")
Back to Journals » Infection and Drug Resistance » Volume 16
Effects of Repeated in-vitro Exposure to Saudi Honey on Bacterial Resistance to Antibiotics and Biofilm Formation
Authors Aldarhami A , Bazaid AS , Qanash H , Ahmad I, Alshammari FH, Alshammari AM, Alshammari AH, Aljanfawe FM, Aldamiri B, Aldawood E , Alghamdi MA, Binsaleh NK , Saeedi NH, Snoussi M
Received 26 February 2023
Accepted for publication 28 June 2023
Published 3 July 2023 Volume 2023:16 Pages 4273—4283
DOI https://doi.org/10.2147/IDR.S410159
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Professor Suresh Antony
Abdu Aldarhami,1 Abdulrahman S Bazaid,2,3 Husam Qanash,2,3 Iqbal Ahmad,4 Fahad H Alshammari,2,3 Abdulrahman M Alshammari,2,3 Abdulrahman H Alshammari,2,3 Fahad M Aljanfawe,3 Bushra Aldamiri,5 Esraa Aldawood,6 Mashail A Alghamdi,7 Naif K Binsaleh,2,3 Nizar H Saeedi,8 Mejdi Snoussi3,9,10
1Department of Medical Microbiology, Qunfudah Faculty of Medicine, Umm Al-Qura University, Al-Qunfudah, 21961, Saudi Arabia; 2Department of Medical Laboratory Science, College of Applied Medical Sciences, University of Ha’il, Hail, 55476, Saudi Arabia; 3Medical and Diagnostic Research Center, University of Ha’il, Hail, 55473, Saudi Arabia; 4Department of Agricultural Microbiology, Faculty of Agricultural Sciences, Aligarh Muslim University, Aligarh, 202002, India; 5Biochemistry Department, College of Science, King Saud University, Riyadh, 12372, Saudi Arabia; 6Clinical Laboratory Sciences Department, College of Applied Medical Sciences, King Saud University, Riyadh, 12372, Saudi Arabia; 7Biology Department, Faculty of Science, King Abdulaziz University, Jeddah, 21589, Saudi Arabia; 8Department of Medical Laboratory Technology, Faculty of Applied Medical Sciences, University of Tabuk, Tabuk, Saudi Arabia; 9Department of Biology, College of Science, University of Ha’il, Hail, 81451, Saudi Arabia; 10Laboratory of Genetics, Biodiversity and Valorization of Bio-Resources, Higher Institute of Biotechnology of Monastir, University of Monastir, Monastir, 5000, Tunisia
Correspondence: Abdulrahman S Bazaid; Husam Qanash, Department of Medical Laboratory Science, College of Applied Medical Sciences, University of Ha’il, Hail, 55476, Saudi Arabia, Tel +966 16 5358200 ext. 1713, Email [email protected]; [email protected]
Introduction: Although Sumra and Sidr Saudi honey is widely used in traditional medicine due to its potent activity, it is unknown whether its prolonged usage has impact upon bacterial virulence or leading to reduced antibiotic sensitivity. Thus, the study aims to investigate the effect of prolonged (repeated) in-vitro exposure to Saudi honey on the antibiotic susceptibility profiles and biofilm formation of pathogenic bacteria.
Methods: Several bacteria, including Staphylococcus aureus, Escherichia coli, and Acinetobacter baumannii, were in-vitro exposed ten times [passaged (P10)]to Sumra and Sider honey individually to introduce adapted bacteria (P10). Antibiotic susceptibility profiles of untreated (P0) and adapted (P10) bacteria were assessed using disc diffusion and microdilution assays. The tendency regarding biofilm formation following in-vitro exposure to honey (P10) was assessed using the Crystal violet staining method.
Results: Adapted (P10) bacteria to both Sumra and Sidr honey showed an increased sensitivity to gentamicin, ceftazidime, ampicillin, amoxycillin/clavulanic acid, and ceftriaxone, when compared with the parent strains (P0). In addition, A. baumannii (P10) that was adapted to Sidr honey displayed a 4-fold increase in the minimal inhibitory concentration of the same honey following in-vitro exposure. 3-fold reduction in the tendency toward biofilm formation was observed for the Sumra-adapted (P10) methicillin resistant S. aureus strain, although there was a lower rate of reduction (1.5-fold) in biofilm formation by both the Sumra- and Sidr-adapted A. baumannii (P10) strains.
Conclusion: The data highlight the positive impact of prolonged in-vitro exposure to Saudi honey (Sumra and Sider) for wound-associated bacteria since they displayed a significant increase in their sensitivity profiles to the tested antibiotic and a reduction in their ability to form biofilm. The increased bacterial sensitivity to antibiotics and a limited tendency toward biofilm formation would suggest the great potential therapeutic use of this Saudi honey (Sumra and Sidr) to treat wound infections.
