")
Back to Journals » Neuropsychiatric Disease and Treatment » Volume 19
Effects of Electroacupuncture on Gut Microbiota and Fecal Metabolites in Rats with Poststroke Depression
Authors Cai W, Wei XF, Hu C, Ji J, Cui HS, Shen WD
Received 19 April 2023
Accepted for publication 21 June 2023
Published 12 July 2023 Volume 2023:19 Pages 1581—1592
DOI https://doi.org/10.2147/NDT.S415098
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Roger Pinder
Wa Cai,1,* Xi-Fang Wei,1,* Chen Hu,1 Jun Ji,2 Hua Shun Cui,1 Wei-Dong Shen1
1Department of Acupuncture, Shanghai Shuguang Hospital Affiliated to Shanghai University of Traditional Chinese Medicine, Shanghai, People’s Republic of China; 2Department of Acupuncture Literature, Shanghai Research Institute of Acupuncture and Meridian, Shanghai, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Wei-Dong Shen; Hua Shun Cui, Email [email protected]; [email protected]
Background: Poststroke depression (PSD) is the most frequent neuropsychiatric consequence of stroke. Electroacupuncture (EA) has been found to be an effective therapy for treating PSD. However, the underlying mechanisms of EA’s efficacy remain unclear. This research aimed to investigate the effects of EA on alterations in gut microbiota and fecal metabolome in PSD rats.
Methods: Analyses of gut microbiome and fecal metabolome were performed to identify gut microbes and their functional metabolites in a sham group, PSD group, and EA group. We conducted enrichment analysis to identify the differential metabolic pathways in three groups. Correlations between altered gut microbes and differential metabolites after EA treatment were studied.
Results: PSD showed decreased species-richness/diversity indices of microbial composition, characterized by an increase in Muribaculaceae, Peptostreptococcaceae, Oscillospiraceae, Ruminococcaceae, and Clostridiaceae and a decrease in Lactobacillaceae, Lachnospiraceae, and Bacteroidaceae. Of these, the abundance of Muribaculaceae, Lactobacillaceae, Lachnospiraceae, Peptostreptococcaceae, and Clostridiaceae were reversed by EA. Furthermore, PSD was associated with 34 differential fecal metabolites, mainly belonging to steroid hormone biosynthesis, that could be regulated by EA.
Conclusion: Regulation of gut microbiome and lipid metabolism could be one of the potential mechanisms for EA treatment for alleviating the depressive behaviors of PSD.
Keywords: electroacupuncture, gut microbiome, fecal metabolome, metabolic pathways, poststroke depression
Introduction
Poststroke depression (PSD) usually predicts poor prognosis, such as cognitive impairment, poor rehabilitation outcomes, and increased mortality.1–4 Nearly 30% of stroke survivors have depression within 5 years of the stroke.2 PSD has been reported to be associated with increased inflammation,5,6 changes in neurotransmitters,7 HPA-axis dysfunction,8,9 and decreased neurotrophic factors.10–12
The gut microbiome consists of bacteria, viruses, microorganisms, fungi, and other life forms. Dysfunctions of the gut microbiome have been implicated in neuropsychiatric disorders, such as depressive disorder, schizophrenia-spectrum disorders, and bipolar disorder.13–16 Previous research revealed that PSD rats had decreased species-richness indices.17 A quantifiable monitor to inform the molecular mechanisms of diseases, metabolomics has been used as a diagnostic, prognostic tool of psychiatric disorders in a number of studies.18–21 Four published papers22–25 identified differential urinary metabolites, including lactic acid, palmitic acid, azelaic acid, and tyrosine, in PSD patients. All of these studies provide a scientific basis for the exploration of therapeutical strategies for PSD.
Based on classical meridian theory, the acupuncture treatment of traditional Chinese medicine is a therapy puncturing needles into acupoints.26,27 Electroacupuncture (EA) treatment was found to be effective for alleviating PSD with fewer side effects in the latest meta-analysis.28 Only one study by Peng et al29 revealed that acupuncture can relieve depression-like behaviors of chronic unpredictable mild stress (CUMS) rats by regulating intestinal flora. However, whether EA can effectively regulate the gut microbiota and its metabolites of PSD remains to be verified and clarified.
This study was designed and conducted to investigate the effects of EA on the gut microbiota and fecal metabolites in rats with PSD to explore the microbiological mechanisms of EA treatment for PSD. We selected acupoints of DU20 and DU24, both of which belong to the DU meridian. The combined use of DU20 and DU24 has been found to effectively relieve depressive behaviors of rats in previous studies.30,31
Methods
Animals
Male 2-month-old 250 g Sprague Dawley rats were purchased from Huan Shuo Laboratory Animal Technology (Shanghai, China). Approved by the Ethics Committee of Shanghai University of Traditional Chinese Medicine (PZOHUTCM210305007), the protocol of animal procedures complied with the ARRIVE guidelines, and procedures were carried out in accordance with the UK Animals (Scientific Procedures) Act 2010 and EU Directive 2010/63/EU for animal experiments. Figure 1 shows the workflow. The temperature of the animal room was controlled at 24°±2°C under a 12-hour light–dark cycle. A total of 39 rats with similar baseline performance were included after the first sucrose preference test (SPT) following 1-week acclimatization. Of them, eight rats were categorized to the sham group, and the rest had middle cerebral artery occlusion surgery performed. In total, 15 rats died within 48 hours after surgery. The survivors were randomly divided into a PSD group (n=8) and EA group (n=8).
 |
Figure 1 Time schedule of the experiment. Abbreviations: CUMS, chronic unpredictable mild stress; W0, beginning of CUMS regimen; W1–W5, CUMS performed daily for 5 weeks; W3–W5, acupuncture treatment conducted daily for 2 weeks; SPT, sucrose preference test; LAT, locomotor activity test; EA, electroacupuncture; FSC, fecal sample collection. |
Establishment of PSD Model
Transient middle cerebral artery occlusion surgery32 was conducted. After anesthetization, a midline neck incision was made at to expose the right common carotid artery (CCA), external carotid artery (ECA), and internal carotid artery (ICA). We placed two closely spaced permanent knots at the distal part of the ECA to prevent the backflow of blood and cut the ECA between the knots. The tied section of the ECA was incised to insert the monoflament after the microvascular clip had been placed in the ICA temporarily proximal to the CCA junction. The filament was advanced up to 17–20 mm into the MCA from the CCA junction. The suture was removed to establish reperfusion after 1.5 hours of ischemia. Rats undergoing the same surgical procedure without suture insertion belonged to the sham group.
