Back to Journals » Infection and Drug Resistance » Volume 12
Enhancing the antibacterial activity of polymyxins using a nonantibiotic drug
Authors Krishnamurthy M, Lemmon MM, Falcinelli EM, Sandy RA, Dootz JN, Mott TM, Rajamani S, Schaecher KE, Duplantier AJ, Panchal RG
Received 3 December 2018
Accepted for publication 6 February 2019
Published 27 May 2019 Volume 2019:12 Pages 1393—1405
DOI https://doi.org/10.2147/IDR.S196874
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Joachim Wink
Malathy Krishnamurthy,1 Margaret M Lemmon,1 Evan M Falcinelli,1 Reuel A Sandy,1 Jennifer N Dootz,1 Tiffany M Mott,1 Sathish Rajamani,1,2 Kurt E Schaecher,3 Allen J Duplantier,1,4 Rekha G Panchal1
1Department of Target Discovery and Experimental Microbiology, Division of Molecular and Translational Sciences, United States Army Medical Research Institute of Infectious Diseases, Frederick, MD, USA; 2General Dynamics Information Technology, Frederick, MD, USA; 3Department of Pathology, Walter Reed National Military Medical Center, Bethesda, MD, USA; 4Cherokee Nation Assurance, Frederick, MD, USA
Purpose: The rapid emergence of multidrug-resistant (MDR) bacteria and the lack of new therapies to eliminate them poses a major threat to global health. With the alarming rise in antimicrobial resistance (AMR), focus has now shifted to the use of the polymyxin class of antibiotics as the last line of defense for treatment of Gram-negative infections. Unfortunately, the growing resistance of bacteria against polymyxins is threatening the treatment of MDR infections, necessitating the need for novel strategies. The objective of this study was to determine if combination of polymyxin (polymyxin B or colistin) with a nonantibiotic small molecule AR-12, a celecoxib derivative that is devoid of cyclooxygenase 2 (COX-2) inhibitory activities, can be an effective strategy against polymyxin-resistant MDR bacteria.
Methods: Growth inhibition studies, time-kill assays and permeability assays were conducted to investigate the effect of AR-12 on the antibacterial activity of polymyxins.
Results: Growth studies were performed on a panel of polymyxin-resistant MDR strains using the combination of AR-12 with either colistin or polymyxin B. The combination treatment had no effect on strains that have inherent polymyxin resistance; however, AR-12 was effective in lowering the minimal inhibitory concentration (MIC) of polymyxins by 4–60-fold in several strains that had acquired polymyxin resistance. Time-kill assays using the combination of AR-12 and colistin with select MDR strains suggest rapid killing and bactericidal activity, while the permeability assays using fluorescently labeled dansylated polymyxin and 1-N-phenylnaphthylamine (NPN) in these MDR strains suggest that AR-12 can potentiate the antibacterial activity of polymyxins by possibly altering the bacterial outer membrane via modification of lipopolysaccharide and thereby improving the uptake of polymyxins.
Conclusion: Our studies indicate that the combination of AR-12 and polymyxin is effective in targeting select Gram-negative bacteria that have acquired polymyxin resistance. Further understanding of the mechanism of action of AR-12 will provide new avenues for developing narrow-spectrum antibacterials to target select Gram-negative MDR bacteria. Importantly, our studies show that the use of nonantibiotic small molecules in combination with polymyxins is an attractive strategy to counter the growing resistance of bacteria to polymyxins.
Keywords: Gram-negative, MDR bacteria, potentiator, antimicrobial resistance
Introduction
The alarming rise of multidrug-resistant (MDR) pathogenic organisms has become a significant global health challenge. According to The Review on Antimicrobial Resistance (AMR), which was established jointly by the United Kingdom (UK) Government and Wellcome Trust in 2014, it is estimated that each year, at least 50,000 people die due to infections caused by MDR pathogens in Europe and US alone.1 The emergence of MDR bacteria has been rampant and is the primary cause for nosocomial infections which are life threatening.2 In particular, among MDR Gram-negative bacteria, nosocomial infections that are caused by Klebsiella pneumoniae, Acinetobacter baumannii, Pseudomonas aeruginosa, Enterobacter species and Escherichia coli pathogens are posing the greatest threat to human and animal health. To treat infections caused by MDR pathogens, focus has now shifted to the use of polymyxins (colistin and polymyxin B), the class of antibiotics typically reserved as the last line of defense. In 2017, the World Health Organization (WHO) classified polymyxins among the most critically important antibiotics with the highest priority for human medicine (
Polymyxins are known to target the outer membrane of Gram-negative bacteria by displacing divalent calcium and magnesium ions from the negatively charged phosphate groups of membrane lipids. This causes destabilization of the lipopolysaccharide (LPS) and the membrane, leading to leakage of cytoplasmic contents and bacterial cell death.4 Although a clear mechanism of action is not known, it is also suggested that polymyxins can bind to endotoxin, which is the lipid A portion of the LPS and neutralize LPS during cell lysis. Polymyxins have been shown to have bactericidal effect against several Gram-negative pathogens such as E.coli, A. baumannii, P. aeruginosa, K. pneumoniae, Enterobacter spp., Salmonella spp. and Shigella spp. Until recently, the mechanisms of reported polymyxin resistance have been chromosomally mediated that result in LPS modifications, formation of capsules and overexpression of outer membrane protein OprH, thereby conferring resistance in Gram-negative bacteria.4,5 In 2015, a plasmid-mediated polymyxin resistance mechanism (MCR-1) was reported in Enterobacteriaceae, which is further compromising and threatening the treatment using polymyxin antibiotics.6
To address the escalating problem of MDR, a global action plan was recommended at the 68th World Health Assembly in 2015, which appeals for development of new antibiotics, diagnostic tools and other approaches to quickly identify and treat MDR infections (
With respect to combating the rise of polymyxin-resistant bacterial pathogens, the use of small molecule adjuvants in combination with polymyxins may be highly valuable for restoring polymyxin sensitivity against MDR Gram-negative bacterial pathogens. The utilization of small molecule adjuvants can also potentially reduce safety concerns such as nephrotoxicity and neurological disturbances associated with the prolonged and systemic use of polymyxin antibiotics by reducing the therapeutic dose of polymyxin required for treatment. Recently, Stokes et al have shown that the combination of an anticoumarin antibiotic (novobiocin) in combination with an antiprotozoal agent (pentamidine) was highly effective in clearing bacterial load in mice infected with colistin-resistant A. baumannii.11 In another example, Harris et al have shown that 2-aminoimidazole-based adjuvants can overcome colistin resistance in A. baumannii and K. pneumoniae strains by interfering with the expression of a two-component system in bacteria.12 The above examples demonstrate that the use of combination therapy is an attractive alternative for the development of new therapies for treating infections caused by polymyxin-resistant bacteria.
