Back to Journals » Journal of Pain Research » Volume 12
Electroacupuncture Alleviates Spared Nerve Injury-Induced Neuropathic Pain And Modulates HMGB1/NF-κB Signaling Pathway In The Spinal Cord
Authors Xia Y, Xue M, Wang Y ![]() , Huang Z
, Huang Z ![]() , Huang C
, Huang C ![]()
Received 20 June 2019
Accepted for publication 12 September 2019
Published 16 October 2019 Volume 2019:12 Pages 2851—2863
DOI https://doi.org/10.2147/JPR.S220201
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Michael Überall
Yang-yang Xia,1 Meng Xue,1 Ying Wang,1 Zhi-hua Huang,1,2 Cheng Huang1,2
1Department of Physiology, Gannan Medical University, Ganzhou 341000, People’s Republic of China; 2Pain Medicine Research Institute, Gannan Medical University, Ganzhou 341000, People’s Republic of China
Correspondence: Cheng Huang; Zhi-hua Huang
Department of Physiology, Gannan Medical University, Ganzhou 341000, People’s Republic of China
Email [email protected]; [email protected]
Background: Neuropathic pain with complications greatly affects patients worldwide. High mobility group box 1 (HMGB1) has been shown to contribute to the pathogenesis of neuropathic pain; thus, suppression of HMGB1 may provide a novel therapeutic option for neuropathic pain. Electroacupuncture (EA) has been indicated to be effective in attenuating neuropathic pain, but the underlying mechanism remains to be fully clarified. We aim to explore whether 2Hz EA stimulation regulates the spinal HMGB1/NF-κB signaling in neuropathic pain induced by spared nerve injury (SNI).
Materials and methods: Paw withdrawal threshold and CatWalk gait analysis were used to assess the effect of 2Hz EA on pain-related behaviors in SNI rats. Administration of 2Hz EA to SNI rats once every other day lasting for 21 days. Expression of spinal protein molecules were detected using Western blot and immunofluorescence staining.
Results: It was found that SNI significantly induced mechanical hypersensitivity and decrease of gait parameters, and subsequently increased the levels of HMGB1, TLR4, MyD88, and NF-κB p65 protein expression. 2Hz EA stimulation led to remarkable attenuation of mechanical hypersensitivity, upregulation of spinal HMGB1, TLR4, MyD88, and NF-κB p65 protein expressions induced by SNI, and significant improvement in gait parameters. Furthermore, immunofluorescence staining also confirmed that 2Hz EA obviously suppressed the co-expression of microglia activation marker CD11b and TLR4 or MyD88, as well as the activation of NF-κB p65 in SNI rats.
Conclusion: This study suggested that blockade of HMGB1/NF-κB signaling in the spinal cord may be a promising therapeutic approach for 2Hz EA management of SNI-induced neuropathic pain.
Keywords: electroacupuncture, neuropathic pain, spared nerve injury, HMGB1/NF-κB signaling, TLR4
Introduction
Neuroinflammation is a pathological mechanism implicated in the initiation and development of neuropathic pain.1 High mobility group box 1 (HMGB1), a potent pro-inflammatory mediator, has been shown to elicit inflammatory responses and plays a crucial role in neuroinflammation.2 Studies have indicated that a crosstalk between HMGB1 and pro-inflammatory cytokines induces and maintains inflammatory activities. HMGB1 may aggravate the persistent pain state in the development of chronic pain.2,3 Peripheral nerve injury induces the elevation of HMGB1 and maintenance of neuropathic pain depends on HMGB1 release.2,3 These findings greatly suggest that HMGB1 is involved in the pathogenesis of neuropathic pain. It is reported that injection of HMGB1 to rats evokes neuropathic pain,4 and administration of anti-HMGB1 antibody effectively ameliorates pain hypersensitivity induced by spinal nerve ligation and partial sciatic nerve ligation in the spinal cord.2,5 It is evident that HMGB1 is acted as a potential therapeutic target for neuropathic pain.
It is clear that peripheral nerve injury induces microglia activation.6,7 TLR4, one of the potent HMGB1 receptors, has been indicated as the initiator and mediator of neuropathic pain,8 and it was highly expressed in the microglia of the spinal cord.9 When bounding to HMGB1, TLR4 activates spinal microglia to promote the release of pro-inflammatory cytokines.7 However, mice lacking TLR4 shows remarkable inhibition of both microglia activation and pain hypersensitivity following peripheral nerve injury.10 Furthermore, suppression of TLR4 can reduce microglia activation and alleviate neuropathic pain.10 Overall, these results suggest that HMGB1-triggered TLR4 activation in spinal microglia contributes to the development of neuropathic pain.
There is evidence that myeloid differentiation factor-88 adaptor protein (MyD88) in the spinal microglia mediated the activation of TLR4 and NF-κB signaling.11 Nerve injury-evoked neuropathic pain significantly upregulated spinal MyD88 protein expression,12 suggesting that MyD88 plays a key role in the pathogenesis of neuropathic pain. After nerve injury, the NF-κB signaling is activated along with TLR4 activation in spinal microglia,13 and subsequently promotes the upregulation of pro-inflammatory cytokines and implicates in the initiation and development of neuropathic pain.14
Neuropathic pain severely impacts the quality of patient life and leads to a wide variety of problems worldwide.15 However, the current pharmacological therapeutics for neuropathic pain are still limited,16 therefore, other managements with little side effects should be considered. It is well known that electroacupuncture (EA) has been used in People’s Republic of China and other oriental countries for the treatment of chronic pain with few side effects.17,18 EA stimulation is shown to attenuate neuropathic pain by activating a numerous of neurotransmitters via peripheral and central mechanisms, such as opioids, which block pro-inflammatory cytokines production, glial activation, and some signaling molecules expression in the spinal cord.19 Furthermore, our recent data revealed that 2Hz EA has been identified as an effective treatment for SNI-induced pain hypersensitivity,20,21 and it alleviates neuropathic pain by suppressing a pro-inflammatory cytokine IL-1β in the spinal cord of SNI rats.20 But the underlying mechanism remains poorly elucidated. Based on the fact that HMGB1 modulates the release of IL-1β, IL-6, and TNF-α through activating spinal microglia, including TLR4/MyD88/NF-κB signaling in neuropathic pain. Thus, in this study, we hypothesized that 2Hz EA has its inhibitory effect on spinal HMGB1/NF-κB signaling in SNI-induced neuropathic pain.
In the present study, paw withdrawal threshold (PWT) and CatWalk gait analysis were used to evaluate the effect of 2Hz EA on pain-related behaviors in SNI rats. To further determine the effect of 2Hz EA on the levels of spinal HMGB1, TLR4, MyD88, and NF-κB p65 induced by SNI, Western blot was used to quantify the amount of them. Moreover, immunofluorescence staining was used to observe the co-localization of TLR4 or MyD88 with microglial activation marker CD11b, as well as the distribution of NF-κB p65 in the spinal cord. Findings from this study may provide a clue that HMGB1/NF-κB signaling serves as a potential therapeutic target for 2Hz EA treating neuropathic pain.