Three days after surgery, rats in the PSD group underwent a CUMS regimen33 for 5 weeks in a single cage. Any two of the following 10 stressors were performed daily: water deprivation for 24 hours, food deprivation for 24 hours, overnight illumination, soiled cage for 24 hours, cage tilt (45°) for 24 hours, restraint for 4 hours, restraint at 4°C for 2 hours, noise for 8 hours, stroboscope lighting for 12 hours, cage shaking for 15 minutes.
Electroacupuncture Treatment
We selected acupoints of DU20 and DU24 for PSD treatment (Figure 2). The selected acupoints were inserted with stainless steel acupuncture needles (Energy, 40 mm in length and 0.25 mm in diameter; Jiajian Medical Supplies, Jiangsu, China) to a depth of about 2 mm with an electric current at a frequency of 2 Hz using an EA device (Hwato SDZ-II; Suzhou Medical Supplies, Jiangsu, China) for about 20 minutes per session. One session was conducted per day. During EA, rats were slightly anesthetized with isoflurane. The selection of acupoints and electric current frequency of EA were determined by our previous studies,30,31 which were verified to have better efficacy for alleviating depressive behaviors of PSD rats. EA treatment started from the end of the third week of CUMS and lasted daily for 14 days. Each EA treatment session was applied to the rats before the CUMS session on the same day. Only rats in the EA group were treated with EA, and rats in the sham and PSD groups did not accept EA intervention.
 |
Figure 2 Acupoint locations of DU20 and DU24 on the rat body. |
Behavioral Tests
The behavioral tests consisted of sucrose preference test and locomotor activity test, both of which were performed on the last day of CUMS and acupuncture treatment.
Sucrose Preference Test
The sucrose preference test (SPT) was used to assess anhedonia in the animals. Before water deprivation for 24 hours, animals were provided with two bottles of 1% sucrose solution for 24 hours followed by one bottle of 1% sucrose solution and another bottle of purified water for 24 hours with bottle positions switched after 12 hours. After 24 hours’ water deprivation, one bottle of 1% sucrose solution and another bottle of purified water were provided to each rat for 1 hour. Sucrose intake (g)/(sucrose intake [g] + purified water intake (g]) was used to calculate the sucrose preference rate.
Locomotor Activity Test
A dark box (100×100×50 cm) and an automatic data-acquisition system (Xinruan Information and Technology, Shanghai, China) were used in the locomotor activity test. There were 25 equal square quadrants at the floor of the box. The system automatically measured locomotor activity when at least three paws were presented in a quadrant. Each rat was monitored individually for 5 minutes. We cleaned the box thoroughly before the next rat was tested.
Sample Collection
At around 8:00 am one day after behavioral tests, fresh feces of rats were collected using the sterilization centrifuge tubes. At least 50 mg fecal samples were collected from each group for the analysis of fecal microbiome and metabolites. Then, the fecal samples were immediately stored in liquid nitrogen before transferal to a −80°C refrigerator.
16S rRNA Gene-Sequence Analysis
An Omega soil DNA kit (Omega Bio-Tek, Norcross, GA, United States) was used to separate and extract microbial DNA from fecal samples. The bacterial 16S rRNA gene was amplified by the primers 338F (5′-ACTCCTACGGGAGGCAGCAG-3′) and 806R (5′-GGACTACHVGGGTWTCTAAT-3′). The PCR program was completed by a thermal cycling PCR system (GeneAmp 9700; ABI, USA). Trimmomatic filtered the quality of the original fastq files. FLASH was used to merge the data. The similarity of operational taxonomic units (OTUs; version 7.1, http://drive5.com/uparse) based on a 97% criterion was classified by UPARSE. UCHIME was used to identify and remove the chimeric sequences. We used the Silva (SSU123) 16S rRNA database to execute the RDP classifier algorithm (http://rdp.cme.msu.edu) program to analyze the classification of each 16S RNA gene sequence. The α-diversity was assessed by species-richness indices (Ace, Chao, and Sobs) and species-diversity indices (Shannon, Simpson). Principal-coordinate analysis was used to evaluate the whole difference and similarity of bacterial communities among three groups. The Wilcoxon rank-sum test was used to identify microbiota discrimination between any two groups.
Fecal Metabolome Analysis
LC-MS analysis was performed using the UHPLC-Q Exactive HF-X system of Thermo Fisher Scientific. Fecal samples were weighed. The metabolites were extracted using a 400 µL methanol:water (4:1 v:v) solution with 0.02 mg/mL L-2-chlorophenylalanine. The mixture was allowed to settle at −10°C and treated with a Wonbio-96c high-throughput tissue crusher (Shanghai Wanbo Biotechnology) at 50 Hz for 6 min, followed by ultrasound at 40 kHz for 30 min at 5°C. The samples were placed at −20°C for 30 min to precipitate proteins. The supernatant was carefully transferred to sample vials for LC-MS analysis after centrifugation at 13,000 g at 4°C for 15 min. All samples were stored at 4°C during the period of analysis. A Thermo UHPLC-Q Exactive HF-X mass spectrometer was used to collect the mass spectrometric data, which was performed in Data Dependent Acquisition mode. The detection was carried out over a mass range of 70–1050 m/z. We used orthogonal partial least-squares discrimination analysis to visually discriminate between groups. The differential metabolites discriminating between groups were identified by orthogonal projections to latent structures discriminant analysis (variable importance plot [VIP] >2, P<0.05). The Kyoto Encyclopedia of Genes and Genomes Pathway database was used to carry out pathway analyses.