To address the problem of polymyxin resistance in MDR bacteria, we have utilized a similar strategy of using a small molecule adjuvant, AR-12 in combination with polymyxin B or colistin against a panel of MDR bacteria. AR-12 is a celecoxib derivative that is devoid of cyclooxygenase 2 (COX-2) inhibitory activities in eukaryotic host.13 It is an autophagy inducing small molecule that was developed by researchers at the Ohio State University (licensed to Arno Therapeutics Inc.) and taken into the clinical settings as it was found to be effective against a wide variety of cancer types.14 Previous studies done by Chiu et al have shown that AR-12 can induce autophagy in THP-1 macrophages and inhibit the intracellular survival of Gram-negative Francisella tularensis and F. novicida bacterial strains in macrophages.15 In these studies, it was found that AR-12 did not have any direct antibacterial activity against the Francisella strains and it was found that the inhibition of intracellular survival of Francisella is mediated indirectly by induction of autophagy of the host cells. Interestingly, in another study done by Booth et al, it was found that AR-12 significantly inhibited the growth of a laboratory-generated ampicillin and kanamycin-resistant E. coli strain and reduced the proliferation of an MDR clinical isolate of N. gonorrhoeae, demonstrating that AR-12 can directly act against certain bacteria.16 In these Gram-negative bacteria, AR-12 was found to reduce the expression of DnaK, which is essential for bacterial growth and mRNA/protein stability and chaperone protein RecA. Though only a select strains have been studied for determining the antibacterial activity of AR-12, the above studies indicate that AR-12 can potentially target certain Gram-negative bacterial species over others and will be a valuable orally bioavailable small molecule that can be evaluated for antibacterial activity against other MDR Gram-negative bacteria.
Considering the urgent need for evaluating new chemical scaffolds against MDR bacteria, the objective of this study was to determine the antibacterial activity of AR-12 in a panel of MDR Gram-negative bacteria with specific focus on polymyxin-resistant strains. Further, we also wanted to determine if susceptibility of bacteria that are resistant to polymyxin can be enhanced by utilizing AR-12 as an adjuvant in combination with polymyxins. Herein, we report that even though AR-12 does not have direct antibacterial activity against polymyxin-resistant MDR strains studied, AR-12 is able to potentiate the antibacterial activity of polymyxins in these strains significantly. Our studies show that the combination of AR-12 with either polymyxin B or colistin is very effective as an antibacterial agent against several pathogens including polymyxin-resistant MDR A. baumannii and MDR K. pneumoniae. Growth inhibition studies, time-kill experiments and permeability assays were conducted to investigate the potentiation of the antibacterial activity of polymyxins by AR-12. Our results highlight the significance of using small molecule adjuvants as an alternative approach to counter MDR pathogens by enhancing the antibacterial activity of polymyxins.
Materials and methods
Bacterial strains and media
Clinical isolates of P. aeruginosa, K. pneumoniae, A. baumannii, E.coli, Serratia marcescens, Proteus mirabilis, Enterobacter cloacae and Providencia stuartii (Tables 1 and S1) were utilized for determining the antimicrobial activity of the antibiotics and the combinations. The isolates were obtained from the culture collections at either JMI Laboratories, Centers for Disease Control and Prevention (CDC), Walter Reed National Military Medical Center (WRNMMC) or internal culture collection at USAMRIID. Mueller-Hinton II broth-Cation Adjusted (CAMHB) (Catalog Number 212322) and Mueller-Hinton II Agar-Cation Adjusted (Catalog Number 211438) were purchased from BD Diagnostics. HEPES 1 M solution (pH 7.0–7.6) (Catalog Number H0887) was purchased from Sigma-Aldrich. Phosphate-Buffered Saline (PBS), 1X without Calcium and Magnesium (Catalog Number 21-040-CV) was purchased from Corning, Inc.
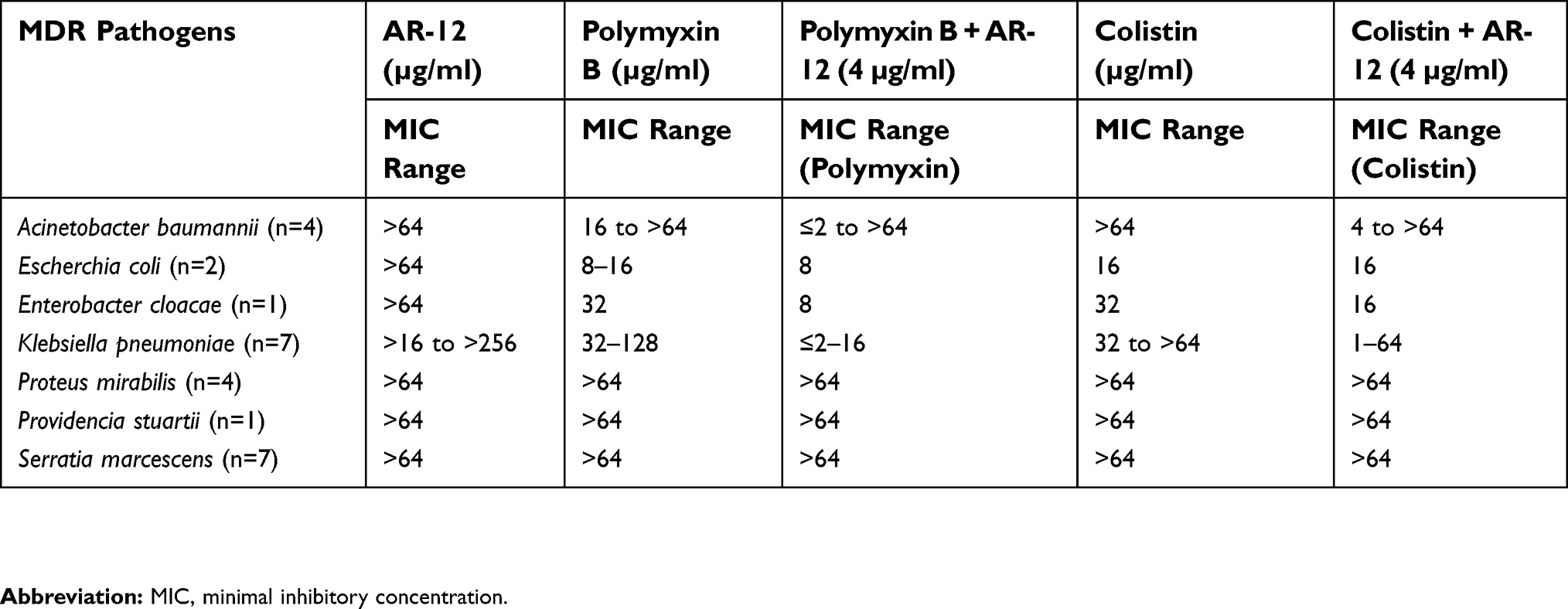 | Table 1 Effect of AR-12 alone and AR-12 in combination with polymyxins (Polymyxin B or Colistin) against Gram-negative MDR pathogens |
Synthetic compounds and antibiotics
Synthetic compound AR-12 was generously provided by Arno Therapeutics, Inc. Dansyl polymyxin was synthesized by Alta Biosciences, UK. N-Phenyl-1-naphthylamine (NPN) (Catalog number 104043) and polymyxin B sulfate salt (Catalog Number P1004) were purchased from Sigma-Aldrich. Stock solution of AR-12 was prepared in Dimethyl sulfoxide (DMSO). Stock solution of NPN was prepared in acetone. Stock solutions of dansyl polymyxin, polymyxin B sulfate, colistinmethate were prepared in sterile water.
