")
Back to Journals » Infection and Drug Resistance » Volume 14
Distribution of β-Lactamase Genes and Genetic Context of blaKPC-2 in Clinical Carbapenemase-Producing Klebsiella pneumoniae Isolates
Authors Liu H , Lin H, Sun Z, Zhu X, Zhang X, Li Q, Lu J, Lin X, Lin L, Li K, Zhu M, Bao Q, Xu T, Hu Y, Zhang H
Received 6 November 2020
Accepted for publication 9 January 2021
Published 26 January 2021 Volume 2021:14 Pages 237—247
DOI https://doi.org/10.2147/IDR.S290434
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Suresh Antony
Hongmao Liu,1– 3,* Hailong Lin,1– 3,* Zhewei Sun,2,3 Xinyi Zhu,1– 3 Xueya Zhang,1– 3 Qiaoling Li,1– 3 Junwan Lu,2,3 Xi Lin,2,3 Li Lin,1,3 Kewei Li,2,3 Mei Zhu,4 Qiyu Bao,2,3 Teng Xu,5 Yunliang Hu,1– 3 Hailin Zhang1,3
1The Second Affiliated Hospital and Yuying Children’s Hospital, Wenzhou Medical University, Wenzhou 325027, People’s Republic of China; 2Key Laboratory of Medical Genetics of Zhejiang Province, Key Laboratory of Laboratory Medicine, Ministry of Education, China, School of Laboratory Medicine and Life Sciences, Wenzhou Medical University, Wenzhou 325035, People’s Republic of China; 3Institute of Biomedical Informatics, School of Laboratory Medicine and Life Sciences, Wenzhou Medical University, Wenzhou 325035, People’s Republic of China; 4Department of Clinical Laboratory, Zhejiang Hospital, Hangzhou, Zhejiang 310013, People’s Republic of China; 5Institute of Translational Medicine, Baotou Central Hospital, Baotou 014040, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Yunliang Hu; Hailin Zhang
The Second Affiliated Hospital and Yuying Children’s Hospital, Wenzhou Medical University, Wenzhou 325027, People’s Republic of China
Tel/Fax +86-577-86699398
; Tel +86-577-88002134
Email [email protected]; [email protected]
Background: This study was designed to characterize the dissemination mechanism and genetic context of Klebsiella pneumoniae carbapenemase (KPC) genes in carbapenem-resistant Klebsiella pneumoniae (CRKP) isolates.
Methods: A retrospective analysis was performed on CRKP strains isolated from a teaching hospital of Wenzhou Medical University during 2015– 2017. Polymerase chain reaction (PCR)-based amplification and whole-genome sequencing (WGS) were used to analyze the genetic context of the blaKPC-2 gene. Conjugation experiments were performed to evaluate the transferability of blaKPC-2-bearing plasmids. Multilocus sequence typing (MLST) and pulsed-field gel electrophoresis (PFGE) were performed to investigate the clonal relatedness of blaKPC-2-producing strains.
Results: The blaKPC-2 gene was identified from 13.61% (40/294) of clinical K. pneumoniae isolates. Three different sequence types (ST11, ST15 and ST656) and 5 PFGE subtypes (A to E) were classified among them. ST11 was the dominant sequence type (92.50%, 37/40). Plasmid-oriented antibiotic resistance genes, such as extended spectrum-β-lactamases (ESBLs) and other antimicrobial resistance genes, were also found in KPC-positive K. pneumoniae (KPC-Kp) isolates. Mapping PCR and genomic sequencing revealed that the blaKPC-2-bearing sequence regions, which are related to different mobile elements, including Tn1721- and IS26-based transposons, were mainly located in but not restricted to IncFII-like plasmids and were structurally divergent.
Conclusion: The blaKPC-2 genes related to divergent mobile genetic elements encoded on transferable plasmids may transfer widely, facilitating the spread of carbapenem resistance among bacteria with different genetic backgrounds. The dissemination of blaKPC-bearing plasmids that collectively carry additional multidrug resistance genes has caused widespread public concern, further limiting the antibiotics available to treat infections caused by KPC-producing pathogens.