Keywords: Sider and Sumra honey, Saudi honey, resistance, biofilm formation, antibiotic susceptibility
Introduction
Antimicrobial resistance (AMR) is a well-defined, continuously emerging phenomenon that poses frightening threats and challenges to human health worldwide.1,2 This is mainly due to the emergence bacterial strains that cannot be treated using the currently available drugs. These are known as multidrug-resistant (MDR) bacteria or “superbugs”.1 Multiple factors, including globalization and poor cleanliness, might trigger or enhance the emergence of AMR, although the term “antibiotics misuse” is most often claimed to be the predisposing factor regarding the establishment and development of AMR.1 The misuse of antibiotics includes the under and over usage of these drugs by different means, such as the incorrect duration or dosage of the prescribed antibacterial agents in different sectors (eg, veterinary medicine, agriculture and aquaculture), including human health.3 In addition, over-the-counter sales, or unguided empirical prescriptions of antibiotics by doctors/specialists, where there was no real need for their use (eg, for patients with a viral infection), are commonly observed examples of unprofessional practices globally across different sectors.4 AMR has led to a worldwide health and economic burden because infections caused by MDR bacteria are associated with the highest rate of mortality and economic losses.1,5,6 In 2006, it was reported that about 50,000 deaths were attributed to hospital-acquired infections (HAIs) caused by drug-resistant strains in the United States of America (USA) alone.7 It was also estimated that globally, by 2050, about 10 million deaths annually will be caused by AMR, unless targeted actions are implemented prior to this date. Furthermore, AMR increases the annual cost of healthcare by 20 billion dollars in the USA alone.8
Although multiple actions are taken to combat the AMR, scientists have focused on antibiotic-alternative approaches and therapies, due to the high probability of their potential effectiveness and limited development of resistance compared with conventional antibiotics.1 These approaches would include the use of plant-based products (eg, honey) as an antibacterial agent to treat drug-resistant bacteria, such as Manuka, or Leptospermum scoparium honey, which is named after a species of tea tree that is native to New Zealand.9 Manuka honey displayed activity toward multiple Gram-positive and negative pathogenic drug resistant bacteria, including Escherichia coli, methicillin-resistant S. aureus (MRSA), antibiotic resistant beta-hemolytic streptococci and vancomycin-resistant Enterococci (VRE).9,10 Despite the considerable tolerance of biofilm to antibiotics and its great challenge to medicine, an in-vitro study showed the successful eradication of biofilm growth by P. aeruginosa and S. aureus when using Manuka honey.11,12 Interestingly, the sensitivity of Manuka-treated pathogenic bacteria increased towards conventional antibiotics, compared with untreated ones.13
Saudi Arabia produces multiple types of honey, that according to the tree, nectar and/or location of hives, including Sider, Talah and Sumra honey.14 Sider and Sumra honey are commonly used in traditional medicine to treat various infections as well as gastric ulcers, probably due to their antioxidant, anti-inflammatory, and anti-bacterial properties.15 Sider honey, with an 80% w/v concentration, displayed potent activity against S. aureus, S. epidermidis and Shigella flexneri, while Dharm honey of the same concentration showed a superior inhibitory effect toward E. coli, Proteus mirabilis, S. epidermidis and Shigella flexneri.16 In addition, the promising sensitivity of Gram-positive and negative bacteria including B. cereus, S. aureus, E. coli, Salmonella enterica as well as Trichophyton mentagrophytes (Fungi) toward Sider honey was also monitored. Moreover, Saudi Sumra honey has been proven to be effective against multidrug-resistant (MDR) S. aureus, E. coli, and Candida albicans in single and/or polymicrobial cultures.17
Although the above-mentioned studies have highlighted the promising activity of Saudi honey and its historical usage in traditional medicine to treat infectious diseases, there is a clear research gap concerning the effect of the prolonged, repeated in-vitro exposure of Saudi honey on bacterial resistance to conventional antibiotics, as well as its ability to create biofilm. This in-vitro study is essential for any potential therapeutic use of Saudi honey to treat infections caused by MDR and biofilm-forming pathogenic bacteria. Accordingly, the current study aims to investigate the antibiotic susceptibility and biofilm formation of honey-treated (adapted) pathogenic bacteria following repeated in-vitro exposure to various types of Saudi honey in comparison with the same parent (untreated) bacterial strains.
Materials and Methods
Collection of Bacterial Isolates, Honey Samples and Antibiotic Discs
Several Gram-positive (n=7) and negative (n= 6) clinical and reference bacterial isolates that are associated with wound infections were used in this study, including Staphylococcus aureus and Staphylococcus epidermidis, as Gram-positive indicators, while Acinetobacter baumannii, Escherichia coli and Pseudomonas aeruginosa were the Gram-negative strains. Clinical isolates were collected from King Khalid hospital in Hail, Saudi Arabia, as part of the routine diagnosis procedure. To generate data with decent reliability and reproducibility for practical comparison, reference Gram-positive and negative bacterial strains were purchased from the American Type Culture Collection (ATCC) (Manassas, United States) and National Collection of Type Cultures [(NCTC) Public Health England, Salisbury, United Kingdom)]. All of the isolates were recovered, cultured and/or maintained in a Mueller-Hinton agar (MHA) and/or Mueller-Hinton broth (MHB) (Thermo-Fisher Scientific, Massachusetts, United States).
Sumra and Sider honey was obtained from an authenticated company that sold honey in Al-Bah city, Saudi Arabia (Al Amari, AL-Baha, Saudi Arabia).17 In addition, medical grade Manuka honey (Methylglyoxal 400) was purchased from MaunukaGuard (Monterey, California, USA),9 which was utilized as a positive control. All of the obtained honey was inoculated into MHA plates and incubated at 37°C for 24 hours to ensure the absence of any contamination (a purity check), then prepared as a 90% stock solution using sterile distilled water (dH2O) before any experimental work was conducted. Antibiotic discs, including gentamicin (120 μg), ceftazidime (30 μg), ampicillin (10 μg), amoxycillin/clavulanic (30 μg), ceftriaxone (30 μg), and imipenem (10 μg), were obtained from Condalab (Madrid, Spain) and stored at −20°C in a dry place.
Long-Term Exposure of Bacterial Isolates to Honey Samples (Adaptation Process)
To evaluate the effect of the long-term, in-vitro exposure of bacterial isolates to various honey samples, each individual bacterium was repeatedly exposed (ten times) to each tested honey sample (Sumra, Sider and Manuka) using an agar-based diffusion method.13 In brief, approximately 6-millimeter (mm) wells were formed at the center of a Mueller Hinton agar plates, after which 200 μL of a sub-lethal concentration of each tested honey (30 mg/mL dissolved in sterile dH2O) was added aseptically into the center of the well/s. Each parent (unexposed) bacterial isolate [Passage zero (P0)] was distributed radially in triplicate around the central well and incubated at 37°C for 24 hours. Following incubation, the bacteria that had grown in close proximity to the well were aseptically re-inoculated onto a plate containing the same or a higher concentration of the tested honey. This procedure was repeated for a total of ten passages to generate the adapted bacteria (P10) for each type of honey tested. The parent strains (P0) were also sub-cultured ten times in MHA (without any honey) as a negative control [(P0) negative control)] to ensure that any potential changes in the antibiotic susceptibility of the adapted bacteria (P10) could be attributed to the honey exposure alone (the treatment) rather than the sub-culturing process itself.