Materials And Methods
Experimental Animals
Healthy adult male Sprague-Dawley rats weighing 160–180g (7 to 8 weeks old) were obtained from Hunan SLAC Laboratory Animal Co., Ltd. (Changsha, People’s Republic of China). The rats were housed five per cage with free access to food and water, and they were kept at a temperature-controlled (22°C–24°C) room and under a 12/12 hrs light/dark cycle. The rats were adapted to the surroundings for 5 days at the beginning of the study. The behavioral experiments were performed in a quiet environment and conducted in a double-blind manner. In all experiments, caution was taken to minimize discomfort to the rats. All animal experimental procedures were approved by the Animal Use and Protection Committee of Gannan Medical University in China and carried out in accordance with the guidelines of the International Association for the Study of Pain and the National Institute of Health Guide for the Care and Use of Laboratory Animals.
Experimental Procedures
Experimental procedures for SNI-induced neuropathic pain can be found in this study. For the first procedure (Figure 1A), rats were randomly divided into 3 groups: sham, SNI, and SNI+EA (n=10–14). The sham rats underwent sham surgery, while the other groups received SNI surgery. The rats in the SNI+EA group were treated with 2Hz EA on day 1 post-SNI surgery and once every other day lasting for 21 days. The rats in the sham and SNI groups were treated in the same way as those in the SNI+EA group except EA stimulation. The PWT and CatWalk gait analysis were performed following administration of 2Hz EA on day 21 post-SNI surgery, respectively. In the second procedure (Figure 1B), to further explore the mechanism underlying 2Hz EA alleviating SNI-induced neuropathic pain in the spinal cord, rats from each group were sacrificed to dissect the L4-L6 segments of spinal cord on day 21 post-surgery after behavioral experiment. The spinal cord segments at L4-L6 were collected for immunofluorescence staining and Western blot.
 |
Figure 1 The schematic diagram of the experimental procedures. Notes: (A) The PWT was assessed after 2Hz EA stimulation on day 21 post-SNI surgery. Rats received 2Hz EA treatment once every other day lasting for 21 days. The CatWalk gait analysis was performed on the same day as the examination of PWT. (B) Following 2Hz EA treatment for 21 days, rats were sacrificed after the measurement of PWT, and the samples were collected from the L4–L6 segment of spinal cord for immunofluorescence staining and Western blot experiments.Abbreviations: PWT, paw withdrawal threshold; EA, electroacupuncture; SNI, spared nerve injury. |
SNI Model Of Neuropathic Pain
After 5 days of adaptation, the SNI-induced neuropathic pain rat model was established according to the method described in the previous report.20 Briefly, rats were anesthetized with 2% to 3% isoflurane, followed by exposing the left lateral sciatic nerve and its three terminal branches (the sural, common peroneal, and tibial nerves). The left lateral common peroneal and the tibial nerves were tightly ligated with a 5–0 silk suture and sectioned distal to the ligation, removing 2–4 mm of the distal nerve stump, and leaving the sural nerve intact. Muscles and skin were closed in two layers under sterile operation. Only rats that developed mechanical hypersensitivity were used in this experiment. Sham-surgery rats underwent all identical surgical procedures except that the tibial and common peroneal nerves were left intact.
EA Stimulation
Rats were housed in specially designed holders with their hind legs and tails exposed as described in our previous study.20 Briefly, the skin of the rat hind legs was sterilized with 75% alcohol. Two stainless-steel needles (4 mm length and 0.4 mm diameter) were inserted into a couple of acupoints of each hind leg in rats. One needle was inserted at the Zusanli acupoint (ST36), which was 5 mm lateral to anterior tubercle of tibia marked by a notch, and the other needle was inserted at the Sanyinjiao acupoint (SP6), which was 3 mm proximal to medial malleolus and at the posterior border of the tibia. The stimulation square waves of EA, generated from Han’s Acupoint Nerve Stimulator (HANS, LH202, Beijing Huawei Industrial Developing Company, People’s Republic of China), were applied to both hind legs of rats simultaneously. Since 2Hz EA stimulation possesses better analgesia on neuropathic pain than 100Hz EA does. In this study, the EA stimulation parameters were: 2Hz with a pulse width of 0.6 ms, and the stimulation intensity was increased in a stepwise manner at 1-2-3 mA, each intensity lasting for 10 mins. Administration of 2Hz EA stimulation to rats once every other day lasting for 21 days. All rats were always kept awaken in the process of 2Hz EA stimulation.
Assessment Of Mechanical Hypersensitivity
Mechanical hypersensitivity was served as a behavioral sign of neuropathic pain. The sensitivity to mechanical stimuli was examined by PWT as described in a previous study.20 In brief, rats were first acclimatized in each individual plastic enclosure (12×22×18 cm) on a metal mesh floor standing for 15 mins before assessment. PWT was detected with a dynamic plantar aesthesiometer (Ugo Basile, 37450, Italy), which comprised a force transductor fitted with a 0.5-mm diameter polypropylene rigid tip. A probe was applied perpendicularly to the mid-plantar surface of the rat left hind paw with an increasing pressure, lasting for 10 s from 0 to 50 g. To avoid injury to rats, the cutoff pressure was set to be 50g, the force that induced the withdrawal response was automatically recorded to the nearest 0.1 g by the anesthesiometer. Three examinations were taken with an interval of 5 mins. The mean PWT for the three measurements was achieved from each rat.
CatWalk-Automated Gait Analysis
The CatWalk system (XT, Noldus Information Technology, the Netherlands) was used for the quantitative assessment of gait parameter and footfalls in rodents. The CatWalk system provides an objective and accurate examination of limb coordination, and it has been used for neuropathic pain study.22 Briefly, the CatWalk XT system has a high-speed digital camera, which transforms scene into digital images. The digital image is transferred to a computer. The brightness of a pixel was the amount of light received from an area by the camera. This experiment was conducted in a darkroom environment, and in the case of minimizing interference factors other than food attraction, each rat was subject to pre-training so that they did not stop and smoothly pass the CatWalk glass plate within 10 s. The baseline value was collected, rats that did not continue to pass the training were excluded. Twenty-one days after 2Hz EA treatment, rats were tested using the CatWalk system. After each group of rats smoothly passed through the glass plate, reflected light, emitted by the fluorescent lamp, in the glass was collected, and a series of gait parameters were automatically generated, and the recordings evaluated off-line. This experiment mainly examined pain-related behavior alterations from the following gait parameters: stand(s) is the duration of ground contact for a single paw, print area (cm2) represents the surface of the complete print of a paw, swing speed (cm/s) is the duration of no contact of a paw with the glass plate in a step cycle, stride length (cm) is the distance between successive placements of the same paw, print length (cm) is the length of the rectangle fitted to the paw print.
Immunofluorescence Staining
Following the assessment of behavioral alterations, rats were anesthetized using 2% to 3% isoflurane and the thoracic cavity was incised to expose the heart. A puncture needle was inserted from the left apex and extended towards the aorta. The right auricle was incised and the rat was transcardially perfused with 0.9% normal saline and then 4% paraformaldehyde solution in 0.1 M PBS (pH 7.4, 4°C). Immunofluorescence staining was performed as described in the previous study.8 Briefly, the spinal cord segments at L4-L6 were immediately removed and put into 4% paraformaldehyde solution for 4 hrs, followed by cryoprotection in 20% and 30% sucrose, respectively, at 4°C until it submerged. Transverse sections (10 μm) were cut on a cryostat, and blocked with 5% BSA for 60 mins at room temperature, after washing with PBS for 15 mins and incubated with 5% BSA for overnight at 4°C, the sections were incubated in the antibody solution containing rabbit anti-TLR4 (1:200, 19811-1-AP, Proteintech, USA), rabbit anti-MyD88 (1:100, ab2064, Abcam, UK) or rabbit anti-phospho-NF-κB p65 (1:100, 3033, CST, UK), and mouse anti-CD11b for microglia (1:50, ab1211, Abcam, UK), respectively. After washing with PBS for 15 mins, the sections were incubated for 120 mins at room temperature with Alexa FluorTM 555 goat anti-rabbit IgG (H+L) (1:1000, A21429, Invitrogen, USA), and Alexa FluorTM 488 goat anti-mouse IgG (H+L) (1:1000, A11029, Invitrogen, USA). The all stained sections were detected and analyzed with a confocal laser scanning fluorescence microscope.