Keywords: carbapenemase, CRKP, blaKPC-2, KPC-Kp, ST11, Tn 1721
Introduction
Carbapenems represent a critical class of β-lactam antimicrobials that are used against multidrug-resistant (MDR) gram-negative bacteria, especially those expressing high levels of AmpC cephalosporinases or extended spectrum-β-lactamases (ESBLs).1 Although Klebsiella pneumoniae is considered an opportunistic pathogen, the prevalence of carbapenem-resistant K. pneumoniae (CRKP) associated with severe nosocomial and systemic infections has become an alarming threat to public health in recent years.2 CRKP may serve as a reservoir for antimicrobial resistance because of its propensity to accumulate and transfer ESBLs and other antibiotic inactivating enzymes, mostly in the form of transferable plasmids and transposons, leading to MDR bacterial infections.3
To date, the CRKP pandemic is primarily driven either by the widespread dissemination of clonal complex (CC) 258, more specifically to multilocus sequence type (ST) 258, or the horizontal dissemination of mobile genetic elements, especially insertion sequences and transposons encoded on the plasmids.4,5 ST258 is an international hyperepidemic lineage clone that contributes to the spread of K. pneumoniae carbapenemase (KPC)-positive K. pneumoniae (KPC-Kp) and has disseminated throughout the USA and European countries.6 However, ST11, a single locus variant of ST258, is frequently reported as the dominant clone of CRKP in Asia due to its association with multidrug resistance determinants, hypervirulence and high transmission ability.7
The most common plasmid-carried Ambler class A carbapenemase is KPC, and the blaKPC-2 gene is the main cause of the prolonged outbreak.3 The blaKPC-2 gene is carried frequently within the conserved Tn3 family transposon Tn4401 on transferable plasmids in Europe and the United States.8,9 However, non-Tn4401 mobile elements, especially Tn1721-like transposons among ST11 K. pneumoniae, are mainly responsible for the effective spread of the blaKPC-2 gene in China.10
Despite the description of several novel structures and detailed data obtained in previous research, the genetic process of the conversion that accounts for the genetic differences in the blaKPC-2 gene has not yet been fully elucidated. The mechanism behind the transmission of this gene deserves further exploration. This research aimed to provide a molecular epidemiological investigation and the genetic features of retrospectively collected blaKPC-2-harboring K. pneumoniae in a Chinese tertiary hospital.
Materials and Methods
Bacterial Strains and Antibiotic Susceptibility Testing
Between January 2015 and December 2017, 294 clinical isolates of K. pneumoniae were obtained from a hospital in Wenzhou, China, and identified as K. pneumoniae by a VITEK 2 automated microbiology analyzer (BioMerieux Corporate, Craponne, France). Antimicrobial susceptibility tests were determined by the agar dilution method for 14 antibiotics and by the broth microdilution method for tigecycline, colistin and ceftazidime/avibactam (CAZ/AVI). The results were interpreted by the guidelines of the Clinical and Laboratory Standards Institute (CLSI, 2018).11 For colistin, tigecycline, and CAZ/AVI, the breakpoints were defined according to the guidelines of the European Committee on Antimicrobial Susceptibility Testing (EUCAST) (http://www.eucast.org/).12 Escherichia coli ATCC 25922 and K. pneumoniae ATCC 700603 were used as quality controls. Strains resistant to either or both carbapenems (imipenem and meropenem) tested were classified as CRKP.13 The carbapenemase phenotype was further confirmed by the Carba NP test.14 MDR, extensively drug-resistant (XDR) and pandrug-resistant (PDR) phenotypes were defined according to a previous report.15
PCR Amplification and Sequencing
Bacterial DNA was extracted using the AxyPrep Bacterial Genomic DNA Miniprep Kit (Axygen Scientific, Union City, CA, United States) from a single colony subcultured on MacConkey agar plates at 37°C for 18 hours. Carbapenemase genes (blaKPC, blaGES, blaIMP, blaVIM, blaNDM, and blaOXA-48), plasmid-mediated AmpC β-lactamase genes (blaMOX, blaCIT, blaDHA, blaACC, blaEBC, and blaFOX) and broad-spectrum β-lactamase genes (blaCTX-M, blaTEM, and blaSHV) were analyzed by PCR as previously described.16 PCR amplicons were sequenced and compared with sequences available in the National Center for Biotechnology Information (NCBI) database using BLAST searches. The coding sequences of the outer membrane protein (Omp) genes OmpK35 and OmpK36 were amplified by PCR as previously described.17 The PCR products were sequenced and compared with the reference sequences of K. pneumoniae strain ATCC13883 (NZ_JOOW00000000.1).
