Back to Journals » International Journal of Nanomedicine » Volume 18
Enhancing Immunity and Modulating Vaginal Microflora Against Candidal Vaginitis Through Nanoemulsion Supplemented with Porphyra Oligosaccharide as an Intravaginal Vaccine Adjuvant
Authors Tsai WC, Liu FL, Huang MH, Huang CH
Received 17 July 2023
Accepted for publication 27 October 2023
Published 6 November 2023 Volume 2023:18 Pages 6333—6346
DOI https://doi.org/10.2147/IJN.S431009
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Yan Shen
Wei-Chung Tsai,1 Fang-Ling Liu,1 Ming-Hsi Huang,2 Chung-Hsiung Huang1,3
1Department of Food Science, National Taiwan Ocean University, Keelung, Taiwan; 2National Institute of Infectious Diseases and Vaccinology, National Health Research Institutes, Miaoli, Taiwan; 3Center for Marine Bioscience and Biotechnology, National Taiwan Ocean University, Keelung, Taiwan
Correspondence: Chung-Hsiung Huang, Department of Food Science, National Taiwan Ocean University, Keelung, Taiwan, Tel +886-2-2462-2192 ext 5116, Email [email protected]
Background: Intravaginal vaccination is an encouraging approach to prevent infectious vaginitis, with nanoemulsions showing effectiveness as mucosal adjuvants.
Purpose: This study aimed to formulate a nanoemulsion incorporating Porphyra oligosaccharide (PO@NE) and assess its effectiveness as a mucosal adjuvant in intravaginal vaccines against candidal vaginitis.
Materials and Methods: PO@NE was prepared, and the stability, immunomodulatory activity and cytotoxicity were screened in vitro. Further, the preventive effect of PO@NE as adjuvants for heat-killed Candida albicans (HK-CA) vaccines was explored in a murine model of candidal vaginitis, in comparison with those supplemented with polysaccharide (PP@NE). The mice were intravaginally vaccinated with 106 HK-CA cells, suspended in 1% NE without or with either PO or PP at a final concentration of 6.5 μg/mL, in a total volume of 20 μL. This vaccination was intravaginally administered once a week for 3 weeks. One week following the final vaccination, the mice underwent an intravaginal challenge with 107 C. albicans cells. One week after the challenge, the mice were euthanized to isolate serum, spleen, vaginal washes, and vaginal tissues for analysis.
Results: PP@NE and PO@NE, with diameters approximately around 100 nm, exhibited exceptional stability at 4°C and low cytotoxicity when used at a concentration of 1% (v/v). Intravaginal vaccination with HK-CA adjuvanted with PO@NE effectively protected against candidal vaginitis evidenced by less Candida hyphae colonization, milder mucosal damage and cell infiltration. Moreover, enhanced mucosal antibody production, induction of T helper (Th)1 and Th17-related immune responses, enlarged the population of CD8+ cells, and elevated vaginal microflora diversity were observed in vaccinated mice. Interestingly, the potency was rather attenuated when PO@NE was replaced with PP@NE.
Conclusion: These findings indicate PO@NE as a HK-CA vaccine adjuvant for candidal vaginitis prevention via enhancement of both cellular and humoral immunity and modulation of vaginal microflora, emphasizing further intravaginal vaccination development.
Keywords: Candida albicans, mucosal vaccine adjuvant, nanoemulsion, Porphyra oligosaccharide, vaginitis
Introduction
Candida albicans commonly coexists as an opportunistic pathogen in the oral cavity, gastrointestinal tract and genitourinary tract. In an immunocompromised host, C. albicans could cause mucosal infections, such as oral and vaginal infection, and even systemic infection.1 About 50,000 cases of Candida systemic infection occur in the United States per year, and the fatality rate is as high as 50%.2 Most of the conventional treatments for C. albicans infection are oral or topical administration of antifungal azole drugs.3 However, fluconazole-resistant C. albicans strains have been isolated and identified.4 Normally, the protective function of the mucosal epithelium involves creating a physical barrier to hinder the entry of Candida. However, Candida has the ability to breach this barrier either through triggering endocytosis or actively invading the epithelial cells. In response to the presence of Candida hyphae, the epithelial cells release cytokines to attract phagocytes to phagocytize and eliminate Candida. Concurrently, pro-inflammatory cytokines, such as TNF-α and IL-6, produced by macrophages and epithelial cells trigger the recruitment and infiltration of neutrophils. Dendritic cells also play a role by migrating to the lymph nodes and contributing to the development of adaptive T helper (Th) cell responses. Th17 cells are particularly crucial in defending against Candida in mucosal tissues by producing IL-17, which recruits and activates neutrophils. Additionally, Th1 cells release IFN-γ, which strongly activates phagocytic cells.5
Vaccination activates the immune system to elicit effective adaptive immune response by simulating the infection process.6 During the reaction, some immune cells are converted into effector cells to induce antigen-specific immune responses against pathogens.7 Most of the current vaccination induces systemic immune responses through subcutaneous or intramuscular injections, and it is difficult to trigger antigen-specific immune responses at mucosal tissues.8 Nevertheless, it is important to note that the mucosal tissues encompassing the oral cavity, respiratory tract, gastrointestinal tract, and genitourinary tract serve as a significant entry for infectious pathogens into the body due to their extensive surface area.9 Moreover, approximately 80% of immune cells are in the mucosa-associated lymphoid tissue, indicating that the mucosa is an ideal route for vaccination.10 Compared to the other mucosal tissues, especially the gastrointestinal mucosa, vaginal mucosa is a promising route for vaccination due to its relatively stable pH, low enzymatic activity, and large surface area.11 As an antigen delivery system, emulsion adjuvant has been proven to be safe and well tolerated.12 We recently demonstrated that intravaginally sensitized with ovalbumin formulated with nanoemulsion as a mucosal adjuvant effectively reinforced mucosal and immunological fingerprints.13
Porphyra sp., a type of red seaweed commonly found in East Asia, holds economic value as a crop and food source. It is rich in various bioactive components, especially polysaccharides primarily composed of galactose, 3,6-anhydrogalactose, and sulfate groups, constituting approximately 20–40% of the dry weight of Porphyra.14,15 Several studies have explored the bioactivities of Porphyra polysaccharides. Specifically, recent research has highlighted the modulatory effects of Porphyra polysaccharides on immune cell activation.16,17 Furthermore, polysaccharides extracted from P. haitanensis have shown potential in alleviating allergic responses in mice.18 Notably, oligosaccharides hydrolyzed from Porphyra polysaccharides have also demonstrated bioactive properties. Osumi et al utilized an enzyme produced by Arthrobacter sp. S-22 to hydrolyze Porphyra polysaccharides from P. yezoensis, resulting in oligosaccharides with the ability to reduce cholesterol levels in rats.19 Similarly, an enzyme solution derived from Aeromonas salmonicida MAEF108 was employed to hydrolyze algal polysaccharides, yielding oligosaccharides with antiviral and prebiotic activity.20 In a recent study, oral administration of Porphyra polysaccharides (PP) and oligosaccharides (PO) could modulate the immune response specific to ovalbumin in mice sensitized with ovalbumin.21 However, the adjuvanticity of PP and PO is unclear. In this study, PP- and PO-containing nanoemulsions, named as PP@NE and PO@NE, were prepared, and their physical property and storage stability were determined. Furthermore, these nanoemulsions were utilized as mucosal adjuvants and to be mixed with heat-killed C. albicans (HK-CA) for the preparation of C. albicans vaccines. The protective efficacy of these vaccines was investigated in a murine model of vaginal C. albicans infection, and the immunological fingerprints and profiles of vaginal microflora were investigated to comprehensively understand their action mechanisms.
Materials and Methods
Chemicals, Reagents, Detection Kits, Antibodies, C. albicans and Porphyra sp
Chemicals and reagents utilized in the study were obtained from Sigma Chemical (St. Louis, MO, USA) unless specified otherwise. Cell culture reagents were purchased from GE Healthcare Life Sciences (Marlborough, MA, USA). ELISA kits for measuring cytokines and immunoglobulins (Igs) were procured from eBioscience, Inc. (San Diego, CA, USA). The C. albicans strain (ATCC 10231) was kindly provided by Prof. Shiow-Yi Chen of National Taiwan Ocean University. Anti-Mouse CD4-PE and anti-mouse CD8-APC antibodies were obtained from Thermo Fisher Scientific Inc. (Waltham, MA, USA). Dried Porphyra sp. was acquired from Xin Meng Cheng company (Penghu, Taiwan) and ground into a powder using an electric grinder (RT-02, Mill Powder Tech Solutions, Tainan City, Taiwan). Prior to use, the powder was filtered through a 60 mesh (0.25 mm) sieve.