Statistical Analysis
SPSS 21.0 was used to carry out the statistical analyses. One-way ANOVA was used to analyze continuous variables, such as sucrose preference and locomotor activity counts, which were presented as means ± SEM. Wilcoxon’s rank-sum test was applied to analyze α-diversity. Adonis analysis was used to analyze β-diversity. The correlation between changes in gut microbes and differential fecal metabolites was determined by Spearman correlation analysis. P<0.05 was considered statistically significant.
Results
EA Treatment Alleviates Depressive Behaviors of PSD Rats
As shown in Figure 3, rats in the PSD group showed decreased sucrose preference and reduced locomotion counts at the end of the CUMS regimen compared with the sham group (n=8 per group, P<0.01). EA treatment was able to substantially increase sucrose preference and locomotion counts of PSD rats (n=8 per group, P<0.01), which indicated that EA treatment could effectively alleviate PSD.
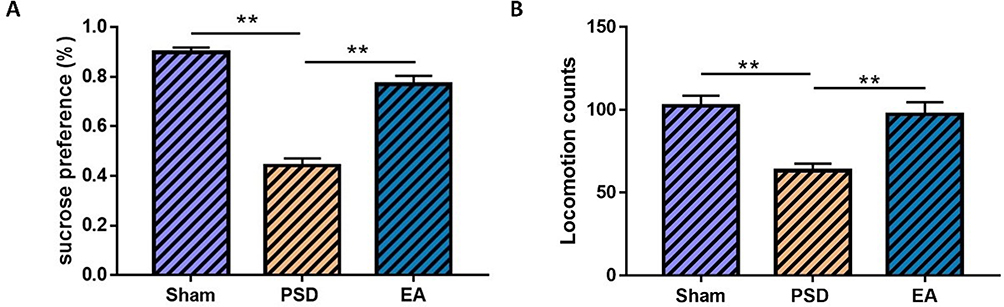 |
Figure 3 The therapeutic effect of EA treatment on depressive behaviors of PSD rats. (A) Sucrose preference; (B) locomotion counts (n=8 per group, one-way ANOVA for (A and B). **P<0.01. Abbreviations: PSD, poststroke depression; EA, electroacupuncture. |
Effects of EA on Species Diversity and Richness in PSD Rats
In sum, 1,477,882 high-quality reads across all samples were obtained, with an average length of 413. All these reads were clustered into 983 OTUs. The α-diversity values consisting of Shannon, Simpson, Ace, Sobs, and Chao were compared among groups. Ace, Chao, and Sobs were used to evaluate species richness, while species diversity was characterized using Shannon and Simpson. Both microbial richness indices and diversity indices were different among groups, with no significant distinction. The PSD group had lower Chao, Ace, Sobs, and Shannon indices than the sham group, indicating that PSD was characterized by decreased species richness and diversity (Figure 4A-C, P>0.05). Compared with the PSD group, EA treatment improved the species richness and diversity of PSD rats by increasing the Chao, Ace, Sobs, and Shannon indices (Figure 4D and E, P>0.05).
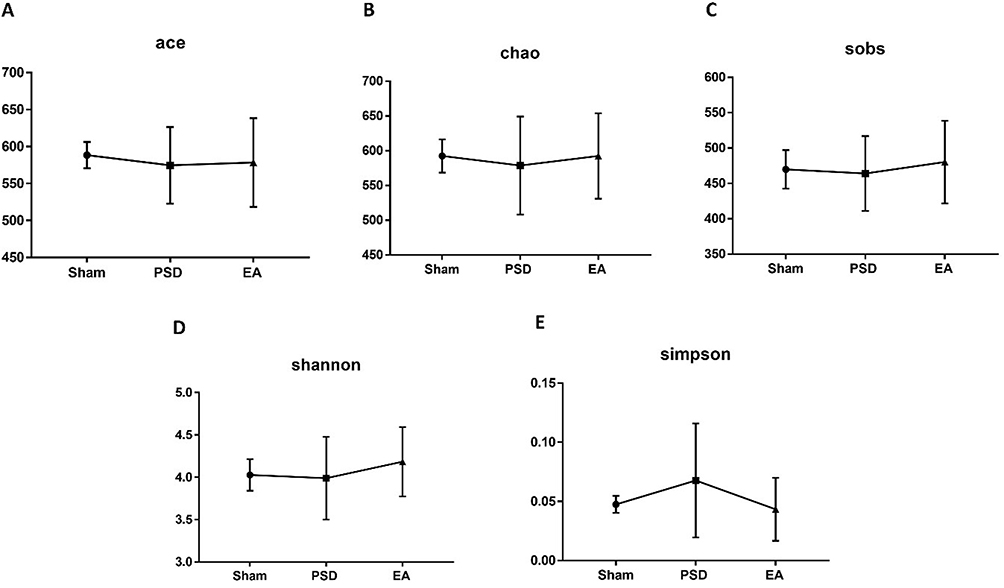 |
Figure 4 Effects of EA on species diversity and richness in PSD rats. (A–C) Species richness indices (Ace, Chao and Sobs); (D and E) species diversity indices (Shannon and Simpson). Eight per group, Wilcoxon rank-sum test for (A-E). Abbreviations: EA, electroacupuncture; PSD, poststroke depression. |
Effects of EA on Microbial Composition in PSD Rats
Analysis of β-diversity was performed to determine the effects of EA on microbial composition in PSD rats. It was revealed on principal-coordinate analysis that gut microbial composition was significantly different among the three groups (Figure 5A; n=8 per group). The differential OTUs between any two groups were analyzed by the Wilcoxon rank-sum test (Figure 5B and C). Compared with the sham group, PSD was characterized by 108 OTUs, which mainly belonged to Tannerellaceae), Ruminococcaceae (eight OTUs), Rikenellaceae (three