Molecular Typing
All CRKP isolates were classified by XbaI (Takara, Dalian, China)-digested pulsed-field gel electrophoresis (PFGE) using a CHEF Mapper System (Bio-Rad) as previously described.13 A phylogenetic dendrogram of PFGE profiles was constructed using UPGMA clustering under appropriate tolerance (2%) settings. Cutoff lines at 85% and 95% were used to delineate PFGE clusters.18 Multilocus sequence typing (MLST) of the K. pneumoniae isolates was performed with PCR, and PCR products of seven housekeeping genes, namely, gapA, infB, mdh, pgi, phoE, rpoB, and tonB, were sequenced.10 All the sequences were compared with information in the MLST database (http://bigsdb.web.pasteur.fr/klebsiella).
Conjugation Experiment
Plasmid conjugation experiments were carried out by filter mating using rifampicin-resistant E. coli EC600 as a recipient.19 The transformants were selected on MacConkey agar supplemented with rifampicin (1,024 mg/L) and meropenem (2 mg/L). Putative transconjugant colonies were selected, and further verifications were performed based on 16S rRNA sequencing and PCR-amplified blaKPC-2 detection of the transconjugants. PCR-based replicon typing was conducted, and replicons were sequenced using previously reported primers.20
Genetic Environment Analysis of the blaKPC-2 Gene
A series of primers based on blaKPC-surrounding sequences were designed. Mapping PCR was performed to compare the genetic context of the blaKPC gene according to previously reported conditions.10,21 When a standard primer pair failed to yield a product, alternative outer primers were used to span the region of variation, and all amplification products obtained were sequenced.
WGS and Bioinformatic Analyses
Genomic DNA was extracted using the aforementioned method. DNA was sequenced by an Illumina HiSeq 2500 and PacBio RS II instrument (Pacific Biosciences) at Personalbio Technology Co., Ltd. (Shanghai, China). The PacBio long reads were initially assembled by Canu v1.622 to obtain contigs of the genome sequences, and then two FASTQ sequence files generated using the Illumina HiSeq 2500 platform were mapped onto the primary assembly to control assembly quality. The potential open reading frames (ORFs) were predicted using Glimmer software (http://ccb.jhu.edu/software/glimmer) and annotated against UniProt/Swiss-Prot. BlastX (https://blast.ncbi.nlm.nih.gov) was used to annotate the predicted protein-coding genes against a nonredundant protein database with an e-value threshold of 1e-5. Annotation of resistance genes was performed using ISfinder, INTEGRALL and ResFinder with default parameters.23
Comparative Genomics Analysis of the blaKPC-2-Bearing Plasmids and blaKPC-2 Gene-Related Regions
The plasmid and chromosome genome sequences used for comparative genomics analysis were downloaded from the NCBI database. The sequences with an identity and coverage of ≥80% with pKPC3020-124 were retained for later ortholog analysis using BlastP and InParanoid.24 A map of the plasmid with GC content and GC skew was drawn using the online CGView Server and local GView 1.7 with a visual interface.23 For the linear comparison of the blaKPC-2 gene-related fragments, wild-type KPC-Kp isolates were mixed, and the plasmids were extracted using a Qiagen Plasmid Mega Kit (Qiagen, Valencia, CA, USA). Sequences containing the blaKPC-2 gene were also obtained from the NCBI nucleotide database using blaKPC-2, Tn1721, ISKpn6, and ISKpn8 as the key words. The results were filtered, and only sequences that contained a complete blaKPC-2 gene and were more than 20 kb in length were retained. Multiple sequence alignments were performed by MAFFT using the 11 kb blaKPC-2 gene-related fragment of pKPC3020-124 as a reference, and the sequences were clustered with an identity of 80%.25 The sequence sharing the greatest similarity to the other sequences in each cluster was chosen as the candidate for ortholog analysis.
Nucleotide Accession Number
The nucleotide sequences reported in this paper have been deposited in the GenBank nucleotide database, and the accession numbers for the genome sequences of K. pneumoniae KP3020 are CP061354 (chromosome), CP061355 (pKPC3020-124), CP061356 (pKPC3020-54), CP061357 (pKPC3020-4) and CP061358 (pKPC3020-2), while CP061347, CP061346 and CP061348 were for the blaKPC-2-bearing plasmid pKPC1880 and sequences contig1846 and 41_pilon, respectively.