Western Blot Analysis
Western blot assay was performed after behavioral experiments for each group. Rats were anesthetized with 2% to 3% isoflurane, the L4-L6 spinal cord segments were rapidly removed for extraction of proteins. Thirty micrograms of protein per sample were denatured and then separated by a 10% SDS-PAGE and Western blotted on a PVDF (Millipore, CA) membrane. The membrane was blocked with 5% non-fat milk in Tris-buffered saline containing 0.1% Tween-20 for 60 mins at room temperature, and subsequently the membrane was immuno-labelled overnight at 4◦C with antibodies of rabbit anti-HMGB1 (1:1000, ab79823, Abcam, UK), mouse anti-TLR4 (1:400, 66350-1-lg, Proteintech, USA), rabbit anti-MyD88 (1:500, ab2064, Abcam, UK), rabbit anti-NF-κB p65 (1:1000, 8242, CST, UK) and rabbit anti-phospho-NF-κB p65 (1:400, 3033, CST, UK), mouse anti-Tubulin (1:1000, M1000140, Solarbio, People’s Republic of China). The blots were washed with Tris-buffered saline and Tween-20 and then incubated with the horseradish peroxidase-conjugated anti-rabbit secondary antibody (1:1000, Cell Signaling Technology, USA) for 120 mins at room temperature. The blots site of the antigen-antibody complex was visualized with the Immobilon Western Chemiluminescent HRP Substrate (Millipore, MA). The bands were analyzed using Quantity One software (Bio-Rad). β-tubulin was used as the internal control. The standardized ratio of HMGB1, TLR4, MyD88, and phospho-NF-κB protein to β-tubulin band density was used to calculate the alteration of corresponding protein expression level.
Data Analysis And Statistics
All experimental data were expressed as the mean±SEM. Statistical analyses were performed using Prism 5.0 software. Differences between groups in behavioral experiments, immunofluorescence staining, and Western blotting tests were analyzed using one-way ANOVA followed by Newman–Keuls post-hoc tests. P<0.05 was considered to represent significant difference.
Results
Effect Of 2Hz EA On SNI-Evoked Mechanical Hypersensitivity
Mechanical hypersensitivity is a typical sign of neuropathic pain, it is more pronounced in SNI-induced mechanical hypersensitivity. To examine the effect of EA stimulation on mechanical hypersensitivity in SNI rats, the PWT was measured following administration of 2Hz EA to rats post-SNI surgery. As shown in Figure 2, the PWT in the SNI group was obviously reduced compared with that of the sham group (P<0.001), indicating the successful establishment of SNI-induced neuropathic pain rat model. In comparison with the SNI group, the PWT in the SNI+EA group was markedly increased (P<0.001), suggesting that 2Hz EA attenuated SNI-induced mechanical hypersensitivity.
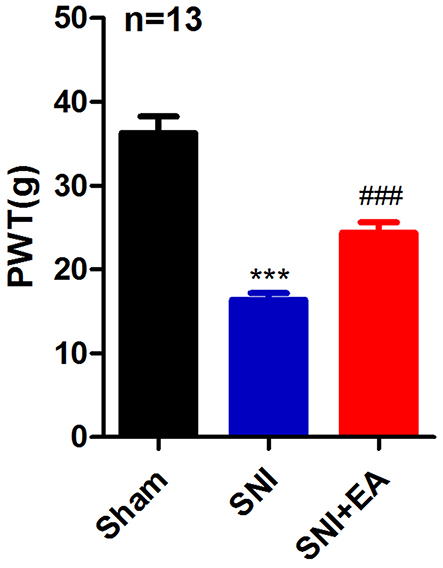 |
Figure 2 Effect of 2Hz EA on paw withdrawal threshold (PWT) induced by spared nerve injury. Notes: Rats were treated with 2Hz EA once every other day for 21 days post-surgery. Sham represents the sham group; SNI represents rats treated with the spared nerve injury; SNI+EA represents rats receiving spared nerve injury and 2Hz EA treatment. ***P<0.001, compared to the sham group, ###P<0.001, compared to the SNI group. All data were expressed as the mean±SEM, n =13 per group.Abbreviations: PWT, paw withdrawal threshold; EA, electroacupuncture; SNI, spared nerve injury; SEM, standard error of the mean. |
Effect Of 2Hz EA On SNI-Induced CatWalk Gait Analysis
CatWalk gait analysis, used to assess gait parameters, is served as an objective method to evaluate sensory neuropathic pain induced by SNI. In this experiment, we measured five gait parameters to display pain-related behaviors in SNI rats. As shown in Figure 3A–E, in comparison with the sham group, SNI significantly decreased stand, print area, swing speed, stride length, and print length on day 21 post-surgery (P<0.001), suggesting that SNI induced sensory impairment to rats. These decreases were significantly attenuated by 2Hz EA treatment to SNI rats (P<0.05, P<0.01, P<0.001). These data demonstrated that 2Hz EA alleviated pain-related behaviors in SNI rats.
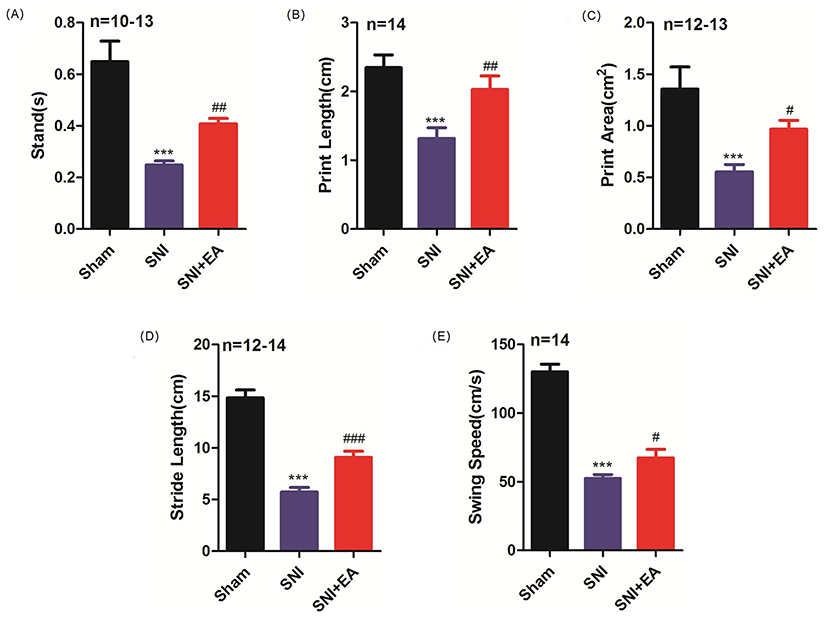 |
Figure 3 Effect of 2Hz EA on Catwalk gait parameters in SNI rats. Notes: Rats were treated with 2Hz EA once every other day for 21 days post-surgery. Sham represents the sham group; SNI represents rats treated with the spared nerve injury; SNI+EA represents rats receiving spared nerve injury and 2Hz EA treatment. The measurement of Catwalk gait parameters of stand(s), print length (cm), print area (cm2), stride length (cm), and swing speed (cm/s) is shown in (A–E). ***P<0.001, compared to the sham group, #P<0.05, ##P<0.01, ###P< 0.001, compared to the SNI group. All data were expressed as the mean±SEM, n=10–14 per group.Abbreviations: EA, electroacupuncture; SNI, spared nerve injury; SEM, standard error of the mean. |
Effect Of 2Hz EA On Spinal HMGB1 Protein Expression In SNI Rats
To explore the effect of 2Hz EA on HMGB1 in the spinal cord of SNI rats, spinal HMGB1 protein expression was determined on day 21 post-surgery. As shown in Figure 4A and B, in comparison with the sham group, HMGB1 in the SNI group was significantly upregulated (P<0.001) on day 21 post-SNI surgery. 2Hz EA stimulation markedly decreased the expression level of HMGB1 protein in SNI rats (P<0.01) on day 21 post-SNI surgery. These findings revealed that 2Hz EA downregulated HMGB1 protein expression in the spinal cord of SNI rats.