Broth microdilution method for determination of minimum inhibitory concentrations (MIC)
MIC assays were performed in 96-well round bottom plates. Each well was pre-dispensed with the appropriate stock concentrations of the drugs or controls for testing. The volumes of the drugs were adjusted in order to get the final required concentration of the drug in each well. Bacterial cultures were prepared from bacteria grown on Mueller-Hinton II agar plates incubated at 37°C overnight. Several colonies of each strain were then suspended in 1X Phosphate Buffered Saline (PBS) and OD 600 was measured. Based on the OD 600 value, the bacterial suspension was diluted in CAMHB to yield the inoculum at a concentration of 106 CFU/ml. Bacterial culture was added to the prepared 96-well plate by either pipetting or using “INTEGRA Viafill” reagent dispenser, resulting in a final bacterial concentration of 5×105 CFU/ml. The plate was incubated at 37°C without shaking for 18–24 hrs and then evaluated visually for the presence or absence of growth. The MIC was defined as the lowest concentration of compound that completely inhibited growth. For checkerboard assays, the concentration ranges for polymyxin B and colistin were selected to encompass the MIC value determined in the preliminary MIC assay. The concentration range for AR-12 was selected to encompass 4 μg/ml. Two-fold serial dilutions of polymyxin B or colistin (dilutions ranging from 0.13–64 or 128 μg/ml) were prepared across the rows of a 96-well round bottom plate containing CAMHB. Two-fold serial dilutions of AR-12 (dilutions ranging from 1–64 or 128 μg/ml) were performed down the columns of a 96-well round bottom plate containing CAMHB. Appropriate volumes of polymyxin B or colistin compound preparations and AR-12 compound preparations were dispensed in a 96-well round bottom plate by either pipetting or using a PerkinElmer JANUS MDT Automated Workstation. Bacterial suspensions (final concentration of 5×105 CFU/ml) were added as described above and the 96-well plates incubated at 37°C without shaking for 16–20 hrs and then evaluated visually for the presence or absence of growth.
Bacterial growth assays
Bacterial growth assays were done using the Bioscreen C Automated Microbiology Growth Curve Analysis System (Growth Curves USA). Bacteria were grown by inoculating colonies from freshly streaked Mueller-Hinton II agar plates into CAMHB at 37°C with shaking for ~16 hrs. A 1:100 dilution was subcultured in CAMHB and grown for additional 2–3 hrs to get log phage cultures. 5×105 CFU/ml of the log phase culture was then dispensed into sterile Honeycomb microwell plates (10 × 10 well plates) containing the various concentrations of drugs in CAMHB pre-dispensed into the plates (in triplicates), such that total volume in each well was 100 μl. The plates were then incubated with shaking at 37°C in a Bioscreen C Analyzer System (Growth Curves USA). The optical density (OD 600) of each microwell was measured automatically at 15 mins intervals for 24 hrs. The MIC was determined to be the lowest concentration of the drug which showed complete growth inhibition. The experiments were repeated two or more independent times to verify results.
Time-kill assays
A. baumannii strain 34299 and K. pneumoniae strain 30835 were grown by inoculating colonies from freshly streaked Mueller-Hinton II agar plates into CAMHB at 37°C with shaking for ~16 hrs. A 1:100 dilution was subcultured in CAMHB and grown for additional 2–3 hrs to get log phage cultures. An inoculum of the log phase culture was diluted in 15 ml CAMHB to get a final culture of 5×105 CFU/ml. The tubes contained CAMHB alone or CAMHB containing AR-12 alone, colistin alone or combinations of colistin and AR-12. All the tubes were incubated at 37°C with shaking and samples were withdrawn at 1, 2, 4, 8 and 24 hrs, following which they were serially diluted and plated on Mueller-Hinton II agar plates in duplicates. Plates were incubated at 37°C overnight and colonies were enumerated next day. The experiment was repeated twice on two independent days for each time point and concentration of drug used.
Dansyl polymyxin permeability assay
A. baumannii strains and K. pneumoniae strains were grown by inoculating colonies from freshly streaked Mueller-Hinton II agar plates into CAMHB at 37°C with shaking for ~16 hrs. A 1:100 dilution was subcultured in CAMHB and grown for additional 2–3 hrs to get log phage cultures. OD 600 was measured and volume of culture amounting to 0.5 OD in 1 ml was centrifuged and re-suspended in 1 mL of 5 mM HEPES buffer and kept at room temperature. Greiner Bio-One 96 well plates (catalog #655096, black with clear bottom) were used for testing the permeability of Dansyl polymyxin in the presence and absence of AR-12. A mix consisting of 5 mM HEPES buffer- pH 7·2, dansyl polymyxin (0.6 μM) and appropriate AR-12 concentration (1.25 × stock) was prepared. 80 μl of this mix was suspended in the wells in triplicates, following which 20 μl aliquots of the bacterial suspension in 5 mM HEPES buffer was added. Fluorescence was read at room temperature in a Tecan Sapphire II multimode plate reader, with filters of 320 nm (bandwidth 10 nm) for excitation and 500 nm (bandwidth 10 nm) for emission. To determine the fluorescence due to dansyl polymyxin uptake, the fluorescence from the wells containing the dansyl polymyxin and AR-12 but with no cells was subtracted from the wells containing the cells and the mix. The experiments were repeated two or more independent times to verify results.
N-phenyl-1-naphthylamine (NPN) assay
Bacterial suspensions were prepared as described for the dansyl polymyxin permeability assay. Greiner Bio-One 96 well plates (catalog #655096, black with clear bottom) were used for testing the permeability of NPN in the presence and absence of AR-12. Stock concentration of NPN (5 mM) was made in acetone, following which a stock concentration of NPN (40 μM) in 5 mM HEPES-pH 7·2 was made from the 5 mM NPN stock. The 5 mM NPN stock can be stored in the −20°C for up to a month. A mix consisting of 5 mM HEPES buffer, NPN (40 μM) and AR-12 (10 × stock) was prepared for each concentration of AR-12 that was tested. 80 μl of this mix was suspended in the wells in triplicates, following which 20 μl aliquots of the bacterial suspension in 5 mM HEPES buffer was added. Fluorescence was read at room temperature in a Tecan Sapphire II multimode plate reader with filters of 355 nm (bandwidth 10 nm) for excitation and 405 nm (bandwidth 10 nm) for emission. To determine the fluorescence due to NPN uptake, the fluorescence from the wells containing the NPN and AR-12 but with no cells was subtracted from the wells containing the cells and the mix. The experiments were repeated two or more independent times to verify results.