Results
Prevalence of Carbapenemase and Distribution of the β-Lactamase Genes
Ten β-lactamase genes were predicted through the sequencing of the pooled genomic DNA of 294 strains and were confirmed by the PCR screening method. Regardless of the intrinsic genes blaSHV, blaOKP, and blaLEN encoded in the chromosome, blaCTX-M-65 and blaTEM-1 were the most prevalent genes, with positive rates of 27.21% and 18.71%, respectively, while blaOXA-10 and blaOXA-1 showed the lowest positive rates of 1.02% and 0.68%, respectively (Table S1). The blaKPC gene was identified in 13.61% (40/294) of the strains, and they all exhibited a carbapenem-resistant phenotype. The sequencing results of the PCR products revealed identical blaKPC-2 alleles (NG_049253) among these carbapenemase-producing K. pneumoniae strains. No other carbapenemase genes were identified in the other two carbapenem nonsusceptible isolates (Table S1). The KPC-Kp isolates also carried at least one other β-lactamase gene, such as the ESBL gene blaCTX-M (90%, 36/40) and narrow-spectrum blaTEM (70%, 28/40) (Figure 1). The blaSHV variants included blaSHV-2 (n=2), blaSHV-12 (n=10), blaSHV-28 (n=1) and blaSHV-11 (n=37), while blaCTX-M-65 and blaTEM-1 were the only variants observed.
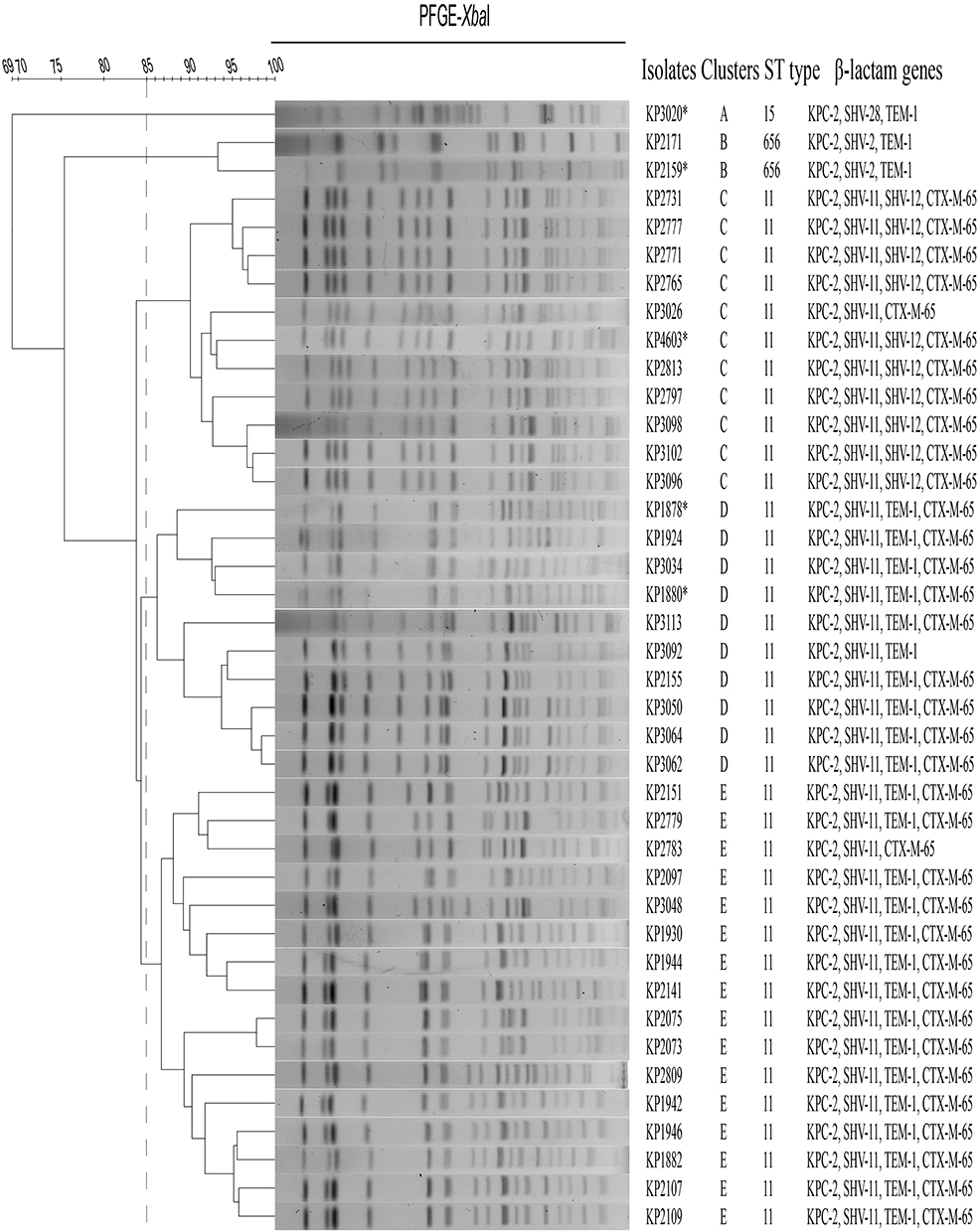 |
Figure 1 Dendrogram of XbaI-digested genomic DNA of 40 KPC-Kp isolates. Five different clusters were identified based on 85% similarity of PFGE profiles. An asterisk (*) indicates that the blaKPC-2 plasmid in the strain was transferable by conjugation. |
Antimicrobial Susceptibility and Conjugation Results of the KPC-Kp Isolates