Antibiotic Susceptibility Testing
To assess the effect of honey on bacterial sensitivity toward antibiotics, the antibiotic susceptibility profiles of the isolates prior to their exposure to honey [(P0) negative control)] as well as those of the isolates that had been treated ten times with honey (P10) were determined.18 This was performed according to the standardized British Society for Antimicrobial Chemotherapy (BSAC) disc diffusion method for antimicrobial susceptibility testing using MHA plates.18 The susceptibility profiles of all of the Gram-positive isolates were determined against five different antibiotics, including gentamicin, ceftazidime, ampicillin, amoxycillin/clavulanic and ceftriaxone, whereas imipenem in addition to these five antibiotics were used for the evaluation of the Gram-negative isolates. The plates were incubated for 24 hours at 37°C, after which the zones of inhibition were measured in millimeters (mm). This testing was carried out in triplicate for each bacterium to provide more reliable and reproducible data. The average of the three reads and standard deviations were also calculated.
Antibacterial Activity of Various Types of Honey Against Adapted (Honey-Treated) and Parent (Untreated) Bacterial Strains
The minimum inhibitory concentrations (MICs) and minimum bactericidal concentrations (MBCs) of Sider and Sumra honey against adapted clinical isolates (P10) were assessed using a broth microdilution method.18 Overnight bacterial cultures of clinical isolates with zero exposure (P0) as well as isolates with long-exposure (P10) were adjusted to a specific optical density (OD600 of 0.8), known as 0.5 MacFarland, followed by a 1:100 dilution using the MHB. Next, after the bacterial suspension (100 µL) had been transferred to a 96-well microtiter plate, multiple concentrations (0–1000 mg/L) of the tested honey were added. The inoculated plates were incubated aerobically at 37°C for 24 hours. Then, the MIC values were determined visually, which is the lowest concentration of tested honey at which the bacterial growth was inhibited. Only then was the MBC assessed by transferring aliquots of 5 μL from wells exhibiting no visible turbidity onto the MHA plate, followed by overnight incubation at 37°C. The MBC values were determined as the lowest concentration of tested honey that led to the complete absence of bacterial growth on the MHA plate.
Biofilm Formation Assay
A crystal violet assay was used to assess the biofilm formation by the clinical bacterial strains that had been treated with each type of honey (P10) and the untreated isolates [(P0) negative control].19 The overnight bacterial cultures were diluted (1:100) using MHB, then transferred to a sterile 96-well microtiter plate and incubated at 37°C for 48 hours. Next, the bacterial suspension was discarded, and the wells were washed twice in a sterile phosphate-buffered saline (PBS) (Thermo-Fisher Scientific, Massachusetts, USA) before staining with 250 μL of 1% (w/v) crystal violet solution (Sigma-Aldrich, Missouri, USAT) at room temperature until dry. Ethanol was added to each well and the absorbance (OD600) was monitored using a plate reader. The experiments were performed in triplicate and the data for the isolates with zero exposure (P0) were compared with the data for the treated strains (P10).
Statistical Analysis
The statistical analysis was conducted using GraphPad Prism (version 9). The analysis was performed on biofilm formation essay reads to determine any potential significant changes between the parent (P0) (untreated) and adapted (honey-treated) bacteria (P10) using the t-test.
Results
Effect of Honey Exposure on Bacterial Sensitivity to Antibiotics
In this study, certain honey-treated bacterial strains, that had previously been exposed in-vitro ten times to a single type of honey, showed a significant increase in the size (mm) of their caused zone of inhibition (ZOI) to the tested antibiotics, which varied from one type of honey to another. Most of the significant changes were observed in the tested reference strains compared to the clinical isolates (Figures 1 and 2). The sensitivity data for each of the tested antibiotics to the types of honey used will be presented in the following sections separately.
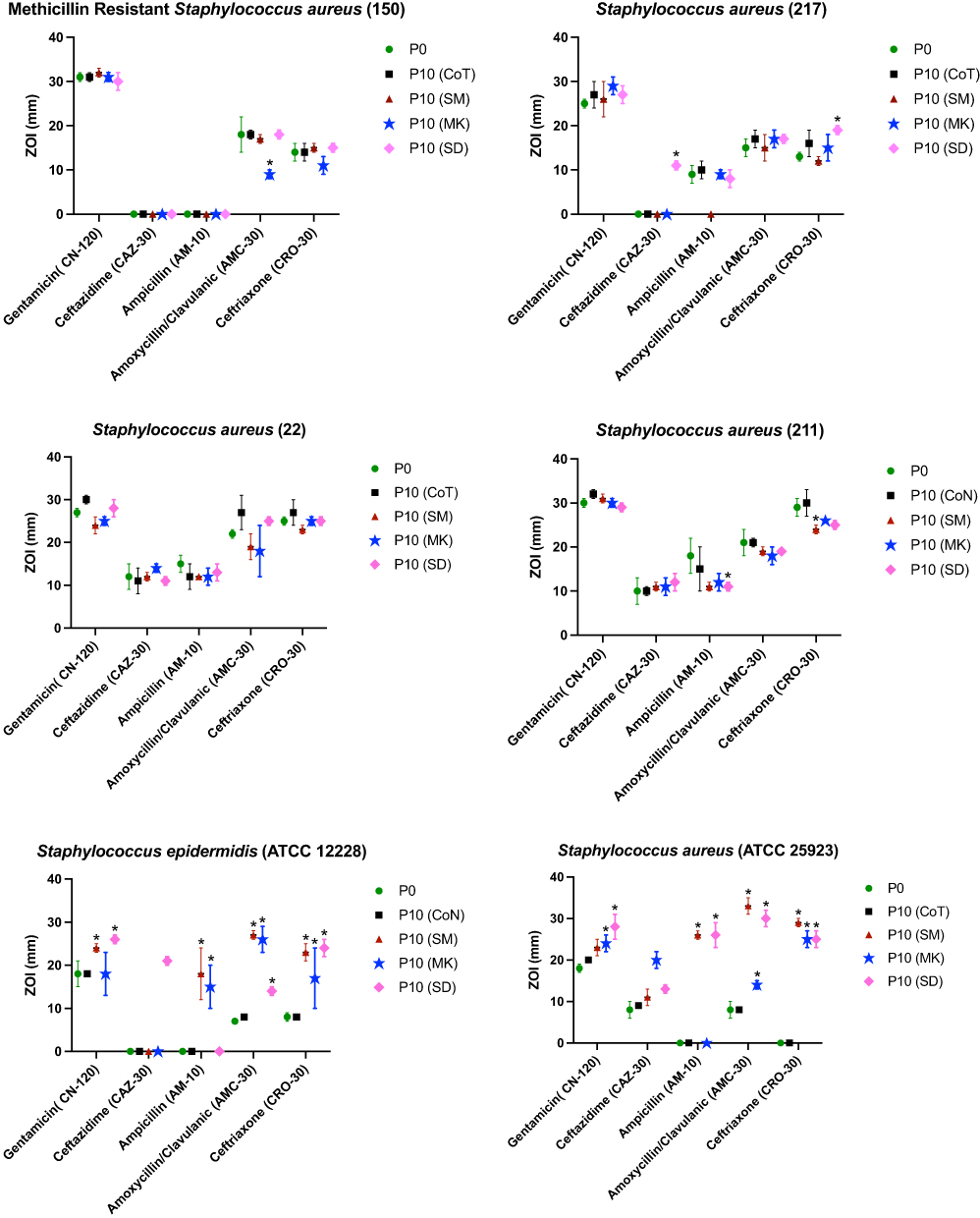 |
