Back to Journals » Infection and Drug Resistance » Volume 15
Distribution of Biocide Resistance Genes and Association with Clonal Complex Genotypes in Staphylococcus aureus Isolated from School-Age Children in Guangzhou
Authors Zhang C, Liang B, Xiong Z, Liang Z, Cai H, Zhong H, Xie Y, Xie Y, Liu X, Xie S, Lan F, Zhou Z
Received 25 August 2022
Accepted for publication 26 November 2022
Published 7 December 2022 Volume 2022:15 Pages 7165—7175
DOI https://doi.org/10.2147/IDR.S387528
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. Héctor Mora-Montes
Chao Zhang,1,2,* Bingshao Liang,1,* Zhile Xiong,1,2 Zhuwei Liang,1,2 Hao Cai,1 Huamin Zhong,1 Yongqiang Xie,1 Yuanwei Xie,2 Xiaochun Liu,2 Shiying Xie,2 Fangjun Lan,2 Zhenwen Zhou2
1Clinical Laboratory, Guangzhou Women and Children’s Medical Center, Guangzhou Medical University, Guangzhou, Guangdong, People’s Republic of China; 2Clinical Laboratory, Longgang District Maternity and Child Healthcare Hospital, Shenzhen, Guangdong, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Zhenwen Zhou, Clinical Laboratory, Longgang District Maternity and Child Healthcare Hospital, Shenzhen, Guangdong, People’s Republic of China, Tel +86 13925097897, Email [email protected]
Purpose: Chlorhexidine and mupirocin are often prescribed to children in affected communities to prevent colonization and transmission of Staphylococcus aureus, but this has led to an increasing rate of biocide resistance. In this study, we aimed to determine the distribution of biocide resistance genes among S. aureus isolates from school-age children in Guangzhou, investigate chlorhexidine gluconate and mupirocin susceptibility and clonal complex (CC) genotypes in strains carrying biocide-resistance genes, and further explore the role of biofilms in this resistance.
Patients and Methods: Antibiotic resistance and multilocus sequence genotyping were performed on 722 S. aureus isolates from previous study. The distribution of nine biocide genes (qacA/B, mupA, mepA, sepA, norA, lmrS, smr, mupB, qacG) was determined by PCR. Isolates carrying qacA/B or mupA genes were further tested for susceptibility to chlorhexidine gluconate (CHG) and mupirocin and biofilm formation abilities.
Results: The most prevalent of the nine biocide resistance genes were mepA (95.57%), followed by norA (78.81%), lmrS (77.01%), and sepA (58.17%). The qacG gene was not detected. Distribution of sepA was significantly decreased in CC30 and CC45 genotypes, and presence of sepA was associated with resistance to antibiotics such as CLI, ERY, TCY, SXT, CIP, and LVX. In addition, 64 (94.1%, n=68) qacA/B+ isolates showed CHG resistance, 12 (100.0%, n=12) mupA+ isolates were mupirocin resistant, and 4 (80%, n=5) and 5 (100%, n=5) qacA/B+mupA+ isolates were CHG and mupirocin resistant, respectively. Of these 85 isolates, 98.8% (n=84) had different degrees of biofilm-forming abilities, which were positively associated with CHG and mupirocin resistance.
Conclusion: The distribution of biocide resistance genes was associated with special CCs. The qacA/B and mupA genes are highly associated with resistance to CHG and mupirocin, and biofilm formation was found to contribute to this biocide resistance.
Keywords: Staphylococcus aureus, biocide resistance genes, qacA/B, children, chlorhexidine, mupirocin, biofilm formation
Introduction
Staphylococcus aureus can cause a variety of infectious diseases, including moderate skin and soft tissue infections, fatal pneumonia, and sepsis. Additionally, the frequent occurrence of antibiotic resistance poses a great challenge for S. aureus prevention, management, and treatment.1,2 Particularly, severe and invasive infections of community-associated methicillin-resistant S. aureus (CA-MRSA) pose a serious threat to one of the most vulnerable groups—children.3–6
S. aureus is reported to frequently colonize populations, and children in many regions have higher rates of S. aureus nasal colonization (up to 40%).2,7–9 This is a significant issue as the colonization of S. aureus contributes to its transmission and increases the risk of infection. Decolonization agents, such as mupirocin and chlorhexidine, have been widely used in healthcare-associated MRSA infections, and have also been prescribed to children in community settings to prevent S. aureus infectious diseases. Mupirocin is a commonly used antiseptic that inhibits isoleucyl tRNA synthetase, thus preventing the protein synthesis of bacteria.10 Chlorhexidine gluconate (CHG) can covalently bind to the S. aureus membrane and ultimately lead to cell depolarization and death. However, in recent decades, widespread biocide use has resulted in a general decrease in antibiotic susceptibility of S. aureus.11–13
With the deepening of our knowledge on the mechanism of MRSA drug resistance, an increasing number of antimicrobial agents, like pyrazole-based analogs, benzimidazole analogues, indole-based derivatives, and arylethenesulfonyl fluorides, have recently proven efficient against MRSA.14–17 At the same time, more factors associated with drug resistance of S. aureus also need to be discussed.
Evidence has shown that S. aureus strains exhibiting resistance to CHG and mupirocin are highly associated with chromosomally-encoded biocide resistance genes and plasmid-encoded multidrug resistance (MDR) efflux pumps. Plasmid-mediated genes such as qacA/B encode the major facilitator superfamily, whereas genes such as smr and qacG encode the small multidrug resistance family, which are the major MDR efflux pump proteins. Other plasmid-mediated genes such as mupA and mupB encode novel isoleucyl tRNA synthetases, thus conferring increased antimicrobial resistance of S. aureus, including to mupirocin.10,18 In addition, chromosomally-encoded efflux genes such as mepA, norA, lmrS, sepA have been reported to provide bacteria with multidrug and biocide resistance or help them survive in severe conditions. The prevalence of biocide resistance genes is high among S. aureus strains worldwide.19–21
In addition, it is believed that biofilms, a type of dynamic and complex biological system, make S. aureus infections more challenging because they can increase the resistance of staphylococcal species to both the immune system and antibiotics.22–24 Although some biocides targeting S. aureus biofilm mechanisms have been successfully developed and have showed promise,25,26 it has also been reported that sub-inhibitory concentrations of CHG or mupirocin used in daily life may promote biofilm formation by S. aureus.27,28 Therefore, the role of S. aureus biofilm formation in its resistance to biocides such as CHG and mupirocin needs to be further investigated.
Herein, we detected the distribution of nine biocide-resistant genes (qacA/B, mupA, mepA, sepA, norA, lmrS, smr, mupB, qacG) in 722 S. aureus isolates from nasal swabs of school-age children in Guangzhou, Southern China, and CHG and mupirocin resistance phenotypes, gene type characteristics, and biofilm formation ability in 85 qacA/B or mupA positive isolates were analyzed.