OTUs), Prevotellaceae (six OTUs), Peptococcaceae (two OTUs), Oscillospiraceae (six OTUs), Muribaculaceae (10 OTUs), Monoglobaceae (three OTUs), Lachnospiraceae (21 OTUs), Helicobacteraceae), Erysipelotrichaceae), Erysipelatoclostridiaceae (three OTUs), Elusimicrobiaceae), Eggerthellaceae (15 OTUs), Deferribacteraceae), Christensenellaceae (four OTUs), Bacteroidaceae), Anaerovoracaceae (four OTUs), Anaerofustaceae), and Actinomycetaceae (one OTU) at the family level (Table S1). Compared with the PSD group, EA treatment was characterized by 52 OTUs, mainly belonging to Ruminococcaceae (four OTUs), Rikenellaceae (three OTUs), Prevotellaceae (three OTUs), Peptococcaceae (one OTU), Oscillospiraceae (one OTU), Muribaculaceae (two OTUs), Monoglobaceae (one OTU), Marinifilaceae (two OTUs), Lachnospiraceae (eight OTUs), Eggerthellaceae (12 OTUs), Desulfovibrionaceae (two OTUs), Christensenellaceae (two OTUs), Butyricicoccaceae (one OTU), Bacteroidaceae(two OTUs), and Anaerovoracaceae (one OTU) (Table S2). Figure 5D presents the microbial compositions at the family level. Muribaculaceae, Lactobacillaceae, Lachnospiraceae, Peptostreptococcaceae, Oscillospiraceae, Ruminococcaceae, Clostridiaceae, Bacteroidaceae, and Prevotellaceae were relatively abundant in all groups. We found an increase in Muribaculaceae, Peptostreptococcaceae, Oscillospiraceae, Ruminococcaceae, and Clostridiaceae and a decrease in Lactobacillaceae, Lachnospiraceae, and Bacteroidaceae in the PSD group compared to the sham group. Of these, the abundance of Muribaculaceae, Lactobacillaceae, Lachnospiraceae, Peptostreptococcaceae, Clostridiaceae was reversed by EA.
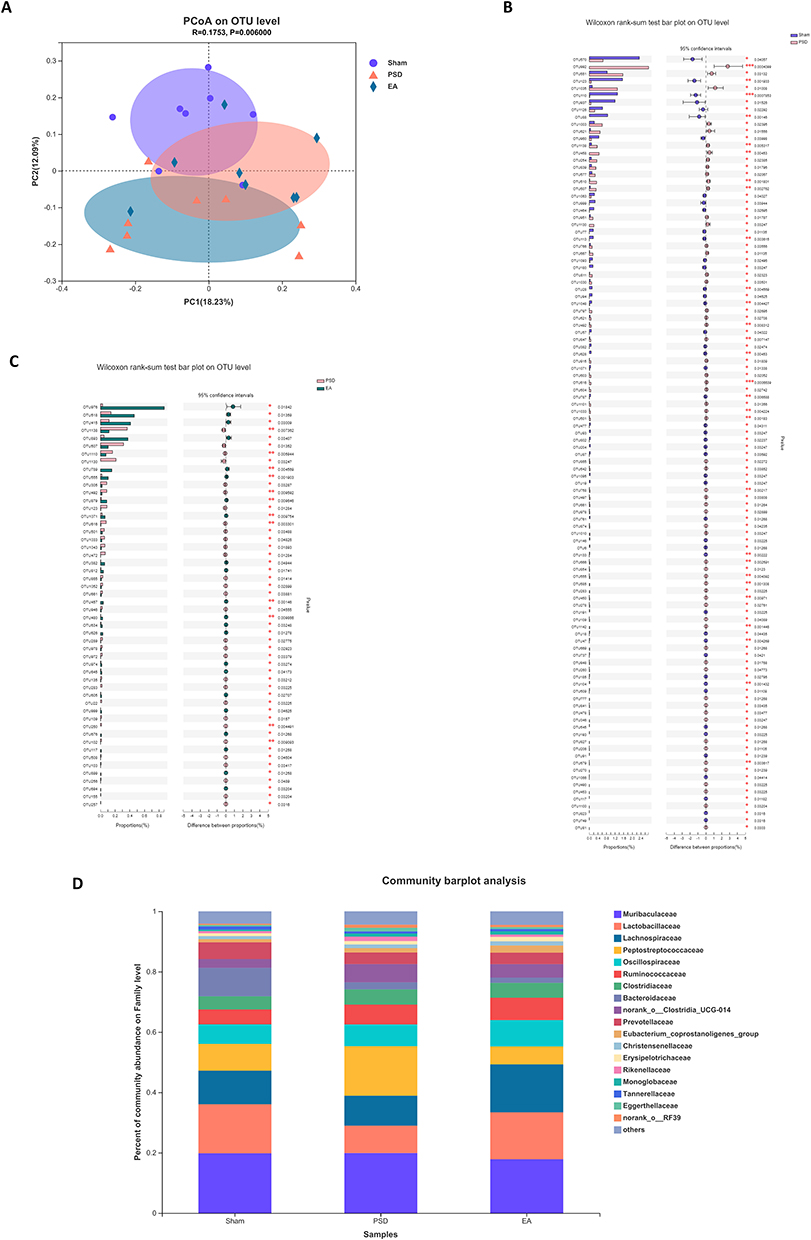 |
Figure 5 Effects of EA on microbial composition in PSD rats. (A) PCoA analysis at the OUT level (n=8 per group, Adonis analysis). (B) Microbiota discrimination (PSD vs sham, n=8 per group, Wilcoxon rank-sum test). (C) Microbiota discrimination (EA vs PSD, n=8 per group, Wilcoxon rank-sum test). (D) Taxonomic composition of gut microbiota at the family level; (n=8 per group, Bonferroni post hoc analysis). *P<0.05; **P<0.01. Abbreviations: EA, electroacupuncture; OTU, operational taxonomic unit; PCoA, principal coordinates analysis; PSD, poststroke depression. |
Effects of EA on Fecal Metabolites in PSD Rats
As shown in Figure 6A, the fecal metabolic phenotypes were significantly different among the three groups (n=8 per group). Compared with sham, 34 differential fecal metabolites were found in PSD rats, with 13 metabolites increased and 21 metabolites decreased (Table S3, VIP >2, P<0.05). Pathway analysis revealed that ovarian steroidogenesis, steroid-hormone biosynthesis, biosynthesis of plant hormones, and chemical carcinogenesis were the most significantly enriched pathways (Figure 6B, P<0.05; Table S5, P<0.05). There were 40 differential fecal metabolites in the EA group compared with the PSD group, of which 22 metabolites were increased and 18 metabolites were decreased (Table S4, VIP >2, P<0.05). The most significantly enriched pathway was steroid-hormone biosynthesis (Figure 6C, P<0.05; Table S6, P<0.05).