Figure 1 Antibiotic susceptibility profiles of Gram-positive bacteria treated ten times (passage 10 (P10)) with different honeys including, Manuka (MK), Sumra (SM) and Sider (SD), as well as parent (untreated) isolates [passage 0 (P0)] and controls for passaging procedure without any treatment (control (P10-CoT). |
 |
Figure 2 Antibiotic susceptibility profiles of Gram-negative bacteria treated ten times [passage 10 (P10)] with different honeys including, Manuka (MK), Sumra (SM) and Sider (SD), as well as parent (untreated) isolates [passage 0 (P0)] and controls for passaging procedure without any treatment [passage control (P10-CoT). |
Alteration in the Susceptibility to Gentamicin
The S. epidermidis [ATCC 12228] that was treated with Sumra and Sider honey showed an increased sensitivity to gentamicin. The largest inhibitory zones to gentamicin were observed in the adapted S. aureus [ATCC 25923], that had been exposed ten times to Manuka and Sider honey individually, while S. pyogenes [ATCC 19615] showed an increased susceptibility to gentamicin following exposure to all types of the honeys tested separately. The Gram-negative strains that had been treated with Manuka honey exhibited a noticeable reduction in their resistance to gentamicin. The adapted (P10) E. coli (ATCC 14169) and E. coli (ATCC 12923) strains displayed a larger zone of inhibition following long exposure (P10) to Manuka and Sider honey, respectively.
Alteration in the Susceptibility to Ceftazidime
Treatment with Sider honey resulted in greater sensitivity toward ceftazidime among the clinical isolates of S. aureus (217), S. epidermidis (ATCC 12228) and S. aureus (ATCC 25923). S. aureus (ATCC 25923) that had been treated (P10) with Manuka honey and S. pyogenes (ATCC 19615) that had been treated with Sumra honey showed a similar increase in their sensitivity to ceftazidime. All of the remaining adapted bacteria, (P10) revealed inhibitory zones to ceftazidime of a similar size to those of the parent strains (P0). In addition, all of the adapted (P10) reference strains of E. coli had larger ZOIs to ceftazidime (from around 8 mm to over 20 mm) following exposure to all three types of honey individually (Sumra, Manuka, and Sider). Moreover, the treated strains of P. aeruginosa (NCTC12903) became more susceptible to ceftazidime having been exposed to Manuka honey ten times.
Alteration in Susceptibility to Ampicillin
Adapted (P10) S. aureus (211) exhibited a reduced susceptibility to ampicillin following exposure to all of the types of honey tested. In contrast, although S. epidermidis (ATCC 12228) was fully resistant to ampicillin, it showed a larger ZOI following treatment with either Sumra or Manuka honey. Noticeably, S. aureus (ATCC 25923) that was exposed (P10) to Sumra and Sider honey separately became sensitive to ampicillin, with an inhibitory zone (26 mm) of identical size compared to full resistance (ZOI of 0 mm) prior to exposure. In addition, S. pyogenes (ATCC 19615) became more sensitive to ampicillin following exposure to Manuka honey. The susceptibility of all of the tested Gram-negative bacteria remained unchanged, except for that of P. aeruginosa (NCTC12903), which had been completely resistant to ampicillin previously, but became sensitive to it, with a large ZOI following in-vitro exposure (P10) to Sumra and Sider honey, individually.
Alteration in Susceptibility to Amoxycillin/Clavulanic
Methicillin-resistant S. aureus (150) that was treated (P10) with Manuka honey showed a slight decrease in its ZOI. All of the tested reference isolates of S. aureus displayed significant changes in their sensitivity profiles to this antibiotic following exposure to the tested types of honey. S. aureus (ATCC 25923) and S. epidermidis (ATCC 12228), having been adapted (P10) to all three types of honey, an exhibited significant increase in the size of their inhibitory zones caused by amoxycillin/clavulanic, although larger zones by the same antibiotic were monitored for S. pyogenes (ATCC 19615) following long exposure (P10) to Manuka honey. The most noticeable change in susceptibility was observed for the isolates that had been treated with Sider honey, including E. coli (NCTC 10418), (E. coli ATCC 14169), (E. coli NCTC 12923) and P. aeruginosa (NCTC 12903), as they become sensitive to amoxycillin/clavulanic, compared to their complete resistance to it prior to their exposure to Sider honey. In addition, adapted E. coli (NCTC 12923) and P. aeruginosa (NCTC12903) showed greater sensitivity to this antibiotic following exposure to Manuka honey.
