Back to Journals » Diabetes, Metabolic Syndrome and Obesity » Volume 12
Dysregulation of DPP4 Is Associated with the AMPK/JAK2/STAT3 Pathway in Adipocytes Under Insulin Resistance Status and Liraglutide Intervention
Authors Cheng F, Yuan G, He J, Shao Y, Zhang J, Guo X
Received 4 September 2019
Accepted for publication 2 December 2019
Published 11 December 2019 Volume 2019:12 Pages 2635—2644
DOI https://doi.org/10.2147/DMSO.S229838
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. Juei-Tang Cheng
Fangxiao Cheng,1 Geheng Yuan,1 Jiao He,2 Yimin Shao,1 Junqing Zhang,1 Xiaohui Guo1
1Department of Endocrinology, Peking University First Hospital, Beijing 100034, People’s Republic of China; 2Department of Endocrinology, Baoding First Central Hospital, Baoding 071000, Hebei Province, People’s Republic of China
Correspondence: Geheng Yuan; Xiaohui Guo
Department of Endocrinology, Peking University First Hospital, No. 8 Xishiku Street, Xicheng District, Beijing 100034, People’s Republic of China
Tel|Fax +86 010 83575103
Email [email protected]; [email protected]
Purpose: Dipeptidyl peptidase 4 (DPP4) is one of the newly identified adipokines, which acts as paracrine in adipose tissue and as endocrine hormones in the liver, muscles and central nervous system. Expression of DPP4 was significantly upregulated in obese patients upon insulin resistance (IR) conditions, but the mechanism underlying the dysregulation of DPP4 remains unclear. This study aimed to investigate the DPP4 expression in adipose tissue and adipocytes under IR conditions or with liraglutide intervention, and explore the potential molecular mechanisms.
Methods: Obesity-associated IR animal and cell models were, respectively, constructed by using high-fat diet and palmitic acid (PA) stimulation. Expression of DPP4 in adipose tissues and adipocytes was estimated by quantitative real-time RT-PCR and Western-blot. Effects of the AMPK/JAK2/STAT3 pathway on DPP4 were examined by regulating the activity of AMPK and the JAK2/STAT signaling. The therapeutic efficacy of liraglutide in the IR models was evaluated, and its regulatory effects on DPP4 expression and the underlying molecular mechanisms were explored.
Results: The expression of DPP4 was markedly upregulated in both the animal and cell IR models. In the adipocyte, DPP4 expression was found to be suppressed by the activation of AMPK, and this inhibition effect was mediated by the JAK2/STAT3 signaling. Moreover, liraglutide could alleviate the obesity-induced IR, and led to the downregulation of DPP4 in IR animal and cell models. Liraglutide intervention resulted in the activation of AMPK and deactivation of the JAK2/STAT3 signaling in the adipocytes.
Conclusion: Taken together, the expression of DPP4 is upregulated in adipose tissues and adipocytes upon IR conditions, but is reduced after liraglutide intervention. The dysregulation of DPP4 in the adipocytes may be performed by the AMPK/JAK2/STAT3 pathway.
Keywords: obesity, adipocyte, DPP4, AMPK, JAK2/STAT3, liraglutide, insulin resistance
Introduction
Obesity epidemic becomes a major global health burden due to the related comorbidities, such as hyperglycemia, diabetes mellitus and insulin resistance (IR).1 Obese individuals, particularly those with excess abdominal adipose tissue, have an elevated risk of developing Type 2 diabetes mellitus (T2DM), cardiovascular disease, and hypertension.2 During obesity, the production of inflammatory cytokines and reactive oxygen species within adipose tissue increases as well as ectopic lipid deposition in liver or skeletal muscle.3,4 These consequences reflect potential causative links between adipose tissue dysfunction and IR.
Emerging studies indicate that adipose tissue is not only an energy storage organ but also an important endocrine organ.5 Adipokines are a group of proteins and peptides released by adipose tissue cells, which contribute to the interconnection with other organs through the complex signaling pathways.6 Obesity with the enlargement of adipose tissues results in dysregulation of the adipokines, leading to the disorder of glucose and lipid metabolism.7 Dipeptidyl peptidase 4 (DPP4) has been identified as a novel adipokines by Daniela in 2011 and was proved to be upregulated in obesity, especially in the cases upon IR condition.8 In T2DM, the deregulated DPP4 plays as an enhancer of IR development by regulating incretins.9,10 In obesity, inhibition of DPP4 prevented the white adipose tissue fibrosis by paracrine effect.11 Although a recent research reported that there is no significant effect of adipose cell-derived DPP4 on obesity-associated inflammation and glucose homeostasis, the obviously increased DPP4 in obesity is an established fact.12 Adipose tissue and specifically adipocytes have been proposed to be a significant circulating source of DPP4.8,13 DPP4 secretion from adipose tissue was also demonstrated in vivo with greater release in obese compared to lean individuals.14 DPP4 release from visceral adipocytes was also higher in obese patients.15 More importantly, obesity with IR patients had markedly higher circulating DPP4 than the matched insulin-sensitive obese patients.14 However, the understanding of molecular mechanisms underlying the dysregulation of DPP4 in obesity remains limited.