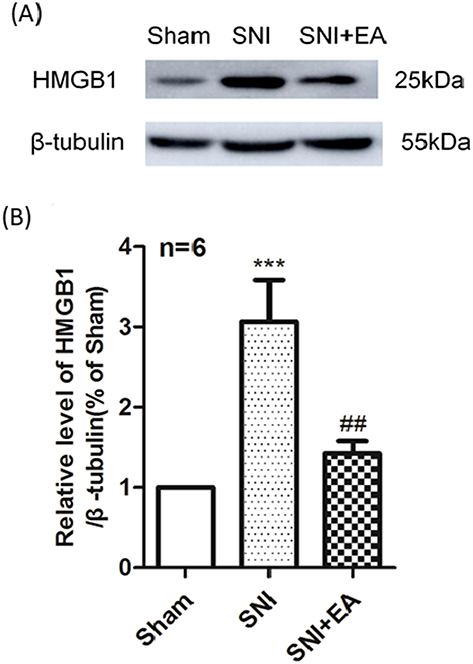 |
Figure 4 Effect of 2Hz EA on SNI-induced HMGB1 protein expression in the spinal cord. Notes: (A) Western blotted band of HMGB1. (B) Relative level of HMGB1/β-tubulin protein expression. Samples were collected from the L4–L6 segment of rat spinal cord on day 21 post-surgery. ***P<0.001, compared to the sham group, ##P<0.01, compared to the SNI group. All data were expressed as the mean±SEM, n=6 per group.Abbreviations: HMGB1, high mobility group box 1; EA, electroacupuncture; SNI, spared nerve injury; SEM, standard error of the mean. |
Effect Of 2Hz EA On TLR4 Protein Expression And Co-Localization Of TLR4 With CD11b In The Spinal Cord Of SNI Rats
To evaluate the role of TLR4 in 2Hz EA treatment to SNI rats, the level of TLR4 protein expression was detected on day 21 post-SNI surgery. As shown in Figure 5A and B, in comparison with the sham group, TLR4 protein expression in the SNI group was markedly increased (P<0.001) on day 21 post-surgery. In comparison with the SNI group, TLR4 protein expression level in the SNI+EA group was significantly reduced (P<0.001) on day 21 post-SNI surgery. To demonstrate the critical role of spinal microglia in the pathogenesis of SNI-induced neuropathic pain. Following 2Hz EA treatment to rats for 21 days post-SNI surgery, activation of microglia in the spinal cord was examined using immunofluorescence staining against the microglia activation marker CD11b. As shown in Figure 5C and D, SNI markedly induced the upregulation of CD11b (P<0.001) in the spinal cord compared with the sham group. 2Hz EA treatment significantly attenuated the upregulation of CD11b evoked by SNI (P<0.01). Similarly, immunofluorescence staining also showed that spinal TLR4 level was significantly increased in the SNI group (P<0.001) compared with that of the sham group. In comparison with the SNI group, 2Hz EA treatment significantly reduced SNI-induced upregulation of spinal TLR4 (P<0.01). The double immunofluorescence staining further showed that SNI increased the co-expression of TLR4 and CD11b in the dorsal horn of spinal cord compared with the sham group. 2Hz EA treatment significantly decreased the co-expression of spinal TLR4 and CD11b in SNI rats. These results confirmed that 2Hz EA decreased the expression of both spinal TLR4 and CD11b, as well as the activation of microglia after SNI.
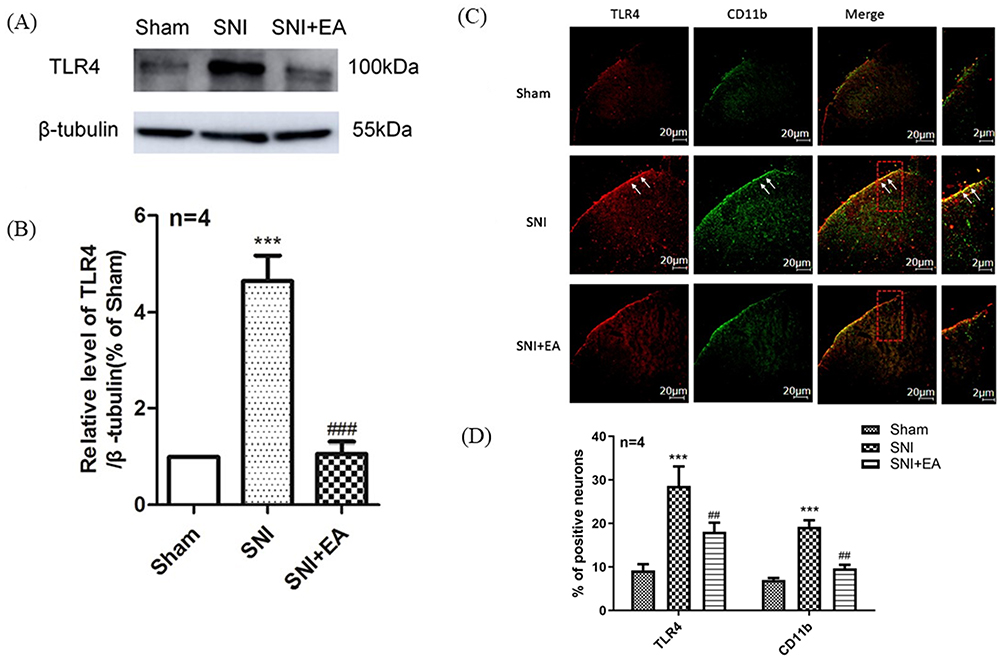 |
Figure 5 Effect of 2Hz EA on TLR4 protein expression and activation of microglia in the spinal cord following SNI.Notes: (A) Western blotted band of TLR4. (B) Relative level of TLR4/β-tubulin protein expression. (C) TLR4 and CD11b were double labeled in the dorsal horn of spinal cord (scale bar=20μm). 2Hz EA treatment significantly downregulated the co-expression of TLR4 (red) and microglial activation indicator CD11b (green). (D) Intensity mean value for TLR4 or CD11b. Samples were collected from the L4–L6 segment of rat spinal cord on day 21 post-surgery. ***P<0.001, compared to the sham group, ##P<0.01, ###P<0.001, in comparison with the SNI group. All data were expressed as the mean ±SEM, n=4 per group. Arrows indicate expressions of TLR4 and CD11b as well as co-localization of TLR4 with CD11b in the spinal dorsal horns.Abbreviations: TLR4, toll-like receptor 4; EA, electroacupuncture; SNI, spared nerve injury; SEM, standard error of the mean. |
Effect Of 2Hz EA On MyD88 Protein Expression And Co-Localization Of MyD88 With CD11b In The Spinal Cord Of SNI Rats
To explore whether MyD88 participates in the effect of 2Hz EA on SNI rats, the spinal MyD88 expression following 2Hz EA treatment was examined on day 21 post-SNI surgery. As shown in Figure 6A and B, in comparison with the sham group, MyD88 protein expression in the SNI group was significantly increased (P<0.05) on day 21 post-surgery. In comparison with the SNI group, 2Hz EA treatment obviously suppressed the upregulation of MyD88 protein expression in SNI rats (P<0.05) on day 21 post-SNI surgery. The role of 2Hz EA treatment on the expression of both MyD88 and CD11b in the spinal cord was examined using immunofluorescence staining. As shown in Figure 6C and D, in comparison with the sham group, SNI induced a significant increase of both MyD88 and CD11b in the dorsal horn of the spinal cord (P<0.001). As expected, co-localization of MyD88 with CD11b in the dorsal horn of spinal cord was observed. In comparison with the SNI group, after 2Hz EA treatment to SNI rats, a significant downregulation of both spinal MyD88 and CD11b were also found (P<0.05), which indicated that 2Hz EA decreased the expression of both MyD88 and CD11b and inhibited SNI-induced microglia activation in the spinal cord.