Preparation and Characterization of Porphyra Polysaccharide (PP) and Oligosaccharide (PO)
The process for preparing PP and PO followed the method described in a previous study.21 However, in this study, a commercial enzyme called Cellulase AP3 (2000 U/g; Amano Enzyme, Aichi, Japan) was used for the hydrolysis of PP. Specifically, 1% PP were hydrolyzed at 37°C and pH 5.4 for 48 h using Cellulase AP3. The molecular weights and functional groups of PP and PO were determined using high-performance liquid chromatography (HPLC) analysis as described in the previous study.21 PP with a molecular weight greater than 10 kDa was collected by an ultrafiltration system, and the molecular weight was approximately 300 kDa (Figure S1A). After enzymatic hydrolysis, PO with a molecular weight less than 3 kDa was collected by an ultrafiltration system, and the molecular weight was approximately 1 kDa (Figure S1B).
Preparation of PP- and PO-Containing Emulsions and Nanoemulsions
The method for emulsions and nanoemulsions preparation was according to the methods reported in the previous study with minor modifications.13 In a typical procedure, 400 μL of PO solution (1.5 mg/mL in aqueous), 470 μL of docosahexaenoic acid, 60 μL of Tween 80® and 85 μL of Span 85® were homogenized for 3 min (23,000 rpm, 4°C) and then sonicated for 30 min (43 KHz, 4°C). Subsequently, the homogeneous mixture was diluted with 100-fold volume of phosphate buffered saline (PBS) to obtain PO-containing emulsion (PO@EM). The emulsion was further passed through a 0.22 µm pore size of sterile filter for 11 times to harvest nanoemulsion (PO@NE). PO was replaced with the same volume of PP to yield PP@EM and PP@NE, respectively. For the purposes of comparison, blank emulsion and nanoemulsion, EM and NE, were prepared in the absence of PP or PO under the same procedure.
The morphology of nanoemulsions were observed by using liquid transmission electron microscope (K-kit; Bio Materials Analysis Technology Inc., Hsinchu, Taiwan). Emulsion and nanoemulsion samples were stored at 4°C and 25°C. The dynamic light scattering (DLS) technique, using a Zetasizer Nano ZS instrument (Malvern Instruments Ltd., Worcestershire, UK), was employed to determine the droplet size, polydispersity index (PDI), and zeta potential of the samples. Measurements of droplet size, PDI, and zeta potential were conducted once a week for a duration of 8 weeks. The freeze-thaw stability testing method followed the procedure described by Esmaeili et al.22 Specifically, 2 mL of emulsions or nanoemulsions were stored at −20°C for 48 h, followed by storage at 25°C for an additional 48 h, constituting one freeze-thaw cycle. The droplet size, PDI, and interface potential of the emulsions and nanoemulsions were measured after each cycle, with a total of 1–3 freeze-thaw cycles being performed.
Preparation of C. albicans Vaccines
C. albicans was inoculated into liquid yeast growth medium (YPD broth, Gibco, Thermo Fisher Scientific Inc., NY, USA) and cultured at 37°C for 24 h. Following centrifugation at 2000 × g for 5 min, the supernatant was carefully decanted, and the cell pellet was washed using sterile PBS. Subsequently, the pellet was resuspended in PBS to a density of 108 cells/mL. The suspensions were then subjected to a boiling water bath for 45 min to generate heat-killed C. albicans (HK-CA). To prepare C. albicans vaccines, the HK-CA pellet (108 cells) was mixed with PBS, emulsions or nanoemulsions to reach a total volume of 1 mL. This mixing process was conducted using a test-tube rotator set at 5 rpm for a minimum duration of 1 h prior to vaccination. The formula of vaccines included HK-CA alone (Control group, C), HK-CA mixed with emulsion (Em), HK-CA mixed with emulsion containing PP (EmP), HK-CA mixed with emulsion containing PO (EmO), HK-CA mixed with nanoemulsion (NEm), HK-CA mixed with nanoemulsion containing PP (NEmP), and HK-CA mixed with nanoemulsion containing PO (NEmO).
In vitro Assessment of Cytotoxicity and Immunomodulatory Activity of C. albicans Vaccines
The spleen samples were isolated from normal BALB/c mice and prepared as splenocyte suspensions. Briefly, the isolated spleen samples were washed three times with sterile RPMI-1640 medium, ground using a cell strainer, and then centrifuged at 11.29 × g and 4°C for 5 min. After removing the supernatant, red blood cell lysis buffer was added and left for 90s. Following this, ten times the volume of RPMI-1640 medium was added to halt the lysis process. After another round of centrifugation, the supernatant was discarded. The final concentration of splenocytes was adjusted to 5×106 cells/mL.23 Splenocytes were cultured with or without C. albicans vaccines individually for 48 h. The concentrations of HK-CA vaccines tested in the in vitro assays were 106 cells/mL of HK-CA with PBS or with 1% (v/v) of the emulsions or nanoemulsions. This concentration was also the highest one that could be prepared for the in vitro experiment. Upon completion of centrifugation at 11.29 × g and 4°C for 5 min, the supernatants were carefully collected for cytokine measurement. The viability of cells was assessed using the MTT assay.24 The levels of IFN-γ, IL-6, IL-17A, and TNF-α in the supernatants of cultured splenocytes were quantified using ELISA, following the instructions provided by the supplier.24
Animal Experiments
Female BALB/c mice (5 weeks old) were procured from the National Laboratory Animal Center of Taiwan, ensuring that they were specific pathogen-free. The mice were housed at the laboratory animal facility of the National Taiwan Ocean University (NTOU) and allowed unlimited access to food and water for a period of 1 week prior to the start of the experiments. The employed murine model of vaginal Candida infection was according to that reported in the previous study.25 The mice were divided into naïve group (NA; intravaginally administrated with 20 μL PBS), control group (C; intravaginally vaccinated with HK-CA), NEm group (vaccinated HK-CA adjuvanted with NE), NEmP group (vaccinated with HK-CA adjuvanted with PP@NE) and NEmO (vaccinated with HK-CA adjuvanted with PO@NE). Before intravaginal administration, the mice were anesthetized with a steady concentration of 2.5% isoflurane. Oxygen, delivered at a pressure of 4 kg/cm² through the XGI-8 Anesthesia System, was used in conjunction with isoflurane. The intravaginal administration of C. albicans suspension or HK-CA vaccines was carried out by inserting the pipette tip approximately 5 mm deep into the vaginal lumen, using a total volume of 20 μL. Each vaccine dose contained 106 HK-CA cells suspended in PBS or 1% nanoemulsions, without or with the addition of either PO or PP to achieve a final concentration of 6.5 μg/mL, in a total volume of 20 μL. The mice were intravaginally vaccinated once a week for 3 weeks. Five days after the last vaccination, the mice were intraperitoneally with 0.2 mg β-estradiol in 0.1 mL sesame oil. Except for the mice in NA group, the mice in the other groups were intravaginally challenged with 107 cells of C. albicans 7 days after the last vaccination. Seven days after the challenge, the mice were euthanized to isolate serum, spleen, vaginal washes and vaginal tissues. To ensure a humane conclusion, the mice were first anesthetized as described earlier and then euthanized through cervical dislocation. Vaginal tissues were prepared for Periodic Acid-Schiff stain (PAS)-stained sections for histological examination. PAS dye could react with aldehyde groups in the fungal cell wall, resulting in pink signals of Candida hyphae.26 Moreover, total genomic DNA from vaginal tissues was extracted for the full-length 16S genes (V1-V9 regions) sequencing analysis as described previously.27,28
Measurement of Splenic Cytokine Production, Cell Population and Immunoglobulin Concentration
The samples of spleen were isolated individually from mice to prepare splenocytes. The splenocyte suspensions (5 × 106 cells/mL) were cultured with heat-treated C. albicans (HK-CA; 2.5×105 cells/mL) for 48 h. Following centrifugation, the liquid above the cell pellet was gathered to assess cytokine levels. The concentrations of serum IgG, vaginal wash IgA, as well as IL-6, TNF-α, IFN-γ, and IL-17 in splenocyte supernatants were determined using an ELISA method in accordance with the provided guidelines. To gain further insights into the phenotype of splenic T cells, the cells was stained with PE-conjugated anti-CD4 and APC-conjugated anti-CD8 antibodies followed by analysis using flow cytometry (Attune® NxT ThermoFisher Scientific).