It is generally considered that the mechanisms that regulate DPP4 may be variant in different cell types. Studies show that many STAT activators play a critical role in the regulation of adipocyte gene expression and exhibit differential expression in conditions of obesity and/or IR.16,17 In chronic b lymphocytic leukemia cells, interferons could stimulate STAT1 to bind to a consensus interferon-γ-activated sequence (GAS), and thereby led to the upregulation of DPP4.18 STAT3 is abundantly expressed in adipocytes19,20 and activated early in the adipogenic program and mediates the action of numerous cytokines in adipocytes.21,22 Notably, the association of the JAK2/STAT3 signaling with DPP4 has been demonstrated in other cells.23,24 The activity of JAK2/STAT3 signaling could be suppressed by AMP-activated protein kinase (AMPK) in adipose tissues and adipocytes, leading to the inhibition of adipogenic processes.22 Additionally, the AMPK signaling has been reported to mediate the function of DPP4 inhibitor in some cells, such as HepG2 cell25 and neuronal cell.26 However, the association of DPP4 with the JAK2/STAT3 signaling pathway and AMPK has been rarely reported in obesity. Liraglutide is a kind of incretin-based drug, which has been widely used in the treatment of T2DM with obesity.27 Cao et al indicated that liraglutide administration in T2DM resulted in abnormal DPP4 expression.28 Our previous studies have reported that AMPK was up-regulated in adipocyte with liraglutide in db/db mice.29 Thus, we suspected that the treatment of liraglutide might also regulate the expression of DPP4 via regulation of AMPK in obesity.
To confirm the expression patterns of DPP4 in obesity upon IR status or received liraglutide treatment, and explore the molecular mechanisms underlying the regulatory effects, we constructed obesity-induced IR models using rats and adipocytes, and assessed the expression of DPP4 in adipose tissues and cells, as well as its relationship with the AMPK/JAK2/STAT3 pathway.
Materials and Methods
Experimental Animals and Treatments
A total of 26 Sprague-Dawley (SD) rats (males, 6 weeks) were purchased from Beijing Vital River Laboratory Animal Technology Company (Beijing, China). All the rats were kept in the departmental animal house under a 12 hrs light-dark cycle and free access to water and food conditions, with humidity of 50–60% and temperature of 22–25°C. The rats were used for experiments after a one-week adaptation to the housing facilities. The protocols for animal experiments were performed following the Principles of Laboratory Animal Care by the National Institute of Health and approved by the Animal Ethics Committee of Peking University First Hospital.
The rats were divided into three groups, including normal control (NC) group (n = 10), IR group (n = 10) and liraglutide group (n = 6). The animals in NC group were fed with regular diet (10% fat). The rats in IR group were fed with D12492 high-fat diet (HFD; 60% fat) for 12 weeks to establish IR animal models. In liraglutide group, the rats were fed with HFD for 12 weeks and subsequently received injection of liraglutide (200 μg/kg twice daily; Novo Nordisk) or normal saline (NS) for 8 weeks. During the treatment, the body weight (BW) was estimated weekly, and the fasting blood glucose (FBG) and fasting serum insulin (FINS) of the rats was measured after the feeding to evaluate the homeostasis model of assessment for IR index (HOMA-IR). The omental adipose tissues were collected from the animal models and immediately frozen in liquid nitrogen for further experiments.
Cell Culture and Treatments
Mouse 3T3-L1 pre-adipocytes were obtained from American Type Culture Collection (ATCC, Manassas, USA) and cultured in Dulbecco’s modified Eagle’s medium (DMEM; Gibco, CA, USA) supplemented with 10% fetal bovine serum (FBS) in a humidified atmosphere of 5% CO2 a 37°C.
After the cells spread to all the bottom of the culture plates, they were incubated in induction medium I, containing 10 μg/mL of insulin, 0.5 mM 1-methyl-3-isobutyl-xanthine and 1 μg/mL of dexamethasone in DMEM supplemented with 10% FBS for 24 hrs. Then, the cells were cultured in fresh DMEM with 10% FBS for other 24 hrs. Thereafter, the cells were placed in induction medium II, including 10μg/mL of insulin in DMEM with 10% FBS for 48 hrs. Lastly, the cells were incubated in DMEM supplemented with 10% FBS, and the medium was renewed every 2 days until about 95% of the cells differentiated into adipocytes. The differentiated 3T3-L1 adipocytes were divided into three groups: normal control (NC) group, palmitic acid (PA) group and liraglutide treated cell group. Cells in PA group were treated with 1 mM of PA for 24 hrs to establish IR cell model. In the liraglutide-treated cell group, the cells were firstly incubated with PA to establish IR model and then received 50 nM liraglutide for 24 h. The JAK2/STAT3 signaling pathway activity was regulated by STAT3 activator Colivelin (COL; 1 μM, 3 hrs) or inhibitor cryptotanshinone (CPT; 5μM, 24 hrs). AMPK activity was modulated by its activator metformin (MET; 2 mM, 5 days).
Oral Glucose Tolerance Test (OGTT) and Intraperitoneal Insulin Tolerance Test (IPITT)
OGTT was carried out before and after the drug treatments. The animals free accessed to water and were fasted for 12 hrs, and then treated with 40% of glucose (2 g/kg BW) via oral injection. A glucose meter (ACCU-CHEK Performa Roche, Germany) was used to examine the blood glucose at the tip of the tail at 0, 30, 60, 90, and 120 mins after the administration of glucose. The area under the curve (AUC) was calculated to evaluate the sensitivity of islet β-cells.
We used IPITT to evaluate the insulin sensitivity before and after the drug treatments. The rats free accessed to water and were fasted for 12 hrs, and then treated with insulin (0.75 IU/kg BW) through intraperitoneal injection. A glucose meter (ACCU-CHEK Performa Roche, Germany) was used to examine the blood glucose at the tip of the tail at 0, 30, 60, 90, and 120 mins after the administration of glucose. The AUC value was computed to evaluate the insulin sensitivity.
