Back to Journals » Nature and Science of Sleep » Volume 14
Effects of Caffeine Intake on Cognitive Performance Related to Total Sleep Deprivation and Time on Task: A Randomized Cross-Over Double-Blind Study
Authors Quiquempoix M ![]() , Sauvet F
, Sauvet F ![]() , Erblang M, Van Beers P, Guillard M, Drogou C, Trignol A, Vergez A, Léger D
, Erblang M, Van Beers P, Guillard M, Drogou C, Trignol A, Vergez A, Léger D ![]() , Chennaoui M, Gomez-Merino D, Rabat A
, Chennaoui M, Gomez-Merino D, Rabat A
Received 11 October 2021
Accepted for publication 7 December 2021
Published 16 March 2022 Volume 2022:14 Pages 457—473
DOI https://doi.org/10.2147/NSS.S342922
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Prof. Dr. Ahmed BaHammam
Michael Quiquempoix,1,2 Fabien Sauvet,1,2 Mégane Erblang,1– 3 Pascal Van Beers,1,2 Mathias Guillard,1,2 Catherine Drogou,1,2 Aurélie Trignol,1,2 Anita Vergez,1,2 Damien Léger,2,4 Mounir Chennaoui,1,2 Danielle Gomez-Merino,1,2 Arnaud Rabat1,2
1Department of Operational Environments, Fatigue and Vigilance Team, French Armed Forces Biomedical Research Institute (IRBA), Paris, France; 2VIFASOM Team (EA 7330), University of Paris - Hôtel Dieu AP-HP Hospital, Paris, France; 3LBEPS, Univ Evry, IRBA, University of Paris-Saclay, Paris, France; 4Centre du sommeil et de la vigilance, Hôpital Hôtel Dieu AP-HP, Paris, 75004, France
Correspondence: Michael Quiquempoix, Department of operational environments, Fatigue and Vigilance Team, Armed Forces Biomedical Research Institute (IRBA), 1, avenue du Général Valérie André, BP 73, Brétigny sur Orge, Paris, 91223, France, Email [email protected]
Introduction: It is widely admitted that both total sleep deprivation (TSD) and extended task engagement (Time-On-Task, TOT) induce a cognitive fatigue state in healthy subjects. Even if EEG theta activity and adenosine both increase with cognitive fatigue, it remains unclear if these modifications are common mechanisms for both sustained attention and executive processes.
Methods: We performed a double-blind counter-balanced (placebo (PCBO) and caffeine (CAF) - 2× 2.5 mg/kg/24 h)) study on 24 healthy subjects (33.7 ± 5.9 y). Subjects participated in an experimental protocol including an habituation/training day followed by a baseline day (D0 and D1) and a total sleep deprivation (TSD) day beginning on D1 at 23:00 until D2 at 21:00. Subjects performed the psychomotor vigilance test (PVT) assessing sustained attention, followed by the executive Go-NoGo inhibition task and the 2-NBack working memory task at 09:15 on D1 and D2.
Results: We showed differential contributions of TSD and TOT on deficits in sustained attention and both executive processes. An alleviating effect of caffeine intake is only observed on sustained attention deficits related to TSD and not at all on TOT effect. The caffeine dose slows down the triggering of sustained attention deficits related to TOT effect.
Discussion: These results suggest that sustained attention deficits induced by TSD rely on the adenosinergic mechanism whereas TOT effect observed for both sustained attention and executive would not.
Keywords: total sleep deprivation, time on task, caffeine, sustained attention, inhibition, working memory, cognitive fatigue, mental fatigue, healthy subjects
Introduction
Cognitive/Mental fatigue, a general process associated with a sensation of exhaustion and lack of energy and resulting in a decrease of cognitive capacity,1 is involved in 4 to 50% of accidents (eg road, aviation, railway, etc.) when human error is engaged (for review see2). The longer subjects are engaged in a demanding cognitive task (Time-On-Task – TOT - effect), the more impaired they will be on sustained attention or other attention and executive processes (for review1,3). Cognitive/mental fatigue could be the outcome of an imbalance between energy demand/mental resource4,5 and/or the outcome of a personal evaluation concerning the cognitive demand of the task.1,6
Accidents related to cognitive fatigue are often observed with sleepy subjects.2 Sleep, a well-regulated physiological process, is under the control of two factors: 1) the homeostatic factor (the longer the time spent awake the greater the drive for sleep) and 2) the circadian one that varies with a 24-hour periodicity independently of the amount of preceding sleep or wakefulness (for review see7). Total sleep deprivation (TSD), which is characterized as a non-physiological prolonged duration of wakefulness is an extreme situation inducing both subjective (complaint) and objective (reduction in latency to fall asleep) sleepiness state (for review see8). TSD is also responsible for deficits both on sustained attention, executive and higher cognitive processes (such as planning, reasoning, etc.) in healthy individuals9,10 (for review see8).
Time on task (TOT) and sleepiness (ie, TSD) both contribute to cognitive/mental fatigue. Sustained attention deficits related to TOT during 20-min of the Psychomotor vigilance test (PVT) have been observed in well-rested subjects, with significant increase of self-reported sleepiness and mental fatigue.11,12 In addition, Gui et al found that impaired spontaneous activity and connectivity in the resting-state DMN (default mode network) are associated with TOT effects and mental fatigue during a PVT of 20 minutes.12 However subjective fatigue and sleepiness were shown increased and driving performance deteriorated during a simulate monotonous night-time rural driving, concomitantly to increased EEG delta, theta and alpha activity, while breaks improved driving and fatigue ratings but did not affect EEG activity and sleepiness.13 During a neuropsychological test battery of approximately 90 minutes, significant increase of EEG-relative power of theta activity was evidenced in frontal-central and parietal regions14 and a positive correlation was found between PVT deficits related to TSD and changes in EEG theta activity particularly in centro-posterior brain areas.15 The effects of TOT and TSD are evidenced in overlapping brain areas at least for sustained attention tasks.16–18 Wascher et al have also observed an increase of frontal theta power of subjects cognitively fatigued (ie, a sensorimotor decision making during 4 hours).16 The same results have also been observed in TSD situations.19–21
Both cognitive deficits related to TSD and TOT are also dependent of brain metabolic resources and this was shown in the frontal cortex notably for TSD.19,22 Interestingly, Martin et al, proposed that mental fatigue causes localized changes in cerebral fuel stores, which in turn contribute to changes in cerebral adenosine that acts on motivation during a prolonged and demanding cognitive task through the interaction between adenosine and dopamine receptors.23 It is interesting to note that TOT effects were firstly revealed during a 20-min PVT task with an increase of response time, and secondly evidenced as associated with the dopamine system (the DAT1 transporter precisely).11 Adenosine is a neuromodulator that reduces neural activity via binding to adenosine receptors, mainly the A1 and A2A receptors.24,25 Adenosinergic mechanisms have been repeatedly shown to regulate sustained and selective attention deficits associated with TSD, sometimes using caffeine, the non-selective competitive adenosine receptor antagonist.25–27 Caffeine administration was evidenced both to reduce subjective sleepiness and to limit the cognitive impairment on sustained attention induced by TSD (for review see25). Nevertheless, we found no data 1) relative to TOT effects during TSD on executive tasks such as inhibition and working memory and 2) regarding caffeine effects on cognitive deficits related to TOT under TSD.
In this study, we sought to (i) determine the effects of TSD and TOT on three different cognitive processes (sustained attention, motor inhibition, and working memory) and EEG theta power, and (ii) further examine their underlying mechanisms through the influence of caffeine intake. Our interest in EEG theta power originates from our previous study, which showed significant positive correlations between theta power in the centro-temporal brain region and the number of PVT lapses after TSD under caffeine or placebo conditions, whereas no significant correlation was observed for alpha power.28
Methods
Participants
24 healthy subjects (33.7±5.9 years), after giving their informed written consent, were included in this randomized cross-over and double-blind study that received the agreement of the Cochin–CPP Ile de France IV (Paris) Ethics Committee and of the French National Agency for Medicines and Health Products Safety (ANSM, ID RCB Number: 2017-A02793-50). It was also conducted according to the principles expressed in the Declaration of Helsinki of 1975 as revised in 2001 and in accordance with the full trial protocol (PERCAF) that has been recorded (NCT03859882, 25/02/2019). Subjects had not travelled between time zones within 7 days prior the study. They were free from medical, psychiatric and sleep disorders and were excluded if they self-reported any use of medications with sleep related side effects and illicit drugs. Subjects were also excluded if they reported a daily caffeine intake greater than 500 mg in order to minimize caffeine withdrawal effects during the protocol, and they were asked to reduce their caffeine intake 48 hours before the protocol. Exclusion criteria additionally included physical or mental health troubles based on (I) Hospital Anxiety and Depression scale, HAD ≥ 16, (II) significant medical history, (III) Epworth Sleepiness Scale, ESS > 11, (IV) Pittsburg sleep quality index, PSQI > 6, (V) morningness-eveningness questionnaire < 31 or > 69, (VI) habitual time in bed per night < 6 hours.10
Subjects completed a sleep/wake schedule for one week prior to the study. Their average daily caffeine consumption was 254 ± 190 mg (mean ± SD). The characteristics of our subjects are summarized in Table 1.