Statistical Analysis
Comparison between groups were conducted by use of ANOVA followed by Fisher’s least significant difference (LSD) method, and comparison between the treatment group and the control group was conducted by use of a Student’s t-test (SigmaPlot V14, Systat Software Inc.). The differences were considered significant at p < 0.05.
Results
In vitro Cytotoxicity and Immunomodulatory Activities of Emulsions and Nanoemulsions
To investigate the safety and application potential of emulsions and nanoemulsions as adjuvants for C. albicans vaccine, splenocytes were cultured with HK-CA formulated with or without various emulsions or nanoemulsions. Compared to untreated splenocytes, HK-CA stimulation obviously elicited TNF-α, IL-6, IFN-γ and IL-17A production without significantly altering cell viability (Figure 1A–E). However, treatment of HK-CA formulated with emulsions, including Em, EmP and EmO, markedly reduced cell viability and cytokine production compared to that of HK-CA alone, indicating the cytotoxic and immunosuppressive properties of emulsions (Figure 1A–E). In contrast, HK-CA formulated with nanoemulsions limitedly influenced cell viability, and NEmO even raised IFN-γ production compared to that of HK-CA alone (Figure 1A and D). These findings reveal that strict control of the nanoemulsion size is crucial for mitigating cytotoxicity. Therefore, nanoemulsions were employed for further experiments.
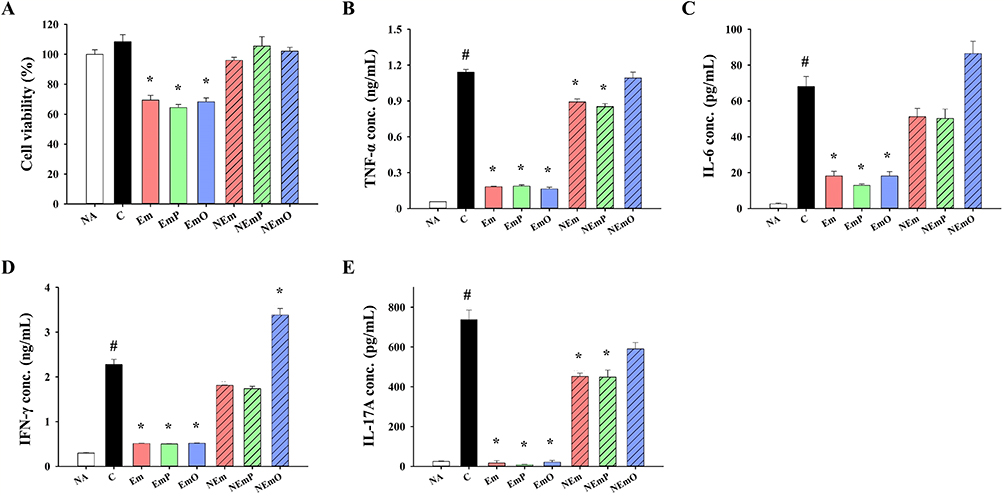 |
Figure 1 Immunostimulatory activity of vaccine formulations on splenocytes viability and cytokine production. HK-CA vaccines were prepared as described in the Materials and method. Primary splenocytes from normal mice were cultured with or without HK-CA vaccines individually for 48 h. (A) Cell viability was determined by MTT assay. The concentrations of (B) TNF-α, (C) IL-6, (D) IFN-γ and (E) IL-17A in supernatants of cultured splenocytes were measured by ELISA. The data are presented as mean ± SEM (n = 3) and represent a representative set of two independent experiments. Statistical significance was denoted as #p < 0.05 compared to the NA group and *p < 0.05 compared to the C group. |
Physical Property and Storage Stability of Nanoemulsions
The morphology of nanoemulsions were sphere with the droplet size between 90–110 nm (Figure 2). Notably, the PDI values (0.177 ± 0.022 and 0.179 ± 0.017) and zeta potentials (−13.23 ± 0.81 mV and −11.37 ± 0.79 mV) of PP@NE and PO@NE were lower than those of NE (0.207 ± 0.009, −7.68 ± 0.47 mV), indicating that PP and PO, with sulfate groups, adsorbed on nanoemulsion are beneficial for nanoemulsion droplets dispersion in the liquid phase (Figure 2).
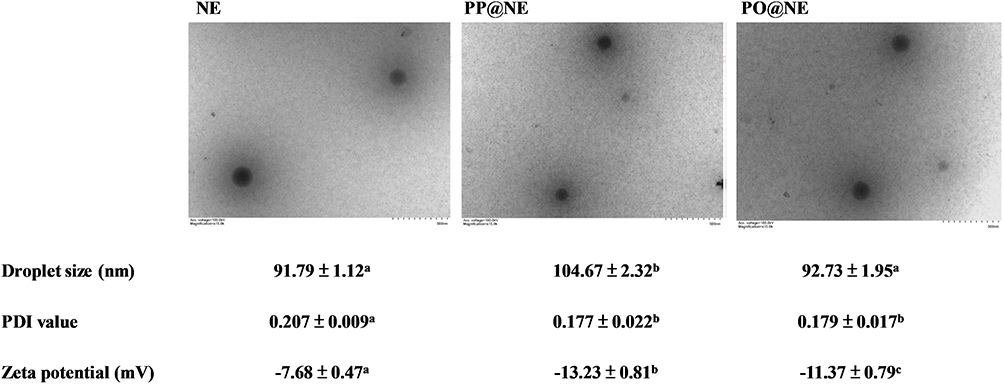 |
Figure 2 Morphology and physical properties of nanoemulsions. NE, PP@NE and PO@NE were prepared as described in the Materials and methods. Representative images of liquid transmission electron microscope are shown. Droplet sizes, PDI values and zeta potentials of nanoemulsions were analyzed by DLS. The data are presented as mean ± SEM (n = 3) and represent a representative set of two independent experiments. Values in each test parameter without a common letter represent a significant difference (p < 0.05). |
In terms of storage stability, the droplet size of NE decreased from approximately 90 nm to about 50 nm within one week. Similarly, the droplet size of PP@NE and PO@NE decreased from 90 − 100 nm to 50 − 60 nm between the first and second weeks at 25°C. Concurrently, the PDI values of the nanoemulsions increased from around 0.2 to 0.3 − 0.4. Interestingly, the droplet size, PDI value and zeta potential of nanoemulsions were not significantly altered after storage for 3 weeks (Figure 3A). On the other hand, the droplet size (around 90 − 110 nm), PDI value (approximately 0.2), and zeta potential (between −7 and −13 mV) of the nanoemulsions remained stable at 4°C for up to 6 weeks. However, after 6 weeks of storage, a slight decrease in droplet size (to 50 − 60 nm) and an increase in PDI value (to 0.3 − 0.4) were observed (Figure 3B). Furthermore, the droplet size (around 90 − 110 nm) and droplet quantity of the nanoemulsions remained unchanged after undergoing 3 cycles of freeze-thaw (Figure 3C), indicating 4°C is an ideal temperature for the storage of nanoemulsions.