 |
Figure 6 Effects of EA on fecal metabolites in PSD rats. (A) PLS-DA analysis (n=8 per group, principal-component analysis). (B) KEGG enrichment pathways (PSD vs sham, n=8 per group, Bonferroni post hoc analysis). (C) KEGG enrichment pathways (EA vs PSD, n=8 per group, Bonferroni post hoc analysis). Abbreviations: EA, electroacupuncture; PLS-DA, partial least squares discriminant analysis; PSD, poststroke depression. |
Correlations Between Gut Microbiota and Fecal Metabolites after EA Treatment
As depicted in Figure 7, correlation analysis was performed to explore the associations between altered gut microbiota with fecal metabolites after EA treatment. Overall, 98.11% (52 of 53 OTUs) of altered microbial OTUs were significantly associated with the 40 differential metabolites (r>0.6, P<0.05), mainly belonging to steroid-hormone biosynthesis (three metabolites), nicotine addiction (two metabolites), synaptic vesicle cycle (two metabolites), C5-branched dibasic acid metabolism (two metabolites), arginine biosynthesis (two metabolites), alanine (two metabolites), aspartate and glutamate metabolism (two metabolites), taste transduction (two metabolites), carbapenem biosynthesis (two metabolites), pathways of neurodegeneration (two metabolites), neuroactive ligand–receptor interaction (two metabolites), biosynthesis of various secondary metabolites (two metabolites), glycerophospholipid metabolism (two metabolites), arginine and proline metabolism (two metabolites), and ABC transporters (two metabolites). EA treatment was characterized by significant changes in gut microbiota and fecal metabolites. The alteration of gut microbiota was closely correlated with the metabolism of PSD rats.
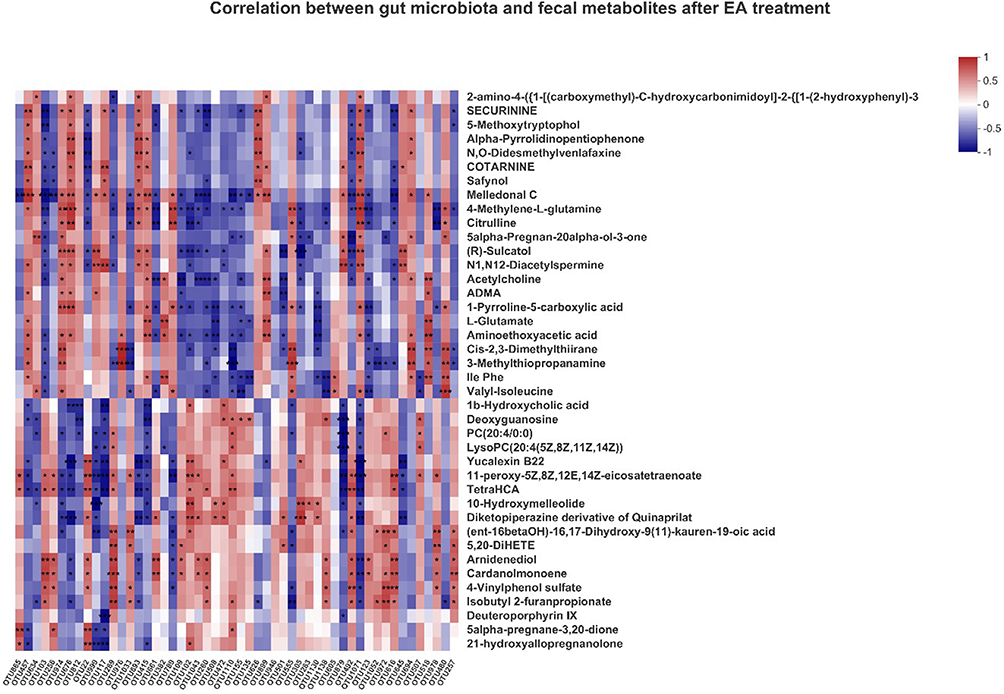 |
Figure 7 Heat map of correlation coefficients of gut microbiota and fecal metabolites (n=8 per group, Spearman correlation analysis). *P<0.05; **P<0.01; ***P<0.001. Abbreviations: EA, electroacupuncture; OTU, operational taxonomic unit; PCoA, principal coordinates analysis. |
Discussion
A highly diverse community of microorganisms that live in the digestive tracts,34,35 gut microbiota are much more abundant than somatic and germ-line cells.36 Gut microbiota can control the central nervous system via various mechanisms, such as changes to neurotrophic factors and neurotransmitters, which regulate metabolite production. Through the microbiota, the brain and gut can influence each other’s functions via neuroendocrine, sensorineural, molecular, and neuroimmune pathways. Brain–gut–microbiota cross talk is closely associated with suchneuropsychiatric disorders as depression, anxiety, bipolar disorder, and schizophrenia.37,38 Gut microbiota can impact brain function and behavior through the brain–gut–microbiota axis.39,40 Jiang et al17 revealed that alterations in both gut microbiome and fecal metabolites were associated with depression after stroke.