Materials and Methods
All methods were carried out in accordance with the relevant guidelines and regulations.
- Bacterial isolates. A total of 722 S. aureus strains isolated from nasal swabs of 2375 school-age students (6–18 years) in Guangzhou were recovered between October and November 201829 (22 isolates were excluded due to quality problems caused by long-term storage). All the participants were recruited voluntarily and their guardians provided written informed consent. This study was approved by The Ethics Committee of Guangzhou Woman and Children’s Medical Center (registration no. 2016081029). All S. aureus isolates were identified and detected for the presence of femB and mecA. Resistance to ten conventional antibiotics had been previously determined (PEN, penicillin; CLI, clindamycin; ERY, erythromycin; TCY, tetracycline; SXT, sulfamethoxazole-trimethoprim; RIF, rifampicin; CIP, ciprofloxacin; LVX, levofloxacin; MFX, moxifloxacin; GEN, gentamicin).29
- Detection of biocide resistance genes by PCR. Total DNA was extracted and nine genes (qacA/B, mupA, mepA, sepA, mupB, smr, norA, lmrS, qacG) were amplified by polymerase chain reaction (PCR) as previously described.30 The nucleotide sequences of the primers used in PCR are listed in Table 1. Positive amplicons were randomly selected and sent for sequencing at BGI (Shenzhen, China), and BLAST on the NCBI website (www.ncbi.nlm.nih.gov) was used to confirm sequence consistency. Finally, partial nucleotide sequences of eight genes (qacA/B, mupA, mepA, sepA, mupB, smr, norA, lmrS) were submitted to GenBank repository (https://www.ncbi.nlm.nih.gov/).
- Chlorhexidine gluconate and mupirocin susceptibility test. Broth microdilution was used to test CHG and mupirocin susceptibility according to the Clinical and Laboratory Standards Institute (CLSI) M07.31 Susceptibility to CHG was tested using a twofold dilution series from 32 to 0.0625 mg/L, starting with a 20% (w/v) CHG solution (Macklin, Shanghai, China).32 Isolates were considered susceptible to CHG if the MIC was <4 mg/L and had a tolerance to CHG at MIC ≥4 mg/L.33 Mupirocin susceptibility was determined by another twofold dilution series from 1024 mg/L to 1 mg/L (Macklin), which was classified into three categories, depending on the MIC to mupirocin: ≤4 mg/L as mupirocin-susceptible, 8–256 mg/L as low-level mupirocin resistance (LLMR), and ≥512 mg/L as high-level mupirocin resistance (HLMR).34
- Biofilm formation using tissue culture plate (TCP) assay. The biofilm formation test in this study was based on Stepanovic’s recommendations and minimally modified.35 Briefly, S. aureus isolates from fresh Columbia blood agar plates were inoculated into 5 mL Trypticase Soy broth (TSB) and incubated on a 220-rpm shaker overnight at 37 ℃. The cultures were then diluted to 1:50 with TSB containing 0.25% glucose, and 200 μL was added (in triplicate) to sterile 96-U-bottom polystyrene plates for 18 h at 37 ℃. Uninoculated wells containing TSB and glucose served as negative controls. Wells were washed with PBS solution three times and dried at 60 ℃ for 1 h; then, 1% crystal violet was used to stain adherent bacteria for 10–15 minutes. After washing three times with distilled water to remove excess stain and allowing them to dry at room temperature, a microplate reader (Multiskan FC, Thermo Fisher, USA) was used to measure optical density (OD) at 570 nm. ODc = average OD of negative control + (3×SD of negative control); biofilm formation ability was divided into four degrees: non (OD≤ODc), weak (ODc<OD≤2ODc), moderate (2ODc<OD≤4ODc), and strong (4ODc<OD).
- Multilocus sequence typing (MLST), clonal complex (CC) genotyping, and data analysis. Multilocus sequence typing numbers were determined previously,29 and the eBURST v.3 program (https://www.mlst.net/eburst/) was used to conduct CCs. All data were entered and analyzed using GraphPad Prism 8.0.1 (GraphPad Software Inc, San Diego, CA, USA) and categorical variables were compared using Fisher’s exact test. The results were considered statistically significant at P<0.05.
 |
Table 1 Sequences of Primers for PCR of Biocide Resistance Genes |
Results
Prevalence and Distribution Characteristics of Biocide Resistance Genes
S. aureus isolates were collected from nasal swabs of school-age students in Guangzhou, where carriers were recruited from senior high school (n=192, 26.59%), junior middle school (n=182, 25.21%), and elementary school (n=348, 48.20%). The overall prevalence of qacA/B, smr, norA, mepA, sepA, mupA, mupB, and lmrS was 10.11% (n=73), 1.39% (n=10), 78.81% (n=569), 95.57% (n=690), 58.17% (n=420), 2.35% (n=17), 0.42% (n=3), 77.01% (n=556), respectively. Of note, five isolates harbored both qacA/B and mupA (qacA/B+mupA+) and four isolates harbored both qacA/B and smr (qacA/B+smr+). The qacG gene was not detected. The prevalence of all these genes in the MRSA and methicillin-susceptible S. aureus (MSSA) populations showed no significant difference (P>0.05) (Figure 1A).
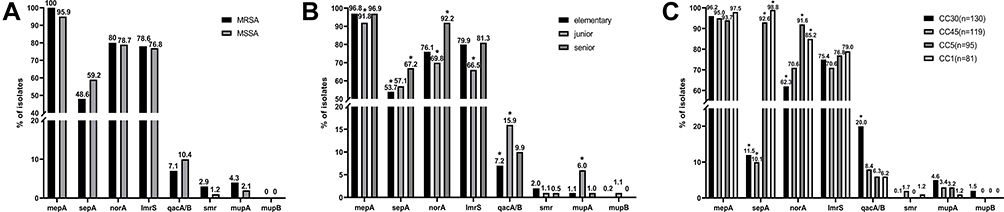 |
Figure 1 The distribution of biocide resistance genes in different groups. (A) MRSA and MSSA, (B) educational ages, and (C) four main clonal complex (CC) genotypes among 722 S. aureus isolates from school-age students in Guangzhou. The data shown above the column are percentages in that population. “*” indicates statistics which were significantly different (P<0.05). |
Compared to the other educational age groups, elementary school children had lower sepA (53.7%) and qacA/B (7.18%) carrier rates, and senior school students had a higher prevalence of sepA (67.18%) and norA (92.18%) (P<0.05). Additionally, a significantly lower prevalence of mepA (91.76%), norA (69.78%), and lmrS (66.48%) and a higher prevalence of qacA/B (15.93%) and mupA (6.04%) were found in junior school children (P<0.05) compared to other populations (Figure 1B). Furthermore, among all CC genotype groups, the distribution of sepA and norA was significantly lower in CC30 groups and higher in CC45 groups (P<0.05), while the distribution of qacA/B was significantly higher in the CC30 genotype (P<0.05) (Figure 1C).