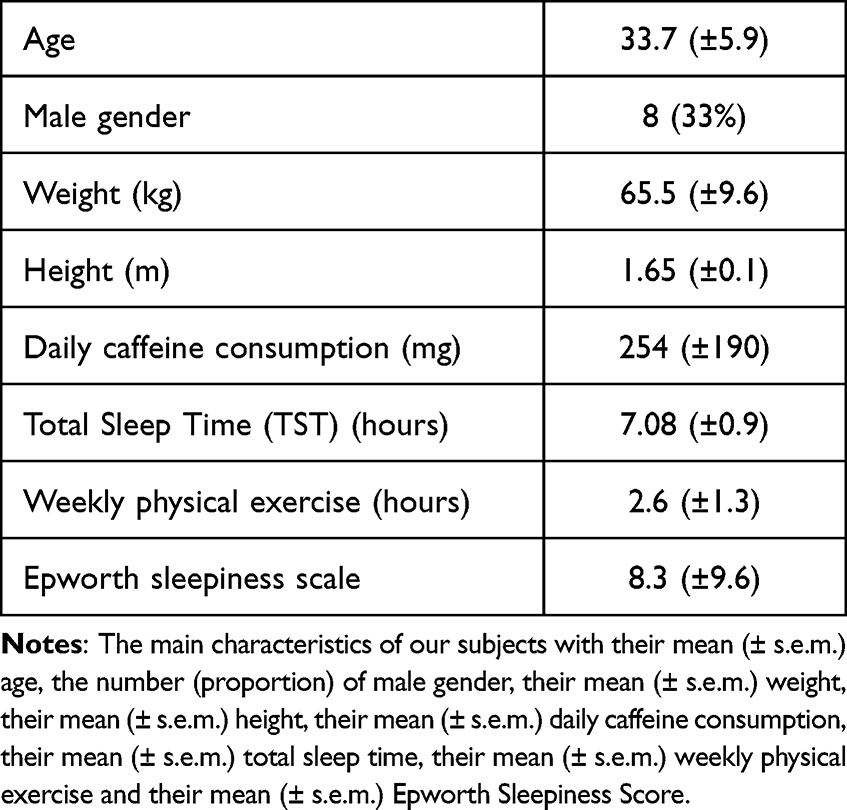 |
Table 1 Subjects Characteristics |
Behavioural Tests and Parameters
Karolinska Sleeping Scale (KSS) for Sleepiness
Subjective sleepiness was assessed on a single-item scale using the Karolinska Sleepiness Scale (KSS). This 9-points scale, based on a self-reported, assess awake/sleepiness rating.29 The different levels used for ratings were: 1 = very alert, 3 = alert, 5 = neither alert nor sleepy, 7 = sleepy (but not fighting sleep), 9 = very sleepy (fighting sleep). Our computer version enables subject to choose out of the nine given options just before starting the PVT.
Psychomotor Vigilance Task (PVT) for Sustained Attention
We utilized a computer-based version of the 10-min PVT,9 that is detailed in previous studies.9,10 Results are expressed as Response Time (RT) and number of lapses. Lapses were defined as Response Time (RT) > 500 ms and RT were ranged from 150 to 500 ms. We have also analyzed both 10% fast and 10% slow RT.30
Executive Task N°1: Go-NoGo for Motor Inhibition
In this first executive task,10 subjects were required either to respond or not to respond when a visual stimulus (a white arrow) arrived on a screen (black). A detailed version of this task is described in a previous studies.10 In our study, the proportion of “Go” and “No-Go” responses were respectively 75% and 25% of total trials. The total duration of the task is 10 min. Results are expressed with the rate of errors by commission (ie, responding “Go” on a “No-Go” trial) or the rate of omission (ie, no response on a “Go” trial) and the Response Time (RT).
Executive Task N°2: 2N-Back for Working Memory
In this second executive (visual working memory) task, pseudo-random sequences of letters appeared in the center of the screen and subjects have to respond to specified letters. A detailed version of this task is described in a previous studies.10 Briefly this task combined two conditions: (I) a control condition “0N-Back” consisting in responding to a specific letter (ie, letter W, also permanently displayed at the bottom of the screen during the block) and (II) a working memory “2N-Back” condition where participants had to respond whenever the current letter is identical to the letter presented two trials back (ie, M-X-M). The total duration is around 10 min. Results are expressed as response time (RT) and percentage of correct responses (CR).10
Time on Task (TOT) Analysis
Time on task (TOT) effect was visually inspected minute by minute and quantitatively assessed by comparing the first 3 and the last 3 minutes on PVT (RT and number of lapses but not for RT 10% fastest and 10% slowest due to insufficient number of trials), Go/No-Go (RT, rate of No-Go errors) and 2N-Back tasks (TR, rate of correct responses). This was chosen i) to approximately estimate time on task kinetic and ii) because a 3-minute task is often used to evaluate cognitive fatigue.31
Study Design and Testing Conditions
This is a laboratory-based, double-blind, placebo-controlled, and crossover study, with participants randomly assigned to either a caffeine or a placebo condition (Figure 1). The full trial protocol can be accessed by request to the corresponding author. The assignment of participants (caffeine (CAF) or placebo (PCBO)) has been made using the order of inscription to the study by an independent member of the staff following a random list. The randomized plan has been made in order to have 2 subjects with caffeine and 2 subjects with placebo in each session. Participant and staff members were blind for treatments. The in-laboratory experimental protocol included: (I) a habituation/training day (D0), (II) a baseline day (D1), (III) a total sleep deprivation (TSD) day beginning on D1 at 23:00 until D2 at 21:00, and (IV) a recovery night (from 21:00 until 09:00) for safety reasons so that the subjects can recover before returning to their usual life. Caffeine or placebo was administrated at 08:30 and 14:30 on D1 and at 08:30 on D2. The psychomotor vigilance task (PVT) was performed and the sleepiness scale (KSS) was completed at 09:15 at baseline (D1) and on the day of TSD (D2). The executive Go-NoGo test was performed just after each PVT on D1 and D2, followed by the 2-NBack task.
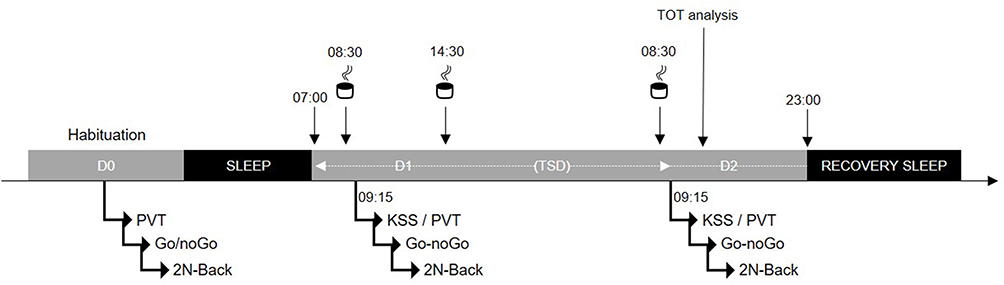 |
Figure 1 Experimental design. It includes an habituation/training day followed by a baseline day (D0 and D1) and a total sleep deprivation (TSD) day beginning on D1 at 23:00 until D2 at 21:00. Subjects completed the Karolinska Sleepiness Scale (KSS) and then performed cognitive tests at 09:15 on D1 and D2 (26h of awakening), and EEG was recorded during each test. (↓) 08:30 and 14:30, Placebo or caffeine (2.5 mg/kg). |
Subjects were maintained in an individual temperature-controlled (22 ± 1°C) room. Restroom and bathroom facilities were collectively available in the sleep-laboratory flat that contains a main living room (34 m2). Illumination was maintained between 150 and 300 lux during the entire experimental period (lights off during sleep periods). Subjects were prohibited from practicing exercise, taking tobacco, alcohol, or other psychoactive substances during the study. When not engaged in any specific testing or meals, subjects followed a standardized activity program (reading, watching videos, and playing games). In addition to the portable or wrist actimeters, investigators were systematically present in the laboratory with at least 1 of them with subjects. When subjects were about to fall asleep (eyes closed, head down), they were gently and immediately woken up (ie, no period of sleep > 30 seconds). During testing periods (morning D1 and morning D2), subjects were individually equipped (electrophysiological measurement) and monitor by an experimenter. All subjects had a systematic habituation/training period (D0 day) for behavioral tests in order to reduce learning or misunderstanding bias during the first set of tests.
Caffeine Administration
Caffeine (CAF) or placebo (PCBO) was administrated in decaffeinated beverage. During caffeine condition, caffeine powder was pre-measured by the project supervisor and amounted to 2.5 mg per kg body mass for each participant, then mixed with decaffeinated beverage. After review of literature, this amount of caffeine powder was chosen because a range from 0.2 to 5.5 mg/kg has been found to enhance response time in sleep-deprived conditions.32
Electrophysiological Measurements
EEG Procedure
EEG was recorded at 19 scalp sites, according to the international 10–20 system (Fp1, Fp2, F7, F3, Fz, F4, F8, T7, C3, Cz, C4, T8, P7, P3, Pz, P4, P8, O1, O2), with a Siesta 802 (Compumedics Limited, Victoria, Australia). EEG was recorded continuously at a sampling rate of 512 Hz referenced with bridged mastoidal electrodes. Data were re-referenced during preprocessing with a common average. Electrodes were interfaced with the scalp using EC2 gel (Grass Technologies, Astro-Med, Inc., West Warwick, RI, USA), and impedances were kept below 10 kOhm during the whole session. EEG was installed approximately 20–30 min before the first cognitive task (PVT), and supplemented with EC2 gel if needed, to prevent dry electrodes during recordings.
EEG Analysis
EEG data were analyzed in MatLab (MathWorks, Natick, MA, USA) with Fieldtrip toolbox33 and custom codes. Data were bandstop filtered between 48 and 52 Hz to remove electrical noise, high pass filtered above 0.1 Hz and locally detrended.