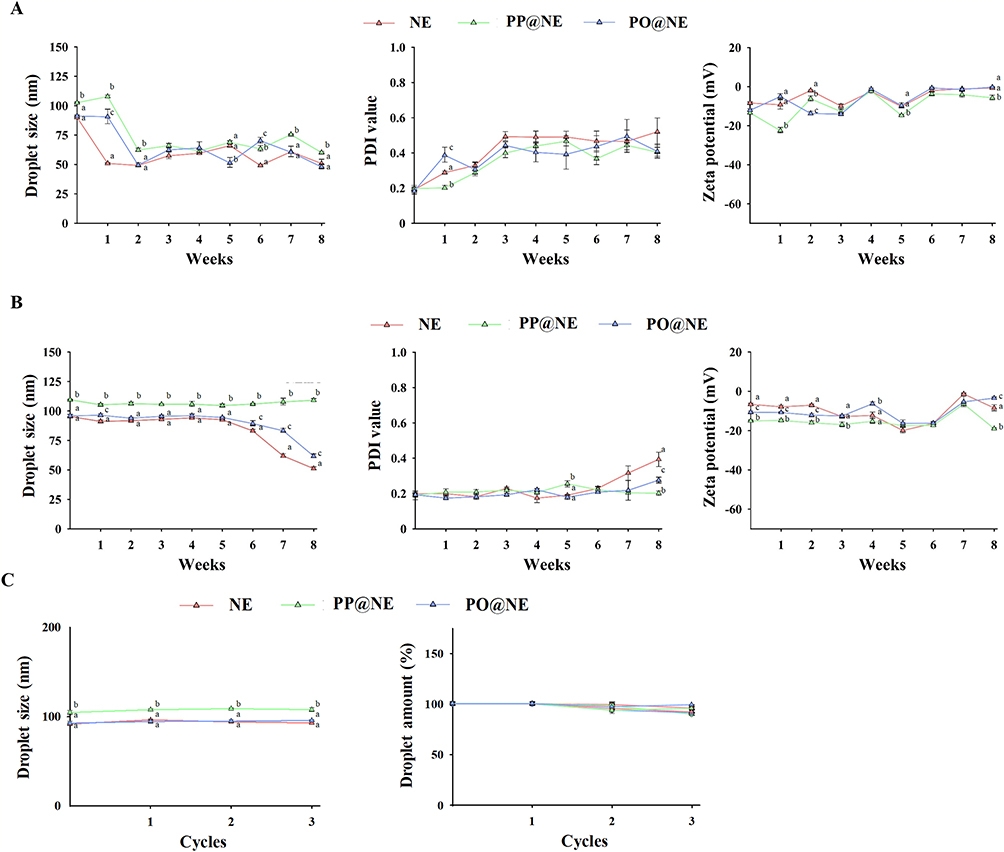 |
Figure 3 The stability of nanoemulsions under storage and freeze-thaw cycles. NE, PP@NE and PO@NE were prepared as described in the Materials and methods. The droplet sizes, PDI values and zeta potentials of nanoemulsions were measured once a week for 8 weeks at the storage temperatures of (A) 25°C and (B) 4°C. (C) In the freeze-thaw test, the droplet sizes, PDI values and zeta potentials were measured for 3 freeze-thaw cycles. The data are presented as mean ± SEM (n = 3) and represent a representative set of two independent experiments. Values in each test parameter without a common letter represent a significant difference (p < 0.05). |
Preventive Effect of Nanoemulsion-Based HK-CA Vaccines Against Candidal Vaginitis
The in vivo preventive effect of nanoemulsion-adjuvanted HK-CA vaccines on vaginal C. albicans infection was investigated using a murine model of candidal vaginitis (Figure 4A). The employed formulation and concentration of nanoemulsion-adjuvanted HK-CA vaccines (1% v/v with 106 cells/mL of HK-CA) was the same as that employed in the in vitro study showing no cytotoxicity. The body weights of all mice were stably increased, and no obvious clinical signs and abnormal activity were observed throughout the experiment, revealing the vaccines did not induce adverse effects (Figure 4B). Compared with that in the NA group, obvious epithelial damage and cell infiltration were observed at the vaginal mucosa of mice in C and NEm groups (Figure 5A). Moreover, a large amount of Candida hyphae could be observed at the vaginal mucosa of mice in the C group (Figure 5B). However, milder severity of histopathological change and limited Candida hypha were observed in mice of NEmO group. The quantitative data show that intravaginal vaccination with NEmP- and NEmO-adjuvanted HK-CA significantly reversed C. albicans-induced decrement of vaginal epithelial thickness and increment of cell infiltration (Figure 5C and D), indicating the prevention against C. albicans infection. As vaccination provides active immunity, the profile of adaptive immunity was further explored. The concentration of serum IgG was comparable between each group, and that of vaginal IgA was markedly elevated in mice of NEmO group (Figure 6A and B), revealing that elicitation of mucosal IgA production is one of the action mechanism of NEmO vaccination. Since Th17 and Th1 cells play a critical role in in mucosal defense against Candida, cell viability, IL-17A and IFN-γ production of splenocytes stimulated by HK-CA were measured. Intravaginal vaccination with NEm, NEmP and NEmO, but not HK-CA alone, significantly increased viability of splenocytes, indicating nanoemulsion is a promising adjuvant for intravaginal vaccination of HK-CA to induce systemic lymphocyte responses (Figure 6C). Compared to the C group, NEmO limitedly influenced IL-17A but augmented IFN-γ production (Figure 6D and E). Furthermore, the population of CD4+ and CD8+ splenocytes was analyzed to understand the profile of Th cells and cytotoxic T cells. Consistently, intravaginal vaccination with NEm, NEmP and NEmO, but not HK-CA alone, increased the population of CD4+ cells (Figure 6F). Restoration of depleted CD8+ cell was only observed in the NEmO group (Figure 6G). To further understand the impact of NEmO on the healthy mucosa, histopathological examination of vaginal tissues from mice intravaginally vaccinated with NEmO without a C. albicans challenge was conducted. However, no obvious histopathological changes were observed (Figure S2).
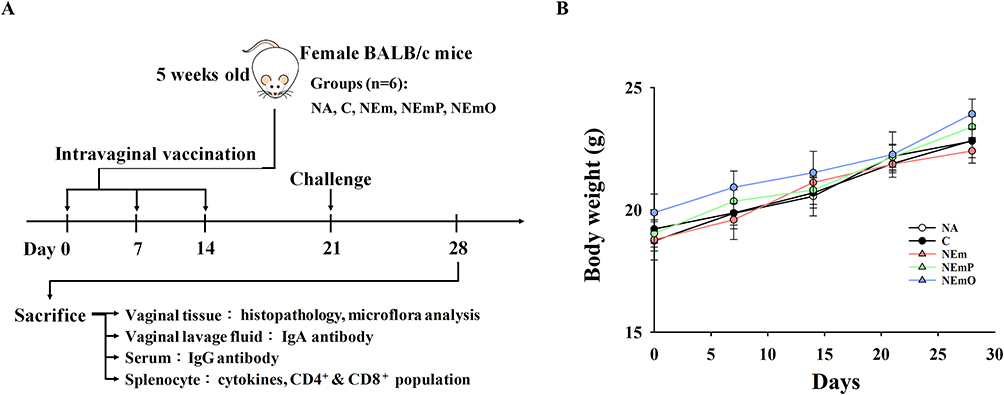 |