Glucose Uptake Assay
In this study, the insulin-stimulated glucose uptake of the 3T3-L1 cells was examined using a glucose colorimetric/fluorometric assay kit (Biovision, Inc., Milpitas, CA, USA). Briefly, the cells were incubated with or without insulin (100 nM) at 37°C for 20 mins. Then, the glucose assay buffer was added into the cells and incubated at 37°C for 10 mins. The absorbance at 570 nm was measured by a microplate reader (BioTek, Beijing, China).
RNA Extraction and Quantitative Real-Time RT-PCR (qRT-PCR)
Total RNA in the tissues and cells was extracted using Trizol reagent (Invitrogen, Carlsbad, CA, USA). Reverse transcription was performed to synthesize cDNA with the obtained RNA using a PrimeScript RT reagent kit (Takara, Shiga, Japan). Quantitative real-time PCR (qPCR) was carried out using SYBR green I Master Mix kit (Invitrogen, Carlsbad, CA, USA) and ABI Prism 7300 system (Thermo Fisher Scientific, Waltham, MA, USA). The GAPDH was used as an endogenous control for the normalization of mRNA, and the final expression value was calculated using 2−ΔΔCt method.
Western Blot Assay
Total protein was extracted from the tissues and cells by RIPA buffer and then separated using 12% SDS polyacrylamide gel electrophoresis. The proteins were transferred to the PVDF membrane (Amersham, Little Chalfont, UK). After the blocking with 3% nonfat milk, the membrane was incubated with first antibodies against DPP4, JAK2, p-JAK2, p-STAT3, and STAT3 at 4°C overnight, and then was incubated with the second antibodies at room temperature for 1 h. The final immunoreactivity was measured using an enhanced chemiluminescence detection system (Amersham, Little Chalfont, UK). The protein bands were quantified with an Image-Pro Plus 6.0 software (Media Cybernetics, Inc., Rockville, MD, USA).
Statistical Analysis
All the statistical analyses were carried out using SPSS 21.0 software (SPSS Inc., Chicago, IL) and GraphPad Prism 5.0 software (GraphPad Software, Inc., USA). Data were expressed as mean ± standard deviation (SD). Differences between groups were compared using Student’s t test or one-way ANOVA analysis followed by Turkey’s post hoc test. A value of P< 0.05 was considered statistically significant.
Results
Establishment of Obese Animal Model and Adipocyte Model with IR
This study, respectively, used HFD and PA treatment to establish the obesity-associated IR animal and cell models. As expected, the BW of the animals was increased in the IR group as compared with the controls (P < 0.05, Figure S1A). To verify that the obesity-induced IR model was established, OGTT and IPITT were carried out. As shown in Figures S1B and 1C, the glucose tolerance and insulin tolerance were significantly impaired in the IR group compared with the NC group, and the AUCs of IR group for OGTT and IPITT were all increased compared with that in NC group (all P < 0.05, Figure S1D). In addition, the IR animals had higher HOMA-IR than the normal controls (P < 0.05, Figure S1E). For the IR cell model, we estimated the ability of glucose uptake. From Figure S1F, we found that the insulin-stimulated glucose uptake was suppressed by PA treatment in the 3T3-L1 cells. These data indicated that the obesity-induced IR models were successfully constructed.
Expression of DPP4 Under IR Conditions
Upregulated expression of DPP4 has been reported in obese patients with IR compared with those insulin sensitive individuals. In the present study, the expression of DPP4 was measured in the adipose tissue and adipocyte. As shown in Figure 1A and B, both the mRNA and protein expression levels of DPP4 were increased in the IR animals compared with the normal controls (all P < 0.05). Similarly, the upregulated expression of DPP4 was also observed in the adipocyte with IR as compared to the normal controls (all P < 0.05, Figure 1C and D).
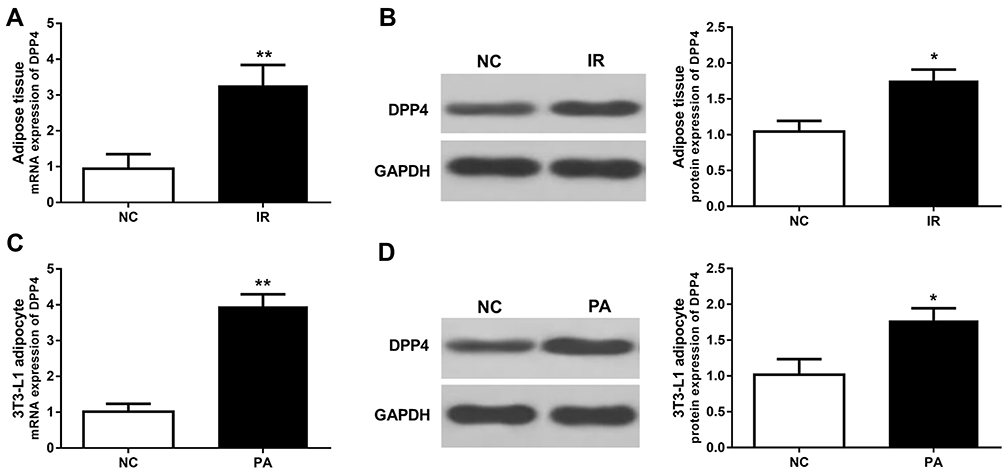 |
Figure 1 Expression of DPP4 in adipose tissues and adipocytes under IR conditions. (A, B) The mRNA and protein expression levels of DPP4 were increased in the IR rats compared with the normal controls. (C, D) The mRNA and protein expression levels of DPP4 were increased in the IR cells compared with the controls. *P < 0.05, **P < 0.01. |
DPP4 Expression Was Regulated by the JAK2/STAT3 Signaling Pathway in Adipocytes
The activity of JAK2/STAT3 signaling pathway was investigated due to its important role in the adipogenesis. According to the Western blot results, we found that the JAK2/STAT3 signaling pathway was activated in the IR cell models, which evidenced by the increased ratio of p-JAK2/JAK2 and p-STAT3/STAT3 (all P < 0.05, Figure 2A and B). Furthermore, the effect of the JAK2/STAT3 signaling pathway on DPP4 was evaluated by using the inhibitor CPT and the activator COL. The expression of p-STAT3 was significantly decreased by CPT, while it was upregulated by COL in the IR cells (P < 0.01, Figure 2B), indicating that the activity of JAK2/STAT3 signaling pathway was blocked by CPT, but was promoted by COL. As shown in Figure 2C, the deactivation of JAK2/STAT3 signaling pathway led to inhibited expression of DPP4, whereas the activation of JAK2/STAT3 signaling pathway promoted the expression of DPP4 in adipocyte (all P < 0.05).