Pharmacokinetic studies in mice
Pharmacokinetic (PK) studies of AR-12 compound in Balb/c mice were performed by Biotranex, LLC. at Project Safety Labs (PSL), located in New Jersey, USA. PSL facility is accredited by the Association for Assessment and Accreditation of Laboratory Animal Care International (AAALAC) and operates in compliance with all US Government regulations and industry standards for animal welfare as they apply to the testing conducted. PSL complies with all applicable provisions of the Animal Welfare Act and other Federal statutes and regulations relating to animals including the Animal and Plant Health Inspection Service (APHIS), US Department of Agriculture (USDA) and The Office of Laboratory Animal Welfare (OLAW). PSL has established and maintains an animal use and veterinary care program for activities involving animals in accordance with the most recent version of the Guide for the Care and Use of Laboratory Animals, Institute of Laboratory Animal Resources Commission on Life Sciences, National Academy Press, Washington, D.C. All study protocols involving the use of animals at PSL are thoroughly reviewed and approved by Biotranex, LLC’s Internal Animal Care and Use Committee (IACUC).
For the PK studies, AR-12 was formulated at 8 mg/mL in 10% Solutol HS 15. Mice (n=3 mice/time point) were administered AR-12 (80 mg/kg) via the oral route and at time points of T=0, 0.5, 1, 2, 4, 8, 10 and 24 hrs. Plasma samples were collected and quantitatively analyzed by HPLC.
Results and discussions
Minimal inhibitory concentration (MIC) assays using polymyxin and AR-12 combination
The potency of AR-12 as an antibacterial was evaluated against a panel of MDR Gram-negative pathogenic bacterial strains, including colistin-resistant clinical isolates, using standard broth dilution assays to determine MIC. We found that AR-12 did not show any antibacterial effect against the Gram-negative strains evaluated (Table 1). As it has been previously shown that AR-12 can reduce the growth of antibiotic-resistant E. coli strain and an MDR clinical isolate of N. gonorrhoeae,16 we wanted to determine if AR-12 could potentiate or act in conjunction with other antibiotics in effectively targeting Gram-negative bacteria. Hence, AR-12 was initially tested either alone or in combination with different classes of antibiotics against a select panel of MDR human clinical isolates (data not shown). AR-12 alone had no MIC value when tested upto 64 µg/ml against a panel of MDR pathogens (Tables 1 and S1). At higher concentrations due to the poor solubility of AR-12, the data could not be interpreted. Interestingly, we found that AR-12 was able to potentiate the antibacterial activity by lowering the MIC of polymyxins, when compared to polymyxins alone. Further, the potentiating effect of AR-12 with polymyxin B or colistin was observed only with certain Gram-negative bacterial strains, namely A. baumannii and K. pneumoniae. Significantly, in the presence of 4 μg/ml of AR-12 (the lowest AR-12 concentration at which the potentiating activity was observed for majority of the MDR strains), the MIC of polymyxin B was reduced by 4–16 fold for MDR A. baumannii (2 out of 4 strains tested) and K. pneumoniae strains (7 out of 7 strains tested) and the MIC of colistin was reduced by 4–16 fold for MDR A. baumannii (2 out of 4 strains tested) and 4–60 fold for MDR K. pneumoniae strains tested (5 out of 7 strains tested) (Table 1, Table S1). Interestingly, no potentiating effect of AR-12 with polymyxins was observed with representative clinical isolates of Gram-negative bacterial strains such as Proteus mirablis, Providencia stuartii, Serratia marcesecens and key biothreat pathogens Burkholderia pseudomallei and Burkholderia mallei (Tables 1 and S1). AR-12 also did not exhibit a combinatorial activity with other classes of antibiotics (data not shown), indicating its narrow spectrum role in potentiating the activity of polymyxins against select bacterial species, including their MDR variants.
Growth studies of MDR strains treated with polymyxin and AR-12 combination
To confirm the effect of AR-12 as a potentiator of colistin, we conducted growth studies on the MDR A. baumannii 34299 and K. pneumoniae 30835 clinical isolates (Figure 1). For MDR A. baumannii 34299, no growth inhibition was observed even at high concentrations of colistin (≥32 μg/ml) (Figure 1B). Similarly, growth inhibition of MDR K. pneumoniae 30835 was observed only at colistin concentration of at least 8 μg/ml but not at 4 μg/ml (Figure 1D). However, in the presence of 4 μg/ml AR-12, growth inhibition of MDR A. baumannii and MDR K. pneumoniae strains were observed at a significantly lower concentration of colistin (1 μg/ml). Similar effect was seen with the polymyxin sensitive American Type Culture Collection (ATCC) strains of A. baumannii 19606 and K. pneumoniae 13833, where growth inhibition of A. baumannii and K. pneumoniae were observed at much lower concentration of colistin in the presence of 4 μg/ml AR-12 (0.015 μg/ml colistin for A. baumannii 19606 and 0.125 μg/ml colistin for K. pneumoniae 13833) in comparison to growth inhibition with colistin alone (0.5 μg/ml colistin for A. baumannii 19606 and 2 μg/ml colistin for K. pneumoniae 13833) (Figure 1A and C). These results clearly demonstrate that AR-12 is able to sensitize certain Gram-negative bacteria to colistin and significantly lower the concentration of colistin required for antibacterial activity against MDR strains.