Figure 4 Vaccine efficacy of heat-killed C. albicans (HK-CA) intravaginal vaccines. (A) Schematic representation of vaccination protocol. BALB/c mice were divided into naïve group (NA; intravaginally administrated with 20 μL PBS), control group (C; intravaginally vaccinated with HK-CA), NEm group (vaccinated HK-CA adjuvanted with NE), NEmP group (vaccinated with HK-CA adjuvanted with PP@NE) and NEmO (vaccinated with HK-CA adjuvanted with PO@NE). Each vaccine dose contained 106 HK-CA cells suspended in PBS or 1% nanoemulsions, without or with the addition of either PO or PP to achieve a final concentration of 6.5 μg/mL, in a total volume of 20 μL. The mice were intravaginally vaccinated once a week for 3 weeks. 5 days after the last vaccination, the mice were intraperitoneally with 0.2 mg β-estradiol in 0.1 mL sesame oil. With the exception of the mice in the NA group, mice in all other groups were subjected to intravaginal challenge with 107 cells of C. albicans 7 days following the final vaccination. Seven days after the challenge, the mice were euthanized to collect serum, vaginal wash, vaginal, and spleen tissues for subsequent experiments. (B) Body weight change after C. albicans challenge. Body weight of the mice was individually measured and recorded on a weekly basis. The data are presented as mean ± SEM (n = 6) and represent a representative set of two independent experiments. |
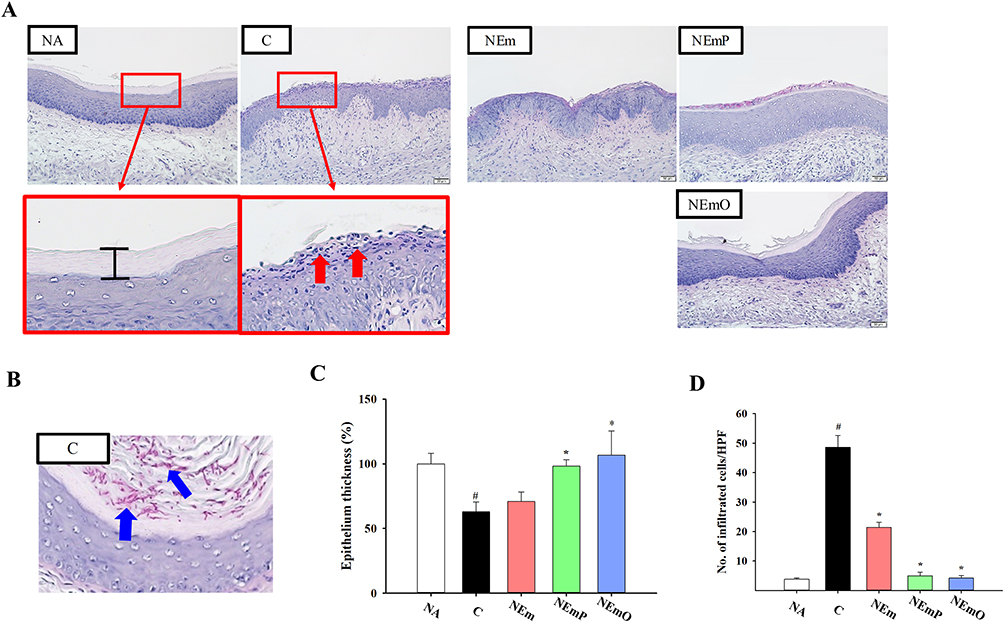 |
Figure 5 Histopathological examination of vaginal tissues. (A) Representative PAS-stained sections are shown. Red arrows indicate mucosal erosion and cell infiltration. (B) Representative photo of colonization of Candida hyphae at the vaginal mucosa, and blue arrows indicate Candida hyphae. Quantification of (C) relative epithelial thickness and (D) number of infiltrated cells per high power field (HPF) were calculated by ImageJ software. The data are presented as mean ± SEM (n = 6) and represent a representative set of three independent experiments. Statistical significance was denoted as #p < 0.05 compared to the NA group and *p < 0.05 compared to the C group. |
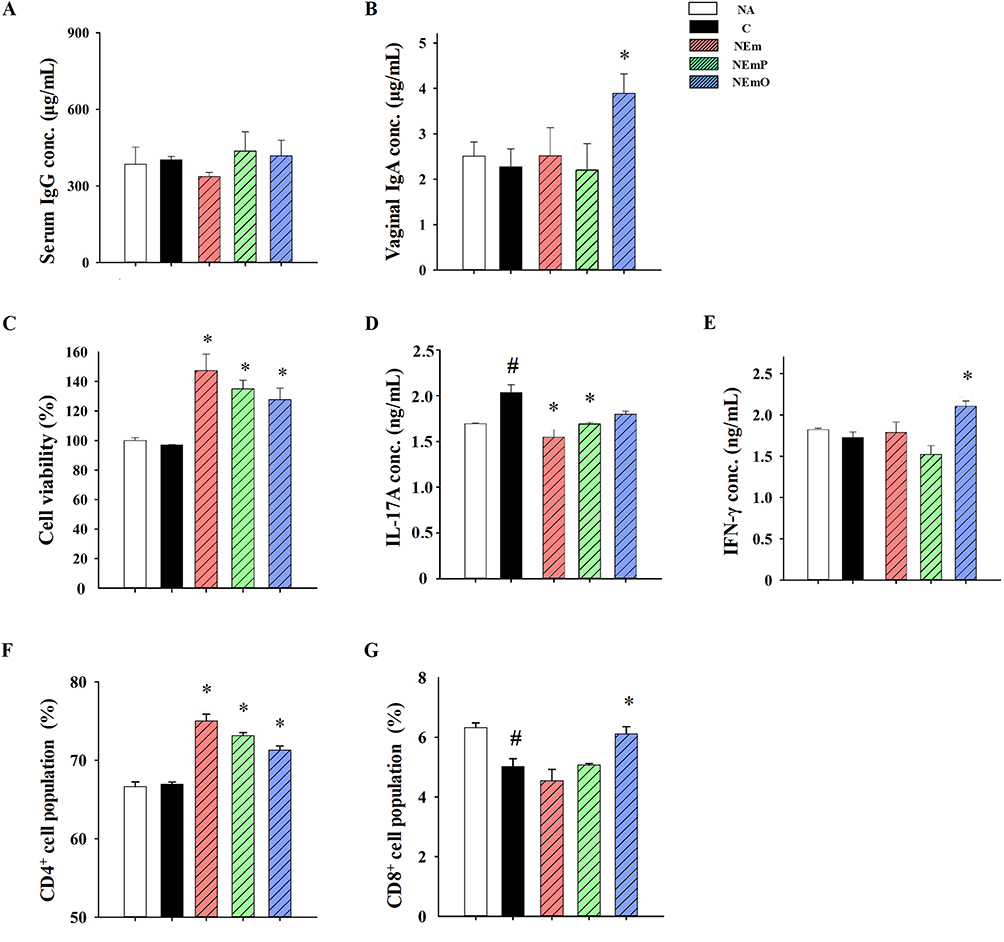 |
Figure 6 Antibody, cytokine production and splenic T cell populations in vaccinated and C. albicans-challenged mice. After euthanasia, the samples of serum, vaginal washes and spleens were collected individually. The concentrations of (A) IgG in serum and (B) IgA in vaginal washes were determined by ELISA. The prepared splenocytes were cultured with HK-CA for 48 h. (C) Cell viability of cultured splenocytes was measured by MTT assay, and the concentrations of (D) IL-17A and (E) IFN-γ in the supernatants of cultured splenocytes were determined by ELISA. The populations of (F) CD4+ and (G) CD8+ cells in splenocytes were measured by flow cytometry. The data are presented as mean ± SEM (n = 6) and represent a representative set of three independent experiments. Statistical significance was denoted as #p < 0.05 compared to the NA group and *p < 0.05 compared to the C group. |
Modulatory Effect of Nanoemulsion-Based HK-CA Vaccines on Vaginal Microflora
The impact of mucosal microflora on immunity has been documented, and natural polysaccharides and oligosaccharides have the potential to act as prebiotics, influencing the composition of the mucosal microflora.29,30 Therefore, the vaginal microflora profile was further analyzed using full-length 16S rRNA gene sequencing. The Bray-Curtis dissimilarity index is commonly employed as a standardized measure to assess the similarity or dissimilarity between two microflora samples.31 Bray-Curtis dissimilarity of C group was the lowest, and that of NEmO was the highest among the groups (Figure 7A), revealing that intravaginal vaccination of NEmO raised the beta diversity of vaginal microflora. Furthermore, the composition of vaginal microflora was analyzed. Interestingly, obvious relative abundance of Actinobacteria was observed only in the NEmO group (Figure 7B), which may explain why the markedly increased beta diversity of vaginal microflora was observed in the NEmO group.