Association Between Biocide Resistance Genes and Antibiotic Resistance
Table 2 shows the association between ten biocide resistance genes and conventional antibiotic resistance among 722 Staphylococcus aureus isolates. The qacA/B and mupA genes were associated with S. aureus resistance to clindamycin (P = 0.0004, P = 0.0253) and erythromycin (P = 0.006, P = 0.0216), the mepA gene was associated with penicillin (P = 0.0163) and clindamycin (P = 0.0001) resistance, and the norA gene was correlated with resistance to penicillin (P = 0.0011), clindamycin (P = 0.0002), and erythromycin (P = 0.0012). In addition, sepA+ isolates were more frequently resistant to clindamycin, erythromycin, tetracycline, sulfamethoxazole-trimethoprim, ciprofloxacin, and levofloxacin (P<0.05). No additional antibiotic associations were found for smr, mupB and lmrS genes.
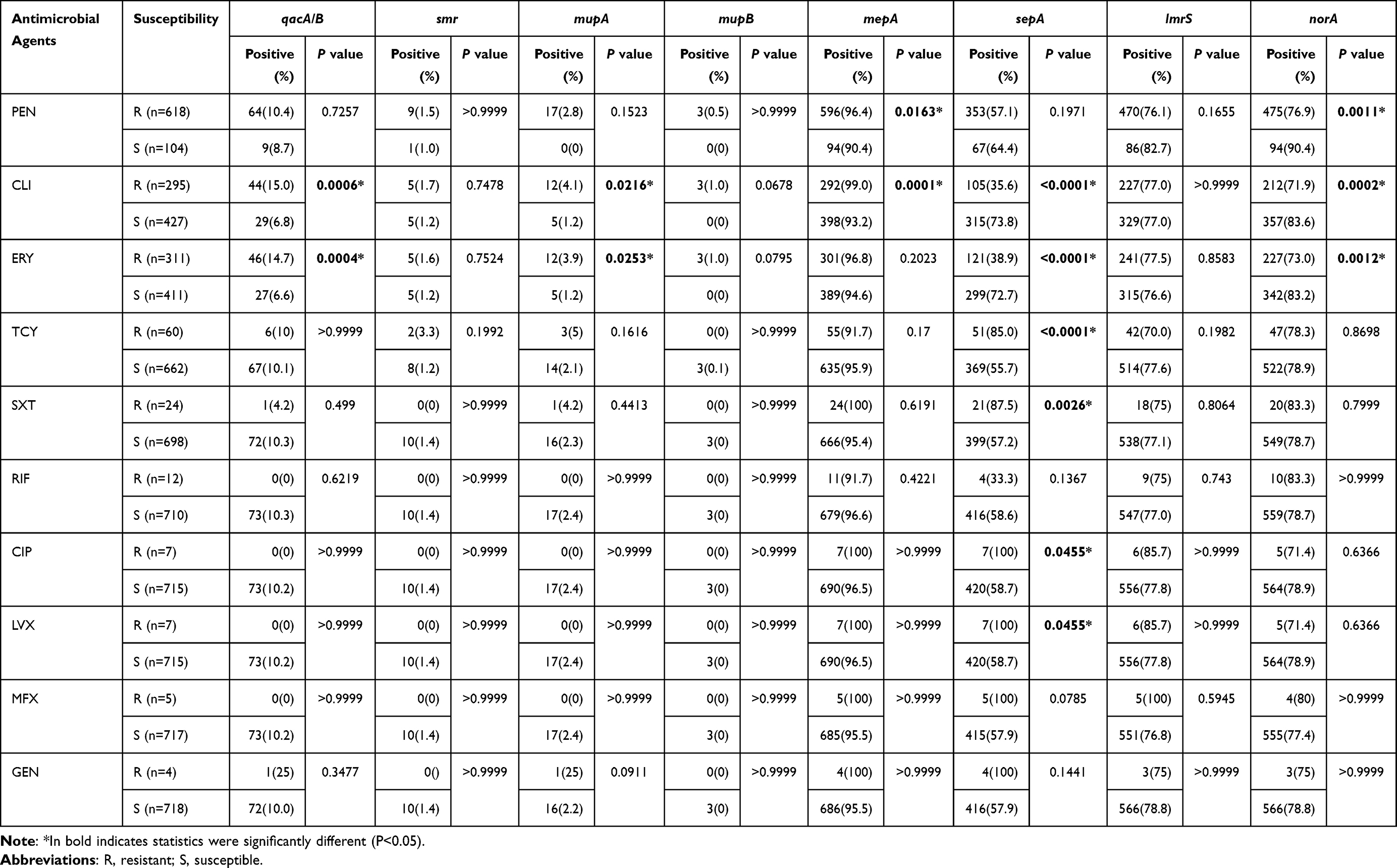 |
Table 2 Association Between ten Biocide Resistance Genes and Conventional Antibiotic Resistance Among 722 Staphylococcus Aureus Isolates |
CHG and Mupirocin Resistance and Its Distribution by CC Genotype
In 722 S. aureus isolates, 85 isolates carrying qacA/B or mupA were screened to test the susceptibility to CHG and mupirocin (68 isolates carried qacA/B but no mupA gene, 12 isolates carried only the latter, and 5 isolates carried both). Among 68 isolates carrying qacA/B only, 64 (94.1%) were resistant to CHG and 4 (5.9%) were susceptible to CHG. All 12 isolates carrying only mupA were resistant to mupirocin, including seven (58.3%) LLMR and five (41.7%) HLMR isolates. Among the five isolates carrying both genes, four (80%) were resistant to CHG, and five (100%) were HLMR (Table 3).