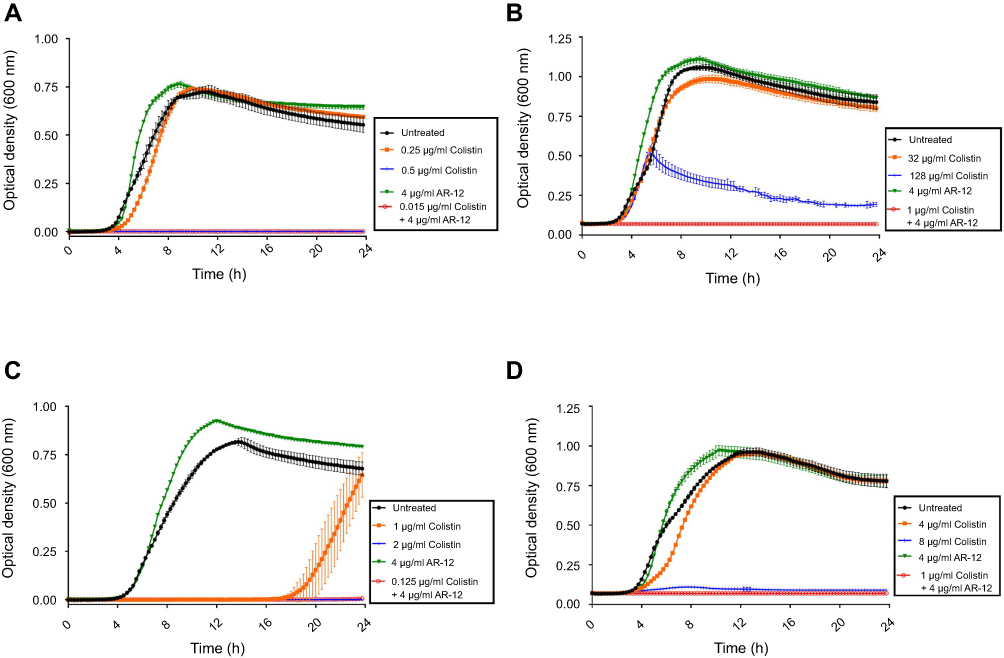 | Figure 1 Growth curves depicting the antibacterial efficacy of colistin-AR12 combination (A) A. baumannii ATCC 19606 polymyxin-sensitive strain; (B) A. baumannii 34299 polymyxin-resistant strain; (C) K. pneumoniae ATCC 13833 polymyxin-sensitive strain; (D) K. pneumoniae 30835 polymyxin-resistant strain. Growth assays demonstrate the growth inhibitory activity of AR-12/colistin combination against A. baumannii and K. pneumoniae strains. The experiments were repeated two or more independent times to verify results. |
Time-kill assays of MDR strains treated with colistin and AR-12 combination
To further determine the rate at which the combination of colistin and AR-12 exerts its antibacterial activity, we performed time-kill assays using the polymyxin-resistant A. baumannii 34299 and K. pneumoniae 30835 clinical isolates (Figure 2). In comparison to colistin alone, treatment with AR-12 and colistin combination resulted in >3log10 fold reduction in CFU/ml at 8 hrs time point, relative to the initial inoculum, for MDR A. baumannii. Similarly, for MDR K. pneumoniae strain, when treated with AR-12 and colistin combination, we observed a 2log10 fold reduction in CFU/ml at 8 hrs time point, relative to the initial inoculum. We observed rapid killing of bacteria within 4 hrs of treatment with the AR-12 and colistin combination and this bactericidal effect was observed up to 8 hrs. In the case of the combination treatment, we observed regrowth after 24 hrs which could be attributed to either degradation or metabolism of AR-12 and thus reduction in efficacy of the combination treatment at 24 hrs time point. Significantly, the data from the time-kill assays demonstrate the potent bactericidal activity of AR-12/colistin combination against MDR pathogens at much lower concentrations of colistin than what is required with colistin alone. Together, these studies suggest that AR-12 could be highly valuable in combination with polymyxins against the colistin-resistant strains, offering an avenue for repurposing these antibiotics that are generally reserved as last resort for treatment of MDR infections.
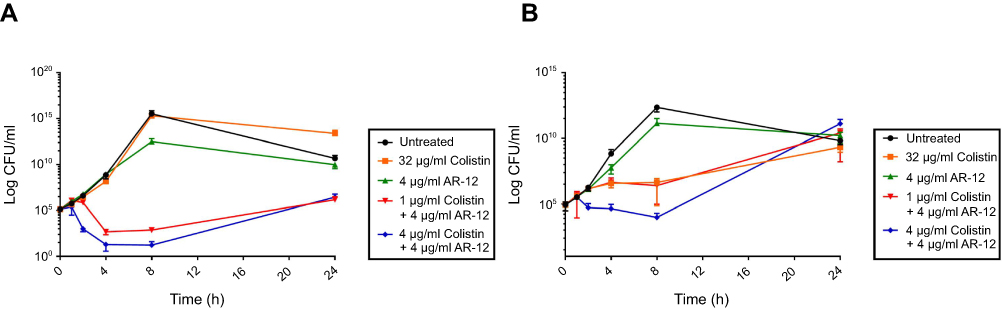 | Figure 2 Time-kill curves for MDR A. baumannii and K. pneumoniae strains by colistin-AR12 combinations: (A) Time-kill kinetics of MDR A. baumannii clinical isolate A. baumannii 34299, (B) Time-kill kinetics of MDR K. pneumoniae clinical isolate K. pneumoniae 30835. Time-kill assays demonstrate the potent bactericidal activity of AR-12/colistin combination against A. baumannii and K. pneumoniae strains. The data represent the averages ± SD (Standard Deviation) for two independent experiments. |
Membrane permeability assays
Antimicrobial peptides such as polymyxins are known to exert their antimicrobial activity by interacting with the lipid A component of the LPS in the outer membrane. The cationic side chains of polymyxin electrostatically bind to the anionic phosphate groups of the lipid A core of LPS and displace the divalent cations (Mg2+, Ca2+) leading to disruption of membrane integrity.4 It is well established that modifications of LPS, leading to overall less negative charge of the outer membrane in bacteria, can lead to polymyxin resistance due to its reduced ability to interact with bacterial membrane.5 Campos et al have shown previously that certain quinolone antibiotics such as ciprofloxacin and neamine derivatives can render Gram-negative bacteria sensitive to antimicrobial peptides.17 Their studies indicate that small molecules can disrupt the LPS by displacing divalent cations from their LPS-binding sites in polymyxin-resistant strains, thereby facilitating the interaction of the peptides with the membrane and leading to cell death.
In this study, we found that the MIC of colistin and polymyxin B can be reduced 4–60 fold in the presence of AR-12 for certain polymyxin-resistant Gram-negative bacterial strains. As the outer membrane of resistant bacteria could be impermeable to polymyxins, we hypothesized that AR-12 might be altering membrane integrity and sensitizing the bacterial strains to polymyxins thus facilitating the interaction of polymyxins with the bacterial membrane. To probe into the mechanism of action of AR-12, membrane permeabilization studies were performed using a fluorescently labeled polymyxin probe, dansyl polymyxin B (DP) (Alta Biosciences, UK) and its permeability was assessed in the presence or absence of AR-12. The fluorescence-based permeability assay was performed using the polymyxin-resistant A. baumannii 34299 and K. pneumoniae 30835 clinical isolates and their respective polymyxin-sensitive ATCC strains adapting the methods reported by Campos et al and Ortega et al.17,18 DP by itself has low fluorescence in aqueous buffer and its fluorescence increases in hydrophobic environments such as phospholipid bilayers. In our assays, notably, we observed that with increasing concentrations of AR-12, the fluorescence of DP was enhanced by 25–50 fold depending on the strain (Figure 3). This indicated that AR-12 is facilitating the uptake of polymyxin in these strains by potentially altering the membrane permeability. At 2 μg/ml and 4 μg/ml of AR-12, the fluorescence increase for the A. baumannii strains was higher than the respective K. pneumoniae strains. This could be due to the differences in the membrane structure of the two bacterial strains.