Blink and saccade artefacts were removed by computing an Independent Component Analysis (ICA, Fieldtrip) and visually discarding components that had either prefrontal distribution and blink-related waveform or a fronto-temporal distribution and a saccade-related waveform. Movement artefacts were removed by visual inspection on a 10 seconds time window basis. If the first 3 min or last 3 min of each cognitive test presented more than 30 seconds of movement artifacts, the electrode was rejected. If during the 10 min of each cognitive test there were more than 120 seconds of cumulated artifacts, the electrode was rejected. Finally, if more than 3 (out of 19) electrodes were removed, the subject was excluded from EEG analysis. EEG theta power (4–8 Hz) was assessed by using continuous Morlet wavelets transform during the entire period of each cognitive test (10 min each). Regions of interest (ROI) represent the mean of grand averaged (all subjects) theta power over frontal (Fp1, Fp2, F7, F3, Fz, F4, F8), Centro-Temporal (T7, C3, Cz, C4, T8) and Parieto-Occipital (P7, P3, Pz, P4, P8, O1, O2) regions. We assessed effects of CAF and TOT (first 3 min vs last 3 min of test, see Time on Task (TOT) Analysis) during D2 for all cognitive tasks.
Statistical Analysis
Data has been collected in the French armed forces biomedical institute (IRBA) in Brétigny sur Orge. All data in text, tables and figures are presented as mean ± standard error of the mean (SEM). Statistical analyses were performed using Statistica 10.0 (StatSoft R). All value’s distributions were tested for their normality (Kolmogorov–Smirnov, Shapiro–Wilk and Lilliefors tests). The primary objective was to investigate the efficacy of caffeine (CAF) compared to placebo (PCBO) for improving sustained attention during a total sleep deprivation (TSD) protocol in healthy subjects, using the psychomotor vigilance test (PVT). In the literature, the 09:15 hour corresponds to the highest decrease of sustained attention during total sleep deprivation, due to a continuous awakening period (sleep pressure) and the nadir of circadian rhythm.34 Secondary objectives were to assess efficacy of CAF on executive functions with Go-NoGo and 2N-Back performances and interaction between CAF and time on task (TOT). With 24 subjects, the study has at least 80% power to detect a significant difference in the primary endpoint when comparisons are made at the unadjusted 2‐sided 5% level. Powering was made assuming an effect size equal to 1.3 and a true within subject standard error of 37.9 ms in response time in the PVT, as reported for placebo treatment in our laboratory.34 When the value’s distribution was normal (at least two negative tests), a two-way repeated measures ANOVAs were conducted in order to test 1) treatment condition (CAF or PCBO), TSD effect and their interaction and 2) treatment condition (CAF or PCBO), Time-On-Task (TOT) effect and their interaction. When value’s distribution was not normal, we used a one way non-parametric ANOVA test with repeated measures (ANOVA of Friedman and Wilcoxon test for post hoc comparison: W) to identify differences inside and between each factor (ie, treatment condition and TOT).
Results
We measured mean response time (RT) and number of lapses during PVT, RT and percentage of errors during Go-NoGo and RT and percentage of correct responses during 2N-Back tasks while performing EEG measurements for each test.
Effect of TSD and CAF on Subjective Sleepiness: Karolinska Sleepiness Scale (KSS)
We observed global day and treatment effects on KSS score that resulted in a significant increase of KSS score after TSD (D2) compared to Baseline (D1) for both PCBO and CAF conditions (Table 2). We did not observe any significant difference between PCBO and CAF at D1 and D2 (respectively p>0.11 and p>0.24).
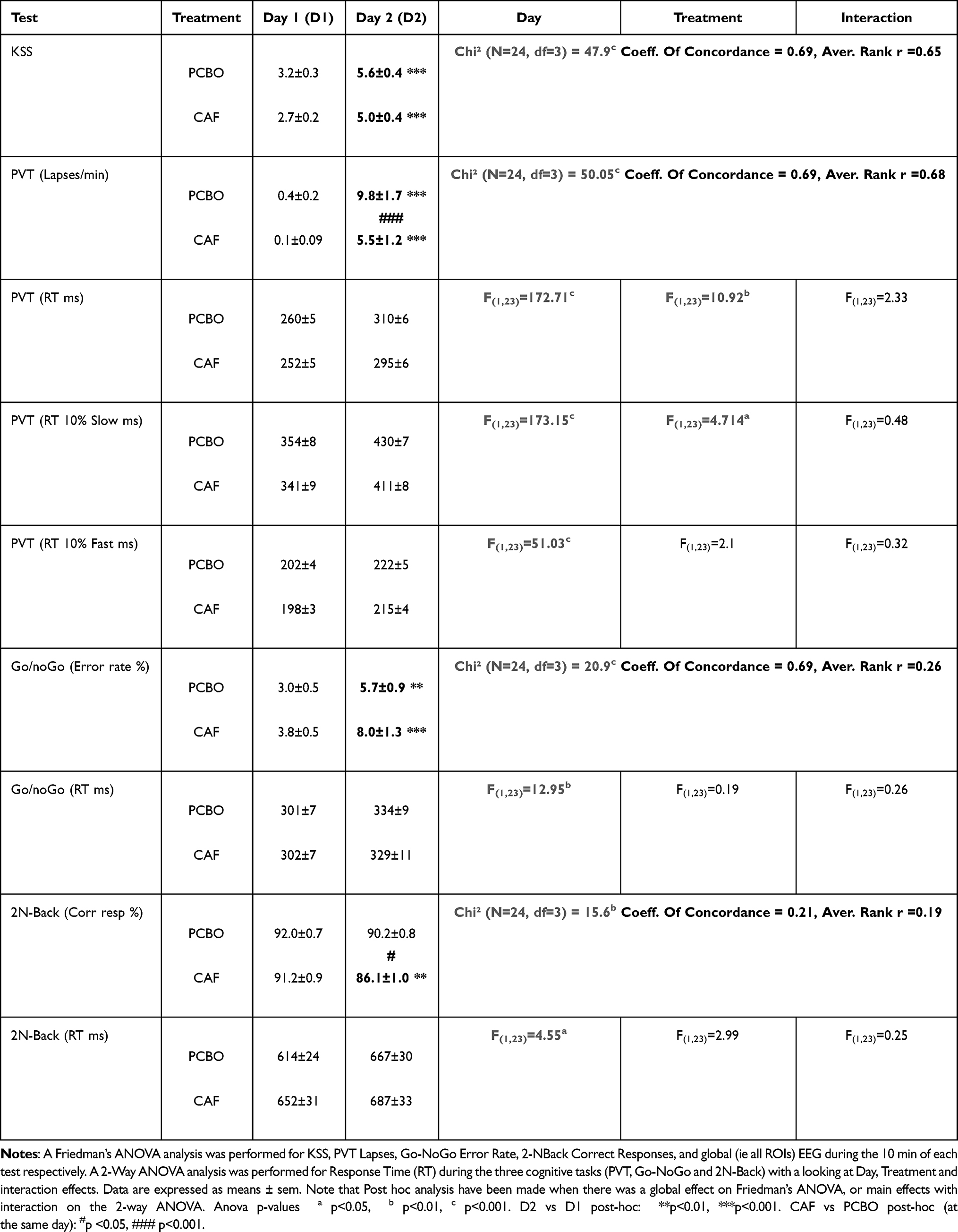 |
Table 2 Global Effects of Total Sleep Deprivation (Day Effect, D2 vs D1) and Treatment (CAF vs PCBO) on Cognitive Performances |
Effects of TSD, CAF and TOT on PVT Lapses
Concerning the number of lapses accumulated during 10 min of PVT, we observed a global day and/or treatment effect (Table 2). Post hoc analyses showed a significant increase in the number of lapses at D2 compared to D1 both in PCBO and CAF conditions (Table 2). We also observed significantly lower lapses in CAF compared to PCBO at D2 (Table 2). Since the number of lapses was affected by TSD in both PCBO and CAF conditions, we looked at Time-On-Task (TOT) and treatment effects and their interaction. To differentiate TSD effects from a mixture of TSD and TOT on the number of lapses, we compared the first 3 min (0–3 min, TSD effect) of PVT with the last 3 min (7–10 min, TSD + TOT effects) (Figure 2A, shaded areas). There was a significant TSD effect (D2 vs D1) during the first 3 min of test in both PBCO and CAF conditions (p<0.001 for PCBO and p=0.015 for CAF condition). Significant TOT and treatment effects were observed on the number of lapses. More precisely at D2, the number of lapses per minute was significantly higher at the last 3 min (7–10min) compared to the first 3 min (0–3min) for both PCBO and CAF conditions (Table 3, Figure 2B). In contrast, we observed that the number of lapses is significantly lower at D2 in the CAF compared to PCBO condition for both the first 3 and the last 3 min of PVT (Figure 2B). We approximately estimated the kinetic of TOT by computing the difference of the number of lapses between 0–3 min and 7–10 min (Delta [7–10 min] – [0–3 min]), and found no significant difference between PCBO and CAF at D2 (p = 1.0; Figure 2C).