 |
Figure 2 Effect of the AMPK/JAK2/STAT3 pathway on the expression of DPP4 in adipocytes. (A) Western blot results for DPP4 and proteins in the JAK2/STAT3 signaling pathway. (B) Activity of the JAK2/STAT3 signaling pathway in adipocytes under IR conditions, and its activity was inhibited by CPT and was promoted by COL. (C) DPP4 expression was suppressed by deactivation of the JAK2/STAT3 signaling pathway, and was enhanced by activation of the JAK2/STAT3 signaling pathway. (D) Western blot results for DPP4 and proteins in the AMPK/JAK2/STAT3 pathway. (E) AMPK was deactivated in adipocytes under IR status, and the activation of AMPK led to the suppression of the JAK2/STAT3 signaling pathway. (F) The decreased expression of DPP4 induced by activation of AMPK was rescued by the activation of the JAK2/STAT3 signaling pathway. *P < 0.05 compared with the NC group; #P < 0.05 compared with the PA group; &P < 0.05 compared with the MET group. |
AMPK Inhibits the Activity of JAK2/STAT3 Signaling and the Expression of DPP4 in Adipocytes
This study also focused on the expression of AMPK, which has been reported to be involved in adipogenesis by regulating the JAK2/STAT3 signaling. As shown in Figure 2D and E, we observed that the p-AMPK protein expression was reduced in the IR cell models (P < 0.05). By using the activator MET, p-AMPK expression was significantly upregulated (P < 0.05), and the activity of JAK2/STAT3 signaling pathway was expected inhibited in the adipocyte, which is evidenced by the decreased p-JAK2 and p-STAT3 (all P < 0.05, Figure 2E). More importantly, the expression of DPP4 was also downregulated by the activation of AMPK in the adipocyte (P < 0.05, Figure 2F).
Activation of the JAK2/STAT3 Signaling Pathway Rescues the Effect of AMPK on the Expression of DPP4
To confirm whether the effect of AMPK on the expression of DPP4 was mediated by the JAK2/STAT3 signaling pathway, this study used the JAK2/STAT3 signaling pathway activator COL in the adipocyte pre-treated with MET. As shown in Figure 2D and E, the inhibition of JAK2/STAT3 signaling pathway induced by AMPK was abrogated after the treatment of COL (all P < 0.05). Notably, the AMPK-induced DPP4 reduction in the adipocyte was markedly rescued by the activation of JAK2/STAT3 signaling pathway (P < 0.05, Figure 2F), indicating that the JAK2/STAT3 signaling pathway mediated the suppressive effect of AMPK on the expression of DPP4 in adipocyte.
Liraglutide Improves the Glucose Homeostasis and Insulin Sensitivity in Obesity-Associated IR
By the treatment with liraglutide, the glucose tolerance and insulin sensitivity of the IR animals were all significantly improved evidenced by the reduced AUCs for OGRR and IPITT compared with the animals in IR group (all P< 0.05, Figure 3A–C). Moreover, the HOMA-IR value in the IR animals was also decreased by the treatment of liraglutide (P< 0.05, Figure 3D). In the IR cell model, the decreased glucose uptake induced by PA was abrogated by the treatment of liraglutide (P< 0.05, Figure 3E), which suggested that liraglutide could improve the insulin-stimulated glucose uptake under IR conditions.