Our study compared the gut microbiome and fecal metabonomics among three groups (sham, PSD and EA). The gut microbial phenotypes of PSD rats were significantly different from sham rats. Furthermore, depressive behaviors of PSD rats were greatly alleviated by EA treatment, which significantly changed the gut microbial phenotypes of PSD rats. Altered gut microbial OTUs after EA treatment were significantly associated with all the metabolites. EA alleviated depression-like behaviors by regulating Lactobacillaceae and Bacteroidaceae,29,41 which is consistent with two previous studies. Crucial metabolic pathways were revealed in further analysis. The results of our study indicate that EA can regulate the gut microbiome, which is associated with changes in the steroid-hormone biosynthesis pathway. Our findings are consistent with Jiang et al’s17 in that the gut microbiome is associated with the development of PSD, the mechanism of which is related to the regulation of lipid metabolism due to PSD rats being particularly enriched with steroid biosynthesis.
Of note, the microbial composition of PSD rats revealed decreased species-richness and -diversity indices compared with the sham group. Gut dysbiosis is characterized as an abnormal alteration in gut microbiome diversity with a decrease in beneficial microbes, which affects neural and immune homeostasis, resulting in depressive behaviors.42,43 In our study, compared with sham, PSD were characterized by Tannerellaceae, Ruminococcaceae, Rikenellaceae, Prevotellaceae, Peptococcaceae, Oscillospiraceae, Muribaculaceae, Monoglobaceae, Lachnospiraceae, Helicobacteraceae, Erysipelotrichaceae, Erysipelatoclostridiaceae, Elusimicrobiaceae, Eggerthellaceae, Deferribacteraceae, Christensenellaceae, Bacteroidaceae, Anaerovoracaceae, Anaerofustaceae and Actinomycetaceae, six of which were consistent with the results of a previous study17 where PSD rats showed altered gut microbiome characterized by Lactobacillaceae, Lachnospiraceae, Erysipelotrichaceae, Streptococcaceae, Veillonellaceae, Ruminococcaceae, Muribaculaceae, Enterococcaceae, Burkholderiaceae, Mycoplasmataceae, Enterobacteriaceae and Eggerthellaceae compared with both sham and stroke group by LEfSe analysis. Compared with sham rats, we observed an increase in Muribaculaceae, Peptostreptococcaceae, Oscillospiraceae, Ruminococcaceae, Clostridiaceae and a decrease in Lactobacillaceae, Lachnospiraceae, Bacteroidaceae of PSD rats. Furthermore, EA treatment reversed the abundances of Muribaculaceae, Lactobacillaceae, Lachnospiraceae, Peptostreptococcaceae and Clostridiaceae, suggesting that EA can improve depressive symptoms of PSD by reducing the abundance of pathogenic microbes and increasing the abundance of beneficial microbes.
In addition, disturbances of fecal metabolomics in PSD were found in our study. Most of the discriminating fecal metabolites in PSD rats belonged to pathways of ovarian steroidogenesis, steroid-hormone biosynthesis, chemical carcinogenesis, and biosynthesis of plant hormones. Three of these four enriched pathways pertain to lipid metabolism, consistent with a previous study that the lipid-related metabolic pathway (steroid biosynthesis) was the most enriched among 19 pathways revealed in PSD. Cristina et al43 reported that metabolic adaptations were motivated, thus increasing caloric efficiency under a series of stress. Ha et al’s study44 found that the livers of animals enhanced energy storage under chronic stress, responding to the great demand of lipid metabolism. Associated with neurological diseases,45 cholesterol can be converted into steroid-related hormones, including estrogens, androgens, and glucocorticoids. Virtanen et al46 found that low high–density lipoprotein cholesterol predicted lower risk of long-term depressive symptoms. Another study47 revealed that corticosteroids were associated with an increased risk of depression. As a result, PSD may be closely related to abnormal lipid metabolism. We found that EA treatment can regulate steroid-hormone biosynthesis. A previous study48 showed similar results: acupuncture treatment effectively alleviated depression by altering hepatic lipid metabolism through attenuating the insensitivity of leptin.
A couple of limitations in this research should be noted. Despite the finding that PSD is associated with gut dysbiosis and EA can improve gut microbiota imbalance, fecal transplantation experiments could be considered to provide more evidence for the causal relation. Moreover, we found the most significantly identified metabolic pathway of PSD, which can be regulated by EA treatment, but the key regulatory targets should be explored in future research.
In conclusion, this study revealed that the gut microbiome and fecal metabolome might play an important role in the pathogenesis of PSD, which could be associated with the brain–gut axis. Differential gut microbiomes may be related to the regulation of lipid metabolism. The regulation of the gut microbiome and lipid metabolism could be one of the potential mechanisms for EA treatment alleviating the depressive behaviors of PSD. Our findings give a fresh perspective on the mechanisms in EA treatment for PSD.
Data Sharing
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding authors.
