmingham D. Global distribution of TEM-1 and ROB-1 beta-lactamases in Haemophilus influenzae. J Antimicrob Chemother. 2005;56(4):773–776. doi:10.1093/jac/dki281
117. Dabernat H, Delmas C, Seguy M, et al. Diversity of beta-lactam resistance-conferring amino acid substitutions in penicillin-binding protein 3 of Haemophilus influenzae. Antimicrob Agents Chemother. 2002;46(7):2208–2218. doi:10.1128/AAC.46.7.2208-2218.2002
118. Garcia-Cobos S, Campos J, Lazaro E, et al. Ampicillin-resistant non-beta-lactamase-producing Haemophilus influenzae in Spain: recent emergence of clonal isolates with increased resistance to cefotaxime and cefixime. Antimicrob Agents Chemother. 2007;51(7):2564–2573. doi:10.1128/AAC.00354-07
119. Qin L, Zhou Z, Hu B, Yamamoto T, Watanabe H. Antimicrobial susceptibility and genetic characteristics of Haemophilus influenzae isolated from community-acquired respiratory tract infection patients in Shanghai City, China. J Infect Chemother. 2012;18(4):508–514. doi:10.1007/s10156-012-0372-0
120. Chen D, Wen S, Feng D, et al. Microbial virulence, molecular epidemiology and pathogenic factors of fluoroquinolone-resistant Haemophilus influenzae infections in Guangzhou, China. Ann Clin Microbiol Antimicrob. 2018;17(1):41. doi:10.1186/s12941-018-0290-9
121. Taha A, Adeline F, Taha MK, Deghmane AE. Haemophilus influenzae drug resistance in France from 2017 to 2021: consideration for treatment of otitis media. J Glob Antimicrob Resist. 2022;31:222–227. doi:10.1016/j.jgar.2022.09.008
122. Ho PL, Chow KH, Mak GC, et al. Decreased levofloxacin susceptibility in Haemophilus influenzae in children, Hong Kong. Emerg Infect Dis. 2004;10(11):1960–1962. doi:10.3201/eid1011.040055
123. Tsang RSW, Shuel M, Whyte K, et al. Antibiotic susceptibility and molecular analysis of invasive Haemophilus influenzae in Canada, 2007 to 2014. J Antimicrob Chemother. 2017;72(5):1314–1319. doi:10.1093/jac/dkw565
124. Puig C, Tirado-Velez JM, Calatayud L, et al. Molecular characterization of fluoroquinolone resistance in nontypeable Haemophilus influenzae clinical isolates. Antimicrob Agents Chemother. 2015;59(1):461–466. doi:10.1128/AAC.04005-14
125. Chang CM, Shih HI, Wu CJ, et al. Fluoroquinolone resistance in Haemophilus influenzae from nursing home residents in Taiwan: correlation of MICs and mutations in QRDRs. J Appl Microbiol. 2020;128(6):1624–1633. doi:10.1111/jam.14580
126. Prymula R, Peeters P, Chrobok V, et al. Pneumococcal capsular polysaccharides conjugated to protein D for prevention of acute otitis media caused by both Streptococcus pneumoniae and non-typable Haemophilus influenzae: a randomised double-blind efficacy study. Lancet. 2006;367(9512):740–748. doi:10.1016/S0140-6736(06)68304-9
127. Saez-Llorens X, Rowley S, Wong D, et al. Efficacy of 10-valent pneumococcal non-typeable Haemophilus influenzae protein D conjugate vaccine against acute otitis media and nasopharyngeal carriage in Panamanian children - A randomized controlled trial. Hum Vaccin Immunother. 2017;13(6):1–16. doi:10.1080/21645515.2017.1287640
128. Novotny LA, Bakaletz LO. Transcutaneous immunization with a nontypeable Haemophilus influenzae dual adhesin-directed immunogen induces durable and boostable immunity. Vaccine. 2020;38(10):2378–2386. doi:10.1016/j.vaccine.2020.01.052
129. Michel LV, Kaur R, Gleghorn ML, et al. Haemophilus influenzae protein D antibody suppression in a multi-component vaccine formulation. FEBS Open Bio. 2022;12(12):2191–2202. doi:10.1002/2211-5463.13498
130. Ysebaert C, Denoel P, Weynants V, et al. A protein E-PilA fusion protein shows vaccine potential against nontypeable Haemophilus influenzae in mice and chinchillas. Infect Immun. 2019;87(8):10–128.
131. Nakahashi-Ouchida R, Mori H, Yuki Y, et al. Induction of mucosal IgA-mediated protective immunity against nontypeable Haemophilus influenzae infection by a cationic nanogel-based P6 nasal vaccine. Front Immunol. 2022;13:819859. doi:10.3389/fimmu.2022.819859
132. Martyn L, Sethia R, Chon R, et al. Antibodies against the DNABII protein integration host factor (IHF) inhibit sinus implant biofilms. Laryngoscope. 2020;130(6):1364–1371. doi:10.1002/lary.28188
133. Clark SL, Seppanen EJ, Kirkham LS, et al. Australian aboriginal otitis-prone children produce high-quality serum IgG to putative nontypeable haemophilus influenzae vaccine antigens at lower titres compared to non-aboriginal children. Front Cell Infect Microbiol. 2022;12:767083. doi:10.3389/fcimb.2022.767083
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.