The minimum inhibitory concentrations (MICs) of 17 antimicrobial agents against all forty KPC-Kp isolates showed multidrug resistance profiles, particularly to β-lactams (including all carbapenems, ertapenem, meropenem and imipenem detected), quinolones and aminoglycosides (Tables 1 and S1). Colistin was shown to be the most active agent (MIC90 = 1 mg/L, 100% susceptible), followed by tigecycline (MIC90 = 8 mg/L, 80% susceptible). The MIC values of aminoglycoside antibiotics were variable: 62.5% (25/40) of isolates showed resistance to gentamicin, and 57.5% (23/40) of isolates showed resistance to both tobramycin and amikacin. Chloramphenicol and fosfomycin exhibited low sensitivity (<20%). On the whole, all isolates were not susceptible to at least one type of agent in three or more antimicrobial categories, while 8 isolates (20%, 8/40) were not susceptible to at least one agent in all categories except colistin, exhibiting MDR and XDR phenotypes. Notably, all isolates were susceptible to CAZ/AVI, except for KP3034, which exhibited low-level resistance to this combination with an MIC of 16/4 mg/L. Sequencing results of the Omp genes amplified from KP3034 revealed that two amino acid duplications (Gly136-Asp137) in the coding sequences of the OmpK36 gene were identified, and no amino acid variation was found in the OmpK35 protein sequence.
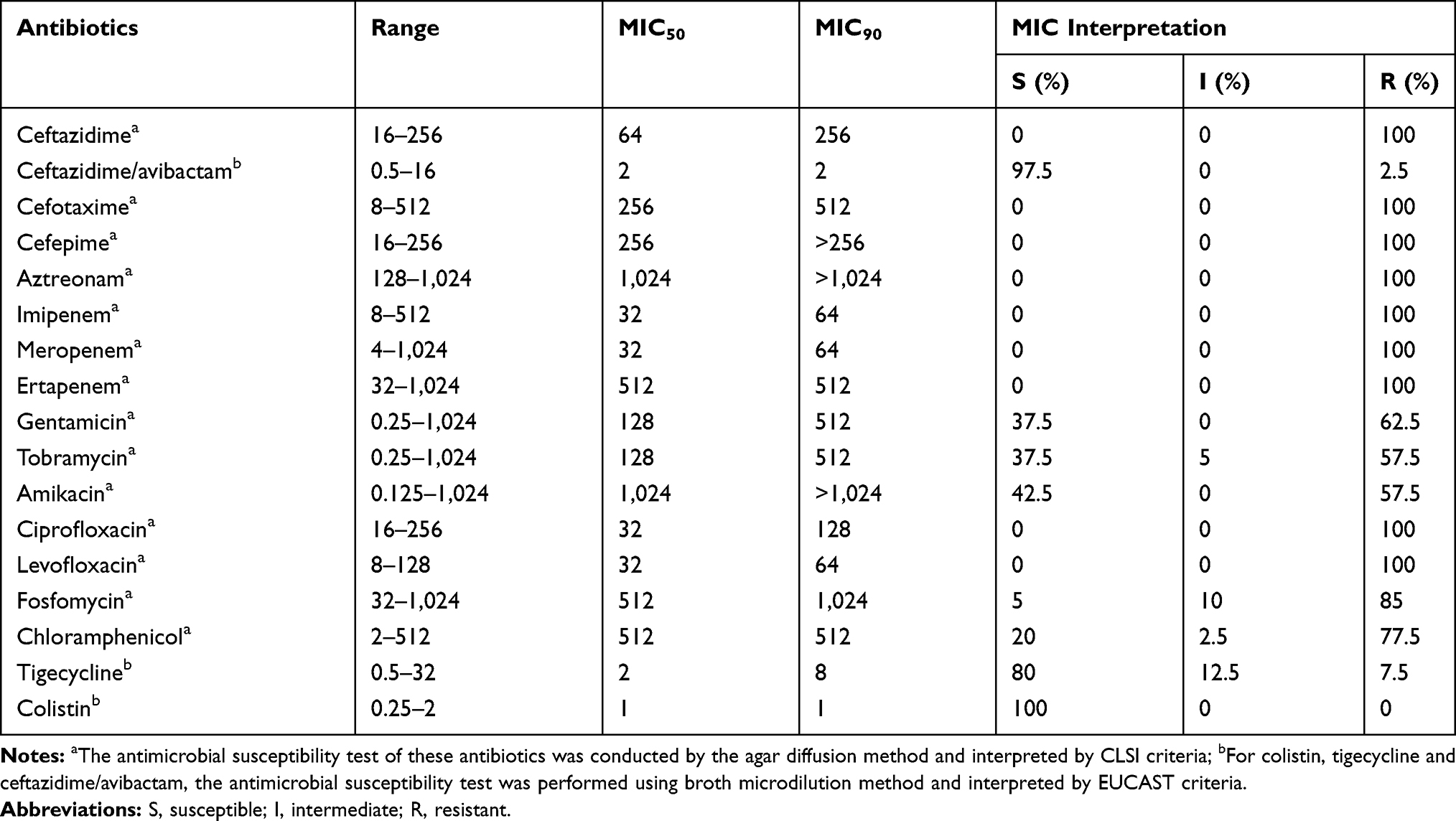 |
Table 1 Antimicrobial Susceptibility Testing Results of Carbapenemase-Producing Klebsiella pneumoniae Isolates from 2015 to 2017 (mg/L) |
According to the conjugation experiment, only 5 of 40 isolates consisting of different ST types successfully transferred carbapenem-resistant plasmids to E. coli EC600 by conjugation. The transconjugants exhibited a phenotype of resistance or reduced susceptibility to carbapenems compared with the recipient E. coli EC600 (Table 2).