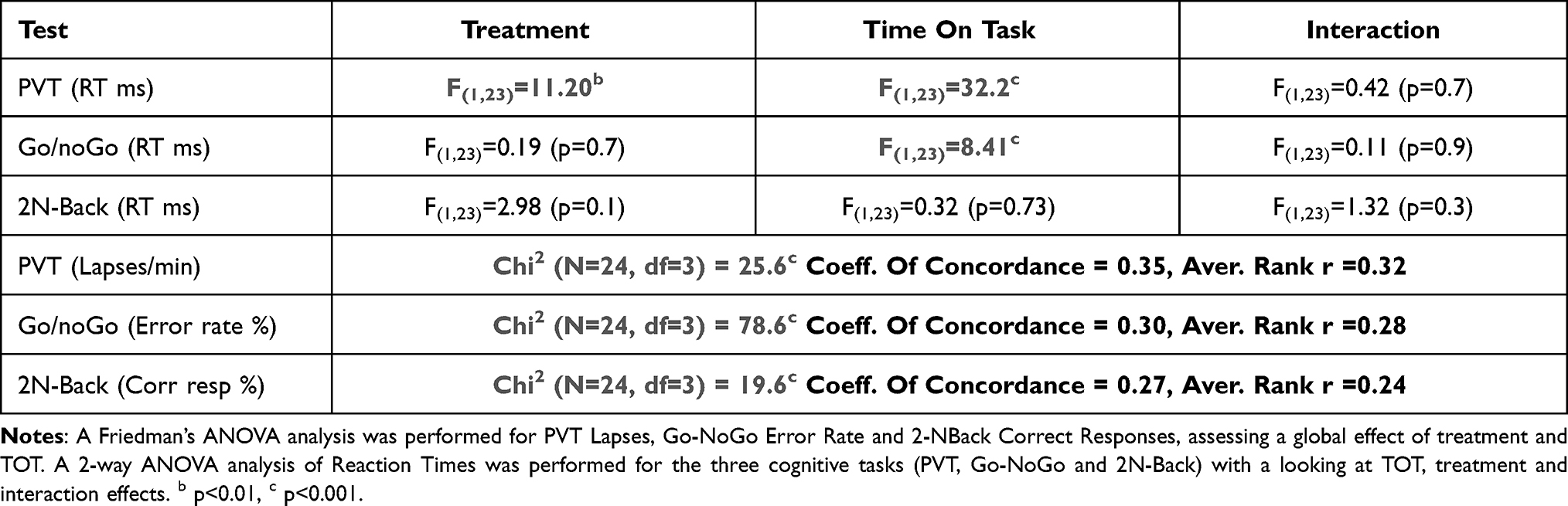 |
Table 3 Effects of Time on Task (TOT, First 3 Min vs Last 3 Min) and Treatment (CAF vs PCBO) on Cognitive Performances at D2 Day (After Total Sleep Deprivation) |
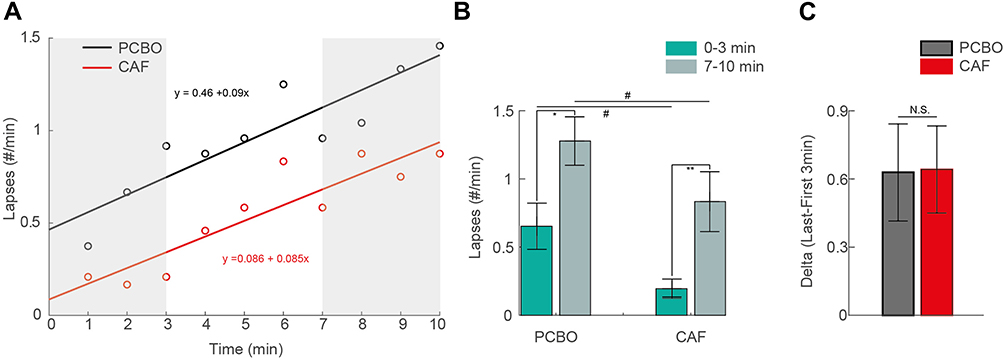 |
Figure 2 Caffeine reduces TSD-related sustained attention deficits without any effects on TOT (A) Kinetic of the number of lapses min−1 as a function of time during the psychomotor vigilance task (PVT) at D2 (after total sleep deprivation (TSD)) for both placebo (PCBO, black) and caffeine (CAF, red) conditions, fitted with a linear regression. Shaded areas show time points of interest further analyzed: the first 3 min (0–3 min) and the last 3 min (7–10 min) of test. (B) Number of lapses min−1 during the first and the last 3 min of PVT at D2 for PCBO and CAF conditions. Error bars show s.e.m. *Significant difference between 0–3 min and 7–10 min of the test (*p<0.05; **p<0.01). #Significant difference between PCBO and CAF conditions (#p<0.05). (C) Algebraic difference in the number of lapses min−1 between 0–3 min and 7–10 min of the task for both PCBO and CAF conditions. Abbreviations: N.S, no significant difference. |
Effects of TSD, CAF and TOT on No-Go Errors
We observed a global day and/or treatment effect for the rate of No-Go errors over the 10 minutes of test (Table 2). More precisely, a significant increase in the rate of No-Go errors was observed at D2 compared to D1 both for PCBO and CAF conditions with no statistical difference between the two conditions (Table 2). We then looked at the influence of TOT and treatment factors and their interaction. Interestingly, we observed no significant increase in the rate of No-Go errors in the first 3 min at D2 compared to D1 for both PBCO and CAF groups (p=0.47 for PCBO and p=0.17 for CAF). At D2 (after TSD), we observed a significant increase of No-Go errors between the first and the last 3 min of the task (Figure 3A, shaded areas) for both PCBO and CAF conditions (Table 3, Figure 3B) but with no statistical difference in the estimated kinetic of TOT between the two conditions (p = 0.98; Figure 3C) even if there was a significantly higher rate of No-Go errors in the first 3 min for CAF compared to PCBO condition (Figure 3B).
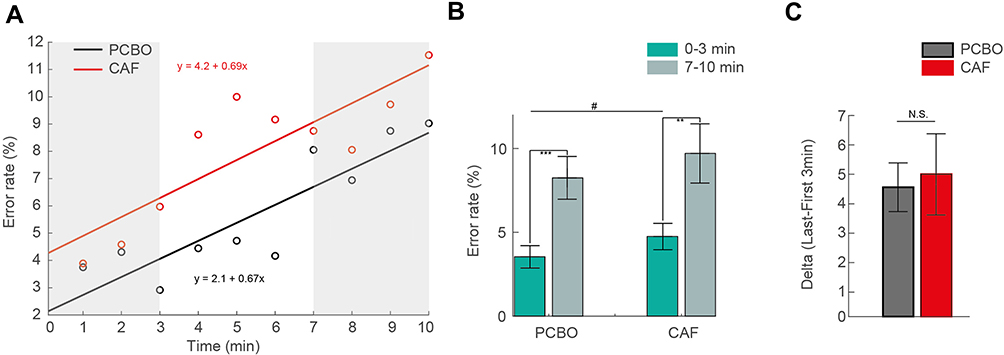 |
Figure 3 Lack of alleviating effects of caffeine on inhibition impairments related to TSD and TOT. (A) Kinetic of rate of No-Go errors (response on « No-Go » trials) during the Go/No-Go task at D2 (after TSD) for both placebo (PCBO, black) and caffeine (CAF, red) conditions, fitted with a linear regression. Shaded areas show the first 3 min (0–3 min) and the last 3 min (7–10 min) of test. (B) Rate of No-Go errors during the first and the last 3 min of test at D2 for both PCBO and CAF. Error bars show s.e.m. *Significant difference between 0–3 min and 7–10 min of the test (**p<0.01; ***p<0.001). #Significant difference between PCBO and CAF conditions (#p<0.05). (C) Algebraic difference in the rate of No-Go errors between the first and the last 3 min for both PCBO and CAF conditions. Abbreviations: N.S, no significant difference. |
Effects of TSD, CAF and TOT on 2N-Back Correct Responses
We observed a global day and/or treatment effect for the percentage of correct answers in 2N condition (Table 2) in PCBO and CAF conditions. A significant decrease of the percentage of correct responses (% CR) at D2 compared to D1 for CAF condition was observed but not for PCBO condition, with a significant difference between PCBO and CAF conditions at D2 (Table 2). We then looked at the influence of TOT and treatment factors and their interaction. We observed no significant increase in the 2N-Back correct responses in the first 3 min at D2 compared to D1 for both PBCO and CAF groups (p=0.261 for PCBO and p=0.215 for CAF). At D2 (after TSD), a significant effect of TOT and/or treatment was noticed (Table 3; Figure 4A). More precisely, a significant decrease of the % of CR in the last 3 min of the task compared to the first 3 min occurred in both PCBO and CAF conditions (Figure 4A and B) but with no significant difference between PCBO and CAF conditions during the first and the last 3 minutes (respectively p>0.10 and p>0.08; Figure 4B). Finally, with the approximated kinetic of TOT, we observed no statistical difference between PCBO and CAF at D2 (p = 0.48; Figure 4C).
 |
Figure 4 No beneficial effects of caffeine on working memory decrement after TSD and TOT (A) Kinetic of percentage of correct answers for the 2N-Back conditions at D2 (after TSD) for both placebo (PCBO, black) and caffeine (CAF, red) conditions, fitted with a linear regression. Shaded areas show the first 3 min (0–3 min) and the last 3 min (7–10 min) of test. (B) Percentage of correct answers during the first and last 3 min of test at D2 for both PCBO and CAF conditions. Error bars show s.e.m. *Significant difference between 0–3 min of 7–10 min (*p<0.05; **p<0.01). (C) Algebraic difference of rate of correct answers between the first 3 min and the last 3 min for PCBO and CAF conditions. Abbreviations: N.S, no significant difference. |
Effects of TSD, CAF and TOT on Response Time (RT) During PVT, Go-NoGo and 2N-Back
Significant main day effects and/or treatment effects on mean RT for these three cognitive tasks and also for the 10% fastest and the 10% slowest RT (just below lapses time period) for PVT were noticed (Table 2). More precisely, we showed that mean RT increased at D2 compared to D1 both for PCBO and CAF conditions during the three tasks (Table 2). We also observed a significant treatment main effect but only for mean RT and the 10% slowest RT for PVT, without any interaction (Table 2). Since all these behavioral parameters were affected by TSD, we looked at TOT and treatment effects and their interaction. We observed significant increase for PVT in the first 3 min at D2 compared to D1 for both PBCO and CAF groups (p<0.001 for PCBO and CAF respectively) and for Go-NoGo (p=0.015 for PCBO and p=0.048 for CAF respectively), and no significance for 2N-Back (p=0.628 for PCBO and p=0.876 for CAF). We observed a TOT effect on mean RT for both PVT and Go-NoGo but not for 2N-Back (Table 3). We also saw a significant treatment effect on mean RT but only for PVT and with any interaction with day (Table 2) or TOT (Table 3). More precisely, we showed at D2 that mean RT (ms) increased with TOT both for PCBO and CAF conditions during PVT (PCBO-D20-3min=301±7 vs PCBO-D27-10min=316±6; CAF-D20-3min=278±6 vs CAF-D27-10min=306±7) and Go-NoGo (PCBO-D20-3min=333±9 vs PCBO-D27-10min=335±9; CAF-D20-3min=328±11 vs CAF-D27-10min=330±12) but with a significant global decrease of PVT mean RT with caffeine (PCBO-D20-3min/7-10min=308±5 vs CAF-D20-3min/7-10min=292±5).