Alteration in Susceptibility to Ceftriaxone
S. aureus (211), S. aureus (ATCC 25923), S. epidermidis (ATCC 12228) and S. pyogenes (ATCC 19615) that had been exposed to Sumra honey presented with a high susceptibility to ceftriaxone. Additionally, elevated activity by ceftriaxone was observed against S. aureus (ATCC 25923) and S. epidermidis (ATCC 12228), that had been treated ten times with Manuka and Sider honey, individually. Moreover, S. pyogenes (ATCC 19615) displayed a larger inhibitory zone following in-vitro exposure to Manuka honey alone. All of the adapted reference isolates of E. coli (NCTC 10418), E. coli (ATCC 14169), E. coli (NCTC 12923) and P. aeruginosa (NCTC12903) exhibited an increased sensitivity to ceftriaxone following exposure to all of the types of honey tested.
Alteration in Susceptibility to Imipenem
The activity of imipenem was tested against Gram-negative clinical and reference isolates, which revealed complete resistance and promising inhibition, respectively. There were no significant changes in the sensitivity profiles towards imipenem for any of the adapted bacteria (P10) to the three types of honey tested, except for P. aeruginosa (NCTC12903), that displayed a larger inhibitory zone caused by imipenem following in-vitro exposure to Sider and Sumra honey separately.
Changes in the Honey’s Antibacterial Activity Profiles Following Exposure to Sider and Sumra Honey
The number of MICs and MBCS of Gram-negative bacteria were slightly increased in the adapted bacteria (P10) compared to the parent strain (P0) against both Sider and Sumra honeys. A. baumannii (20) that had been exposed (P10) to Sider honey displayed a 4-fold increase in its MIC value as well as slightly higher MBCs in comparison with the MIC and MBC values of the parent strain against the same honey. The Gram-positive bacteria showed no changes in their MIC values to both types of honey while the values for the MBCs were somewhat elevated.
Effect of Bacterial Adaption to Honey on Biofilm Formation
The Sumra and Sider honey-adapted bacterial strains were tested for biofilm formation by comparing their adherence to the used surface with an untreated (P0) bacterial strain (Figure 3). The following bacterial strains were tested: S. aureus (211), S. aureus (22), MRSA, S. aureus (217), E. coli (41) and A. baumannii (20). An approximately 3-fold reduction in biofilm formation among Sumra-adapted MRSA (p value=0.011) was observed. Moreover, there was an approximately 1.5-fold reduction in biofilm formation among both the Sumra and Sider-adapted A. baumannii (20) (p value=0.008 and 0.003 respectively).
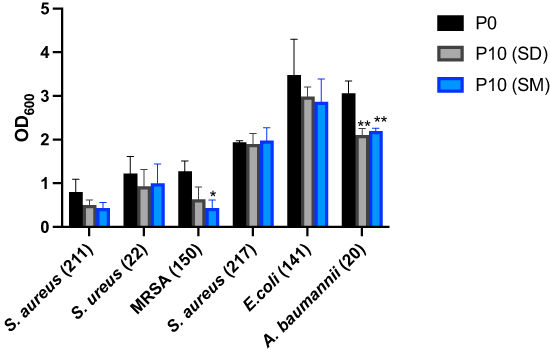 |
Figure 3 The effect of bacterial adaptation to Sider and Sumra honey upon its formation of biofilm. Sumra honey-adapted Methicillin-resistant Staphylococcus aureus (150) and Sumra honey-adapted Acinetobacter baumannii (20) resulted in a significant reduction of biofilm formation compared to the control, evidenced by reduced optical density. Also, Sider honey-adapted Acinetobacter baumannii (20) showed great inhibition of biofilm formation when compared to the control. Error bars represent the standard error of the mean. The results represent three (triplicates) independent experimental replicates. Statistical significances are *Indicated by and **When the p value ≤ 0.05 and ≤ 0.01, respectively. |
Discussion
Honey is well-valued and considered as one of nature’s finest gifts. Honey has been used for medicinal and dietary purposes for a long time in different nations all over the world.20 In conventional medicine, honey is used to treat/control various non-infectious illness (eg, cancer and asthma) and numerus infectious diseases, including, throat and wound infections, tuberculosis, and hepatitis.21 Honey contains hundreds of compounds, such as amino acids, phenol, sugar, minerals, flavonoids, vitamins, and antioxidants.17 Sumra and Sider are famous types of honey in Arab countries, and their antimicrobial activity has been documented against a wide range of pathogens, including drug-resistant bacteria and fungi.17 Honey’s antimicrobial activity has been attributed to several factors, such as the disruption of the bacterial cell membrane.22 However, no information about potential bacterial resistance to honey has ever been reported.23 Although, bacterial adaption to Manuka honey has been well-described13 and studied, there is no published data regarding the adaptation effect of Saudi honey (the ≥ ten times in-vitro exposure of the tested types of Saudi honey to bacterial strains individually) on its antibiotic sensitivity profiles and ability regarding biofilm formation. Therefore, the current project aimed to assess any potential changes in antibiotic sensitivity and biofilm formation for several clinical isolates following long-term in-vitro exposure to Sider and Sumra honey, individually.
Several clinical isolates were repeatedly exposed (ten times) to Sumra and Sider honey separately to generate the adapted bacteria (P10), after which their antibiotic susceptibility and biofilm formation were evaluated. The majority of adapted bacterial strains (P10) showed an increased sensitivity based on their zone size to five different antibiotics following repeated exposure to honey, although these changes were commonly observed in the adapted reference strains rather than the clinical isolates (Figures 1 and 2). This could be because reference isolates are less historically exposed to antimicrobial compounds, including honey (less selection pressure for resistance) compared with clinical isolates that have been treated with various doses of numerous antibiotic drugs.17 Thus, the effect of in-vitro exposure to honey on the reference strains in this study is more obvious and significant than that for the clinical strains. This suggests that further or longer exposure/treatment of the tested clinical isolates to these types of Saudi honey could have a more positive effect upon their sensitivity.