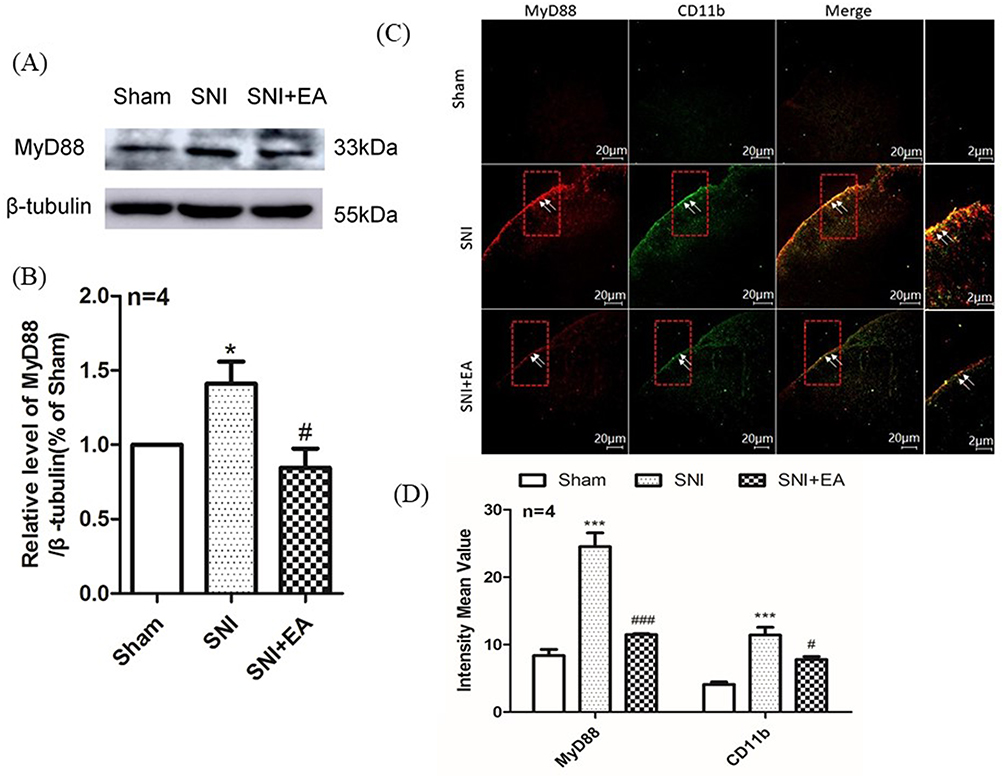 |
Figure 6 Effect of 2Hz EA on MyD88 protein expression and activation of microglia in the spinal cord following SNI. Notes: (A) Western blotted band of MyD88. (B) Relative level of MyD88/β-tubulin protein expression. (C) MyD88 and CD11b were double labeled in the dorsal horn of spinal cord (scale bar=20μm). 2Hz EA treatment significantly downregulated the co-expression of MyD88 (red) and microglial activation indicator CD11b (green). (D) Intensity mean value for MyD88 or CD11b. Samples were collected from the L4–L6 segment of rat spinal cord on day 21 post-surgery. *P< 0.05, ***P<0.001, compared to the sham group, #P<0.05, ###P<0.001, compared to the SNI group. All data were expressed as the mean±SEM, n=4 per group. Arrows indicate expressions of MyD88 and CD11b as well as co-localization of MyD88 with CD11b in the spinal dorsal horns.Abbreviations: MyD88, myeloid differentiation factor-88; EA, electroacupuncture; SNI, spared nerve injury; SEM, standard error of the mean. |
Effect Of 2Hz EA On Spinal NF-κB Activation In SNI Rats
To evaluate the role of spinal NF-κB signaling in 2Hz EA treatment to SNI rats, the activation of NF-κB was measured on day 21 post-SNI surgery through the detection of NF-κB p65 phosphorylation and its intracellular distribution in the spinal cord. As shown in Figure 7A and B, Western blot results showed that SNI significantly induced NF-κB p65 phosphorylation, compared to the sham group (P<0.01), whereas 2Hz EA treatment markedly inhibited SNI-induced NF-κB p65 phosphorylation (P<0.05). Furthermore, as shown in Figure 7C and D, immunofluorescence staining revealed that SNI induced NF-κB p65 nuclear translocation in the spinal dorsal horn and this effect was significantly suppressed by 2Hz EA stimulation. These findings suggested that 2Hz EA treatment inhibited SNI-induced activation of NF-κB signaling in the spinal cord.