Effects of TSD, CAF and TOT on 0N-Back Correct Responses and RT
We observed a day effect for the percentage of % CR in 0N condition (Chi2(24,3)=26.5, p=1.3*10−5, Concordance Coeff. = 0.37, Aver. Rank = 0.34) in PCBO and CAF conditions. There was a significant decrease of correct responses at D2 compared to D1 for both PCBO (PCBO-D2=96.3±0.7% vs PCBO-D1=98.8±0.2%, p=1.9*10−3) and CAF conditions (CAF-D2=95.5±0.7% vs CAF-D1=98.3±0.4%, p=2.6*10−4), with no significant difference between conditions at D2 (PCBO-D2=96.3±0.7% vs CAF-D2=95.5±0.7%, p>0.20). At D2, no significant effect of TOT and treatment on the percentage of % CR was seen (Chi2(24,3)=5.0, p>0.17, Concordance Coeff.=0.07, Aver. Rank=0.03). Concerning mean RT, a significant day effect (F(1,23)=22,13; p=1.10−4) with no significant treatment effect (F(1,23)=0.905; p>0.35) and no interaction (F(1,23)=1,31; p>0.26) were noticed. More precisely, we showed that mean RT increased at D2 both for PCBO and CAF conditions (PCBOD2=465±18 vs PCBOD1=409±14, p<1.10−4; CAFD2=453±15 vs CAFD1=409±12, p<1.10−4).
Effects of TSD, TOT and CAF on EEG Theta Power
A global effect of day and/or treatment over the 10 min of the 3 tests was observed for EEG theta power in the 3 ROIs (Parieto-Occipital (PO), Centro-Temporal (CT) and Frontal (F)) (Table 4). Additional statistical effects of day and treatment on Alpha and Beta power are shown in the Supplementary Table S1.
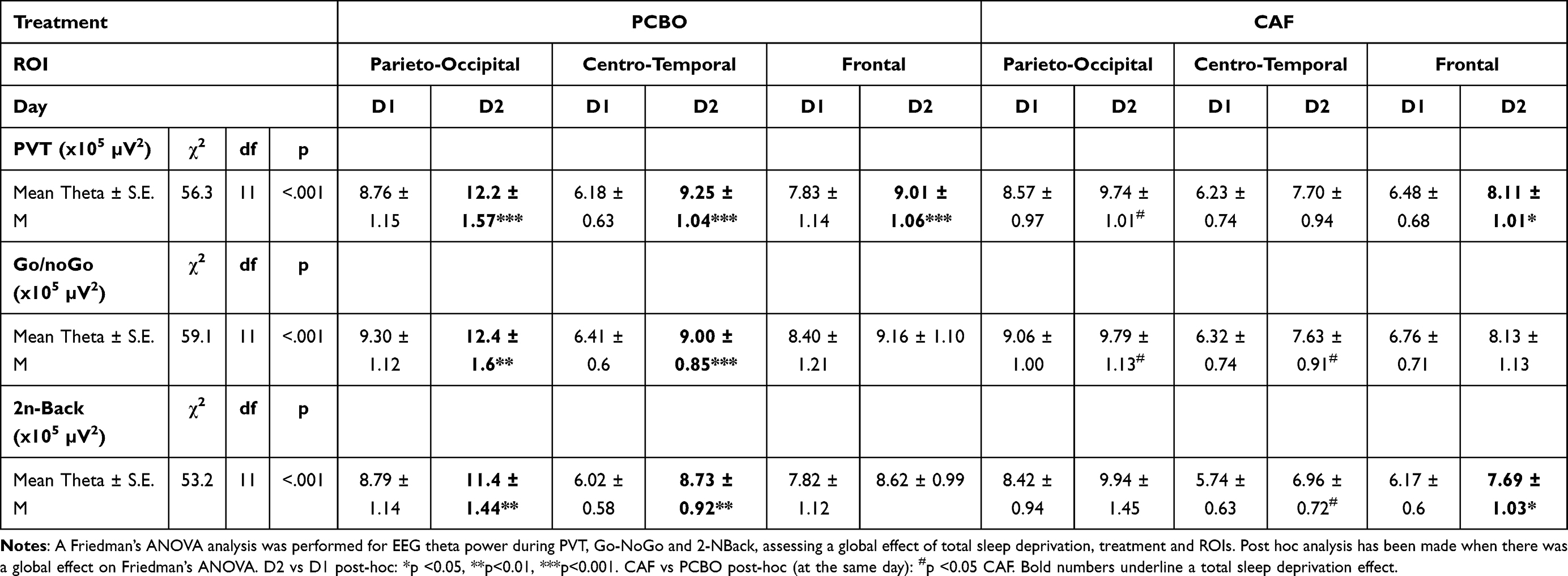 |
Table 4 Global Effects of Total Sleep Deprivation (Day Effect, D2 vs D1), Treatment (CAF vs PCBO) and Region of Interest (ROIs: Parieto-Occipital, Centro-Temporal, Frontal) on EEG Theta Power During the 3 Cognitive Tasks |
During the PVT Task
The theta power increase significantly in the 3 ROIs during PVT in the PCBO condition and only in the F brain area in the CAF condition (Table 4). At D2 (after TSD), a significant difference between PCBO and CAF is observed in the PO brain region. We then look at influence of TOT and treatment factors and their interaction among the three scalp ROI at D2 (Figure 5). Concerning PO brain region, we observed a TOT effect (F(1,20)=4,55; p <0.05), with no treatment (caffeine) effect (F(1,20)=3,62; p=0.07) and no interaction (F(1,20)=0,18; p=0.67). Concerning CT and F brain regions, we observed no effect of TOT (respectively, F(1,20)=1,46; p=0.24; F(1,20)=3,94; p=0.06) or treatment (caffeine) (respectively, F(1,20)=3,46; p=0.07; F(1,20)=1,18; p=0.29), and not interaction (respectively, F(1,20)=0,76; p>0.39; F(1,20)=0,15; p>0.70) (Figure 5A and D).
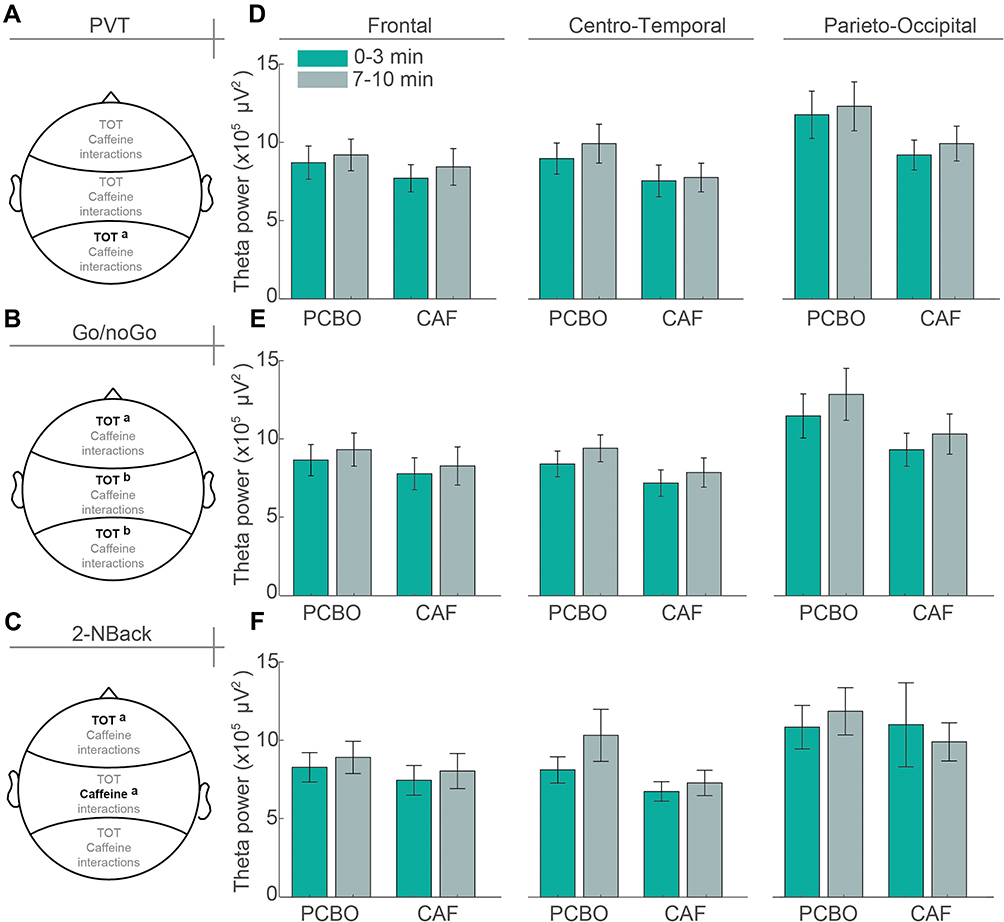 |
Figure 5 Caffeine and TOT effects on theta activity during the three cognitive tasks after TSD. On the left panel (A–C), topographical EEG representation of Time-On-Task (TOT) and caffeine significant (black in bold) or not significant (grey) effects at D2 (after TSD), and on the right panel (D–F), bar histograms of raw theta power in the 3 regions of interest (ROI) of the first 3 min and of the last 3 min, for PVT (upper), Go-NoGo (middle) and 2N-Back (lower). Anova p-values; ap<0.05, bp<0.01. |
During the Go-NoGo Task
The theta power increase significantly in the PO and CT brain regions during Go-NoGo in the PCBO condition, and a significant difference between PCBO and CAF is observed in the two regions at D2 (after TSD) (Table 4). We then look at influence of TOT and treatment factors and their interaction among the three scalp ROI at D2 (Figure 5). Concerning PO, CT and F brain ROIs, we observed a TOT effect (respectively, F(1,20)=8,16; p <0.01; F(1,20)=11,94; p <0.01 and F(1,20)=5,29; p<0.05), with no treatment (caffeine) effect (respectively, F(1,20)=2,87; p>0.10; F(1,20)=2,94; p>0.10 and F(1,20)=0,97; p>0.33) and no interaction (respectively, F(1,20)=0,32; p>0.58; F(1,20)=0,83; p>0.37 and F(1,20)=0,19; p>0.66) (Figure 5B and E).