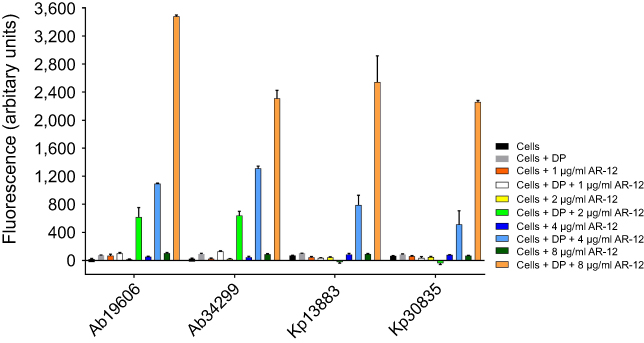 | Figure 3 Outer membrane permeability of Dansyl polymyxin (DP) in the presence and absence of AR-12. Outer membrane permeability of DP increases in the presence of AR-12 as seen with increase in fluorescence of DP with increasing concentrations of AR-12. A. baumannii 19606 and K. pneumoniae 13883 are ATCC strains that are polymyxin sensitive. A. baumannii 34299 and K. pneumoniae 30835 are MDR clinical isolates that are polymyxin resistant. The experiments were repeated two or more independent times to verify results. |
We next assessed the outer membrane permeability of AR-12 using 1-N-Phenyl-naphthylamine (NPN) assay developed by Hancock and Wong.7,19,20 Similar to DP, NPN is a hydrophobic fluorescent probe and its fluorescence increases when it is transferred from aqueous to hydrophobic environment. To test if AR-12 facilitates the uptake of NPN in bacterial strains, we measured the fluorescence of NPN in the presence of AR-12. As hypothesized, we found that the addition of AR-12 indeed increased the fluorescence of NPN (Figure 4). At 4 μg/ml and 8 μg/ml of AR-12, increase in NPN fluorescence was observed for both A. baumannii and K. pneumoniae strains; however, a much bigger increase in fluorescence was observed for the A. baumannii strains over the K. pneumoniae strains, an observation similar to the DP assay. Based on the observations from the above studies, we propose that AR-12 plays a role in altering the bacterial outer membrane by modifying the LPS and improving the uptake of polymyxins in colistin-resistant strains.
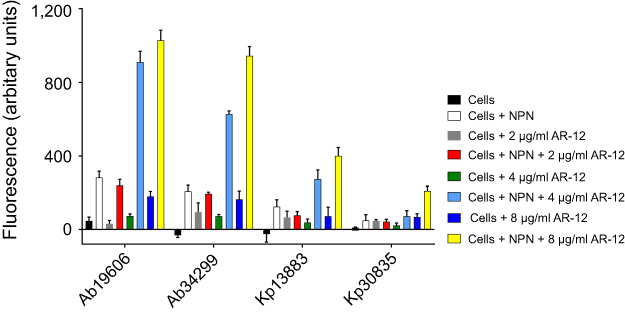 | Figure 4 Membrane permeabilization assay with 1-N-phenylnapththylamine (NPN). Outer membrane permeability of NPN increases in the presence of AR-12 as seen with increase in fluorescence of NPN with increasing concentrations of AR-12. A. baumannii 19606 and K. pneumoniae 13883 are ATCC strains that are polymyxin sensitive. A. baumannii 34299 and K. pneumoniae 30835 are MDR clinical isolates that are polymyxin resistant. The experiments were repeated two or more independent times to verify results. |
Results from the in vitro assays suggested that the combination of AR-12 and colistin could potentially be useful for developing combination therapies for treating infections resulting from MDR pathogens. In order to determine if we could test the combination of AR-12 and colistin in animal efficacy studies, we conducted preliminary pharmacokinetic (PK) studies of AR-12 in mice for evaluation of plasma levels of AR-12 that can be achieved in mice. The maximum tolerated dose of AR-12 was found to be 100 mg/kg, p.o. in mice and a subsequent PK study in mice (80 mg/kg, p.o., q.d.) showed that plasma levels of 1.5–2.1 μg/mL could be sustained over a 24 hrs period (Cmax =2.1 μg/mL, Tmax =10 hrs, T1/2=22 hrs, AUC (0–24 hrs)=41.4 μg/mL*h) (Table S2 and Figure S1). However, this plasma concentration in mice is much lower than the concentration of AR-12 required for the antibacterial effect of the AR-12 combination with colistin in vitro (4 μg/mL). Furthermore, AR-12 was also found to be highly protein bound in mice (99%) making it impossible to achieve unbound plasma levels in mice that would be effective for the in vivo combination studies. Therefore it will be challenging to demonstrate the in vivo efficacy of the polymyxin/AR-12 combination due to the poor bioavailability of AR-12 in mice. Nevertheless, our in vitro results indicate that the combination of nonantibiotic small molecule and polymyxins will be an effective strategy to combat the rising issue of polymyxin resistance in bacteria. Furthermore, development of analogs of AR-12 that are more bioavailable could be highly valuable in screening of new AR-12 scaffolds and polymyxin combination against polymyxin-resistant MDR bacteria.
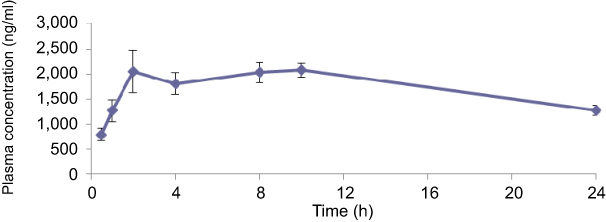 | Figure S1 Time-concentration profile of AR-12 in mice following a single PO dose. PK studies in mice indicate that plasma levels of ~2 μg/mL of AR-12 can be sustained over a 24 hrs period. |
Conclusion
The rapid emergence of resistance in polymyxin-sensitive strains is putting at risk our ability to treat common infections, particularly hospital-acquired infections, which are the sixth leading cause of deaths in the United States.21 Hence, novel therapies are needed to combat bacterial pathogens. The growing problem of AMR is a great concern in the military as well, where the major challenge is the elimination of MDR strains Acinetobacter, Klebsiella and Pseudomonas, the leading causative agents of infections in war wounds. The development of effective combination therapies for targeting nosocomial infections will also prove promising for treating troops wounded in wars and protecting the soldiers. The increasing threat of bacterial resistance to polymyxins has further accelerated the development of new approaches to solve this global health challenge. The use of nonantibiotic small molecules in combination with polymyxin is an attractive approach to counter bacterial resistance and make it more susceptible to polymyxins (Eg: Pentamidine). In our study, we have utilized AR-12 as a small molecule adjuvant of polymyxins against a panel of polymyxin sensitive and resistant strains. Using growth studies and broth dilution assays involving combination of polymyxins and AR-12, we have shown that the MIC of polymyxins can be greatly reduced for MDR A. baumannii and K. pneumoniae strains in the presence of AR-12. Further, time-kill assays using AR-12 and colistin indicate that there is a significant reduction in colony forming units for both MDR A. baumannii and K. pneumoniae strains, when treated with AR-12/colistin combination. Additionally, permeabilization studies of AR-12 using fluorescently labeled DP and NPN probes indicate that AR-12 might be facilitating the uptake of polymyxins in these strains, thereby enhancing the antibacterial activity.