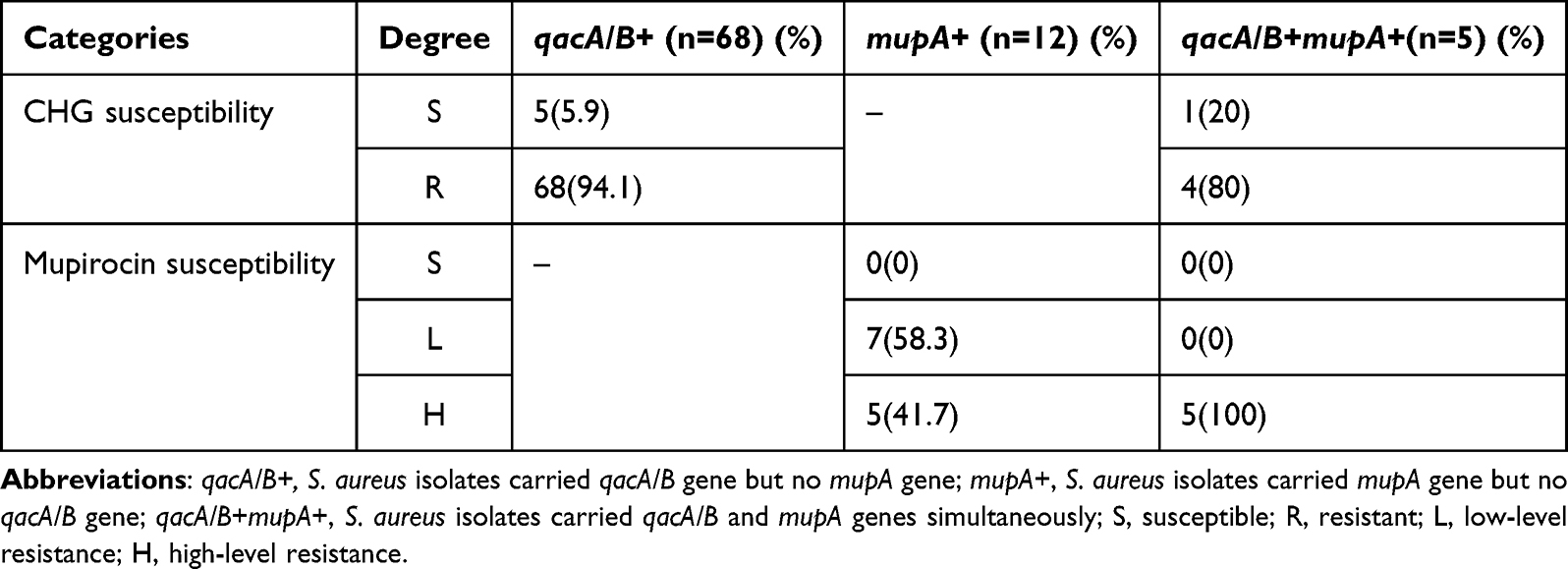 |
Table 3 The Susceptibility to CHG & Mupirocin and Biofilm Formation Ability of qacA/B and mupA Genes Positive Isolates |
Next, 68 and 17 isolates resistant to CHG and mupirocin, respectively, were divided to determine their distribution by CC genotype. Among the isolates resistant to CHG, the top nine CCs were CC30 (36.76%), CC45 (14.72%), CC5 (8.82%), CC1 (7.35%), CC398 (7.35%), CC22 (4.41%), CC59 (4.41%), CC15 (4.41%), and CC7 (4.41%), whereas the other CCs included only one isolate (Figure 2A). Among the 17 isolates resistant to mupirocin, the main CCs were CC30 (29.41%), CC45 (23.53%), and CC5 (17.65%), and the others included only one isolate (Figure 2B).
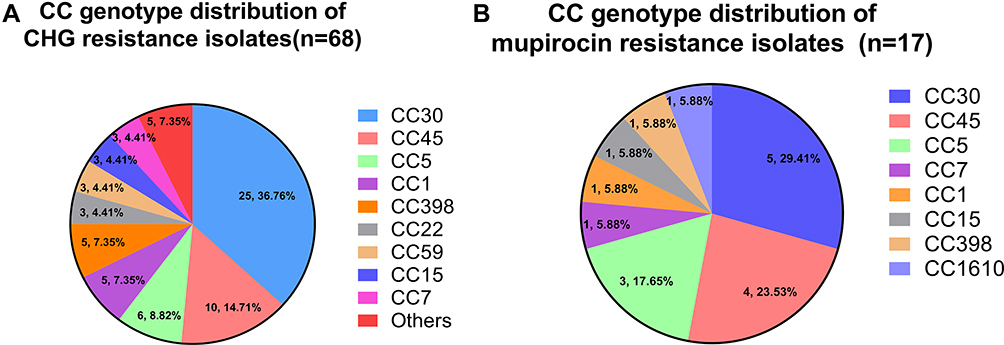 |
Figure 2 CC genotype distribution of (A) CHG and (B) mupirocin resistance isolates. “Others” represent CC8, CC25, CC121, CC944 and CC1610 genotypes; each includes only one isolate. |
Biofilm Formation Among qacA/B and mupA Positive Isolates
A tissue culture plate (TCP) assay was used to assess the biofilm-forming ability of 85 isolates carrying qacA/B or mupA. Overall, 84 (98.8%) isolates produced biofilms, and one (1.2%) isolate did not. Among the biofilm-producing isolates, 40 (47.1%) produced weak biofilms, 37 (43.5%) produced moderate biofilms, and seven (8.2%) isolates produced strong biofilms.
Among the 73 qacA/B positive isolates, 7 (9.6%) were firmly adherent, 34 (47.2%) were moderately adherent, 31 (42.5%) were weakly adherent, and 1 (1.4%) was non-adherent; among the 17 mupA positive isolates, 8 (47.1%) were moderate biofilm producers, and 9 (52.9%) isolates produced weak biofilms. To meet the condition of all expected values being greater than 1.0 and at least 20% of the expected values being greater than 5, we combined non and weak biofilm producers into a “+” group and moderate and strong biofilm producers into a “++” group (Table 4). The relationship between biofilm formation ability and the resistance of S. aureus to CHG and mupirocin was discovered. The results showed that among qacA/B- and mupA- positive isolates, biofilm formation ability was positively associated with resistance to CHG and mupirocin (P<0.05).