During the 2N-Back Task
The theta power increase significantly in the PO and CT brain regions during 2N-Back in the PCBO condition and in the F region in the CAF condition (Table 4). A significant difference between PCBO and CAF is observed in the CT region at D2 (after TSD). We then look at influence of TOT and treatment factors and their interaction among the three scalp ROI at D2 (Figure 5). Concerning CT brain region, we observed a treatment (caffeine) effect (F(1,20)=7,98; p<0.05), with no TOT effect (F(1,20)=3,87; p>0.06) and no interaction (F(1,20)=1,61; p>0.22). Concerning F brain region, we observed no treatment (caffeine) effect (F(1,20)=1,16; p>0.29), but a TOT effect (F(1,20)=6,14; p<0.05) and no interaction (F(1,20)=0,01; p>0.91). Concerning PO brain region, we observed neither treatment (caffeine) effect (F(1,20)=0,19; p>0.66) nor TOT effect (F(1,20)=0.001; p>0.97) and no interaction (F(1,20)=1,11; p>0.30) (Figure 5C and F).
Discussion
Aims of our study were to determine the contribution of both TSD (Total Sleep Deprivation) and TOT (Time-On-Task) on sustained attention and executive processes (ie, inhibition and working memory) using caffeine and the contribution of EEG theta activity in such modifications. In this double-blind counter-balanced (PCBO placebo and CAF caffeine - 2.5 mg/kg) study, we demonstrate for the first-time differential effects of TSD and TOT on deficits of sustained attention and of two executive processes (inhibition and working memory). These cognitive capabilities were impaired by TSD and TOT when expressed in number of lapses (PVT), rate of errors (NoGo) and percentage of correct responses (2N-Back) tasks. More precisely, at the D2 day compared to the D1 day (corresponding to 26 hours of prolonged wakefulness), we observed a significant increase in the number of PVT lapses, the rate of No-Go errors and a decrease in the percentage of 2N-Back correct responses (only significant in the caffeine condition). In addition, we found a TSD per-se effect (D2 compared to D1 on the first three minutes of test) and a TOT effect (last vs first three minutes of the 10-min testing) in placebo and caffeine condition but differently regarding the three cognitive tasks. More precisely, the deleterious effect of TSD per-se was only observed for the increase in the number of Lapses (PVT) whereas the deleterious effect of TOT was observed for all the three cognitive tasks parameters (number of lapses, rate of No-Go errors and percentage of correct responses). This TOT effect is confirmed by the significant increase of theta power in at least one of the three brain regions of interest (ROI) during the three cognitive tasks and in the three ROIs during the Go-NoGo task, as previously shown in subjects engaged in sustained attention and executive tasks and fatigued with TOT.15,16 When we looked at response time, subjects under TSD presented a TOT effect only during PVT and Go-NoGo tasks.
Caffeine consumption significantly reduced sustained attention deficits (number of Lapses) related to TSD and the corresponding EEG theta spectral power without any reduction due to TOT (the last minus the first three minutes). Indeed, the linear downward shift of the number of lapses in the caffeine condition as shown in Figure 2A) and no caffeine effect on kinetics (slopes of regression lines that tend to be similar) suggested that under caffeine a subject that has not slept will degrade at the same rate but starting from a lower level, so caffeine delays the onset of sustained attention deficits due to TOT. The caffeine alleviating effect on the median and the 10% slowest RT (just below lapses time period) confirmed the delayed effect of caffeine on sustained attention deficits with also a significant decrease of theta power (during PVT) on the whole scalp. No such beneficial effect of caffeine was observed 1) on executive (inhibition and working memory) deficits related to TOT under TSD associated with just a global decrease of the EEG theta power on the whole scalp and 2) on the increase of subjective sleepiness.
All these results are in accordance with previous ones showing that a continuous period of wakefulness (upper than 24 hours) is associated with deficits of sustained attention and executive capacities (ie, inhibition and working memory) with higher sleepiness ratings9,10,15,35,36 (for review see8). We added the information that TOT contributes to sustained attention and executive processes deficits related to TSD as previously evidenced with studies using a 10-min PVT task.34–37 Even if post-hoc analysis showed that working memory capabilities were not significantly affected by TSD in the placebo condition as previously described,38,39 our results revealed a TOT effect under TSD. In agreement with Frenda and Fenn (2016), we argue that TSD impairs cognitive processes by primarily inhibiting the ability of individuals to be alert and to sustain their attention and allows other higher executive functions under compensatory mechanisms, explaining why working memory deficits observed in this study are mainly related to TOT (under TSD).40 This differential effect of caffeine on cognitive processes under TSD with a benefit for sustained attention deficits and not for two executive ones (inhibition and working memory) are also in accordance with our previous studies showing that countermeasures such as sleep extension and regular physical activity are ineffective to limit such executive deficits related to TSD.9,10,41 Lack of beneficial effect of caffeine on executive deficits and corresponding EEG theta power related to TSD and also on sleepiness could be due to the lower dose used (175 mg compared to 200–600 mg in other studies42,43) (for review see25). In any case, caffeine was found to be rarely or not at all effective in counteracting executive deficits related to TSD42 (for review see25). In contrast, previous studies have reported, as for us with RT during PVT, an improvement of response time for both simple and choice reaction time task related to TSD with caffeine.44
Sleep pressure that increases with TSD (for review see7) exert a deleterious effect on the capacity of subjects to stay awake, to be alert and thus to sustain their attention. With sustained attention, while waiting for the occurrence of a stimulus, subjects will be more easily distracted by their internal thoughts.45 Indeed, sleepy subjects that are often unaware of their own environment have a higher frequency of self-reported mind-wandering, an experience of thoughts both stimulus-independent and task-unrelated that seems to frequently co-occur with sleepiness.46 Interestingly, the brain default mode network (DMN) – a brain network whose activity is high when the mind is not engaged in specific behavioral tasks and low during focused attention on the external environment47 – is both associated with mind-wandering state47,48 and anti-correlated with a fronto-parietal network (central executive network: CEN) associated with cognitive control,49 especially in TSD situations.45,50 As we found a specific caffeine alleviating effect on the number of lapses related to TSD, we can make the assumption that under TSD, caffeine may help the brain of sleepy subjects to less often switch from a goal-directed to a goal-irrelevant brain function (ie, DMN) and to be less often mind wandered. This hypothesis is reinforced by our observation that caffeine alleviates TSD increases on i) the mean RT, ii) the 10% slowest RT on PVT, a behavioural parameter which has been linked to an activation of DMN30 and iii) the theta power of the subject’s brain during PVT (D2 vs D1). The increase of theta power during PVT following TSD has already been observed in sleepy subjects accompanied with sustained attention decrements15,51 or in a state of mind-wandering.52 Furthermore, a recent study demonstrated that taking a 200 mg caffeine pill increases brain entropy nearly in the entire cerebral cortex with higher effects in DMN and sensorimotor networks.53 These authors suggested that such pharmacological effects of caffeine on the human brain may result in an improvement of vigilance, attention and others functions which are mainly sub-served by the aforementioned brain regions.
In our study, the benefit of a low dose of caffeine (175 mg), a well-known antagonist of adenosine receptors, on the number of lapses, response time (mean and 10% slower) and theta power of subjects engaged in a PVT highlights the involvement of adenosinergic systems in sustained attention deficits related to TSD per-se.54 Executive deficits observed with this TSD protocol were mainly revealed with TOT. It thus seems that TSD per-se and TOT differentially contribute to cognitive deficits related to TSD. These results are in accordance with studies showing that TOT is responsible for inducing a mental fatigue state of subjects engaged in an executive task55–57 (for review see1). Furthermore, it has been observed that 35 hours of continuous wakefulness are associated with an increase of cerebral responses within parieto-frontal networks and with better working memory performance at most difficult load in a verbal learning or in logical reasoning.35,58,59 The frontal increase of theta power on the scalp related to TOT for both executive tasks is also in accordance with studies showing an increase of theta power of subjects engaged in an executive task and fatigued with TOT.14 The lack of beneficial effects of caffeine on such cognitive deficits (ie, executive) and on theta power related to TSD suggests that the neurobiological mechanisms implicated would be different and/or less sensitive to the subsequent increase of extracellular adenosine classically related to sustained attention deficits.25,54 Nevertheless, since working memory deficits related to TSD were worsen under caffeine consumption we can exclude the implication of adenosine on such executive process. Indeed, in a previous study with healthy subjects, caffeine administration (6 mg/kg) lowered the increase of perceived exertion (RPE) and amplitude of motor-related cortical potential observed in the second half compared with the first half of a 15-min sub-maximal intermittent isometric knee-extension exercise protocol (ie, as a TOT effect in a physical task).60 In the herein presented study, we can at least suggest that TOT effects themselves do not depend directly on the adenosinergic mechanisms. It would be interesting in future studies to look at potential benefit of higher doses of caffeine (> 3 mg per kg of body mass) to counteract cognitive deficits of totally sleep-deprived subjects related to TOT. With respect to sustained attention, sleep pressure (ie, TSD) may thus be considered as a threshold modulator of TOT mechanisms. Longer duration tasks (60 min) are generally used to reveal a cognitive fatigue when subjects are not submitted to TSD.61 In our conditions, a task of 10 minutes is sufficient to elicit TOT effect after TSD, suggesting that sleep pressure may trigger TOT earlier by an additive modulation of an input-output linear relationship (for instance, the number of lapses per min as a function of time).