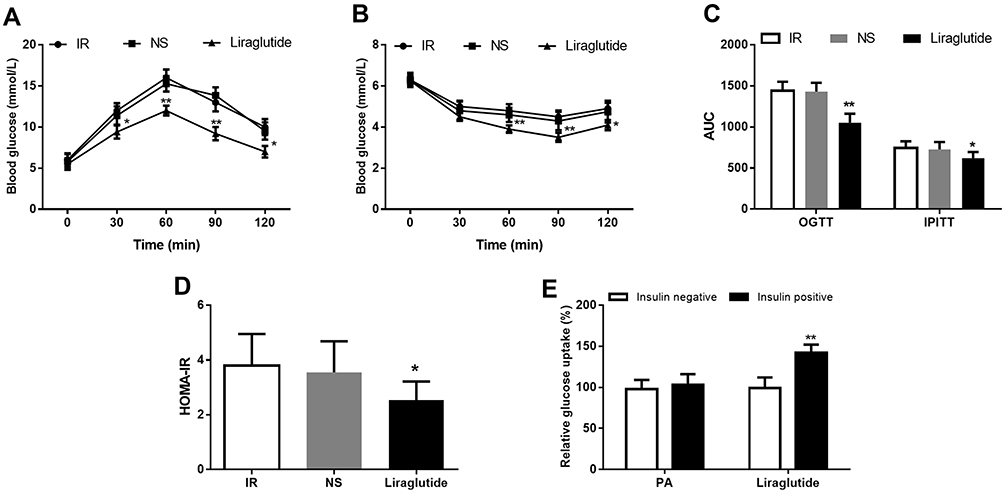 |
Figure 3 Therapeutic efficacy of liraglutide against obesity-associated IR. (A) The impaired glucose tolerance was significantly improved by liraglutide. (B) The decreased insulin sensitivity in IR animals was increased following the treatment of liraglutide. (C) AUCs for OGTT and IPITT significantly decreased after liraglutide treatment. (D) The increased HOMA-IR in IR animals was reduced by liraglutide. (E) The insulin-stimulated glucose uptake ability was improved by the treatment of liraglutide. *P < 0.05, **P < 0.01. |
Liraglutide Inhibits the Expression of DPP4 in IR Animal and Cell Models
The regulatory role of liraglutide in the expression of DPP4 has been previously reported in T2DM patients.1 In the current study, we found that the increased DPP4 in the IR animal and cell models was significantly downregulated by the treatment of liraglutide (all P < 0.05, Figure 4).
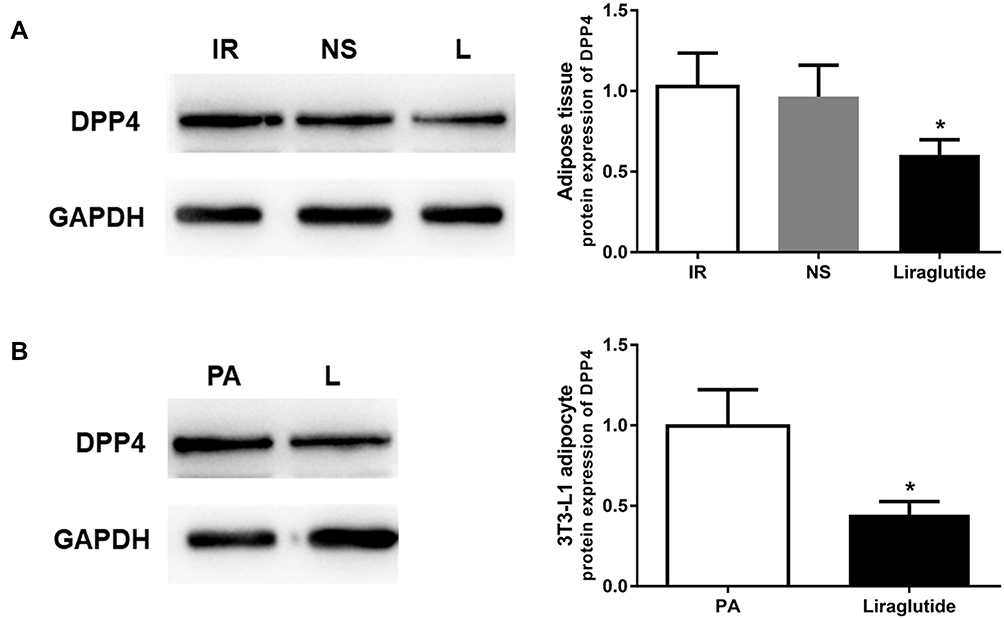 |
Figure 4 Liraglutide treatment reduced the expression of DPP4 in IR animal and cell models. (A) DPP4 expression was decreased by liraglutide in adipose tissues under IR conditions. (B) Liraglutide intervention in adipocytes led to decreased DPP4 expression. *P < 0.05. |
Liraglutide Regulates the Activity of AMPK and the JAK2/STAT3 Signaling Pathway in Adipocytes
The effects of liraglutide on AMPK and the JAK2/STAT3 signaling were also investigated. In the adipocyte under IR conditions, we found that liraglutide could lead to enhanced expression of p-AMPK, while suppressed the expression of p-JAK2 and p-STAT3 (all P < 0.05, Figure 5), suggesting that liraglutide in IR cells resulted in activation of AMPK and inhibition of the JAK2/STAT3 signaling pathway.
 |
Figure 5 Liraglutide regulated the activity of AMPK and the JAK2/STAT3 signaling pathway. (A) Western blot results for activity of AMPK and the JAK2/STAT3 signaling pathway. (B) Liraglutide intervention in adipocytes resulted in the activation of AMPK and deactivation of the JAK2/STAT3 signaling pathway. *P < 0.05. |
Discussion
This study focused on the expression patterns of DPP4 in adipose tissues and adipocytes under IR, and found that DPP4 expression was elevated in the obesity-associated IR animal and cell models. More importantly, the molecular mechanisms underlying the dysregulation of DPP4 were explored, and AMPK/JAK2/STAT3 pathway was proved to exist critical regulatory role in the expression of DPP4 in adipocytes. In addition, our study also focused on the effects of liraglutide on DPP4 in the IR models, and considered that liraglutide administration in adipocytes led to decreased DPP4 expression through the AMPK/JAK2/STAT3 pathway.