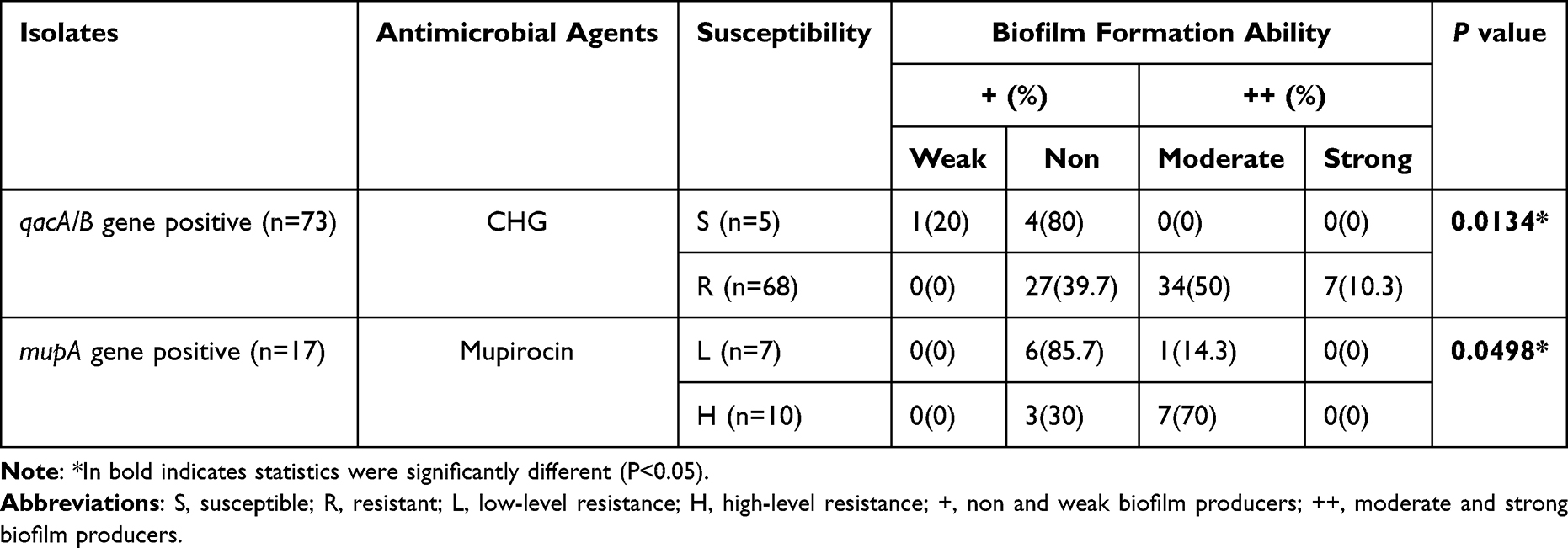 |
Table 4 Relationship Between CHG & Mupirocin Susceptibility and Biofilm Formation Ability Among qacA/B & mupA Positive Isolates |
Discussion
Over the past decade, the susceptibility of S. aureus to CHG and mupirocin has been decreasing, mainly owing to the prevalence of a variety of biocide resistance genes.34,36,37 It has been reported that these genes on plasmids or chromosomes are distributed worldwide and differ significantly throughout the different regions infected with S. aureus.38 From the data available, the plasmid-mediated qacA/B and smr genes prevailed at 35.4–41.6%, with 0–31% of S. aureus in the southern Asian region.38–40 In our study, only 10.11% and 1.39% of isolates carried qacA/B and smr genes, respectively, and 0.55% (n=5) of isolates carried both genes simultaneously, which was lower than that reported by McNeil et al for the Texas children’s hospital at 22.7% and 33.1% of isolates harboring qacA/B and smr genes, respectively, and 10.9% carrying both simultaneously.41 This could likely be owing to the lower prevalence of these genes in community-associated isolates compared to hospital-associated isolates. In addition, sporadic harboring of the smr gene is thought to confer less advantageous resistance compared to qacA/B.40 Plasmid-mediated genes, mupA and mupB, were detected in 2.35% and 0.42% of the isolates, respectively, and this prevalence was similar to that of clinical S. aureus isolates in children from Iran and France.42,43 The qacG gene has been rarely reported, except in strains isolated from food sources and animals,44,45 and likewise, no qacG gene was found in any of the 722 S. aureus isolates analyzed in this study.
In parallel, a high distribution of chromosomally-encoded efflux genes, mepA (95.57%) and norA (78.81%), was determined in the present study, which was similar to those found in Europe, Africa, and Asia.19–21 However, compared to some European and African studies,19,20 a lower prevalence of lmrS (77.01%) and sepA (58.17%) was found in S. aureus isolates from children’s nasal swabs in Guangzhou, suggesting that the distribution of these genes varied by region. Overall, we provided updated data on the prevalence of biocide resistance genes among S. aureus colonies in the nasal cavity of school-aged children in Guangzhou, which showed large regional differences.
In addition, we found that the distribution of biocide resistance genes among S. aureus was related to the educational age and CC groups in this study. Although qacA/B or smr genes were reported to be more common in MRSA populations,46,47 our results showed that MRSA and MSSA populations have a similar distribution of these genes (P>0.05).
To the best of our knowledge, the relationship between biocide resistance gene distribution among S. aureus and children’s educational age has scarcely been investigated. We found that among school-age students from elementary school through to senior school, junior school students had a significantly higher prevalence of qacA/B and mupA genes and senior school students had a higher prevalence of norA and sepA genes (P<0.05), suggesting that there may be a wide distribution of biocide resistance genes in children of specific age groups.
MDR efflux pumps encoded by genes such as qacA/B, mepA, sepA, norA, lmrS, smr, qacG may lead to high-level resistance of S. aureus to multiple antibiotics.38 We found that qacA/B was related to resistance to erythromycin and clindamycin, which was consistent with the findings of Teresa et al19 However, unlike the known roles of mepA and norA in S. aureus to confer fluoroquinolone resistance (norfloxacin and ciprofloxacin),38,48 the present study discovered that the mepA gene in S. aureus was positively associated with penicillin and clindamycin resistance, whereas the norA gene was inversely associated with resistance to penicillin, clindamycin, and erythromycin. However, these findings need to be further verified using a larger sample size. The smr gene has been reported to be related to ciprofloxacin resistance in other studies,41 but it was not detected in our study, probably owing to its low prevalence.
Previously, Isa et al revealed that higher MICs of chlorhexidine were exhibited among S. aureus isolates from CC5 and CC22 groups, which originated from humans and animals in Germany,49 suggesting that the phenotype or genotype of biocide resistance may also be related to CC groups.
It is worth noting that among all CC genotypes of the 722 S. aureus isolates, the distribution of sepA was significantly low in CC30 (11.5%) and CC45 (10.1%) groups and high in CC5 (92.6%) and CC1 (98.8%) groups (P<0.05), while the distribution of qacA/B (20.0%) and mupA (4.6%) genes had the highest proportion in the CC30 group. In addition, in this study we found that sepA was inversely associated with resistance to CLI and ERY, while qacA/B and mupA were positively associated with resistance to these antibiotics. These results explain our previous findings that CC30 genotypes had the highest proportion of ERY-and CLI-resistant S. aureus isolates, while CC1 possessed the lowest proportion thereof.29 Moreover, we also revealed that the reason for the lower prevalence of sepA (58.17%) among S. aureus isolates found in Guangzhou compared to other regions (often as high as 90%)19,20 was owing to varying distributions of this gene by region. The sepA gene was prevalent in CC5 (92.6%) and CC1 (98.8%) groups, but sporadically distributed in CC30 (11.5%) and CC45 (10.1%) groups (P<0.05), while CC30 and CC45 were the main S. aureus CC genotypes carried by children in Guangzhou.
Considering qacA/B and mupA are well known to be highly associated with resistance to CHG and mupirocin,30,50 85 isolates carrying qacA/B or mupA were selected to further test their phenotypes. The three major CC genotypes for CHG and mupirocin resistance were CC30, CC45, and CC5; this trend was consistent with the distribution of the whole population.29 In this study, 64 qacA/B genes were associated with CHG resistance in 94.1% (n=68) of S. aureus isolates and the mupA gene was associated with resistance to mupirocin in 100% (n=12) of the isolates (including 7 LLMR and 5 HLMR), which verified that the qacA/B and mupA genes were highly resistant to chlorhexidine and mupirocin. Notably, we found five S. aureus isolates harboring both qacA/B and mupA genes, four (80%) resistant to CHG, and five (100%) highly resistant to mupirocin, suggesting that the qacA/B gene may also increase high-level mupirocin resistance among mupA+ S. aureus isolates. Furthermore, we discovered whether the co-occurrences of qacA/B and mupA may also promote resistance to multiple antibiotics; among the five qacA/B+mupA+ isolates, three (60%) isolates were resistant to ERY and CLI, one (20%) isolate was resistant to OXA, SXT, and TCY, and two (40%) were intermediately resistant to RIF; this resistance rate was higher than when the two genes were present alone, but the underlying mechanism needs to be further explored.