Our studies show that the combination of AR-12 and colistin is effective only against certain Gram-negative bacterial strains that have acquired polymyxin resistance but not effective against strains that may have inherent polymyxin resistance. Novel small molecules based on the AR-12 scaffold will be valuable for use in combination with polymyxins as narrow spectrum antibacterial agents. To delineate the mechanism of action of AR-12, we are currently pursuing a systems biology approach (employing genomics, transcriptomics, lipidomics and metabolomics). To understand the nature of gene mutations and modifications that might be contributing to polymyxin resistance in the clinical isolates used in our study, we are performing whole genome sequencing of select clinical isolates used in this study to compare it with the genomes of polymyxin-sensitive isolates. The analysis and comparison of the genomes will facilitate the determination of mutations in genes known to be involved in polymyxin-resistant pathways or other biological pathways. We are further studying the effect of AR-12 on select clinical isolates by conducting transcriptomics studies to facilitate systematic analysis of the bacterial gene regulation and cellular functions when exposed to the AR-12/polymyxin combination. It is well established that modification of lipid A component of bacterial membrane can lead to polymyxin resistance. It will be highly valuable to conduct mass spectrometric analysis of lipid A of MDR isolates with and without treatment with AR-12/polymyxin combination to determine if AR-12 is modifying the lipid A component of the MDR isolates and thereby making them more susceptible to polymyxins. In parallel, we are also exploring the design and synthesis of new AR-12-based scaffolds that are more bioavailable in host. Identification of other nonantibiotic small molecules employing similar mechanism of action will prove highly valuable in developing treatment options with combination therapies against infections caused by MDR pathogens. Furthermore, a systematic study of the role of the bacterial polymyxin resistance genes and other cellular functions that result in acquisition of polymyxin resistance in Gram-negative bacteria will facilitate the discovery of small molecules that can be used for targeted combination therapies.
Acknowledgments
This work was supported by the Department of Defense Chemical Biological Defense Program through the Defense Threat Reduction Agency (DTRA) under United States Army Medical Research Institute of Infectious Diseases (USAMRIID) project number 13267645. Opinions, interpretations, conclusions and recommendations are those of the authors and are not necessarily endorsed by the US Army.
The authors would like to acknowledge JMI Laboratories, Iowa, for providing MDR A. baumannii and K. pneumoniae strains and CPT Steven D. Zumbrun (USAMRIID) for providing K. pneumoniae strains 101434, 101522 and 101783 (Table S1). The authors would like to thank Arno Therapeutics, Inc. for providing AR-12 to conduct the experiments described in this manuscript. The authors would further like to acknowledge Ms. Christy L. Thoma, Mr. Douglas J. Lane, Mr. Glenn Y. Gomba, Ms. Katlin M. Recabo and Dr. Jennifer M. Scarff for their assistance with MIC assays.
Disclosure
Dr. Malathy Krishnamurthy was supported by the National Research Council (NRC) Research Associateship Award at USAMRIID. Mr. Evan M Falcinelli was supported by Science and Engineering Apprentice Program (SEAP) at USAMRIID. Mr. Reuel A Sandy, Ms. Jennifer N Dootz and Dr. Tiffany M Mott were supported by the Oak Ridge Institute for Science and Education (ORISE) Fellowship. The authors report no other conflicts of interest in this work.
References
1. O’Neill J. Antimicrobial resistance: tackling a crisis for the health and wealth of nations. Rev Antimicrob Resist. 2014. Available from: https://amr-review.org/. Accessed April 23, 2019.
2. Mehrad B, Clark NM, Zhanel GG, Lynch JP
3. Al-Tawfiq JA, Laxminarayan R, Mendelson M. How should we respond to the emergence of plasmid-mediated colistin resistance in humans and animals? Int J Infect Dis. 2017;54:77–84. doi:10.1016/j.ijid.2016.11.415
4. Poirel L, Jayol A, Nordmann P. Polymyxins: antibacterial activity, susceptibility testing, and resistance mechanisms encoded by plasmids or chromosomes. Clin Microbiol Rev. 2017;30(2):557–596. doi:10.1128/CMR.00064-16
5. Olaitan AO, Morand S, Rolain JM. Mechanisms of polymyxin resistance: acquired and intrinsic resistance in bacteria. Front Microbiol. 2014;5:643. doi:10.3389/fmicb.2014.00547
6. Liu YY, Wang Y, Walsh TR, et al. Emergence of plasmid-mediated colistin resistance mechanism MCR-1 in animals and human beings in China: a microbiological and molecular biological study. Lancet Infect Dis. 2016;16(2):161–168. doi:10.1016/S1473-3099(15)00424-7
7. Ejim L, Farha MA, Falconer SB, et al. Combinations of antibiotics and nonantibiotic drugs enhance antimicrobial efficacy. Nat Chem Biol. 2011;7(6):348–350. doi:10.1038/nchembio.559
8. Worthington RJ, Melander C. Combination approaches to combat multidrug-resistant bacteria. Trends Biotechnol. 2013;31(3):177–184. doi:10.1016/j.tibtech.2012.12.006
9. Ball P. The clinical development and launch of amoxicillin/clavulanate for the treatment of a range of community-acquired infections. Int J Antimicrob Agents. 2007;30(Suppl 2):S113–S117. doi:10.1016/j.ijantimicag.2007.07.037
10. Annamanedi M, Varma GYN, Anuradha K, Kalle AM. Celecoxib enhances the efficacy of low-dose antibiotic treatment against polymicrobial sepsis in mice and clinical isolates of ESKAPE pathogens. Front Microbiol. 2017;8:805. doi:10.3389/fmicb.2017.00805
11. Stokes JM, MacNair CR, Ilyas B, et al. Pentamidine sensitizes gram-negative pathogens to antibiotics and overcomes acquired colistin resistance. Nat Microbiol. 2017;2:17028. doi:10.1038/nmicrobiol.2016.251
12. Harris TL, Worthington RJ, Hittle LE, Zurawski DV, Ernst RK, Melander C. Small molecule downregulation of PmrAB reverses lipid A modification and breaks colistin resistance. ACS Chem Biol. 2014;9(1):122–127. doi:10.1021/cb400490k
13. Zhu J, Huang JW, Tseng PH, et al. From the cyclooxygenase-2 inhibitor celecoxib to a novel class of 3-phosphoinositide-dependent protein kinase-1 inhibitors. Cancer Res. 2004;64(12):4309–4318. doi:10.1158/0008-5472.CAN-03-4063
14. Weng SC, Kashida Y, Kulp SK, et al. Sensitizing estrogen receptor-negative breast cancer cells to tamoxifen with OSU-03012, a novel celecoxib-derived phosphoinositide-dependent protein kinase-1/Akt signaling inhibitor. Mol Cancer Ther. 2008;7(4):800–808. doi:10.1158/1535-7163.MCT-07-0434
15. Chiu HC, Soni S, Kulp SK, et al. Eradication of intracellular Francisella tularensis in THP-1 human macrophages with a novel autophagy inducing agent. J Biomed Sci. 2009;16:110. doi:10.1186/1423-0127-16-110
16. Booth L, Robert JL, Cash DR, et al. GRP78/BiP/HSPA5/Dna K is a universal therapeutic target for human disease. J Cell Physiol. 2015;230(7):1661–1676.
17. Campos MA, Morey P, Bengoechea JA. Quinolones sensitize gram-negative bacteria to antimicrobial peptides. Antimicrob Agents Chemother. 2006;50(7):2361–2367. doi:10.1128/AAC.01437-05
18. Ortega X, Silipo A, Saldias MS, Bates CC, Molinaro A, Valvano MA. Biosynthesis and structure of the Burkholderia cenocepacia K56-2 lipopolysaccharide core oligosaccharide: truncation of the core oligosaccharide leads to increased binding and sensitivity to polymyxin B. J Biol Chem. 2009;284(32):21738–21751. doi:10.1074/jbc.M109.008532
19. Helander IM, Mattila-Sandholm T. Fluorometric assessment of gram-negative bacterial permeabilization. J Appl Microbiol. 2000;88(2):213–219.