The obtained data show that the majority of the bacteria that adapted to Saudi honey became more sensitive to the cell wall inhibitors, including ceftazidime, ampicillin, amoxycillin/clavulanic, ceftriaxone, and imipenem (Figures 1 and 2). In line with a previous report,24 the increased sensitivity of the adapted bacteria to these antibiotics might be due to possible alterations in their cell wall permeability25 or/and efflux systems,26 following treatment with the tested honey. It was claimed that the in-vitro exposure of bacterial isolates to an antimicrobial agent (eg, honey) can affect cell membrane integrity, which has been observed in bacteria that have been adapted to antimicrobials.18,26 For instance, exposure to the antibiotic gentamicin has led to changes in cell wall permeability.27 Another possible cause of the increased sensitivity of the adapted bacteria to antibiotics could be that the repeated in-vitro exposure to the honey used (known as the adaptation process) leads to a reduction in bacterial growth and fitness,28 which in turns causes the adapted bacteria to become more sensitive to antibiotics.
The MICs and MBCs values of the Gram-negative and positive adapted bacteria against both types of honey tested (Sumra and Sider) were identical to those of the parent strains, except for A. baumannii (20) that was adapted (P10) to Sider honey, since a 4-fold increase in its MIC value toward this honey was monitored when compared to the parent strain (P0) (Table 1). This observation is in line with a previous study that described how bacterial strains that had adapted to Manuka honey showed similar MIC values to the parent strains against Manuka hone.13 However, the biofilm growth of adapted P. aeruginosa to Manuka honey showed a higher MIC value for Manuka honey as well as an alteration in its sensitivity toward certain antibiotic drugs.29 Honey contains multiple compounds, that engage in antibacterial activities through various modes of actions, so low/limited resistance development is highly expected.24,30 Nevertheless, it would be worth conducting protein expression and whole genome analysis of bacteria that had been treated with Saudi honey to reveal its bacterial targets as well as any possible genomic and proteomic changes.
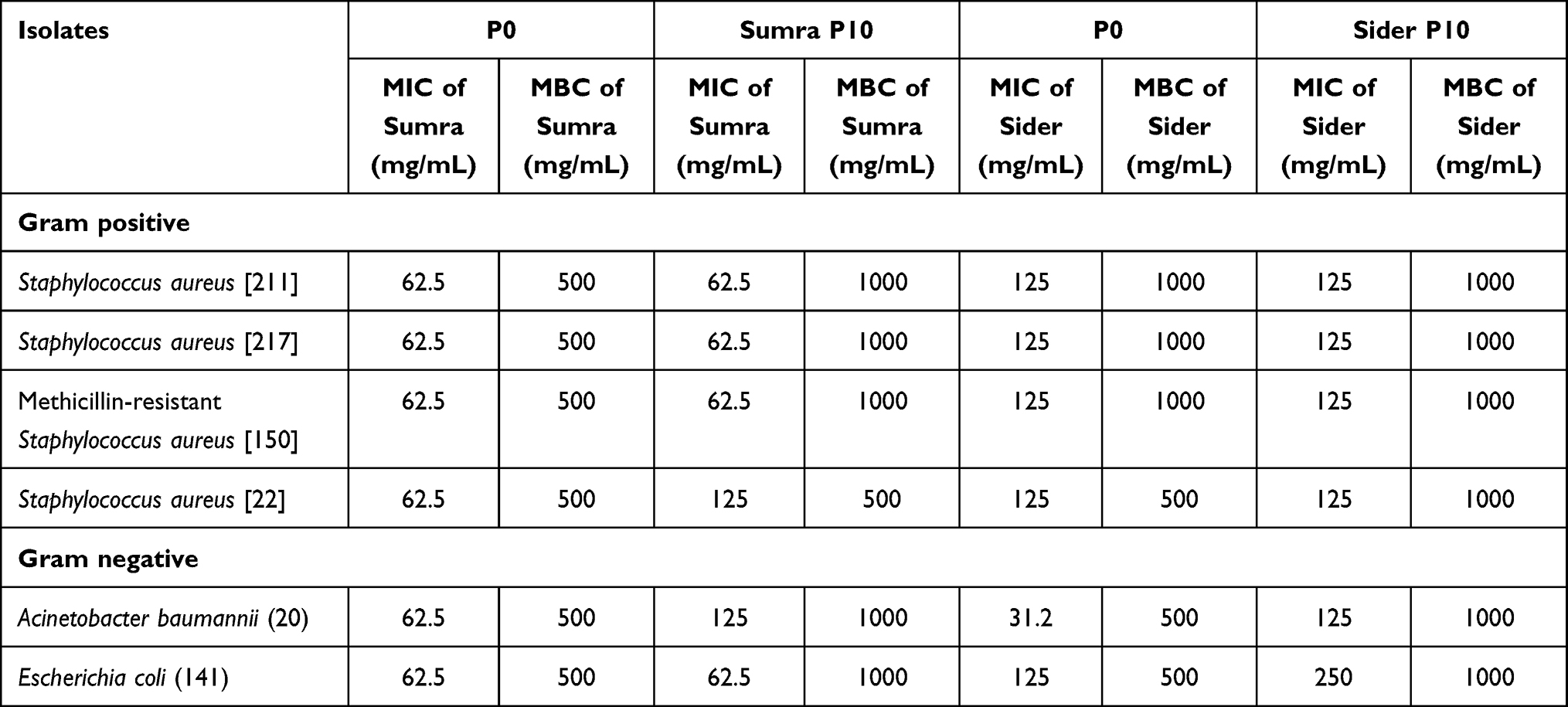 |
Table 1 Minimum Inhibitory Concentration (MIC) and Minimum Bactericidal Concentration (MBC) of Sumra and Sider Honeys of Untreated Bacteria [Passage 0 (P0)] and ten Times Treated Honey [Passage 10 (P10)] |
Biofilm formation is a well-known bacterial virulence factor that hampers wound healing and antimicrobial activity, leading to a severe form of infection, serious complications, treatment failure and death.31 Former studies have reported the promising antibiofilm activity of multiple types of tested Saudi honey (eg, Sumra honey) to treat infections caused by biofilm forming bacteria.15,17
The current findings show a 3-fold reduction in biofilm formation for Sumra-adapted MRSA as well as an approximately 1.5-fold reduction in biofilm formation for both the Sumra and Sider-adapted A. baumannii (20) strains (Figure 3). A reduction in biofilm formation of S. aureus has been reported following treatment with triclosan, which has been attributed to a reduced growth rate following exposure.19 This is in line with previous work that reported a reduction in biofilm formation among Gram-positive bacteria that had adapted to Manuka honey, including S. aureus.32 In contrast, a higher tendency toward biofilm formation was observed among all of the tested Gram-negative bacteria that had adapted to Manuka honey, although A. baumannii was not tested.13 A. baumannii tends to be more resistant to antibiotics and is one of the most common bacteria associated with hospital-acquired infections, especially wound infections. A wound is a complex environment, in which A. baumannii usually form biofilm, leading to a delayed healing process and/or treatment failure.33 The tested A. baumannii exhibited a reduction in biofilm formation and low MIC for both types of Saudi honey. Although more strains of A. baumannii need to be tested for the further validation and confirmation of these results, this finding is promising and supports the potential therapeutic use of the tested Saudi honey as an antimicrobial agent for treating wound infections caused by this bacterium.