Ethics Approval
The protocol of animal procedures was approved by the Ethics Committee of Shanghai University of Traditional Chinese Medicine (PZOHUTCM210305007).
Acknowledgments
Wa Cai and Xi-Fang Wei are joint first authors of this study. Hua Shun Cui and Wei-Dong Shen are joint correspondence authors for this study.
Author Contributions
All authors made a significant contribution to the work reported, whether in conception, study design, execution, acquisition of data, analysis, interpretation, or all these areas, took part in drafting, revising, or critically reviewing the article, gave final approval to the version to be published, have agreed on the journal to which the article has been submitted, and agree to be accountable for all aspects of the work.
Funding
The National Natural Science Foundation of China (82004444), National Natural Science Foundation of China (82074535), Shanghai Municipal Health Commission (20204Y0472), Youth Medical Talents Specialist Program of Shanghai “Rising Stars of Medical Talents” Youth Development Program, Shanghai University of Traditional Chinese medicine (2022YG-55/2022YG-56), Shanghai Municipal Health Commission (2021LPTD-004), Shanghai Municipal Health Commission (ZY [2021–2023]-0209-10), Clinical Technology Innovation Project of Municipal Hospital (SHDC22021210), and Shanghai Science and Technology Commission (20Y21902900).
Disclosure
The authors declare that there are no conflicts of interest in this work.
References
1. Hadidi N, Treat-Jacobson DJ, Lindquist R. Poststroke depression and functional outcome: a critical review of literature. Heart Lung. 2009;38(2):151–162. doi:10.1016/j.hrtlng.2008.05.002
2. Hackett ML, Pickles K. Part I: frequency of depression after stroke: an updated systematic review and meta-analysis of observational studies. Int J Stroke. 2014;9(8):1017–1025. doi:10.1111/ijs.12357
3. Bartoli F, Lillia N, Lax A, et al. Depression after stroke and risk of mortality: a systematic review and meta-analysis. Stroke Res Treat. 2013;2013:862978. doi:10.1155/2013/862978
4. Cai W, Mueller C, Li YJ, Shen WD, Stewart R. Post stroke depression and risk of stroke recurrence and mortality: a systematic review and meta-analysis. Ageing Res Rev. 2019;50:102–109.
5. Danese A, Pariante CM, Caspi A, Taylor A, Poulton R. Childhood maltreatment predicts adult inflammation in a life-course study. Proc Natl Acad Sci U S A. 2007;104(4):1319–1324.
6. Setiawan E, Wilson AA, Mizrahi R, et al. Role of translocator protein density, a marker of neuroinflammation, in the brain during major depressive episodes. JAMA Psychiatry. 2015;72(3):268–275.
7. Villa RF, Ferrari F, Moretti A. Post-stroke depression: mechanisms and pharmacological treatment. Pharmacol Ther. 2018;184:131–144.
8. Barca ML, Eldholm RS, Persson K, et al. Cortisol levels among older people with and without depression and dementia. Int Psychogeriatr. 2019;31(4):597–601.
9. Doolin K, Farrell C, Tozzi L, Harkin A, Frodl T, O’Keane V. Diurnal hypothalamic-pituitary-adrenal axis measures and inflammatory marker correlates in major depressive disorder. Int J Mol Sci. 2017;18(10):65.
10. Chen HH, Zhang N, Li WY, et al. Overexpression of brain-derived neurotrophic factor in the hippocampus protects against post-stroke depression. Neural Regeneration Res. 2015;10(9):1427–1432.
11. Li J, Zhao YD, Zeng JW, Chen XY, Wang RD, Cheng SY. Serum Brain-derived neurotrophic factor levels in post-stroke depression. J Affect Disord. 2014;168:373–379.
12. Zhang E, Liao P. Brain-derived neurotrophic factor and post-stroke depression. J Neurosci Res. 2020;98(3):537–548.
13. Bastiaanssen TFS, Cussotto S, Claesson MJ, Clarke G, Dinan TG, Cryan JF. Gutted! Unraveling the role of the microbiome in major depressive disorder. Harv Rev Psychiatry. 2020;28(1):26–39.
14. Genedi M, Janmaat IE, Haarman B, Sommer IEC. Dysregulation of the gut-brain axis in schizophrenia and bipolar disorder: probiotic supplementation as a supportive treatment in psychiatric disorders. Curr Opin Psychiatry. 2019;32(3):185–195.
15. Nguyen TT, Hathaway H, Kosciolek T, Knight R, Jeste DV. Gut microbiome in serious mental illnesses: a systematic review and critical evaluation. Schizophr Res. 2021;234:24–40.
16. Szeligowski T, Yun AL, Lennox BR, Burnet PWJ. The gut microbiome and schizophrenia: the current state of the field and clinical applications. Front Psychiatry. 2020;11:156.
17. Jiang W, Gong L, Liu F, Ren Y, Mu J. Alteration of gut microbiome and correlated lipid metabolism in post-stroke depression. Front Cell Infect Microbiol. 2021;11:663967.
18. Beger RD, Dunn W, Schmidt MA, et al. Metabolomics enables precision medicine: “A White Paper, Community Perspective”. Metabolomics. 2016;12(10):149.
19. Chen JJ, Zhou CJ, Zheng P, et al. Differential urinary metabolites related with the severity of major depressive disorder. Behav Brain Res. 2017;332:280.
20. Gupta M, Neavin D, Liu D, et al. TSPAN5, ERICH3 and selective serotonin reuptake inhibitors in major depressive disorder: pharmacometabolomics-informed pharmacogenomics. Mol Psychiatry. 2016;21(12):1717–1725.
21. Zhu H, Bogdanov MB, Boyle SH, et al. Pharmacometabolomics of response to sertraline and to placebo in major depressive disorder - possible role for methoxyindole pathway. PLoS One. 2013;8(7):e68283.