20. Hancock REW, Wong PGW. Compounds which increase the permeability of the pseudomonas-aeruginosa outer-membrane. Antimicrob Agents Ch. 1984;26(1):48–52. doi:10.1128/AAC.26.1.48
21. Peleg AY, Hooper DC. Hospital-acquired infections due to gram-negative bacteria. N Engl J Med. 2010;362(19):1804–1813. doi:10.1056/NEJMra0904124
Supplementary materials
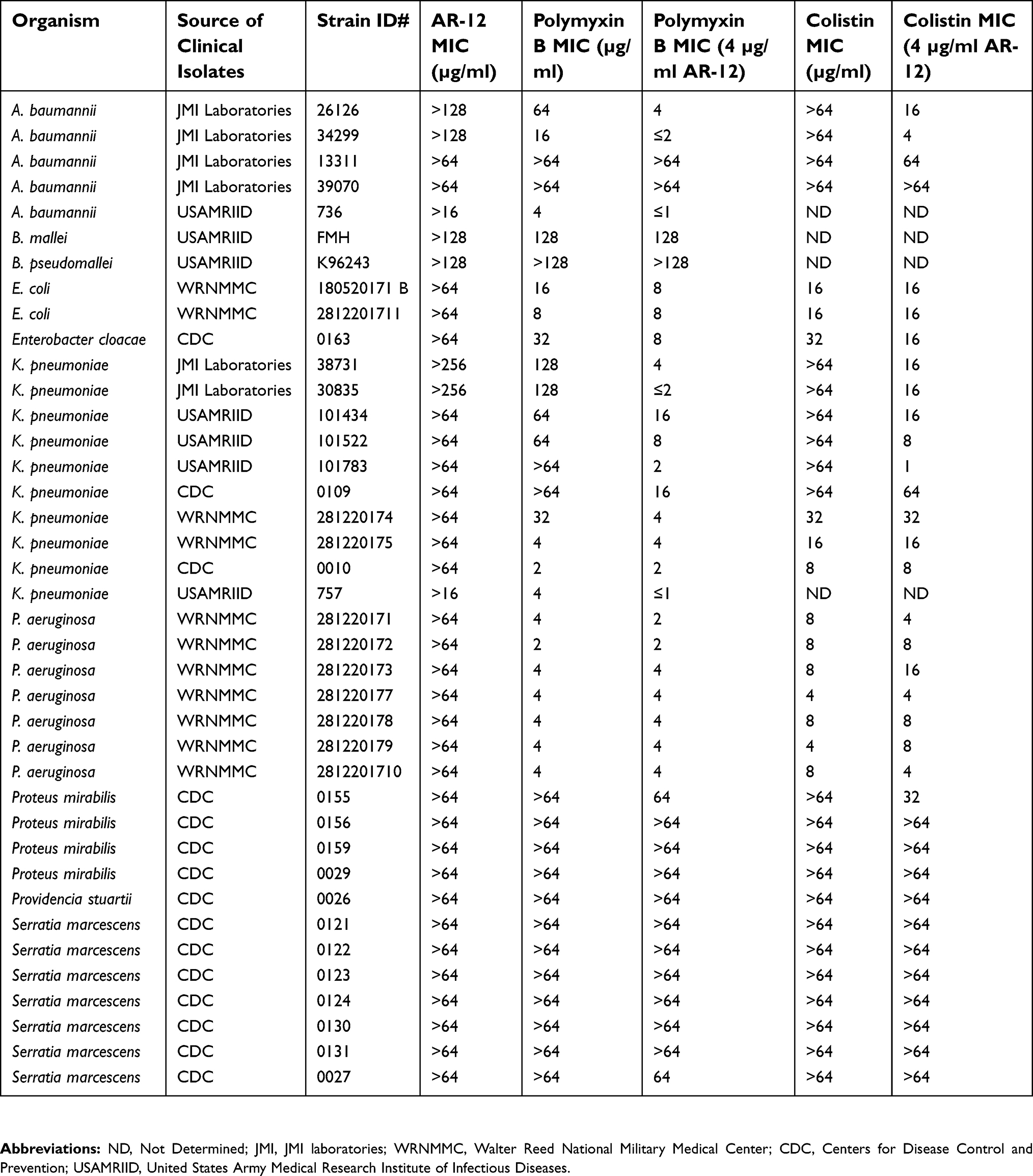 | Table S1 Effect of AR-12 on the antibacterial activity of polymyxin B and colistin in Gram-negative bacterial pathogens |
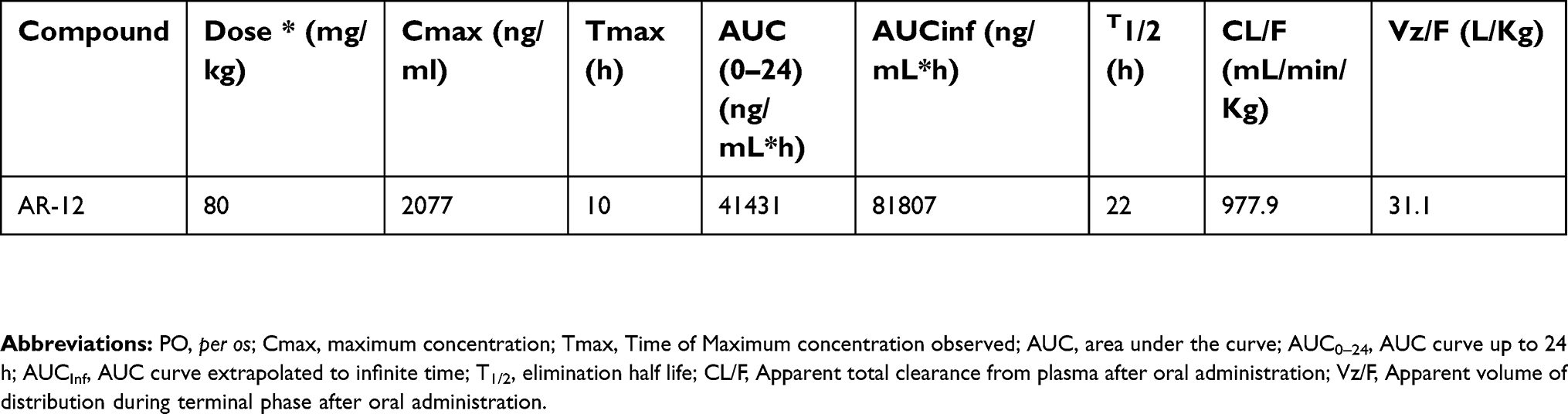 | Table S2 Pharmacokinetic (PK) parameters of AR-12 in female Balb/c mice after a single PO dose |
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.