Although the current study might has successfully assessed the changes in antibiotic sensitivity and biofilm formation of the tested bacteria following repeated in-vitro exposure to multiple types of Saudi honey, a few factors can be regarded as limitations of this study. The bacterial virulence could be evaluated using an in-vivo model (eg, larvae) to gain more knowledge about the pathogenesis of the bacteria following exposure to honey. In addition, an exploration of the growth rate and genomic analysis would facilitate the identification of any potential emergence of variance or mutation occurring at the genomic level.
Conclusion
In summary, the repeated in-vitro exposure of clinical bacteria to Saudi Sumra and Sider honey led to variable changes in both their antimicrobial sensitivity profiles and tendency toward biofilm formation, while very limited resistance to the used honey compared to the parent strains was observed. The increased antibiotic sensitivity of the adapted pathogenic bacteria (eg, S. aureus and A. baumannii) to Saudi honey (Sumra and Sider) and a reduced tendency toward biofilm formation are the main findings of this study. Although the exact mechanisms behind the resulting reduction in antibiotic sensitivity and biofilm formation remain undefined and so further investigation is required, this positive effect on the sensitivity profiles and biofilm formation of the tested bacteria that adapted to Saudi honey highlight the great potential therapeutic use of these tested types of Saudi honey as antimicrobial agents for treating bacterial infections, especially those associated with chronic wounds, since dressing these for a long period of time is mandatory.
Acknowledgments
All authors thank the Scientific Research Deanship at University of Ha’il- Saudi Arabia for their grant through a project number (MDR-22002).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Prestinaci F, Pezzotti P, Pantosti A. Antimicrobial resistance: a global multifaceted phenomenon. Pathog Glob Health. 2015;109(7):309–318. doi:10.1179/2047773215y.0000000030
2. Qanash H, Yahya R, Bakri MM, et al. Anticancer, antioxidant, antiviral and antimicrobial activities of Kei Apple (Dovyalis caffra) fruit. Sci Rep. 2022;12(1):5914. doi:10.1038/s41598-022-09993-1
3. Economou V, Gousia P. Agriculture and food animals as a source of antimicrobial-resistant bacteria. Infect Drug Resist. 2015;8:49–61. doi:10.2147/idr.S55778
4. Alhomoud F, Almahasnah R, Alhomoud FK. ”You could lose when you misuse” - factors affecting over-The-counter sale of antibiotics in community pharmacies in Saudi Arabia: a qualitative study. BMC Health Serv Res. 2018;18(1):915. doi:10.1186/s12913-018-3753-y
5. Al-Rajhi AMH, Qanash H, Almuhayawi MS, et al. Molecular interaction studies and phytochemical characterization of Mentha pulegium L. constituents with multiple biological utilities as antioxidant, antimicrobial, anticancer and anti-hemolytic agents. Molecules. 2022;27(15):4824. doi:10.3390/molecules27154824
6. Abdelghany YR, Bakri MM, Ganash M, Amin BH, Qanash H. Effect of Thevetia peruviana seeds extract for microbial pathogens and cancer control. Int J Pharmacol. 2021;17(8):643–655. doi:10.3923/ijp.2021.643.655
7. Curtis LT. Prevention of hospital-acquired infections: review of non-pharmacological interventions. J Hosp Infect. 2008;69(3):204–219. doi:10.1016/j.jhin.2008.03.018