Biofilms have been receiving much attention because of their association with resistance to antibiotic agents in S. aureus; strains within biofilms are thought to be hundreds of times more resistant to antibiotics than planktonic strains.23,51 It was previously reported that a larger proportion (up to 90%) of S. aureus produced biofilms from nasal swabs compared to clinical isolates;52 in this study, we also found a high proportion of biofilm producers (98.8%) among qacA/B-positive and mupA-positive S. aureus isolates from nasal swabs of school-age children. Similar to the study by Barakat et al,53 we suggested that the biofilm formation ability of S. aureus was positively correlated with its resistance to CHG and mupirocin (P<0.05). This finding raises additional concerns about the potential risks of S. aureus nasal colonization in school-aged children.
This study has several limitations. We only performed susceptibility to CHG and mupirocin and biofilm formation tests among S. aureus isolates carrying qacA/B and mupA. Thus, comparative analysis or a larger sample size will enhance the reliability of our results. Subsequently, we plan to further determine the distribution characteristics of S. aureus resistance to mupirocin and chlorhexidine in school-age children in Guangzhou and investigate the underlying mechanisms of mupirocin and CHG resistance in S. aureus. Otherwise, in the face of the complex mechanism of S. aureus’ antibiotic resistance, the development of drugs targeting its structure-activity relationship is expected in future.
Conclusion
This study revealed the distribution characteristics of biocide resistance genes among S. aureus isolates colonizing the nasal cavity of school-aged children in Guangzhou. A significant association was observed between the distribution of these genes, educational age, and specific CCs. Furthermore, we verified that qacA/B and mupA could be highly resistant to chlorhexidine and mupirocin, and preliminarily discovered that biofilms formed by S. aureus contribute to this resistance. Our results suggest that chlorhexidine and mupirocin are still effective against the decolonization of S. aureus in children in Guangzhou, but it should be noted that some biocide resistance genes are widely carried and closely related to multidrug resistance. Moreover, this study indicated that the biofilm-forming ability of S. aureus should receive more attention for the prevention and management of community and nosocomial infections.
Abbreviations
S. aureus, Staphylococcus aureus; CA-MRSA, community-associated methicillin-resistant Staphylococcus aureus; CHG, chlorhexidine gluconate; MDR, multidrug resistance; MRSA, methicillin-resistant Staphylococcus aureus; MSSA, methicillin-sensitive Staphylococcus aureus; TCP, tissue culture plate; TSB, trypticase soy broth; PBS, phosphate-buffered saline; PEN, penicillin; CLI, clindamycin; ERY, erythromycin; TCY, tetracycline; SXT, sulfamethoxazole-trimethoprim; RIF, rifampicin; CIP, ciprofloxacin; LVX, levofloxacin; MFX, moxifloxacin; GEN, gentamicin; PCR, polymerase chain reaction; OD, optical density; LLMR, low-level mupirocin resistance; HLMR, high-level mupirocin resistance.
Data Sharing Statement
The datasets generated and/or analyzed during the current study are available in the GenBank repository (https://www.ncbi.nlm.nih.gov/), the GenBank accession numbers for the eight partial nucleotide sequences in this study were below: ON448386 (sepA), ON448387 (lmrS), ON448388 (mupA), ON448389 (qacA/B), ON448390 (mepA), ON448391 (norA), ON448393 (smr), ON448392 (mupB).
Ethics Approval and Consent to Participate
This study was approved by the Ethics Committee of Guangzhou Women and Children’s Medical Center (registration no. 2016081029). All the participants were recruited voluntarily and their guardians have provided written informed consent. We declare that our study complies with the Declaration of Helsinki.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This study was funded by the National Natural Science Foundation of China (No. 82002202), Guangzhou Science Technology and Innovation Commission (No. 201707010010), Basic Research Project of Shenzhen Municipality (Natural Science Foundation) (JCYJ20220530162415036) and Longgang Medical and Health Science and Technology Project (LGKCYLWS2022029).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Tong SYC, Davis JS, Eichenberger E, Holland TL, Fowler VG. Staphylococcus aureus infections: epidemiology, pathophysiology, clinical manifestations, and management. Clin Microbiol Rev. 2015;28(3):603–661. doi:10.1128/CMR.00134-14
2. Lee AS, de Lencastre H, Garau J, et al. Methicillin-resistant Staphylococcus aureus. Nat Rev Dis Primers. 2018;4:18033. doi:10.1038/nrdp.2018.33
3. Centers for Disease Control and Prevention. Four pediatric deaths from community-acquired methicillin-resistant Staphylococcus aureus — Minnesota and North Dakota, 1997–1999. MMWR Morb Mortal Wkly Rep. 1999;48(32):707–710.
4. David MZ, Daum RS. Community-associated methicillin-resistant Staphylococcus aureus: epidemiology and clinical consequences of an emerging epidemic. Clin Microbiol Rev. 2010;23(3):616–687. doi:10.1128/CMR.00081-09
5. Lorton F, Chalumeau M, Martinot A, et al. Epidemiology of community-onset severe bacterial infections in children and its evolution: a population-based study in France. Pediatr Crit Care Med. 2020;21(6):e325–e32. doi:10.1097/PCC.0000000000002300
6. Agyeman PKA, Schlapbach LJ, Giannoni E, et al. Epidemiology of blood culture-proven bacterial sepsis in children in Switzerland: a population-based cohort study. Lancet Child Adolesc Health. 2017;1(2):124–133. doi:10.1016/S2352-4642(17)30010-X
7. Gorwitz RJ, Kruszon-Moran D, McAllister SK, et al. Changes in the prevalence of nasal colonization with Staphylococcus aureus in the United States, 2001–2004. J Infect Dis. 2008;197(9):1226–1234. doi:10.1086/533494
8. Wertheim HFL, Melles DC, Vos MC, et al. The role of nasal carriage in Staphylococcus aureus infections. Lancet Infect Dis. 2005;5(12):751–762. doi:10.1016/S1473-3099(05)70295-4
9. Datta F, Erb T, Heininger U, et al. A multicenter, cross-sectional study on the prevalence and risk factors for nasal colonization with Staphylococcus aureus in patients admitted to children’s hospitals in Switzerland. Clin Infect Dis. 2008;47(7):923–926. doi:10.1086/591700
10. Patel JB, Gorwitz RJ, Jernigan JA. Mupirocin resistance. Clin Infect Dis. 2009;49(6):935–941. doi:10.1086/605495
11. Septimus EJ, Schweizer ML. Decolonization in prevention of health care-associated infections. Clin Microbiol Rev. 2016;29(2):201–222. doi:10.1128/CMR.00049-15
12. Loeb MB, Main C, Eady A, Walker-Dilks C. Antimicrobial drugs for treating methicillin-resistant Staphylococcus aureus colonization. Cochrane Database Syst Rev. 2003;2003(4):CD003340.