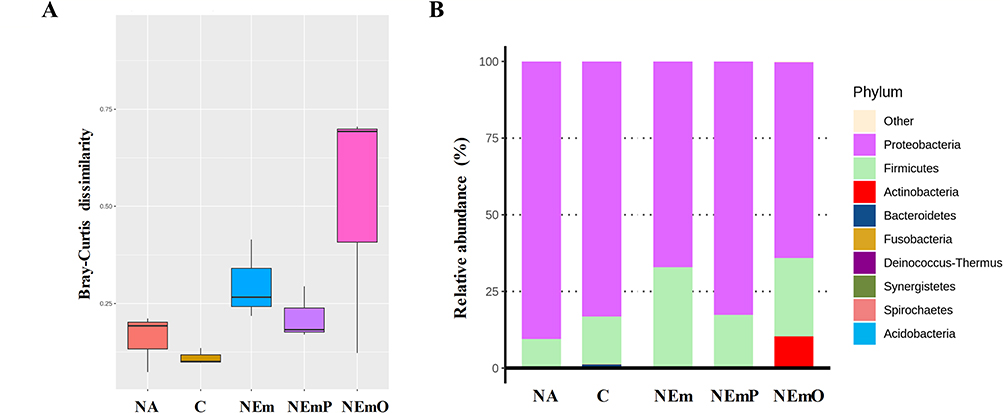 |
Figure 7 Full-length 16S rRNA gene amplicon analysis of vaginal microflora. After euthanasia, the fresh sample of vaginal tissues from each mouse were collected, and DNA of the samples were extracted for full-length 16S rRNA gene by NGS analysis. The results of (A) Bray-Curtis dissimilarity and (B) phylum-level composition, are shown to understand the composition, richness and abundance of vaginal microflora between each group. |
Discussion
A previous study reported that dried Porphyra powder contained 12.79% moisture, 55.89% carbohydrates, 24.61% crude protein, 0.25% crude fat, and 6.46% ash.14 In this study, the employed dry Porphyra powder had 10% moisture, 59.78% carbohydrates, 22.86% crude protein, 0.18% crude fat, and 7.06% ash. Gong et al have extracted Porphyra using 80°C water and obtained polysaccharides with molecular weights of 329 kDa, 203 kDa, 128 kDa, and 10 kDa.32 Isaka et al have extracted Porphyra using aqueous ethanol, obtaining polysaccharide with a molecular weight of 224 kDa.33 It is important to note that the extraction method can affect the molecular weight of the polysaccharides. In this study, a similar extraction method to that reported by Gong et al was used, resulting in a molecular weight of approximately 300 kDa. Furthermore, Hou et al have prepared Porphyra oligosaccharides by hydrolyzing 2.5% (w/v) Porphyra polysaccharides with 0.5M sulfuric acid at 80°C for 2 h. The average molecular weight of the obtained Porphyra oligosaccharide was 1.43 kDa.34 It has been reported that the modulatory effects of PP and PO on the immune response and gut microbiota were distinct.21,35 Therefore, both PP and PO were used in this study, and their impact on the adjuvanticity of the nanoemulsion was clarified. Since PP and PO are not pure compounds and exist within a size range, further investigation will be required to understand whether the polydispersity of PO and PP affects adjuvanticity of nanoemulsions.
In our pilot study, we explored various concentrations of PO and PP, along with different ratios of the aqueous phase, lipid phase, and emulsifiers to create the emulsion. However, only the formulation utilized in the current study, which included the highest concentrations of PO and PP, demonstrated successful emulsion preparation without separation for up to 8 weeks. This indicates that the employed formulation exhibited relative stability. Additional research into the nanoemulsification process could be advantageous for optimizing the nanosystem. When formulating PP or PO into nanoemulsions, the zeta potentials were significantly reduced, indicating the adsorption of PP or PO with negatively charged sulfate groups on the droplets. Similarly, Hirano et al formulated Porphyra polysaccharide into corn oil-based emulsions and observed a significant decrease in zeta potential from −23 mV to −33 mV.36 Interestingly, both PP and PO improved the storage stability of nanoemulsions. This suggests that PP or PO adsorbed on the droplet surface reduced the surface tension between the oil and water phases, preventing aggregation through electrostatic interaction.37 Additionally, when dispersed in the continuous phase, PP or PO may prevent droplet flocculation by thickening or forming a three-dimensional network, thereby enhancing the stability of nanoemulsions.38 However, it is worth mentioning that the size of the nanoemulsions changed within 8 weeks, suggesting that the age of the nanoemulsion may have a significant impact on its adjuvant properties. Further investigation will be needed to explore this issue.
To adhere to the 3R principle, in vitro experiments were conducted to screen the cytotoxicity and immunomodulatory effects of HK-CA vaccines before conduction of animal experiments. The cell wall of C. albicans contains lipopolysaccharides and β-1,3-glucan, which can induce immune cell activation and inflammation.39 Consistently, HK-CA treatment increased the viability of splenocytes and cytokine production. However, emulsion-based vaccines decreased splenocyte viability and cytokine production, indicating their cytotoxic and immunosuppressive potential. In contrast, nanoemulsion-based vaccines had limited impact on cell viability and cytokine production. Furthermore, PO@NE-adjuvanted vaccines significantly increased the production of IFN-γ, a crucial cytokine for eliciting adaptive immune responses and preventing C. albicans infection. Therefore, nanoemulsion-based vaccines were selected for further investigations.
In an earlier investigation, the administration of ovalbumin combined with a nanoemulsion containing squalene through the nasal route resulted in an augmented presence of CD4+ and CD8+ T cells, along with a heightened production of IFN-γ and IL17.40 The same nanoemulsion was subsequently used for intravaginal immunization, leading to increased levels of IFN-γ, IL17, and IgA production in vaginal wash samples of mice.13 Although the antigen used in this study was different from that employed in the above studies, similar results, including increment of CD4+ and CD8+ T cell populations, IgA and IFN-γ production, were observed in mice intravaginally vaccinated with NEmO-adjuvanted HK-CA in this study. These findings further emphasize the importance of the aqueous phase in nanoemulsion for its immunomodulatory effects. Despite prior research indicating that the mechanism of action for squalene-based nanoemulsion involves prolonging antigen retention in the genital tract, facilitating antigen delivery across the mucosal barrier, and promoting the recruitment and activation of antigen-presenting cells, additional investigations are necessary to determine if the adjuvant effect of PO@NE functions through similar mechanisms.13,40
Natural polysaccharides and oligosaccharides are the main sources of prebiotics, which influence the composition of commensal microflora.29 Maintaining the homeostasis of vaginal microflora is crucial for the health of the urogenital tract.30,41 In order to investigate the effect of vaginal administration of PP@NE- and PO@NE-adjuvanted HK-CA on vaginal microflora, vaginal samples were collected, and the bacterial species were identified. The vaginal microflora of untreated mice primarily consisted of Proteobacteria and Firmicutes. Barfod et al pointed out that the vaginal microflora of BALB/cJ mice is mainly composed of Proteobacteria, Firmicutes, and a small amount of Actinobacteria, Bacteroidetes, and Fusobacteria.42 In this study, the NEmO group had the highest Bray-Curtis index, indicating that the intravaginal vaccination of PO@NE-adjuvanted HK-CA could increase the number or variety of vaginal bacteria. Although no variation in the lactobacilli abundance was observed, the relative abundance of Actinobacteria was increased significantly in the NEmO group. Actinobacteria has been found to exhibit potent activity against C. albicans.43,44 Therefore, modulation of vaginal microflora is considered one of the potential mechanisms of PO@NE-adjuvanted HK-CA in improving candidal vaginitis. On the other hand, consistent with the results of this study, recent studies have shown no significant differences in the vaginal microbiome between women with and without frequent vulvovaginal candidiasis.45,46 However, an increase in vaginal microflora diversity was observed in the mice of the NEmO group in the current study. The process of PO preparation included autoclaving, and the emulsion was passed through a sterile 0.22 µm filter to harvest the nanoemulsion. Therefore, the introduction of bacteria into the vaginal tract through PO@NE treatment was impossible. An obvious relative abundance of Actinobacteria was observed only in the NEmO group, which may explain the markedly increased beta diversity of vaginal microflora in the NEmO group. Accordingly, it is suggested that PO@NE selectively enhanced the growth of certain vaginal microbes that were beneficial for the improvement of candidal vaginitis.