Some limitations in the present study must be taken into consideration when interpreting our results. Indeed, the sample size is small and may have masked subtle results on the effects of caffeine on cognitive performance related to the TOT effect for example. In addition, there is variability in daily caffeine consumption that cannot be accounted for with the creation of subgroups given the sample size.
This study demonstrated that cognitive deficits (sustained attention and executive processes) related to TSD are under the influence not only of the lack of sleep itself (TSD) but also of Time-On-Task (TOT). A low dose of caffeine (2.5 mg/kg) is only beneficial for sustained attention deficits related to TSD per-se, by slowing down the trigger due to the TOT factor. Such dose of caffeine was neither efficient to counteract sustained attention deficits related to TOT nor executive processes deficits that seem mainly affected by TOT and thus less dependent to the wakefulness instability state due to an increase of the homeostatic sleep pressure. Finally, the increase of global EEG theta power during TSD and increase in three brain regions during TOT highlighted the implication of brain theta rhythm in revealing a mental/cognitive fatigue state.
Conclusion
Our results demonstrated, to our knowledge for the first time, that cognitive deficits related to a continuous wakefulness of 26 hours are not only due to a lack of sleep itself but also to a time on task (TOT) effect putting forward the idea 1) to look more carefully at the commitment time of a subject under sleep debt that is engaged in a cognitive task and 2) to question for a common psychophysiological substrate of a mental/cognitive fatigue state. Our results also pointed out a differential benefit of a low dose of caffeine (175 mg) that only slows down the triggering of sustained attention deficits related to TOT. They reinforce the idea that caffeine effectiveness in counteracting sleepiness and sustained attention deficits related to TSD would rely on the adenosinergic mechanism whereas TOT effect observed for both sustained attention and executive would not.
Abbreviations
BMI, body mass index; CAF, caffeine; EFs, executive functions; KSS, Karolinska sleepiness scale; PCBO, placebo; PSQI, Pittsburgh Sleep Quality Index; PVT, psychomotor vigilance test; ROI, region of interest; TSD, total sleep deprivation; TOT, time-on-task.
Clinical Trials Details
Clinical Trials Brief Title: Protocol PERCAF 2018. Clinical Trials Number: NCT03859882. Clinical Trials, First submission date: 25/02/2019.
Statement of Significance
It is widely admitted that total sleep deprivation (TSD) is responsible for a large range of cognitive deficits in healthy adults. However, the contribution of sleep debt per se, time on task and benefit of caffeine on cognitive deficits have received little investigation. Here we showed that the contribution of sleep debt per se, time on task and caffeine were not equivalent depending on the cognitive process engaged (ie, sustained attention versus executive processes). These results suggest that cognitive fatigue 1) would be differently supplied by sleep debt and time on task and 2) would not rely on the same neurophysiological changes according to the process involved. They would also open discussion upon concepts of cognitive fatigue and fatigability.
Data Sharing Statement
The authors do not intend to share individual data from de-identified participants, and no study documents will be made available.
Acknowledgments
The authors thank Philippe Colin, Benoît Lepetit, Cyprien Bourrilhon, Pierre Fabriès, Rodolphe Dorey and Bruno Schmid for their technical and logistic contribution to this work.
Funding
Financial supports were provided by the French General Directorate for Armament (Direction Générale de l’Armement), Department of Defense, Contract N°: PDH-1-SMO-2-509/SAN-1-509.
Disclosure
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest. Damien Léger reports grants from Sanofi, grants from Idorsia, grants from Jazz, grants from Bioserenity, outside the submitted work. The authors report no other potential conflicts of interest for this work.
References
1. Hockey R. The Psychology of Fatigue: Work, Effort and Control. Cambridge. UK: Cambridge University Press; 2013.
2. Caldwell JA, Caldwell JL, Thompson LA, Lieberman HR. Fatigue and its management in the workplace. Neurosci Biobehav Rev. 2019;96:272–289. doi:10.1016/j.neubiorev.2018.10.024
3. Boksem MA, Tops M. Mental fatigue: costs and benefits. Brain Res Rev. 2008;59(1):125–139. doi:10.1016/j.brainresrev.2008.07.001
4. Baumeister RF, Bratslavsky E, Muraven M, Tice DM. Ego depletion: is the active self a limited resource? J Pers Soc Psychol. 1998;74(5):1252–1265. doi:10.1037/0022-3514.74.5.1252
5. Pattyn N, Neyt X, Henderickx D, Soetens E. Psychophysiological investigation of vigilance decrement: boredom or cognitive fatigue? Physiol Behav. 2008;93(1–2):369–378. doi:10.1016/j.physbeh.2007.09.016
6. Matthews G. Personality and individual differences in cognitive fatigue. In: Association AP, editor. Decade of Behavior/Science Conference. Cognitive Fatigue: Multidisciplinary Perspectives on Current Research and Future Applications. Washington, DC, US: American Psychological Association; 2011:209–227.
7. Borbely AA, Daan S, Wirz-Justice A, Deboer T. The two-process model of sleep regulation: a reappraisal. J Sleep Res. 2016;25(2):131–143. doi:10.1111/jsr.12371
8. Krause AJ, Simon EB, Mander BA, et al. The sleep-deprived human brain. Nat Rev Neurosci. 2017;18(7):404–418.
9. Arnal PJ, Sauvet F, Leger D, et al. Benefits of sleep extension on sustained attention and sleep pressure before and during total sleep deprivation and recovery. Sleep. 2015;38(12):1935–1943. doi:10.5665/sleep.5244
10. Rabat A, Arnal PJ, Monnard H, et al. Limited benefit of sleep extension on cognitive deficits during total sleep deprivation: illustration with two executive processes. Front Neurosci. 2019;13:591. doi:10.3389/fnins.2019.00591
11. Lim J, Ebstein R, Tse CY, et al. Dopaminergic polymorphisms associated with time-on-task declines and fatigue in the psychomotor vigilance test. PLoS One. 2012;7(3):e33767. doi:10.1371/journal.pone.0033767
12. Gui D, Xu S, Zhu S, et al. Resting spontaneous activity in the default mode network predicts performance decline during prolonged attention workload. Neuroimage. 2015;120:323–330. doi:10.1016/j.neuroimage.2015.07.030
13. Phipps-Nelson J, Redman JR, Rajaratnam SM. Temporal profile of prolonged, night-time driving performance: breaks from driving temporarily reduce time-on-task fatigue but not sleepiness. J Sleep Res. 2011;20(3):404–415. doi:10.1111/j.1365-2869.2010.00900.x
14. Barwick F, Arnett P, Slobounov S. EEG correlates of fatigue during administration of a neuropsychological test battery. Clin Neurophysiol. 2012;123(2):278–284. doi:10.1016/j.clinph.2011.06.027
15. Gorgoni M, Ferlazzo F, Ferrara M, et al. Topographic electroencephalogram changes associated with psychomotor vigilance task performance after sleep deprivation. Sleep Med. 2014;15(9):1132–1139. doi:10.1016/j.sleep.2014.04.022
16. Wascher E, Rasch B, Sanger J, et al. Frontal theta activity reflects distinct aspects of mental fatigue. Biol Psychol. 2014;96:57–65. doi:10.1016/j.biopsycho.2013.11.010
17. Asplund CL, Chee MWL. Time-on-task and sleep deprivation effects are evidenced in overlapping brain areas. Neuroimage. 2013;82:326–335. doi:10.1016/j.neuroimage.2013.05.119
18. Veksler BZ, Gunzelmann G. Functional equivalence of sleep loss and time on task effects in sustained attention. Cogn Sci. 2018;42(2):600–632. doi:10.1111/cogs.12489
19. Xu J, Zhu Y, Fu C, et al. Frontal metabolic activity contributes to individual differences in vulnerability toward total sleep deprivation-induced changes in cognitive function. J Sleep Res. 2015;25(2):169–180. doi:10.1111/jsr.12354
20. Yeo BT, Tandi J, Chee MW. Functional connectivity during rested wakefulness predicts vulnerability to sleep deprivation. Neuroimage. 2015;111:147–158. doi:10.1016/j.neuroimage.2015.02.018
21. Lei Y, Shao Y, Wang L, et al. Large-scale brain network coupling predicts total sleep deprivation effects on cognitive capacity. PLoS One. 2015;10(7):e0133959. doi:10.1371/journal.pone.0133959
22. Thomas M, Sing H, Belenky G, et al. Neural basis of alertness and cognitive performance impairments during sleepiness. I. Effects of 24 h of sleep deprivation on waking human regional brain activity. J Sleep Res. 2000;9(4):335–352. doi:10.1046/j.1365-2869.2000.00225.x
23. Martin K, Thompson KG, Keegan R, Ball N, Rattray B. Mental fatigue does not affect maximal anaerobic exercise performance. Eur J Appl Physiol. 2015;115(4):715–725. doi:10.1007/s00421-014-3052-1
24. Porkka-Heiskanen T, Strecker RE, Thakkar M, Bjorkum AA, Greene RW, McCarley RW. Adenosine: a mediator of the sleep-inducing effects of prolonged wakefulness. Science. 1997;276(5316):1265–1268. doi:10.1126/science.276.5316.1265
25. Urry E, Landolt HP. Adenosine, caffeine, and performance: from cognitive neuroscience of sleep to sleep pharmacogenetics. Curr Top Behav Neurosci. 2015;25:331–366.