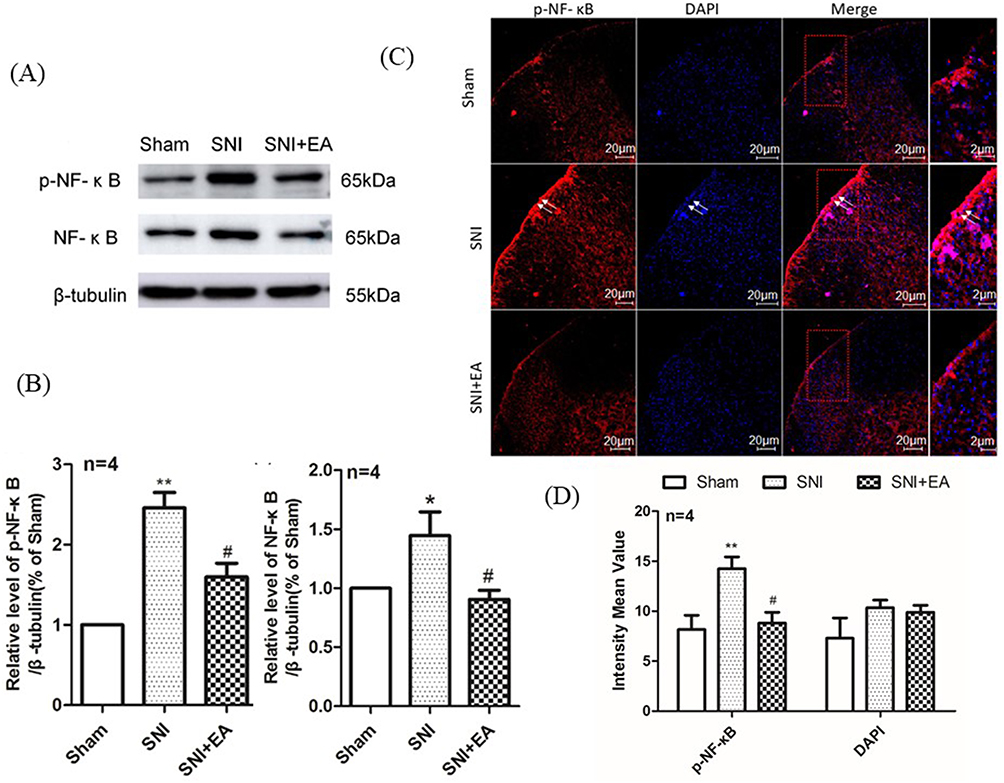 |
Figure 7 Effect of 2Hz EA on SNI-induced NF-κB activation in the spinal cord of rats. Notes: (A) Western blotted bands of p-NF-κB and NF-κB. (B) Relative levels of p-NF-κB/β-tubulin and NF-κB/β-tubulin protein expression. (C) p-NF-κB and DAPI were double labeled in the dorsal horn of spinal cord (scale bar=20μm). 2Hz EA treatment significantly downregulated the distribution of p-NF-κB (red) but not DAPI (blue). (D) Intensity mean value for p-NF-κB and DAPI. Samples were collected from the L4–L6 segment of rat spinal cord on day 21 post-surgery. *P<0.05, **P<0.01, compared to the sham group, #P<0.05, compared to the SNI group. All data were expressed as the mean±SEM, n=4 per group. Arrows indicate expressions of p-NF-κB and DAPI as well as co-localization of p-NF-κB with DAPI in the spinal dorsal horns.Abbreviations: NF-κB, nuclear factor kappa beta; EA, electroacupuncture; SNI, spared nerve injury; SEM, standard error of the mean. |
Discussion
Evidence has shown that HMGB1 exerts its pro-inflammatory effect to contribute to neuropathic pain through triggering TLR4 and adaptor protein MyD88, subsequently leading to activation of NF-κB signaling and resulting in pro-inflammatory cytokines production and pain hypersensitivity.23 In this study, 2Hz EA could ameliorate pain-related behaviors such as mechanical hypersensitivity and elevation of gait parameters in SNI rats, and subsequently reduced the upregulation of spinal HMGB1, TLR4, MyD88, and NF-κB p65 protein expression induced by SNI. Furthermore, we observed the downregulation of co-expression of TLR4 and CD11b, MyD88 and CD11b, as well as the inhibition of NF-κB p65 activation by 2Hz EA in SNI rats. Findings from this study provided novel insight into the molecular mechanism underlying 2Hz EA relieving neuropathic pain.
Neuropathic pain remains a major public health challenge that impacts millions of individuals. Novel and more effective managements should be further investigated. Numerous studies have confirmed that EA stimulation significantly alleviated neuropathic pain with few side effects.17,20 More importantly, EA stimulation produced analgesia greatly relied on EA frequency and EA stimulation interval.18 It was reported that 2Hz EA elicited better analgesic effect on neuropathic pain than 100Hz EA did,18,19 and administration of 2Hz EA to SNI rat once every other day lasting for 21 days induced a cumulative analgesic effect.24 Thus, in this study, 2Hz EA stimulation was selected to treat SNI-induced neuropathic pain as described in our previous studies.20,21 Similarly, 2Hz EA could relieve SNI-induced mechanical hypersensitivity. Additionally, CatWalk gait analysis was used to assess the alterations of gait parameters after SNI in this experiment.22 We found that 2Hz EA significantly improved SNI-induced decrease of gait parameters. These investigations implied that 2Hz EA could attenuate pain-related behaviors in SNI rats.
Currently, the mechanism underlying neuropathic pain remains poorly understood. Neuroinflammation plays a pivotal role in the development of neuropathic pain.25 HMGB1 was reported to be a critical regulator of inflammation and be activated under pathological conditions.26 Evidence showed that nerve injury led to the activation of spinal microglia and astrocytes.27 HMGB1 binds to TLR4 from glial cells and induces an inflammatory response.13 Moreover, administration of an anti-HMGB1 antibody alleviates neuropathic pain via blocking activation of glial cells in the spinal cord,24,27 which indicated that the interaction between HMGB1 and glial cells in the spinal cord contributes to neuropathic pain. Furthermore, the upregulation of spinal HMGB1 expression was found in nerve injury-induced neuropathic pain,24 suggesting that HMGB1 may be synthesized and secreted from spinal cord neurons and/or glial cells and leads to the development of neuropathic pain following nerve injury.2 Consistent with previous reports, our findings demonstrated that SNI elicited increase of HMGB1 protein expression in the spinal cord, which further implied that HMGB1 participated in SNI-induced neuropathic pain. It was shown that intrathecal injection of an antibody against HMGB1 blocked mechanical allodynia and reduced pro-inflammatory cytokines, such as IL-1β, IL-6, and TNF-α in the spinal cord,2 suggesting that a positive feedback may be presented between HMGB1 and pro-inflammatory cytokines, which may improve the effect of HMGB1 in neuropathic pain.2 We have previously reported that 2Hz EA decreased the upregulation of spinal IL-1β mRNA and protein expressions induced by SNI.17 Meanwhile, in this study, 2Hz EA markedly downregulated the level of spinal HMGB1 protein expression in SNI rats. This further confirmed that 2Hz EA stimulation exhibits its analgesic effect on neuropathic pain through reducing HMGB1 and pro-inflammatory cytokines expression in the spinal cord. Therefore, suppressing HMGB1 by 2Hz EA treatment may be a promising therapeutic strategy for neuropathic pain.
HMGB1 signals leading to neuropathic pain may be mainly dependent on either of two receptors for TLR4 and receptor for advanced glycation end products.28–30 Based on the other and our previous data, the present study focused on investigating whether HMGB1-TLR4 signaling participated in the effect of 2Hz EA on SNI-induced neuropathic pain. TLR4 has been indicated to play a critical role in neuropathic pain.31,32 Spinal microglia was involved in the initiation and maintenance of neuropathic pain,32–34 and TLR4 was predominantly expressed in spinal microglia,32,35 suggesting that TLR4 was closely associated with microglia activation in the pathogenesis of neuropathic pain. Consistently, in this study, we found that SNI induced the enhancement of spinal TLR4 protein expression. Moreover, double immunofluorescence staining results showed co-localization of TLR4 with microglia activation marker CD11b in the spinal cord, and the expression levels of both TLR4 and CD11b were also found to elevate in SNI rats, which indicated that SNI evoked microglia activation in the spinal cord. Overall, our findings further confirmed that spinal TLR4 played an important role in microglia activation and the induction of SNI-evoked neuropathic pain. Meanwhile, the present study demonstrated that 2Hz EA treatment could reduce the upregulation of spinal TLR4 protein expression induced by SNI, and immunofluorescence staining also revealed co-expression levels of TLR4 and CD11b were decreased by 2Hz EA in the spinal cord following SNI. These results suggested that 2Hz EA inhibited upregulation of TLR4 and microglia activation induced by SNI in the spinal cord.