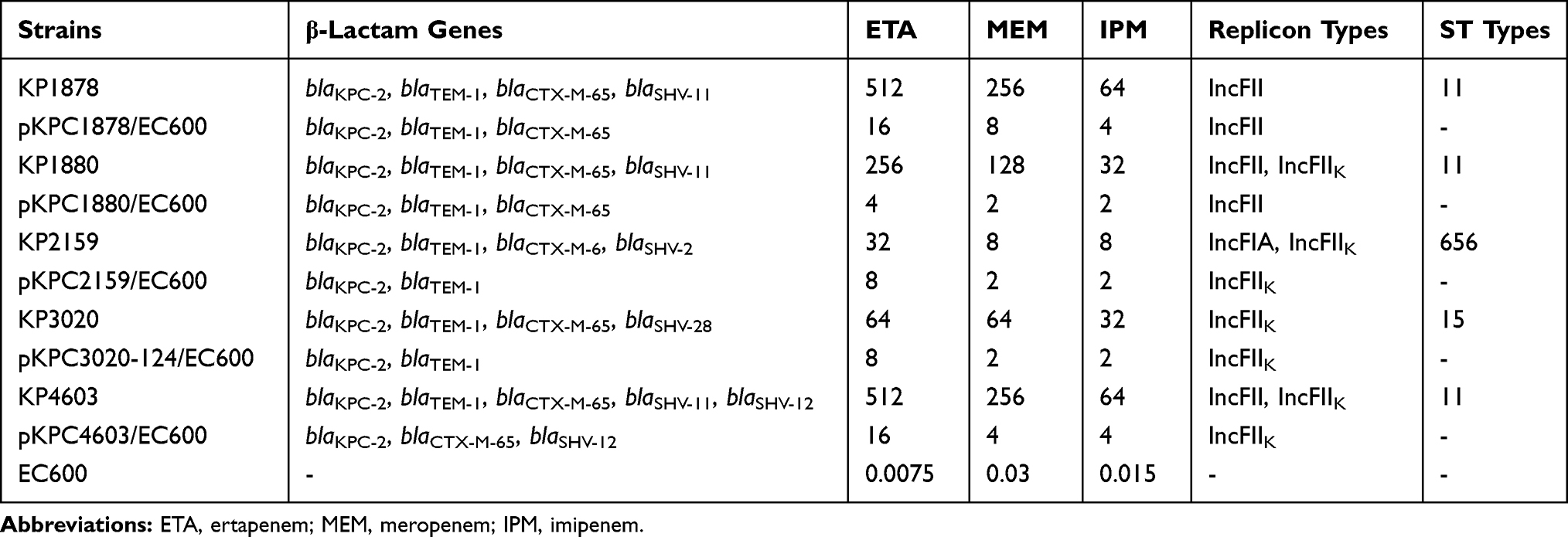 |
Table 2 Resistance Genes and Carbapenem MIC Levels (mg/L) of Wild Strains and Transconjugants |
Genetic Relatedness of the KPC-Kp Isolates
The results of MLST and PFGE analyses divided 40 carbapenemase producers into three different ST types (ST11, ST15 and ST656) and five PFGE pulsotypes (A to E) (Figure 1). ST11 (n=37) was the most predominant sequence type, which consisted of all strains of PFGE pulsotypes C (n=11), D (n=10) and E (n=16), while ST15 (n=1) and ST656 (n=2) consisted of isolates of PFGE pulsotypes A and B, respectively. All the KPC-Kp isolates harbored one or more other β-lactamase genes along with the blaKPC-2 gene, especially blaCTX-M-65 and/or blaTEM-1 in most ST11 K. pneumoniae isolates. The blaTEM-1 gene was not identified in any strains of PFGE cluster C.
Comparative Genomics Analysis of the Plasmid pKPC3020-124 with Homologous Plasmids
Sequencing analysis of the conjugative blaKPC-bearing IncFIIK plasmid pKPC3020-124 from KP3020 showed that the plasmid was 124,452 bp in length with an average GC content of 53.2% and was predicted to encode 152 ORFs. Using the plasmid sequence as a reference query, 15 sequences with the highest similarities (>80% coverage and >80% identity) with pKPC3020-124 were retrieved from the NCBI nucleotide database. Fourteen of them were IncFIIK plasmid sequences, including 2, 10 and 2 sequences from the Enterobacteriaceae species of E. coli, K. pneumoniae and Serratia marcescens, respectively, and one was from the chromosome sequence of K. pneumoniae 91a83dc8-b809-11e8-aae5-3c4a9275d6c8 (Table S2). These blaKPC-bearing plasmids could mainly be divided into two functional regions: a variable region and a