Accumulated studies indicate that the expression of DPP4 is upregulated in T2DM patients with IR, and the inhibition of DPP4 represents an efficient therapeutic strategy to maintain the balance of glucose.30 As a kind of adipokine, DPP4 has been demonstrated to be closely associated with obesity progression.31 Ghorpade et al found that the increased hepatocyte-secreted DPP4 resulted in promoted adipose inflammation and IR in obese mice.32 In addition, the upregulated expression of DPP4 in obese IR patients might serve as a regulator of insulin signaling in adipocytes.33 A recent study by Varin et al also found the deregulated DPP4 expression in obesity, but no significant association was observed between the adipose tissues derived DPP4 and regulation of inflammation and glucose homeostasis.34 Ana and his colleagues revealed that inhibition of DPP4 by vildagliptin in adipose tissue and obese mice prevented the white adipose tissue fibrosis.11 Collectively, the DPP4 expression is remarkably deregulated in adipose tissues and adipocytes, and may exist paracrine effects.
In the present study, both the mRNA and protein expression levels of DPP4 were found to be upregulated in the IR animal and cell models compared with the normal controls, which was consistent with the results in the previous studies.8 Emerging studies reveal the dysregulation of STAT family members in obesity and IR, and demonstrated their regulatory role on the expression of adipocyte gene.35 A study by Wunderlich et al showed that the activation of JAK2/STAT3 signaling by increased leptin could lead to the resistance of leptin, and IL-6-induced JAK2/STAT3 in obesity resulted in impaired insulin action.12 In human adipocytes, Kim et al found that Tau-Ribose could decrease the expression of adipokines through the inhibition of STAT3 signaling pathway.36 In our study, the JAK2/STAT3 signaling pathway was found to be activated in adipocyte by PA treatment, which implied that the activity of this signaling pathway was elevated under the IR conditions. Interestingly, we found that the increased DPP4 expression in the IR cells was suppressed after blocking the JAK2/STAT3 signaling pathway, indicating that the increased expression of DPP4 in adipocytes under IR conditions might arise from the activation of JAK2/STAT3 signaling pathway. Furthermore, this study examined the activity of AMPK, which is a regulator of the JAK2/STAT3 signaling pathway during adipogenesis. The results show that AMPK activity was suppressed in the adipocytes following the stimulation of PA, and the activation of AMPK led to inhibition of the JAK2/STAT3 signaling and reduced DPP4 expression. The rescue experiments proved that the dysregulation of DPP4 in adipocytes upon IR conditions was regulated by the AMPK/JAK2/STAT3 pathway. AMPK can be phosphorylated and activated in response to energy deficit during fasting or cellular stress, and is co-localized with STAT3. Leptin, as a known activator of the JAK2/STAT3, has been reported to be related with the decreased activity of AMPK.37,38 These aforementioned results in combination with our data inspired us to suspect that the increased leptin in obesity might be involved in the regulation of the increased DPP4. Future investigations are necessary to confirm this hypothesize to further understand the systemic regulatory mechanisms of DPP.
Liraglutide is one of the widely used incretin-based drug, and performs well treatment outcomes in diabetes. This study exhibited the liraglutide administration in the constructed IR animal and cell models, and confirm that liraglutide had a good therapeutic effect in obesity-associated IR. Previous studies have reported the regulatory effect of liraglutide on the expression of DPP4 in T2DM.28 In the current study, we observed that the increased DPP4 in the adipose tissues and adipocytes under IR status was significantly reduced by the treatment of liraglutide, which is consistent with the previous study. The stimulation of glucagon-like peptide receptor (GLP-1R), which is achieved by liraglutide, has been reported to promote brown adipose tissue thermogenesis and browning by activating AMPK.39 Our previous studies also have reported that AMPK was up-regulated in adipocyte with liraglutide in db/db mice.29 Thus, we further examined the effects of liraglutide on AMPK and the downstream JAK2/STAT3 signaling pathway, and found that liraglutide intervention in adipocytes led to the activation of AMPK and deactivation of JAK2/STAT3 signaling. Thus, we suspected that liraglutide, as a GLP-1R agonist, might inhibit the expression of DPP4 via the AMPK/JAK2/STAT3 pathway.
In addition to the AMPK, which was demonstrated to regulate the JAK2/STAT3 signaling, PKA was also determined as a regulatory of STAT3 signaling in obesity and obesity-associated hepatocellular carcinoma.40 Another study by Zitman et al reported that liraglutide significantly down-regulated p-STAT3 protein expression through SIRT1 in diabetic kidney disease in db/db mice and in cultured endothelial cells.41 Therefore, we considered that the expression of DPP4 in adipocytes might also modulate by the PKA JAK2/STAT3 pathway or SIRT1 JAK2/STAT3 pathway, which warrants further in-vitro and -vivo studies.
Conclusion
Taken together, all the data in this study revealed that the expression of DPP4 was upregulated in adipose tissues and adipocytes under IR conditions, and this dysregulation was achieved by the AMPK/JAK2/STAT3 pathway. Additionally, liraglutide intervention in led to decreased DPP4 expression also via the regulation of the AMPK/JAK2/STAT3 pathway. This study provides a novel insight into the regulatory mechanism of DPP4 in adipocytes upon IR status, and the AMPK/JAK2/STAT3 pathway may be a novel therapeutic target for adiposity. Although this study investigated the expression changes of DPP4, the levels of soluble DPP4 (sDPP4) in the circulation of IR animal and in cell culture supernatant were not examined, and the activity of sDPP4 was also not evaluated, which are the limitations of our study. Thus, further studies are necessary to uncover the activity of DPP4 and the related regulatory mechanisms.