13. Poovelikunnel T, Gethin G, Humphreys H. Mupirocin resistance: clinical implications and potential alternatives for the eradication of MRSA. J Antimicrob Chemother. 2015;70(10):2681–2692. doi:10.1093/jac/dkv169
14. Verma R, Verma SK, Rakesh KP, et al. Pyrazole-based analogs as potential antibacterial agents against methicillin-resistance staphylococcus aureus (MRSA) and its SAR elucidation. Eur J Med Chem. 2021;212:113134. doi:10.1016/j.ejmech.2020.113134
15. Zha G-F, Preetham HD, Rangappa S, et al. Benzimidazole analogues as efficient arsenals in war against methicillin-resistance staphylococcus aureus (MRSA) and its SAR studies. Bioorg Chem. 2021;115:105175. doi:10.1016/j.bioorg.2021.105175
16. Qin H-L, Liu J, Fang W-Y, Ravindar L, Rakesh KP. Indole-based derivatives as potential antibacterial activity against methicillin-resistance Staphylococcus aureus (MRSA). Eur J Med Chem. 2020;194:112245. doi:10.1016/j.ejmech.2020.112245
17. Zha G-F, Wang S-M, Rakesh KP, et al. Discovery of novel arylethenesulfonyl fluorides as potential candidates against methicillin-resistant of Staphylococcus aureus (MRSA) for overcoming multidrug resistance of bacterial infections. Eur J Med Chem. 2019;162:364–377. doi:10.1016/j.ejmech.2018.11.012
18. McNeil JC, Hulten KG, Kaplan SL, Mason EO. Mupirocin resistance in Staphylococcus aureus causing recurrent skin and soft tissue infections in children. Antimicrob Agents Chemother. 2011;55(5):2431–2433. doi:10.1128/AAC.01587-10
19. Conceição T, de Lencastre H, Aires-de-Sousa M. Prevalence of biocide resistance genes and chlorhexidine and mupirocin non-susceptibility in Portuguese hospitals during a 31-year period (1985–2016). J Glob Antimicrob Resist. 2021;24:169–174. doi:10.1016/j.jgar.2020.12.010
20. Conceição T, Coelho C, de Lencastre H, Aires-de-Sousa M. High prevalence of biocide resistance determinants in staphylococcus aureus isolates from three African countries. Antimicrob Agents Chemother. 2016;60(1):678–681. doi:10.1128/AAC.02140-15
21. Liu Q, Zhao H, Han L, Shu W, Wu Q, Ni Y. Frequency of biocide-resistant genes and susceptibility to chlorhexidine in high-level mupirocin-resistant, methicillin-resistant Staphylococcus aureus (MuH MRSA). Diagn Microbiol Infect Dis. 2015;82(4):278–283. doi:10.1016/j.diagmicrobio.2015.03.023
22. McCarthy H, Rudkin JK, Black NS, Gallagher L, O’Neill E, O’Gara JP. Methicillin resistance and the biofilm phenotype in Staphylococcus aureus. Front Cell Infect Microbiol. 2015;5:1. doi:10.3389/fcimb.2015.00001
23. Schilcher K, Horswill AR. Staphylococcal biofilm development: structure, regulation, and treatment strategies. Microbiol Mol Biol Rev. 2020;84(3). doi:10.1128/MMBR.00026-19
24. Günther F, Blessing B, Tacconelli E, Mutters NT. MRSA decolonization failure-are biofilms the missing link? Antimicrob Resist Infect Control. 2017;6:32. doi:10.1186/s13756-017-0192-1
25. Zhang X, Manukumar HM, Rakesh KP, et al. Role of BP*C@AgNPs in Bap-dependent multicellular behavior of clinically important methicillin-resistant Staphylococcus aureus (MRSA) biofilm adherence: a key virulence study. Microb Pathog. 2018;123:275–284. doi:10.1016/j.micpath.2018.07.025
26. Mohammed YHE, Manukumar HM, Rakesh KP, Karthik CS, Mallu P, Qin H-L. Vision for medicine: staphylococcus aureus biofilm war and unlocking key’s for anti-biofilm drug development. Microb Pathog. 2018;123:339–347. doi:10.1016/j.micpath.2018.07.002
27. Sritharadol R, Hamada M, Kimura S, Ishii Y, Srichana T, Tateda K. Mupirocin at subinhibitory concentrations induces biofilm formation in Staphylococcus aureus. Microb Drug Resist. 2018;24(9):1249–1258. doi:10.1089/mdr.2017.0290
28. Park KH, Jung M, Kim DY, et al. Effects of subinhibitory concentrations of chlorhexidine and mupirocin on biofilm formation in clinical meticillin-resistant Staphylococcus aureus. J Hosp Infect. 2020;106(2):295–302. doi:10.1016/j.jhin.2020.07.010
29. Liang B, Liang X, Gao F, et al. Active surveillance, drug resistance, and genotypic profiling of among school-age children in China. Front Med. 2021;8:701494. doi:10.3389/fmed.2021.701494
30. Fritz SA, Hogan PG, Camins BC, et al. Mupirocin and chlorhexidine resistance in Staphylococcus aureus in patients with community-onset skin and soft tissue infections. Antimicrob Agents Chemother. 2013;57(1):559–568. doi:10.1128/AAC.01633-12