It was reported that peripheral nerve injury significantly upregulated spinal MyD88 protein expression, and which was found to be present in the spinal microglia.20 Evidence showed that inhibition of MyD88-dependent signaling could alleviate peripheral nerve injury-elicited neuropathic pain.20,34 Furthermore, TLR4 triggered MyD88 to lead to the activation of NF-κB signaling and the release of pro-inflammatory cytokines, such as IL-1β.3,35,36 In accordance with previous studies, our results revealed that SNI significantly led to the increase of MyD88 protein expression in the spinal cord. Moreover, double immunofluorescence staining also showed spinal MyD88 was co-localized with CD11b, and both MyD88 and CD11b expressions were also upregulated in the spinal cord of SNI rats. These data suggested that spinal MyD88 indeed contributed to SNI-induced microglia activation and the development of neuropathic pain; whereas 2Hz EA treatment to SNI rats downregulated spinal MyD88 protein expression, and reduced the co-expression of MyD88 and CD11b in the spinal cord, which indicated inhibition of SNI-induced activation of both microglia and MyD88 signaling by 2Hz EA in the spinal cord.
Neuropathic pain due to peripheral nerve injury led to the release of pro-inflammatory mediators, which activated spinal microglia via TLR4/NF-κB signaling pathway.37 It was known that NF-κB is one of the most important transcription factors in the regulation of pro-inflammatory mediators.38 The activated NF-κB p65 was translocated to the nucleus and recruited to the targeted genes, and subsequently improved the transcription of those genes.38 Increasing evidence demonstrated that activation of NF-κB signaling participated in neuroinflammation through regulating the expression of pro-inflammatory cytokines.38,39 Suppression of NF-κB p65 expression significantly alleviated CCI-induced pain hypersensitivity.40 Collectively, these data demonstrated that NF-κB signaling takes part in the pathogenesis of neuropathic pain, and it was served as a molecular target for treating neuropathic pain.41 Our previous study has shown that 2Hz EA stimulation markedly decreased the upregulation of a pro-inflammatory cytokine IL-1β induced by SNI in the spinal cord.20 Thus, further investigation for the effect of NF-κB signaling in 2Hz EA ameliorating neuropathic pain is an interesting question. Consistent with previous reports, our results revealed the upregulation of spinal NF-κB p65 protein expression following SNI, and immunofluorescence staining also showed that SNI induced NF-κB p65 nuclear translocation in the spinal dorsal horn, which suggested SNI-evoked NF-κB activation in the spinal cord. While the present findings demonstrated that 2Hz EA decreased the upregulation of NF-κB p65 protein expression and inhibited NF-κB p65 nuclear translocation elicited by SNI in the spinal cord, suggesting that 2Hz EA could suppress the SNI-induced activation of NF-κB signaling in the spinal cord.
This study also existed some limitations. For example, suppression of HMGB1, TLR4, MyD88, NF-κB signaling, and microglia activation in the spinal cord of SNI rats by 2Hz EA treatment was taken into consideration. However, in this study, there were no further investigations to clarify the relationship between alterations of HMGB1, TLR4, MyD88, NF-κB p65 expression levels and attenuation of 2Hz EA on neuropathic pain with inhibitors, antibodies or antagonists.
Conclusion
The present study revealed that 2Hz EA treatment alleviated pain-related behaviors induced by SNI, such as ameliorating mechanical hypersensitivity and promoting gait parameters. Furthermore, 2Hz EA stimulation reduced the expression levels of HMGB1, TLR4, MyD88, and NF-κB p65 in the spinal cord of SNI rats. Thus, blockade of HMGB1 may be a novel therapeutic target for 2Hz EA alleviating SNI-induced neuropathic pain.
Acknowledgments
This work was supported by the grants from the National Natural Science Foundation of China (no. 31060144), and the Talent Project of Department of Scientific and Technology, Jiangxi Province, People’s Republic of China (no. 20142BCBC22008). The authors thank Dr Tao Gan in the Department of Biochemistry, Gannan Medical University, People’s Republic of China for his help in preparing this manuscript.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Moalem G, Tracey DJ. Immune and inflammatory mechanisms in neuropathic pain. Brain Res Rev. 2006;51(2):240–264. doi:10.1016/j.brainresrev.2005.11.004
2. Maeda T, Ozaki M, Kobayashi Y, Kiguchi N, Kishioka S. HMGB1 as a potential therapeutic target for neuropathic pain. J Pharmacol Sci. 2013;123(4):301–305. doi:10.1254/jphs.13r08cp
3. Chen XL, Sun L, Guo F, et al. High-mobility group box-1 induces proinflammatory cytokines production of Kupffer cells through TLRs-dependent signaling pathway after burn injury. PLoS One. 2012;7(11):e50668.
4. Chacur M, Milligan ED, Gazda LS, et al. A new model of sciatic inflammatory neuritis (SIN): induction of unilateral and bilateral mechanical allodynia following acute unilateral peri-sciatic immune activation in rats. Pain. 2001;94(3):231–244. doi:10.1016/s0304-3959(01)00354-2
5. Wan W, Cao L, Khanabdali R, Kalionis B, Tai X, Xia S. The emerging role of HMGB1 in neuropathic pain: a potential therapeutic target for neuroinflammation. J Immunol Res. 2016;2016:6430423. doi:10.1155/2016/6430423
6. Watkins LR, Milligan ED, Maier SF. Glial activation: a driving force for pathological pain. Trends Neurosci. 2001;24(8):450–455. doi:10.1016/s0166-2236(00)01854-3
7. Tanga FY, Nutile-McMenemy N, DeLeo JA. The CNS role of Toll-like receptor 4 in innate neuroimmunity and painful neuropathy. Proc Natl Acad Sci U S A. 2005;102(16):5856–5861. doi:10.1073/pnas.0501634102
8. Cao L, Tanga FY, Deleo JA. The contributing role of CD14 in toll-like receptor 4 dependent neuropathic pain. Neuroscience. 2009;158(2):896–903. doi:10.1016/j.neuroscience.2008.10.004
9. Lehnardt S, Massillon L, Follett P, et al. Activation of innate immunity in the CNS triggers neurodegeneration through a Toll-like receptor 4-dependent pathway. Proc Natl Acad Sci U S A. 2003;100(14):8514–8519. doi:10.1073/pnas.1432609100
10. Piao Y, Gwon DH, Kang DW, et al. TLR4-mediated autophagic impairment contributes to neuropathic pain in chronic constriction injury mice. Mol Brain. 2018;11(1):11. doi:10.1186/s13041-018-0354-y