22. Chen J, Lv YN, Li XB, et al. Urinary Metabolite Signatures for Predicting Elderly Stroke Survivors with Depression. Neuropsychiatr Dis Treat. 2021;17:925–933.
23. Liang ZH, Jia YB, Li ZR, et al. Urinary biomarkers for diagnosing poststroke depression in patients with type 2 diabetes mellitus. Diabetes Metab Syndr Obes. 2019;12:1379–1386.
24. Xie J, Han Y, Hong Y, et al. Identification of Potential Metabolite Markers for Middle-Aged Patients with Post-Stroke Depression Using Urine Metabolomics. Neuropsychiatr Dis Treat. 2020;16:2017–2024.
25. Zhang W, Zhang XA. A novel urinary metabolite signature for non-invasive post-stroke depression diagnosis. Cell Biochem Biophys. 2015;72(3):661–667.
26. Yang ES, Li PW, Nilius B, Li G. Ancient Chinese medicine and mechanistic evidence of acupuncture physiology. Pflugers Arch. 2011;462(5):645–653.
27. Highfield ES, Lama P, Grodin MA, Kaptchuk TJ, Crosby SS. Acupuncture and traditional Chinese medicine for survivors of torture and refugee trauma: a descriptive report. J Immigrant Minority Health. 2012;14(3):433–440.
28. Wang X, Cai W, Wang Y, Huang S, Zhang Q, Wang F. Is electroacupuncture an effective and safe treatment for poststroke depression? An updated systematic review and meta-analysis. Biomed Res Int. 2021;2021:8661162.
29. Li P, Huang W, Yan YN, et al. Acupuncture can play an antidepressant role by regulating the intestinal microbes and neurotransmitters in a rat model of depression. Med Sci Monit. 2021;27:e929027.
30. Cai W, Ma W, Wang GT, Li YJ, Shen WD. Antidepressant, anti-inflammatory, and antioxidant effects of electroacupuncture through sonic hedgehog-signaling pathway in a rat model of poststroke depression. Neuropsychiatr Dis Treat. 2019;15:1403–1411.
31. Zhang K, Liu R, Zhang J, et al. Electroacupuncture Ameliorates Depression-Like Behaviour in Rats by Enhancing Synaptic Plasticity via the GluN2B/CaMKII/CREB Signalling Pathway. Evid Based Complement Alternat Med. 2021;2021:2146001.
32. Belayev L, Alonso OF, Busto R, Zhao W, Ginsberg MD. Middle cerebral artery occlusion in the rat by intraluminal suture. Neurological and pathological evaluation of an improved model. Stroke. 1996;27(9):1616–1622.
33. Willner P, Muscat R, Papp M. Chronic mild stress-induced anhedonia: a realistic animal model of depression. Neurosci Biobehav Rev. 1992;16(4):525–534.
34. Gomaa EZ. Human gut microbiota/microbiome in health and diseases: a review. Antonie Van Leeuwenhoek. 2020;113(12):2019–2040.
35. Passos M, Moraes-Filho JP. Intestinal Microbiota in Digestive Diseases. Arq Gastroenterol. 2017;54(3):255–262.
36. Adak A, Khan MR. An insight into gut microbiota and its functionalities. Cell Mol Life Sci. 2019;76(3):473–493.
37. Limbana T, Khan F, Eskander N. Gut Microbiome and Depression: how Microbes Affect the Way We Think. Cureus. 2020;12(8):e9966.
38. Nikolova VL, Smith MRB, Hall LJ, Cleare AJ, Stone JM, Young AH. Perturbations in gut microbiota composition in psychiatric disorders: a review and meta-analysis. JAMA Psychiatry. 2021;78(12):1343–1354.
39. Cryan JF, Dinan TG. Mind-altering microorganisms: the impact of the gut microbiota on brain and behaviour. Nat Rev Neurosci. 2012;13(10):701–712.
40. Cryan JF, O’Riordan KJ, Cowan CSM, et al. The microbiota-gut-brain axis. Physiol Rev. 2019;99(4):1877–2013.
41. Qiu X, Li Z, Huang S, et al. Electroacupuncture improves depression-like behavior by regulating the abundance of lactobacillus and staphylococci in mice. J Integr Neurosci. 2023;22(2):28.
42. Chidambaram SB, Essa MM, Rathipriya AG, et al. Gut dysbiosis, defective autophagy and altered immune responses in neurodegenerative diseases: tales of a vicious cycle. Pharmacol Ther. 2022;231:107988.
43. Zheng P, Zeng B, Zhou C, et al. Gut microbiome remodeling induces depressive-like behaviors through a pathway mediated by the host’s metabolism. Mol Psychiatry. 2016;21(6):786–796.
44. Ha HY, Kim KS, Yeom YI, Lee JK, Han PL. Chronic restraint stress massively alters the expression of genes important for lipid metabolism and detoxification in liver. Toxicol Lett. 2003;146(1):49–63.
45. Hussain G, Wang J, Rasul A, et al. Role of cholesterol and sphingolipids in brain development and neurological diseases. Lipids Health Dis. 2019;18(1):26.
46. Virtanen M, Ferrie JE, Akbaraly T, et al. Metabolic syndrome and symptom resolution in depression: a 5-year follow-up of older adults. J Clin Psychiatry. 2017;78(1):e1–e7.
47. Weina H, Yuhu N, Christian H, Birong L, Feiyu S, Le W. Liraglutide attenuates the depressive- and anxiety-like behaviour in the corticosterone induced depression model via improving hippocampal neural plasticity. Brain Res. 2018;1694:55–62.
48. Jung J, Lee SM, Lee MJ, et al. Lipidomics reveals that acupuncture modulates the lipid metabolism and inflammatory interaction in a mouse model of depression. Brain Behav Immun. 2021;94:424–436.
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.