The majority of commercial products and available researches pertaining to intravaginal vaccine primarily concentrate on sexually transmitted infections such as human papillomavirus, human immunodeficiency virus, herpes simplex virus, Chlamydia trachomatis, and Neisseria gonorrhoeae. However, there is a dearth of comprehensive knowledge concerning intravaginal vaccines targeting Candida infections.47 Mucosal adjuvants play a critical role in successful vaccination, and nanoemulsions have been investigated in several studies due to their advantages in enhancing vaginal retention, mucus penetration, cellular targeting, and permitting versatile formulation approaches.48 Nanoemulsions consist of emulsifiers, oil phase, and aqueous phase, with each component playing a crucial role in adjuvanticity. In this study, Tween 80® and Span 85® were employed as emulsifiers. It has been reported that the release of ATP from muscle, which enhances the vaccination response, can be attributed to the presence of Tween 80® and Span 85® in MF59 adjuvants.49 Tween 80® and Span 85®-based adjuvants are commonly used in injectable vaccines. However, the effect of intranasal immunization with MF59-adjuvanted vaccines in enhancing anti-HIV plasma and vaginal antibody responses, as well as peripheral blood IFN-γ responses, have been reported.50 DHA, employed in this study as the core carrier oil, is an immunomodulatory lipid.51 Recently, Bakkari et al designed DHA nanovesicles as vaccine adjuvants and demonstrated their ability to enhance antigen-specific immune responses.52 Moreover, injection of DHA-based vaccine enhanced tumor immunotherapy by augmenting tumor-specific immune responses.53 On the other hand, water-soluble algal polysaccharides are known to have modulatory effects on immune cells, highlighting their potential as vaccine adjuvants.54 For example, fucoidan, as a vaccine adjuvant, increased the viability and cytokine production of spleen cells and raised the production of antigen-specific antibodies.55 The above-mentioned studies provide promising evidence of the adjuvanticity of Tween 80®, Span 85®, DHA, and algal polysaccharides in injectable vaccines individually. However, their impact and even synergy on the efficacy of intravaginal vaccines remains unclear. Innovatively, these potential components were formulated into nanoemulsions, and their adjuvanticity for intravaginal HK-CA vaccines was substantiated in this study.
Conclusion
The findings of this investigation offer initial evidence supporting the protective effects of intravaginal PO@NE-adjuvanted HK-CA against candidal vaginitis. These effects may be attributed to upregulation of mucosal antibody production, activation of Th1- and Th17-related immune responses, expansion of CD8+ cell populations, and enhancement of vaginal microflora diversity, offering insight into the design and development of intravaginal vaccine.
Ethical Statements
All procedures and protocols were conducted in accordance with the guidelines outlined in the National Research Council’s Guide for the Care and Use of Laboratory Animals. Additionally, the experimental protocols were approved by the NTOU Institutional Animal Care and Use Committee (NTOU IACUC-109057).
Acknowledgments
Financial support for this research was provided through grants from the National Science and Technology Council of Taiwan (MOST 109-2320-B-019-007-MY3 and MOST 109-2221-E-019-032-MY3). The authors would like to thank professor Shiow-Yi Chen (Department of Bioscience and Biotechnology, National Taiwan Ocean University, Keelung, Taiwan) for the giving of C. albicans (ATCC 10231). We also would like to thank the Animal Technology Research Center of Agricultural Technology Research Institute (Miaoli, Taiwan), the Pathology Core Laboratory of National Health Research Institutes (Miaoli, Taiwan) and the National Laboratory Animal Center of National Applied Research Laboratories (Taipei, Taiwan) for the assistance of histopathological examination, and BIOTOOLS Co., Ltd. (New Taipei, Taiwan) for the assistance of NGS analysis.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Pfaller MA, Diekema D. Epidemiology of invasive candidiasis: a persistent public health problem. Clin Microbiol Rev. 2007;20(1):133–163. doi:10.1128/CMR.00029-06
2. Aballéa S, Guelfucci F, Wagner J, et al. Subjective health status and health-related quality of life among women with recurrent vulvovaginal candidosis (RVVC) in Europe and the USA. Health Qual Life Outcomes. 2013;11(1):1–13. doi:10.1186/1477-7525-11-169
3. Berkow EL, Lockhart SR. Fluconazole resistance in Candida species: a current perspective. Infect Drug Resist. 2017;10:237–245. doi:10.2147/IDR.S118892
4. Popp C, Hampe IA, Hertlein T, Ohlsen K, Rogers PD, Morschhäuser J. Competitive fitness of fluconazole-resistant clinical Candida albicans strains. Antimicrob Agents Chemother. 2017;61(7):e00584–00517. doi:10.1128/AAC.00584-17
5. Netea MG, Joosten LA, Van Der Meer JW, Kullberg B-J, Van De Veerdonk FL. Immune defence against Candida fungal infections. Nat Rev Immunol. 2015;15(10):630–642. doi:10.1038/nri3897
6. Medzhitov R, Janeway CA. Innate immunity: impact on the adaptive immune response. Curr Opin Immunol. 1997;9(1):4–9. doi:10.1016/S0952-7915(97)80152-5
7. Pashine A, Valiante NM, Ulmer JB. Targeting the innate immune response with improved vaccine adjuvants. Nat Med. 2005;11(Suppl 4):S63–S68. doi:10.1038/nm1210
8. Lycke N. Recent progress in mucosal vaccine development: potential and limitations. Nat Rev Immunol. 2012;12(8):592–605. doi:10.1038/nri3251
9. Marasini N, Skwarczynski M, Toth I. Oral delivery of nanoparticle-based vaccines. Expert Rev Vaccines. 2014;13(11):1361–1376. doi:10.1586/14760584.2014.936852
10. Czerkinsky C, Holmgren J. Topical immunization strategies. Mucosal Immunol. 2010;3(6):545–555. doi:10.1038/mi.2010.55
11. Ramirez JEV, Sharpe LA, Peppas NA. Current state and challenges in developing oral vaccines. Adv Drug Deliv Rev. 2017;114:116–131. doi:10.1016/j.addr.2017.04.008
12. Black S. Safety and effectiveness of MF-59 adjuvanted influenza vaccines in children and adults. Vaccine. 2015;33:B3–B5. doi:10.1016/j.vaccine.2014.11.062
13. H-M H, Huang C-Y, Cheng Y-J, et al. Squalene nanoemulsion reinforces mucosal and immunological fingerprints following intravaginal delivery. Biomed Pharmacother. 2021;141:111799.
14. Cian RE, Fajardo MA, Alaiz M, Vioque J, González RJ, Drago SR. Chemical composition, nutritional and antioxidant properties of the red edible seaweed Porphyra columbina. Int J Food Sci Nutr. 2014;65(3):299–305. doi:10.3109/09637486.2013.854746
15. Yu P, Zhang Y. Separation and purification of Porphyra haitanensis polysaccharide and its preliminary structural characterization. Sep Sci Technol. 2017;52(11):1835–1842. doi:10.1080/01496395.2017.1296464
16. Wang Y, Hwang J-Y, H-b P, Yadav D, Oda T, Jin J-O. Porphyran isolated from Pyropia yezoensis inhibits lipopolysaccharide-induced activation of dendritic cells in mice. Carbohydr Polym. 2020;229:115457. doi:10.1016/j.carbpol.2019.115457
17. Wang Y, Hwang J, Yadav D, Oda T, Lee PC-W, Jin J-O. Inhibitory effect of porphyran on lipopolysaccharide-induced activation of human immune cells. Carbohydr Polym. 2020;232:115811. doi:10.1016/j.carbpol.2019.115811
18. Shi C, Pan T, Cao M, Liu Q, Zhang L, Liu G. Suppression of Th2 immune responses by the sulfated polysaccharide from Porphyra haitanensis in tropomyosin-sensitized mice. Int Immunopharmacol. 2015;24(2):211–218. doi:10.1016/j.intimp.2014.11.019
19. Osumi Y, Kawai M, Amano H, Noda H. Physiological activities of oligosaccharides derived from marine algae Porphyra yezoensis Porphyran. Fisher Sci. 2002;68(sup2):1441–1444. doi:10.2331/fishsci.68.sup2_1441
20. Wu S-C, Kang S-K, Kazlowski B, C-j W, Pan C-L. Antivirus and prebiotic properties of seaweed-oligosaccharide-lysates derived from agarase AS-II. J Fish Soc Taiwan. 2012;39(1):11–21.
21. Wei Y-J, Fang R-E, J-y O, Pan C-L, Huang C-H. Modulatory effects of Porphyra-derived polysaccharides, oligosaccharides and their mixture on antigen-specific immune responses in ovalbumin-sensitized mice. J Funct Foods. 2022;96:105209. doi:10.1016/j.jff.2022.105209
22. Esmaeili F, Rajabnejhad S, Partoazar AR, et al. Anti-inflammatory effects of eugenol nanoemulsion as a topical delivery system. Pharm Dev Technol. 2016;21(7):887–893. doi:10.3109/10837450.2015.1078353
23. Huang C-H, Huang C-Y, Huang M-H. Unsaturated squalene content in emulsion vaccine adjuvants plays a crucial role in ROS-mediated antigen uptake and cellular immunity. Mol Pharm. 2018;15(2):420–429. doi:10.1021/acs.molpharmaceut.7b00800
24. Huang C-H, Lin Y-C, Jan T-R. Lactobacillus reuteri induces intestinal immune tolerance against food allergy in mice. J Funct Foods. 2017;31:44–51. doi:10.1016/j.jff.2017.01.034
25. Yano J, Fidel PL. Protocols for vaginal inoculation and sample collection in the experimental mouse model of Candida vaginitis. JoVE. 2011;58:e3382.