conserved region (Figure 2). The conserved region could be further divided into two fragments, one fragment encoding genes related to the type IV secretion system (conjugative transfer region) and the other encoding backbone genes related to plasmid replication and stability (plasmid maintenance region). The variable region is full of insert sequences (such as ISKpn14, IS5075, ISStma11, and ISEc38) and some resistance genes (MDR region), including blaKPC-2, blaTEM-1 and four mercury resistance genes (merACPT). The difference between pKPC3020-124 and the other plasmids is that the former has a unique region of approximately 15.5 kb in length encoding 5 insert sequences and four DNA metabolism enzyme genes (resA, agp, nqrC and lold) (Figure 2, unknown functional region).
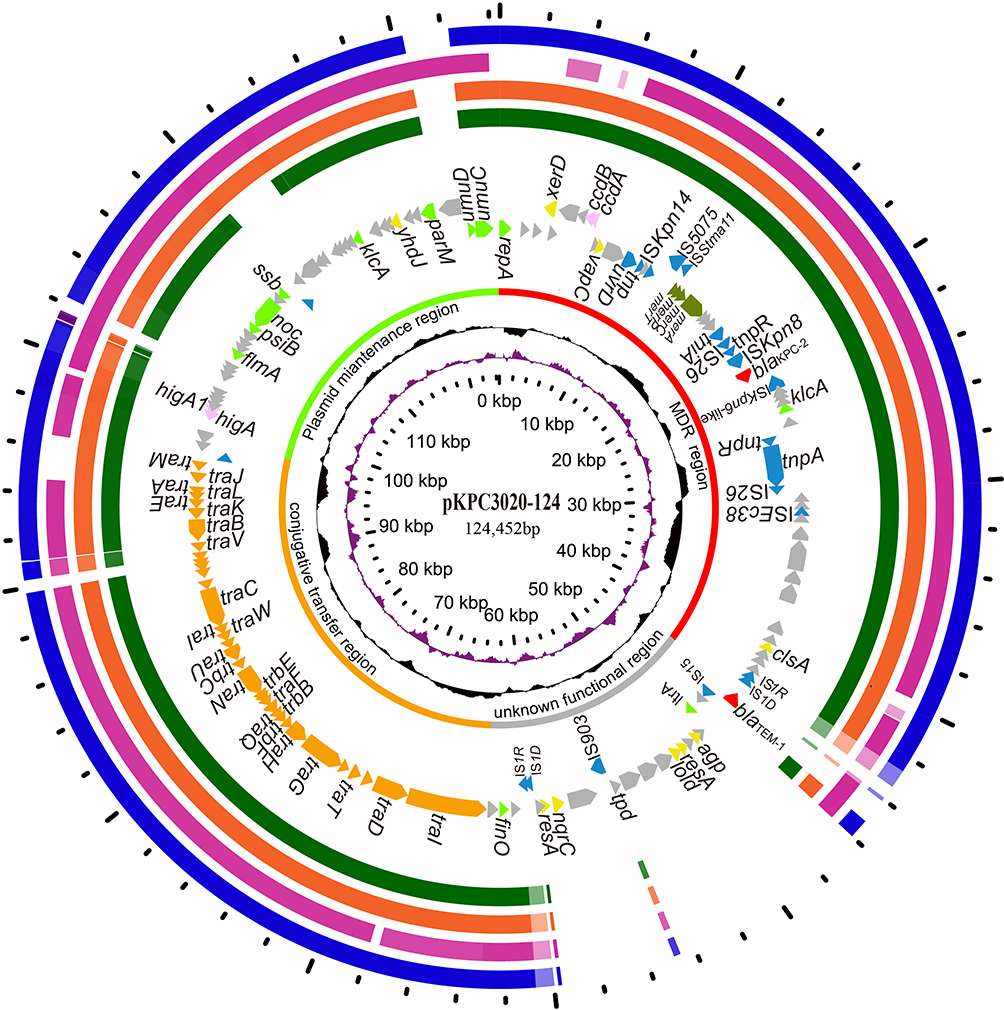 |
Figure 2 Circular map of the plasmid pKPC3020-124 and comparative genomics analysis with its humongous plasmids. Counting from the center toward outside: (1) the scale in bp. (2) GC skew (G-C/G+C), with a positive GC skew toward the outside and a negative GC skew toward the inside. (3) GC content, with an average of 50%, whereby a G+C content of more than 50% is shown toward the outside, otherwise, inward. (4) Functional regions of multidrug resistance, conjugation, maintenance and unknown function. (5) Genes encoded in the leading strand (outwards) or the lagging strand (inwards). The plasmid pKPC3020-124 (CP061355) was used as the reference sequence and was compared to the sequences of (6) pBK34397 (KU295132.1), (7) p628-KPC (KP987218.1), (8) pKPHS2 (CP003224.1) and (9) pS1-KPC2 (MN615880.1). Genes with different functions are shown in different colors: red, antibiotic resistance; blue, mobile genetic elements; orange, transfer conjugation; light green, plasmid stability and replication; brown, heavy metal resistance; pink, toxin-antitoxin system; yellow, DNA metabolism; and gray, hypothetical proteins/genes with unknown functions. |