31. CLSI. Methods for Dilution Antimicrobial Susceptibility Tests for Bacterial That Grow Aerobically CLSI Standard M07.
32. Lu Z, Chen Y, Chen W, et al. Characteristics of qacA/B-positive Staphylococcus aureus isolated from patients and a hospital environment in China. J Antimicrob Chemother. 2015;70(3):653–657. doi:10.1093/jac/dku456
33. McDanel JS, Murphy CR, Diekema DJ, et al. Chlorhexidine and mupirocin susceptibilities of methicillin-resistant staphylococcus aureus from colonized nursing home residents. Antimicrob Agents Chemother. 2013;57(1):552–558. doi:10.1128/AAC.01623-12
34. Hogue JS, Buttke P, Braun LE, Fairchok MP. Mupirocin resistance related to increasing mupirocin use in clinical isolates of methicillin-resistant Staphylococcus aureus in a pediatric population. J Clin Microbiol. 2010;48(7):2599–2600. doi:10.1128/JCM.02118-09
35. Stepanović S, Vuković D, Hola V, et al. Quantification of biofilm in microtiter plates: overview of testing conditions and practical recommendations for assessment of biofilm production by staphylococci. APMIS. 2007;115(8):891–899. doi:10.1111/j.1600-0463.2007.apm_630.x
36. Azrad M, Shmuel C, Leshem T, et al. Reduced susceptibility to chlorhexidine among isolates in Israel: phenotypic and genotypic tolerance. Antibiotics. 2021;10(3):342. doi:10.3390/antibiotics10030342
37. Madden GR, Sifri CD. Antimicrobial resistance to agents used for Staphylococcus aureus decolonization: is there a reason for concern. Curr Infect Dis Rep. 2018;20(8):26. doi:10.1007/s11908-018-0630-0
38. Hassanzadeh S, Ganjloo S, Pourmand MR, Mashhadi R, Ghazvini K. Epidemiology of efflux pumps genes mediating resistance among Staphylococcus aureus; A systematic review. Microb Pathog. 2020;139:103850. doi:10.1016/j.micpath.2019.103850
39. Sheng W-H, Wang J-T, Lauderdale T-L, Weng C-M, Chen D, Chang S-C. Epidemiology and susceptibilities of methicillin-resistant Staphylococcus aureus in Taiwan: emphasis on chlorhexidine susceptibility. Diagn Microbiol Infect Dis. 2009;63(3):309–313. doi:10.1016/j.diagmicrobio.2008.11.014
40. Noguchi N, Suwa J, Narui K, et al. Susceptibilities to antiseptic agents and distribution of antiseptic-resistance genes qacA/B and smr of methicillin-resistant Staphylococcus aureus isolated in Asia during 1998 and 1999. J Med Microbiol. 2005;54(Pt 6):557–565. doi:10.1099/jmm.0.45902-0
41. McNeil JC, Kok EY, Vallejo JG, et al. Clinical and molecular features of decreased chlorhexidine susceptibility among nosocomial staphylococcus aureus isolates at Texas children’s hospital. Antimicrob Agents Chemother. 2016;60(2):1121–1128. doi:10.1128/AAC.02011-15
42. Mahmoudi S, Mamishi S, Mohammadi M, et al. Phenotypic and genotypic determinants of mupirocin resistance among isolates recovered from clinical samples of children: an Iranian hospital-based study. Infect Drug Resist. 2019;12:137–143. doi:10.2147/IDR.S185610
43. Trouillet-Assant S, Flammier S, Sapin A, et al. Mupirocin resistance in isolates of Staphylococcus spp. from Nasal Swabs in a tertiary hospital in France. J Clin Microbiol. 2015;53(8):2713–2715. doi:10.1128/JCM.00274-15
44. Bjorland J, Steinum T, Kvitle B, Waage S, Sunde M, Heir E. Widespread distribution of disinfectant resistance genes among staphylococci of bovine and caprine origin in Norway. J Clin Microbiol. 2005;43(9):4363–4368. doi:10.1128/JCM.43.9.4363-4368.2005
45. Costa SS, Falcão C, Viveiros M, et al. Exploring the contribution of efflux on the resistance to fluoroquinolones in clinical isolates of Staphylococcus aureus. BMC Microbiol. 2011;11:241. doi:10.1186/1471-2180-11-241
46. Opacic D, Lepsanovic Z, Sbutega-Milosevic G. Distribution of disinfectant resistance genes qacA/B in clinical isolates of meticillin-resistant and -susceptible Staphylococcus aureus in one Belgrade hospital. J Hosp Infect. 2010;76(3):266–267. doi:10.1016/j.jhin.2010.04.019
47. Zhang M, O’Donoghue MM, Ito T, Hiramatsu K, Boost MV. Prevalence of antiseptic-resistance genes in Staphylococcus aureus and coagulase-negative staphylococci colonising nurses and the general population in Hong Kong. J Hosp Infect. 2011;78(2):113–117. doi:10.1016/j.jhin.2011.02.018
48. Kaatz GW, McAleese F, Seo SM. Multidrug resistance in Staphylococcus aureus due to overexpression of a novel multidrug and toxin extrusion (MATE) transport protein. Antimicrob Agents Chemother. 2005;49(5):1857–1864. doi:10.1128/AAC.49.5.1857-1864.2005
49. Kernberger-Fischer IA, Krischek C, Strommenger B, et al. Susceptibility of methicillin-resistant and -susceptible Staphylococcus aureus isolates of various clonal lineages from Germany to eight biocides. Appl Environ Microbiol. 2018;84(13). doi:10.1128/AEM.00799-18
50. Batra R, Cooper BS, Whiteley C, Patel AK, Wyncoll D, Edgeworth JD. Efficacy and limitation of a chlorhexidine-based decolonization strategy in preventing transmission of methicillin-resistant Staphylococcus aureus in an intensive care unit. Clin Infect Dis. 2010;50(2):210–217. doi:10.1086/648717
51. Mah TF, O’Toole GA. Mechanisms of biofilm resistance to antimicrobial agents. Trends Microbiol. 2001;9(1):34–39. doi:10.1016/S0966-842X(00)01913-2
52. Sharabiani HR, Sadeghi J, Pirzade T, et al. Comparison of superantigens and attachment factors genes of Staphylococcus aureus in clinical isolates and nasal colonizers in the same patients. Microb Pathog. 2021;154:104860. doi:10.1016/j.micpath.2021.104860
53. Seah C, Alexander DC, Louie L, et al. MupB, a new high-level mupirocin resistance mechanism in Staphylococcus aureus. Antimicrob Agents Chemother. 2012;56(4):1916–1920. doi:10.1128/AAC.05325-11
54. Barakat GI, Nabil YM. Correlation of mupirocin resistance with biofilm production in methicillin-resistant Staphylococcus aureus from surgical site infections in a tertiary centre, Egypt. J Glob Antimicrob Resist. 2016;4:16–20. doi:10.1016/j.jgar.2015.11.010
 © 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Associated Outcomes of Different Intravenous Antibiotics Combined with 2% Mupirocin Ointment in the Treatment of Pediatric Patients with Staphylococcal Scalded Skin Syndrome
You C, Wu Z, Liao M, Ye X, Li L, Yang T
Clinical, Cosmetic and Investigational Dermatology 2023, 16:1691-1701
Published Date: 28 June 2023
Cross-sectional Hospital-based Investigation on Clinical Characteristics of Pediatric Staphylococcus aureus Isolates in a Beijing Hospital from 2013 to 2022
Wang L, Zhen JH, Dong F, Lyu ZY
Infection and Drug Resistance 2024, 17:4899-4912
Published Date: 6 November 2024
Synergistic Anti-Biofilm and Bactericidal Activity of Ethanol and Chlorhexidine Combined with EDTA Against Staphylococcus aureus Isolates from Healthcare-Associated Carriers
Mohammadi M, Nikkhahi F, Charkhchian M, Kiaheyrati N, Javadi A, Fard Sanei A, Fardsanei F
Infection and Drug Resistance 2026, 19:580129
Published Date: 15 April 2026