26. Hebbar PB, Pai A, D S. Mycological and histological associations of Candida in oral mucosal lesions. J Oral Sci. 2013;55(2):157–160. doi:10.2334/josnusd.55.157
27. Sroka-Oleksiak A, Gosiewski T, Pabian W, et al. Next-Generation Sequencing as a tool to detect vaginal microbiota disturbances during pregnancy. Microorganisms. 2020;8(11):1813. doi:10.3390/microorganisms8111813
28. Wang L-J, Yang C-Y, Kuo H-C, Chou W-J, Tsai C-S, Lee S-Y. Effect of Bifidobacterium bifidum on clinical characteristics and gut microbiota in Attention-Deficit/Hyperactivity Disorder. J Personal Med. 2022;12(2):227. doi:10.3390/jpm12020227
29. Chee WJY, Chew SY, Than LTL. Vaginal microbiota and the potential of Lactobacillus derivatives in maintaining vaginal health. Microb Cell Fact. 2020;19(1):203. doi:10.1186/s12934-020-01464-4
30. Balakrishnan SN, Yamang H, Lorenz MC, Chew SY, Than LTL. Role of vaginal mucosa, host immunity and microbiota in vulvovaginal candidiasis. Pathogens. 2022;11(6):618. doi:10.3390/pathogens11060618
31. Faith DP, Minchin PR, Belbin L. Compositional dissimilarity as a robust measure of ecological distance. Vegetatio. 1987;69(1–3):57–68. doi:10.1007/BF00038687
32. Gong G, Dang T, Fang J, et al. Preparation, structural characterization, and bioactivity of PHPD-IV-4 derived from Porphyra haitanensis. Food Chem. 2020;329:127042. doi:10.1016/j.foodchem.2020.127042
33. Isaka S, Cho K, Nakazono S, et al. Antioxidant and anti-inflammatory activities of porphyran isolated from discolored nori (Porphyra yezoensis). Int J Biol Macromol. 2015;74:68–75. doi:10.1016/j.ijbiomac.2014.11.043
34. Hou Y, Wang J, Simerly T, Jin W, Zhang H, Zhang Q. Hydrogen peroxide released from Pyropia yezoensis induced by oligo-porphyrans: mechanisms and effect. J Appl Phycol. 2015;27(4):1639–1649. doi:10.1007/s10811-014-0471-7
35. Wei YJ, Fang RE, Liu JS, et al. Influence of Porphyra-derived polysaccharides and oligosaccharides on attenuating food allergy and modulating enteric microflora in mice. Food Agric Immunol. 2023;34(1):2248419. doi:10.1080/09540105.2023.2248419
36. Hirano Y, Hattori M, Takahashi K. Interaction of porphyran with a hydrophobic surface and stabilization of liposomes. J Agric Food Chem. 2005;53(25):9800–9804. doi:10.1021/jf050793e
37. Xu X, Luo L, Liu C, McClements DJ. Utilization of anionic polysaccharides to improve the stability of rice glutelin emulsions: impact of polysaccharide type, pH, salt, and temperature. Food Hydrocoll. 2017;64:112–122. doi:10.1016/j.foodhyd.2016.11.005
38. Shao P, Feng J, Sun P, Xiang N, Lu B, Qiu D. Recent advances in improving stability of food emulsion by plant polysaccharides. Food Res Int. 2020;137:109376. doi:10.1016/j.foodres.2020.109376
39. Issara-Amphorn J, Dang CP, Saisorn W, Limbutara K, Leelahavanichkul A. Candida administration in bilateral nephrectomy mice elevates serum (1→ 3)-β-D-glucan that enhances systemic inflammation through energy augmentation in macrophages. Int J Mol Sci. 2021;22(9):5031. doi:10.3390/ijms22095031
40. Huang C-H, Huang C-Y, H-m H, et al. Nanoemulsion adjuvantation strategy of tumor-associated antigen therapy rephrases mucosal and immunotherapeutic signatures following intranasal vaccination. J Immunother Cancer. 2020;8(2):e001022. doi:10.1136/jitc-2020-001022
41. Lehtoranta L, Ala-Jaakkola R, Laitila A, Maukonen J. Healthy vaginal microbiota and influence of probiotics across the female life span. Front Microbiol. 2022;13:787.
42. Barfod KK, Roggenbuck M, Hansen LH, et al. The murine lung microbiome in relation to the intestinal and vaginal bacterial communities. BMC Microbiol. 2013;13(1):1–12. doi:10.1186/1471-2180-13-303
43. Pavan Kumar J, Gomathi A, Vasconcelos V, Gothandam K. Bioactivity assessment of Indian origin—mangrove actinobacteria against Candida albicans. Mar Drugs. 2018;16(2):60. doi:10.3390/md16020060
44. Mendes TD, Borges WS, Rodrigues A, et al. Anti-Candida properties of urauchimycins from actinobacteria associated with Trachymyrmex ants. Biomed Res Int. 2013;2013:835081. doi:10.1155/2013/835081
45. Tortelli BA, Lewis WG, Allsworth JE, et al. Associations between the vaginal microbiome and Candida colonization in women of reproductive age. Am J Obstet Gynecol. 2020;222(5):471. doi:10.1016/j.ajog.2019.10.008
46. Sun Z, Ge X, Qiu B, et al. Vulvovaginal candidiasis and vaginal microflora interaction: microflora changes and probiotic therapy. Front Cell Infect Microbiol. 2023;13:1123026. doi:10.3389/fcimb.2023.1123026
47. McIntosh EDG. Development of vaccines against the sexually transmitted infections gonorrhoea, syphilis, Chlamydia, herpes simplex virus, human immunodeficiency virus and Zika virus. Therap Adv Vaccines Immunother. 2020;8:2515135520923887. doi:10.1177/2515135520923887
48. Huang M, Zhang M, Zhu H, Du X, Wang J. Mucosal vaccine delivery: a focus on the breakthrough of specific barriers. Acta Pharm Sin B. 2022;12(9):3456–3474. doi:10.1016/j.apsb.2022.07.002
49. Vono M, Taccone M, Caccin P, et al. The adjuvant MF59 induces ATP release from muscle that potentiates response to vaccination. Proce Natl Acad Sci. 2013;110(52):21095–21100. doi:10.1073/pnas.1319784110
50. Vajdy M, Singh M, Kazzaz J, et al. Mucosal and systemic anti-HIV responses in rhesus macaques following combinations of intranasal and parenteral immunizations. AIDS Res Hum Retroviruses. 2004;20(11):1269–1281. doi:10.1089/aid.2004.20.1269
51. Johnson WT, Dorn NC, Ogbonna DA, Bottini N, Shah NJ. Lipid‐based regulators of immunity. Bioeng Transl Med. 2022;7(2):e10288. doi:10.1002/btm2.10288
52. Bakkari MA, Moni SS, Alshammari A, et al. Design, characterization, and immune augmentation of docosahexaenoic acid nanovesicles as a potential delivery system for recombinant HBsAg protein. Vaccines. 2022;10(6):954. doi:10.3390/vaccines10060954
53. Zhang L, Huang J, Chen X, et al. Self-assembly nanovaccine containing TLR7/8 agonist and STAT3 inhibitor enhances tumor immunotherapy by augmenting tumor-specific immune response. J Immunother Cancer. 2021;9(8):e003132. doi:10.1136/jitc-2021-003132
54. Sanina N. Vaccine adjuvants derived from marine organisms. Biomolecules. 2019;9(8):340. doi:10.3390/biom9080340
55. Kim S-Y, Joo H-G. Evaluation of adjuvant effects of fucoidan for improving vaccine efficacy. J Vet Sci. 2015;16(2):145–150. doi:10.4142/jvs.2015.16.2.145
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.