Acknowledgments
The authors thank Hong Zhang from the Department of Endocrinology of Peking University First Hospital for material support; technicians from the Laboratory Animal Center for animal operational assistance, including Shenshen Kong, Guosheng Yang, Yinghong Tao, Yuanyuan Ma, and Yongyan Hu.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Szulinska M, Gibas-Dorna M, Miller-Kasprzak E, et al. Spirulina maxima improves insulin sensitivity, lipid profile, and total antioxidant status in obese patients with well-treated hypertension: a randomized double-blind placebo-controlled study. Eur Rev Med Pharmacol Sci. 2017;21:2473–2481.
2. Samuel VT, Shulman GI. The pathogenesis of insulin resistance: integrating signaling pathways and substrate flux. J Clin Invest. 2016;126:12–22. doi:10.1172/JCI77812
3. Rebiger L, Lenzen S, Mehmeti I. Susceptibility of brown adipocytes to pro-inflammatory cytokine toxicity and reactive oxygen species. Biosci Rep. 2016;36. doi:10.1042/BSR20150193
4. Wei Y, Chen K, Whaley-Connell AT, et al. Skeletal muscle insulin resistance: role of inflammatory cytokines and reactive oxygen species. Am J Physiol Regul Integr Comp Physiol. 2008;294:R673–R680. doi:10.1152/ajpregu.00561.2007
5. McGown C, Birerdinc A, Younossi ZM. Adipose tissue as an endocrine organ. Clin Liver Dis. 2014;18:41–58. doi:10.1016/j.cld.2013.09.012
6. Giralt M, Cereijo R, Villarroya F. Adipokines and the endocrine role of adipose tissues. Handb Exp Pharmacol. 2016;233:265–282.
7. Gong Z, Tas E, Yakar S, Muzumdar R. Hepatic lipid metabolism and non-alcoholic fatty liver disease in aging. Mol Cell Endocrinol. 2017;455:115–130. doi:10.1016/j.mce.2016.12.022
8. Lamers D, Famulla S, Wronkowitz N, et al. Dipeptidyl peptidase 4 is a novel adipokine potentially linking obesity to the metabolic syndrome. Diabetes. 2011;60:1917–1925.
9. Yanagimachi T, Fujita Y, Takeda Y, et al. Dipeptidyl peptidase-4 inhibitor treatment induces a greater increase in plasma levels of bioactive GIP than GLP-1 in non-diabetic subjects. Mol Metab. 2017;6:226–231. doi:10.1016/j.molmet.2016.12.009
10. Baumeier C, Schluter L, Saussenthaler S, et al. Elevated hepatic DPP4 activity promotes insulin resistance and non-alcoholic fatty liver disease. Mol Metab. 2017;6:1254–1263. doi:10.1016/j.molmet.2017.07.016
11. Marques AP, Cunha-Santos J, Leal H, et al. Dipeptidyl peptidase IV (DPP-IV) inhibition prevents fibrosis in adipose tissue of obese mice. Biochim Biophys Acta Gen Subj. 2018;1862:403–413. doi:10.1016/j.bbagen.2017.11.012
12. Wunderlich CM, Hovelmeyer N, Wunderlich FT. Mechanisms of chronic JAK-STAT3-SOCS3 signaling in obesity. JAKSTAT. 2013;2:e23878.
13. Rohrborn D, Eckel J, Sell H. Shedding of dipeptidyl peptidase 4 is mediated by metalloproteases and up-regulated by hypoxia in human adipocytes and smooth muscle cells. FEBS Lett. 2014;588:3870–3877. doi:10.1016/j.febslet.2014.08.029
14. Sell H, Bluher M, Kloting N, et al. Adipose dipeptidyl peptidase-4 and obesity: correlation with insulin resistance and depot-specific release from adipose tissue in vivo and in vitro. Diabetes Care. 2013;36:4083–4090. doi:10.2337/dc13-0496
15. Svensson H, Oden B, Eden S, Lonn M. Adiponectin, chemerin, cytokines, and dipeptidyl peptidase 4 are released from human adipose tissue in a depot-dependent manner: an in vitro system including human serum albumin. BMC Endocr Disord. 2014;14:7. doi:10.1186/1472-6823-14-7
16. Cristancho AG, Lazar MA. Forming functional fat: a growing understanding of adipocyte differentiation. Nat Rev Mol Cell Biol. 2011;12:722–734. doi:10.1038/nrm3198
17. Sarjeant K, Stephens JM. Adipogenesis. Cold Spring Harb Perspect Biol. 2012;4:a008417. doi:10.1101/cshperspect.a008417
18. Bauvois B, Djavaheri-Mergny M, Rouillard D, Dumont J, Wietzerbin J. Regulation of CD26/DPPIV gene expression by interferons and retinoic acid in tumor B cells. Oncogene. 2000;19:265–272. doi:10.1038/sj.onc.1203292
19. Yuan Y, Xi Y, Chen J, et al. STAT3 stimulates adipogenic stem cell proliferation and cooperates with HMGA2 during the early stage of differentiation to promote adipogenesis. Biochem Biophys Res Commun. 2017;482:1360–1366. doi:10.1016/j.bbrc.2016.12.042
20. Cantwell MT, Farrar JS, Lownik JC, et al. STAT3 suppresses Wnt/beta-catenin signaling during the induction phase of primary Myf5+ brown adipogenesis. Cytokine. 2018;111:434–444. doi:10.1016/j.cyto.2018.05.023