11. Medzhitov R, Preston-Hurlburt P, Kopp E, et al. MyD88 is an adaptor protein in the hToll/IL-1 receptor family signaling pathways. Mol Cell. 1998;2(2):253–258. doi:10.1016/s1097-2765(00)80136-7
12. Liu F, Wang Z, Qiu Y, et al. Suppression of MyD88-dependent signaling alleviates neuropathic pain induced by peripheral nerve injury in the rat. J Neuroinflammation. 2017;14(1):70. doi:10.1186/s12974-017-0822-9
13. Grace PM, Hutchinson MR, Maier SF, Watkins LR. Pathological pain and the neuroimmune interface. Nat Rev Immunol. 2014;14(4):217–231. doi:10.1038/nri3621
14. Fonken LK, Frank MG, Kitt MM, et al. The alarmin HMGB1 mediates age-induced neuroinflammatory priming. J Neurosci. 2016;36(30):7946–7956. doi:10.1523/JNEUROSCI.1161-16.2016
15. Basbaum AI, Bautista DM, Scherrer G, Julius D. Cellular and molecular mechanisms of pain. Cell. 2009;139(2):267–284. doi:10.1016/j.cell.2009.09.028
16. Varrassi G, Muller-Schwefe G, Pergolizzi J, et al. Pharmacological treatment of chronic pain - the need for CHANGE. Curr Med Res Opin. 2010;26(5):1231–1245. doi:10.1185/03007991003689175
17. Huang C, Li HT, Shi YS, Han JS, Wan Y. Ketamine potentiates the effect of electroacupuncture on mechanical allodynia in a rat model of neuropathic pain. Neurosci Lett. 2004;368(3):327–331. doi:10.1016/j.neulet.2004.07.073
18. Han JS. Acupuncture: neuropeptide release produced by electrical stimulation of different frequencies. Trends Neurosci. 2003;26(1):17–22. doi:10.1016/s0166-2236(02)00006-1
19. Zhang R, Lao L, Ren K, Berman BM. Mechanisms of acupuncture-electroacupuncture on persistent pain. Anesthesiology. 2014;120(2):482–503. doi:10.1097/ALN.0000000000000101
20. Wang Y, Jiang Q, Xia YY, Huang ZH, Huang C. Involvement of alpha7nAChR in electroacupuncture relieving neuropathic pain in the spinal cord of rat with spared nerve injury. Brain Res Bull. 2018;137:257–264. doi:10.1016/j.brainresbull.2018.01.002
21. Wang Y, Xue M, Xia Y, Jiang Q, Huang Z, Huang C. Electroacupuncture treatment upregulates alpha7nAChR and inhibits JAK2/STAT3 in dorsal root ganglion of rat with spared nerve injury. J Pain Res. 2019;12:1947–1955. doi:10.2147/JPR.S203867
22. Chen YJ, Cheng FC, Sheu ML, et al. Detection of subtle neurological alterations by the Catwalk XT gait analysis system. J Neuroeng Rehabil. 2014;11:62. doi:10.1186/1743-0003-11-62
23. da Silva JR, da Silva ML, Prado WA. Electroacupuncture at 2/100 hz activates antinociceptive spinal mechanisms different from those activated by electroacupuncture at 2 and 100 hz in responder rats. Evid Based Complement Alternat Med. 2013;2013:205316. doi:10.1155/2013/205316
24. Ren PC, Zhang Y, Zhang XD, et al. High-mobility group box 1 contributes to mechanical allodynia and spinal astrocytic activation in a mouse model of type 2 diabetes. Brain Res Bull. 2012;88(4):332–337. doi:10.1016/j.brainresbull.2012.03.002
25. Reichling DB, Green PG, Levine JD. The fundamental unit of pain is the cell. Pain. 2013;154 Suppl 1.
26. Lu B, Wang C, Wang M, et al. Molecular mechanism and therapeutic modulation of high mobility group box 1 release and action: an updated review. Expert Rev Clin Immunol. 2014;10(6):713–727. doi:10.1586/1744666X.2014.909730
27. Nakamura Y, Morioka N, Abe H, et al. Neuropathic pain in rats with a partial sciatic nerve ligation is alleviated by intravenous injection of monoclonal antibody to high mobility group box-1. PLoS One. 2013;8(8):e73640. doi:10.1371/journal.pone.0073640
28. Zhu XM, Yao YM, Liang HP, et al. High mobility group box-1 protein regulate immunosuppression of regulatory T cells through toll-like receptor 4. Cytokine. 2011;54(3):296–304. doi:10.1016/j.cyto.2011.02.017
29. Tian J, Avalos AM, Mao SY, et al. Toll-like receptor 9-dependent activation by DNA-containing immune complexes is mediated by HMGB1 and RAGE. Nat Immunol. 2007;8(5):487–496. doi:10.1038/ni1457
30. Mazarati A, Maroso M, Iori V, Vezzani A, Carli M. High-mobility group box-1 impairs memory in mice through both toll-like receptor 4 and receptor for advanced glycation end products. Exp Neurol. 2011;232(2):143–148. doi:10.1016/j.expneurol.2011.08.012
31. Su M, Ran Y, He Z, et al. Inhibition of toll-like receptor 4 alleviates hyperalgesia induced by acute dural inflammation in experimental migraine. Mol Pain. 2018;14:1744806918754612. doi:10.1177/1744806918754612
32. Kim D, Kim MA, Cho IH, et al. A critical role of toll-like receptor 2 in nerve injury-induced spinal cord glial cell activation and pain hypersensitivity. J Biol Chem. 2007;282(20):14975–14983. doi:10.1074/jbc.M607277200
33. Bettoni I, Comelli F, Rossini C, et al. Glial TLR4 receptor as new target to treat neuropathic pain: efficacy of a new receptor antagonist in a model of peripheral nerve injury in mice. Glia. 2008;56(12):1312–1319. doi:10.1002/glia.20699
34. Wang Z, Liu F, Wei M, et al. Chronic constriction injury-induced microRNA-146a-5p alleviates neuropathic pain through suppression of IRAK1/TRAF6 signaling pathway. J Neuroinflammation. 2018;15(1):179. doi:10.1186/s12974-018-1220-7
35. Peirs C, Seal RP. Targeting Toll-like receptors to treat chronic pain. Nat Med. 2015;21(11):1251–1252. doi:10.1038/nm.3986
36. Muzio M, Ni J, Feng P, Dixit VM. IRAK (Pelle) family member IRAK-2 and MyD88 as proximal mediators of IL-1 signaling. Science. 1997;278(5343):1612–1615. doi:10.1126/science.278.5343.1612
37. Lin JJ, Du Y, Cai WK, et al. Toll-like receptor 4 signaling in neurons of trigeminal ganglion contributes to nociception induced by acute pulpitis in rats. Sci Rep. 2015;5:12549. doi:10.1038/srep12549
38. Sakaue G, Shimaoka M, Fukuoka T, et al. NF-kappa B decoy suppresses cytokine expression and thermal hyperalgesia in a rat neuropathic pain model. Neuroreport. 2001;12(10):2079–2084. doi:10.1097/00001756-200107200-00008
39. Maeda S, Omata M. Inflammation and cancer: role of nuclear factor-kappaB activation. Cancer Sci. 2008;99(5):836–842. doi:10.1111/j.1349-7006.2008.00763.x
40. Sun T, Song WG, Fu ZJ, Liu ZH, Liu YM, Yao SL. Alleviation of neuropathic pain by intrathecal injection of antisense oligonucleotides to p65 subunit of NF-kappaB. Br J Anaesth. 2006;97(4):553–558. doi:10.1093/bja/ael209
41. Niederberger E, Geisslinger G. The IKK-NF-kappaB pathway: a source for novel molecular drug targets in pain therapy? Faseb J. 2008;22(10):3432–3442. doi:10.1096/fj.08-109355
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.