26. Killgore WDS, Kamimori GH. Multiple caffeine doses maintain vigilance, attention, complex motor sequence expression, and manual dexterity during 77 hours of total sleep deprivation. Neurobiol Sleep Circadian Rhythms. 2020;9:100051. doi:10.1016/j.nbscr.2020.100051
27. Aidman E, Balin M, Johnson K, et al. Caffeine may disrupt the impact of real-time drowsiness on cognitive performance: a double-blind, placebo-controlled small-sample study. Sci Rep. 2021;11(1):4027. doi:10.1038/s41598-021-83504-6
28. Erblang M, Sauvet F, Drogou C, et al. Genetic determinants of neurobehavioral responses to caffeine administration during sleep deprivation: a randomized, cross over study (NCT03859882). Genes. 2021;12:555. doi:10.3390/genes12040555
29. Åkerstedt T, Gillberg M. Subjective and objective sleepiness in the active individual. Int J Neurosci. 1990;52:29–37. doi:10.3109/00207459008994241
30. Drummond SP, Bischoff-Grethe A, Dinges DF, Ayalon L, Mednick SC, Meloy MJ. The neural basis of the psychomotor vigilance task. Sleep. 2005;28(9):1059–1068.
31. Bougard C, VanBeers P, Sauvet F, et al. Motorcycling performance and sleepiness during an extended ride on a dynamic simulator: relationship with stress biomarkers. Physiol Meas. 2020;41(10):104004. doi:10.1088/1361-6579/abb75e
32. McLellan TM, Caldwell JA, Lieberman HR. A review of caffeine’s effects on cognitive, physical and occupational performance. Neurosci Biobehav Rev. 2016;71:294–312. doi:10.1016/j.neubiorev.2016.09.001
33. Oostenveld R, Fries P, Maris E, Schoffelen JM. FieldTrip: open source software for advanced analysis of MEG, EEG, and invasive electrophysiological data. Comput Intell Neurosci. 2011;2011:156869. doi:10.1155/2011/156869
34. Sauvet F, Erblang M, Gomez-Merino D, et al. Efficacy of THN102 (a combination of modafinil and flecainide) on vigilance and cognition during 40-hour total sleep deprivation in healthy subjects: glial connexins as a therapeutic target. Br J Clin Pharmacol. 2019;85(11):2623–2633. doi:10.1111/bcp.14098
35. Lythe KE, Williams SC, Anderson C, Libri V, Mehta MA. Frontal and parietal activity after sleep deprivation is dependent on task difficulty and can be predicted by the fMRI response after normal sleep. Behav Brain Res. 2012;233(1):62–70. doi:10.1016/j.bbr.2012.04.050
36. Satterfield BC, Wisor JP, Schmidt MA, Van Dongen HPA. Time-on-task effect during sleep deprivation in healthy young adults is modulated by dopamine transporter genotype. Sleep. 2017;40(12). doi:10.1093/sleep/zsx167
37. Maire M, Reichert CF, Gabel V, et al. Time-on-task decrement in vigilance is modulated by inter-individual vulnerability to homeostatic sleep pressure manipulation. Front Behav Neurosci. 2014;8:59. doi:10.3389/fnbeh.2014.00059
38. Lo JC, Groeger JA, Santhi N, et al. Effects of partial and acute total sleep deprivation on performance across cognitive domains, individuals and circadian phase. PLoS One. 2012;7(9):e45987. doi:10.1371/journal.pone.0045987
39. Slama H, Chylinski DO, Deliens G, Leproult R, Schmitz R, Peigneux P. Sleep deprivation triggers cognitive control impairments in task-goal switching. Sleep. 2017;41(2):zsx200.
40. Frenda SJ, Fenn KM. Sleep less, think worse: the effect of sleep deprivation on working memory. J Appl Res Mem Cogn. 2016;5:463–469. doi:10.1016/j.jarmac.2016.10.001
41. Sauvet F, Arnal PJ, Tardo-Dino PE, et al. Beneficial effects of exercise training on cognitive performances during total sleep deprivation in healthy subjects. Sleep Med. 2020;65:26–35. doi:10.1016/j.sleep.2019.07.007
42. Killgore WD, Kahn-Greene ET, Grugle NL, Killgore DB, Balkin TJ. Sustaining executive functions during sleep deprivation: a comparison of caffeine, dextroamphetamine, and modafinil. Sleep. 2009;32(2):205–216. doi:10.1093/sleep/32.2.205
43. Wesensten NJ, Killgore WD, Balkin TJ. Performance and alertness effects of caffeine, dextroamphetamine, and modafinil during sleep deprivation. J Sleep Res. 2005;14(3):255–266. doi:10.1111/j.1365-2869.2005.00468.x
44. Wyatt JK, Cajochen C, Ritz-de Cecco A, Czeisler CA, Dijk DJ. Low-dose repeated caffeine administration for circadian-phase-dependent performance degradation during extended wakefulness. Sleep. 2004;27(3):374–381. doi:10.1093/sleep/27.3.374
45. Ward AM, McLaren DG, Schultz AP, et al. Daytime sleepiness is associated with decreased default mode network connectivity in both young and cognitively intact elderly subjects. Sleep. 2013;36(11):1609–1615. doi:10.5665/sleep.3108
46. Stawarczyk D, D’Argembeau A. Conjoint influence of mind-wandering and sleepiness on task performance. J Exp Psychol Hum Percept Perform. 2016;42(10):1587–1600. doi:10.1037/xhp0000254
47. Anticevic A, Cole MW, Murray JD, Corlett PR, Wang XJ, Krystal JH. The role of default network deactivation in cognition and disease. Trends Cogn Sci. 2012;16(12):584–592. doi:10.1016/j.tics.2012.10.008
48. Andrews-Hanna JR. The brain’s default network and its adaptive role in internal mentation. Neuroscientist. 2012;18(3):251–270. doi:10.1177/1073858411403316
49. Menon V. Large-scale brain networks and psychopathology: a unifying triple network model. Trends Cogn Sci. 2011;15(10):483–506. doi:10.1016/j.tics.2011.08.003
50. Xu H, Shen H, Wang L, et al. Impact of 36h of total sleep deprivation on resting-state dynamic functional connectivity. Brain Res. 2018;1688:22–32. doi:10.1016/j.brainres.2017.11.011
51. Cajochen C, Wyatt JK, Czeisler CA, Dijk DJ. Separation of circadian and wake duration-dependent modulation of EEG activation during wakefulness. Neuroscience. 2002;114(4):1047–1060. doi:10.1016/S0306-4522(02)00209-9
52. van Son D, de Rover M, De Blasio FM, van der Does W, Barry RJ, Putman P. Electroencephalography theta/beta ratio covaries with mind wandering and functional connectivity in the executive control network. Ann N Y Acad Sci. 2019;1452(1):52–64. doi:10.1111/nyas.14180
53. Chang D, Song D, Zhang J, Shang Y, Ge Q, Wang Z. Caffeine caused a widespread increase of resting brain entropy. Sci Rep. 2018;8(1):2700. doi:10.1038/s41598-018-21008-6
54. Retey JV, Adam M, Gottselig JM, et al. Adenosinergic mechanisms contribute to individual differences in sleep deprivation-induced changes in neurobehavioral function and brain rhythmic activity. J Neurosci. 2006;26(41):10472–10479. doi:10.1523/JNEUROSCI.1538-06.2006
55. Borragan G, Slama H, Bartolomei M, Peigneux P. Cognitive fatigue: a time-based resource-sharing account. Cortex. 2017;89:71–84. doi:10.1016/j.cortex.2017.01.023
56. Kato Y, Endo H, Kizuka T. Mental fatigue and impaired response processes: event-related brain potentials in a Go/NoGo task. Int J Psychophysiol. 2009;72(2):204–211. doi:10.1016/j.ijpsycho.2008.12.008
57. van der Linden D, Eling P. Mental fatigue disturbs local processing more than global processing. Psychol Res. 2006;70(5):395–402. doi:10.1007/s00426-005-0228-7
58. Drummond SP, Brown GG, Salamat JS, Gillin JC. Increasing task difficulty facilitates the cerebral compensatory response to total sleep deprivation. Sleep. 2004;27(3):445–451.
59. Drummond SP, Meloy MJ, Yanagi MA, Orff HJ, Brown GG. Compensatory recruitment after sleep deprivation and the relationship with performance. Psychiatry Res. 2005;140(3):211–223. doi:10.1016/j.pscychresns.2005.06.007
60. de Morree HM, Klein C, Marcora SM. Cortical substrates of the effects of caffeine and time-on-task on perception of effort. J Appl Physiol. 2014;117(12):1514–1523. doi:10.1152/japplphysiol.00898.2013
61. Luque-Casado A, Perakakis P, Ciria LF, Sanabria D. Transient autonomic responses during sustained attention in high and low fit young adults. Sci Rep. 2016;6:27556. doi:10.1038/srep27556
 © 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.