21. Liu Z, Gan L, Zhou Z, Jin W, Sun C. SOCS3 promotes inflammation and apoptosis via inhibiting JAK2/STAT3 signaling pathway in 3T3-L1 adipocyte. Immunobiology. 2015;220:947–953. doi:10.1016/j.imbio.2015.02.004
22. Mancini SJ, White AD, Bijland S, et al. Activation of AMP-activated protein kinase rapidly suppresses multiple pro-inflammatory pathways in adipocytes including IL-1 receptor-associated kinase-4 phosphorylation. Mol Cell Endocrinol. 2017;440:44–56. doi:10.1016/j.mce.2016.11.010
23. Kubota A, Takano H, Wang H, et al. DPP-4 inhibition has beneficial effects on the heart after myocardial infarction. J Mol Cell Cardiol. 2016;91:72–80. doi:10.1016/j.yjmcc.2015.12.026
24. Nagamine A, Hasegawa H, Hashimoto N, et al. The effects of DPP-4 inhibitor on hypoxia-induced apoptosis in human umbilical vein endothelial cells. J Pharmacol Sci. 2017;133:42–48. doi:10.1016/j.jphs.2016.12.003
25. Hwang HJ, Jung TW, Kim BH, et al. A dipeptidyl peptidase-IV inhibitor improves hepatic steatosis and insulin resistance by AMPK-dependent and JNK-dependent inhibition of LECT2 expression. Biochem Pharmacol. 2015;98:157–166. doi:10.1016/j.bcp.2015.08.098
26. Kornelius E, Lin CL, Chang HH, et al. DPP-4 inhibitor linagliptin attenuates abeta-induced cytotoxicity through activation of AMPK in neuronal cells. CNS Neurosci Ther. 2015;21:549–557. doi:10.1111/cns.12404
27. Ladenheim EE. Liraglutide and obesity: a review of the data so far. Drug Des Devel Ther. 2015;9:1867–1875. doi:10.2147/DDDT.S58459.
28. Cao X, Zhou X, Liu XM, Zhou LH. Liraglutide alters DPP4 in the circumvallate papillae of type 2 diabetic rats. J Mol Endocrinol. 2016;57:13–21. doi:10.1530/JME-16-0001
29. Shao Y, Yuan G, Zhang J, Guo X. Liraglutide reduces lipogenetic signals in visceral adipose of db/db mice with AMPK activation and Akt suppression. Drug Des Devel Ther. 2015;9:1177–1184. doi:10.2147/DDDT.S79175
30. Ahmed RH, Huri HZ, Muniandy S, et al. Altered circulating concentrations of active glucagon-like peptide (GLP-1) and dipeptidyl peptidase 4 (DPP4) in obese subjects and their association with insulin resistance. Clin Biochem. 2017;50:746–749. doi:10.1016/j.clinbiochem.2017.03.008
31. Shin J, Fukuhara A, Onodera T, et al. Regulation of dipeptidyl peptidase-4, its substrate chemokines, and their receptors in adipose tissue of ob/ob mice. Horm Metab Res. 2017;49:380–387. doi:10.1055/s-0043-100115
32. Ghorpade DS, Ozcan L, Zheng Z, et al. Hepatocyte-secreted DPP4 in obesity promotes adipose inflammation and insulin resistance. Nature. 2018;555:673–677. doi:10.1038/nature26138
33. Rohrborn D, Bruckner J, Sell H, Eckel J. Reduced DPP4 activity improves insulin signaling in primary human adipocytes. Biochem Biophys Res Commun. 2016;471:348–354. doi:10.1016/j.bbrc.2016.02.019
34. Varin EM, Mulvihill EE, Beaudry JL, et al. Circulating levels of soluble dipeptidyl peptidase-4 are dissociated from inflammation and induced by enzymatic DPP4 inhibition. Cell Metab. 2019;29(320–334):e325. doi:10.1016/j.cmet.2018.10.001
35. Richard AJ, Stephens JM. The role of JAK-STAT signaling in adipose tissue function. Biochim Biophys Acta. 2014;1842:431–439. doi:10.1016/j.bbadis.2013.05.030
36. Kim KS, Kim SH. Effect of N-(D-Ribopyranosyl) taurine sodium salt on the differentiation of human preadipocytes and expression of adipokines through inhibition of STAT-3 signaling in differentiated human adipocytes. Adv Exp Med Biol. 2017;975:667–674.
37. Minokoshi Y, Alquier T, Furukawa N, et al. AMP-kinase regulates food intake by responding to hormonal and nutrient signals in the hypothalamus. Nature. 2004;428(6982):569–574. doi:10.1038/nature02440
38. Yadav A, Kataria MA, Saini V, Yadav A. Role of leptin and adiponectin in insulin resistance. Clin Chim Acta. 2013;417:80–84. doi:10.1016/j.cca.2012.12.007
39. Beiroa D, Imbernon M, Gallego R, et al. GLP-1 agonism stimulates brown adipose tissue thermogenesis and browning through hypothalamic AMPK. Diabetes. 2014;63(10):3346–3358. doi:10.2337/db14-0302
40. Zhou M, Mok MT, Sun H, et al. The anti-diabetic drug exenatide, a glucagon-like peptide-1 receptor agonist, counteracts hepatocarcinogenesis through cAMP-PKA-EGFR-STAT3 axis. Oncogene. 2017;36:4135–4149. doi:10.1038/onc.2017.38
41. Zitman-Gal T, Einbinder Y, Ohana M, et al. The impact of liraglutide on the JAK-STAT pathway in diabetic kidney disease in db/db mice and in cultured endothelial cells. J Diabetes. 2018.
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.