8. CDC. Antibiotic Resistance Threats in the United States, 2013. US Department of Health and Human Services CDC; 2013.
9. Bazaid AS, Alamri A, Almashjary MN, et al. Antioxidant, anticancer, antibacterial, antibiofilm properties and gas chromatography and mass spectrometry analysis of manuka honey: a nature’s bioactive honey. Appl Sci. 2022;12(19):9928. doi:10.3390/app12199928
10. Rani GN, Budumuru R, Bandaru NR. Antimicrobial activity of honey with special reference to Methicillin Resistant Staphylococcus aureus (MRSA) and Methicillin Sensitive Staphylococcus aureus (MSSA). J Clin Diagn Res. 2017;11(8):Dc05–dc08. doi:10.7860/jcdr/2017/30085.10347
11. Singh S, Singh SK, Chowdhury I, Singh R. Understanding the mechanism of bacterial biofilms resistance to antimicrobial agents. Open Microbiol J. 2017;11:53–62. doi:10.2174/1874285801711010053
12. Lu J, Cokcetin NN, Burke CM, et al. Honey can inhibit and eliminate biofilms produced by Pseudomonas aeruginosa. Sci Rep. 2019;9(1):18160. doi:10.1038/s41598-019-54576-2
13. Mokhtar JA, McBain AJ, Ledder RG, Binsuwaidan R, Rimmer V, Humphreys GJ. Exposure to a manuka honey wound gel is associated with changes in bacterial virulence and antimicrobial susceptibility. Front Microbiol. 2020;11:2036. doi:10.3389/fmicb.2020.02036
14. Bazaid AS, Aldarhami A, Gattan H, Aljuhani B. Saudi honey: a promising therapeutic agent for treating wound infections. Cureus. 2021;13(10):e18882. doi:10.7759/cureus.18882
15. Al-Ghamdi AA, Ansari MJ. Biological and therapeutic roles of Saudi Arabian honey: a comparative review. Science. 2021;33(2):101329. doi:10.1016/j.jksus.2020.101329
16. Ghramh HA, Khan KA, Alshehri AMA. Antibacterial potential of some Saudi honeys from Asir region against selected pathogenic bacteria. Saudi J Biol Sci. 2019;26(6):1278–1284. doi:10.1016/j.sjbs.2018.05.011
17. Bazaid AS, Aldarhami A, Patel M, et al. The antimicrobial effects of Saudi sumra honey against drug resistant pathogens: phytochemical analysis, antibiofilm, anti-quorum sensing, and antioxidant activities. Pharmaceuticals. 2022;15(10):1212. doi:10.3390/ph15101212
18. Bazaid AS, Forbes S, Humphreys GJ, Ledder RG, O’Cualain R, McBain AJ. Fatty acid supplementation reverses the small colony variant phenotype in triclosan-adapted staphylococcus aureus: genetic, proteomic and phenotypic analyses. Sci Rep. 2018;8(1):3876. doi:10.1038/s41598-018-21925-6
19. Latimer J, Forbes S, McBain AJ. Attenuated virulence and biofilm formation in Staphylococcus aureus following sublethal exposure to triclosan. Antimicrob Agents Chemother. 2012;56(6):3092–3100. doi:10.1128/aac.05904-11
20. Eteraf-Oskouei T, Najafi M. Traditional and modern uses of natural honey in human diseases: a review. Iran J Basic Med Sci. 2013;16(6):731–742.
21. Samarghandian S, Farkhondeh T, Samini F. Honey and health: a review of recent clinical research. Pharmacognosy Res. 2017;9(2):121–127. doi:10.4103/0974-8490.204647
22. Kwakman PHS, te Velde AA, de Boer L, Vandenbroucke-Grauls CMJE, Zaat SAJ. Two major medicinal honeys have different mechanisms of bactericidal activity. PLoS One. 2011;6(3):e17709. doi:10.1371/journal.pone.0017709
23. Dixon B. Bacteria can’t resist honey. Lancet Infect Dis. 2003;3(2):116. doi:10.1016/s1473-3099(03)00524-3
24. Bischofberger AM, Pfrunder Cardozo KR, Baumgartner M, Hall AR. Evolution of honey resistance in experimental populations of bacteria depends on the type of honey and has no major side effects for antibiotic susceptibility. Evol Appl. 2021;14(5):1314–1327. doi:10.1111/eva.13200
25. Brudzynski K, Sjaarda C. Antibacterial compounds of Canadian honeys target bacterial cell wall inducing phenotype changes, growth inhibition and cell lysis that resemble action of β-lactam antibiotics. PLoS One. 2014;9(9):e106967. doi:10.1371/journal.pone.0106967
26. Combarros-Fuertes P, Estevinho LM, Teixeira-Santos R, et al. Evaluation of physiological effects induced by manuka honey upon staphylococcus aureus and Escherichia coli. Microorganisms. 2019;7(8):258. doi:10.3390/microorganisms7080258
27. Marro FC, Abad L, Blocker AJ, Laurent F, Josse J, Valour F. In vitro antibiotic activity against intraosteoblastic Staphylococcus aureus: a narrative review of the literature. J Antimicrob Chemother. 2021;76(12):3091–3102. doi:10.1093/jac/dkab301
28. Schulz Zur Wiesch P, Engelstädter J, Bonhoeffer S. Compensation of fitness costs and reversibility of antibiotic resistance mutations. Antimicrob Agents Chemother. 2010;54(5):2085–2095. doi:10.1128/aac.01460-09
29. Camplin AL, Maddocks SE. Manuka honey treatment of biofilms of Pseudomonas aeruginosa results in the emergence of isolates with increased honey resistance. Ann Clin Microbiol Antimicrob. 2014;13:19. doi:10.1186/1476-0711-13-19
30. Combarros-Fuertes P, Fresno JM, Estevinho MM, Sousa-Pimenta M, Tornadijo ME, Estevinho LM. Honey: another alternative in the fight against antibiotic-resistant bacteria? Antibiotics. 2020;9(11):774. doi:10.3390/antibiotics9110774
31. Gilbert P, Allison DG, McBain AJ. Biofilms in vitro and in vivo: do singular mechanisms imply cross-resistance? J Appl Microbiol. 2002;92:98s–110s. doi:10.1046/j.1365-2672.92.5s1.5.x
32. Kot B, Sytykiewicz H, Sprawka I, Witeska M. Effect of manuka honey on biofilm-associated genes expression during methicillin-resistant Staphylococcus aureus biofilm formation. Sci Rep. 2020;10(1):13552. doi:10.1038/s41598-020-70666-y
33. Eze EC, Chenia HY, El Zowalaty ME. Acinetobacter baumannii biofilms: effects of physicochemical factors, virulence, antibiotic resistance determinants, gene regulation, and future antimicrobial treatments. Infect Drug Resist. 2018;11:2277–2299. doi:10.2147/idr.S169894
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.