Comparison of Genetic Environments of blaKPC-2-Encoding Regions
Based on the results of the mapping PCR approach, four divergent forms of blaKPC-2-harboring structures were identified among these carbapenem-resistant plasmids (Table S3). The primers used and their positional counterparts are summarized in Table S4 and Figure S1. The mapping PCR results were confirmed by subsequent sequencing and aligning with WGS and plasmid sequencing results. To analyze the structure of the blaKPC-2-encoding region, all the sequences (including four obtained in this work) of approximately 5–20 kb in length (with the blaKPC-2 gene at the center) were retrieved from the NCBI nucleotide database. According to the similarities of the core sequences adjacent to the blaKPC-2 gene, 10 sequences were chosen as representatives and are illustrated in Figure 3, revealing that an approximately 4-kb fragment encoding tnpR-ISKpn8-blaKPC-2-ΔISKpn6 was conserved and presented in the majority of the sequences. According to whether Tn1721 (or its derivative) was adjacent to the blaKPC-2-encoding fragment, these sequences were roughly categorized into two groups. The first group consisted of the sequences without Tn1721 (or its derivative), while the second group contained Tn1721 (or its derivative) next to the blaKPC-2-encoding fragment. The sequences of the first group existed in various species, such as K. pneumoniae, E. coli, and Citrobacter freundii, while those in the second group were mainly from K. pneumoniae.
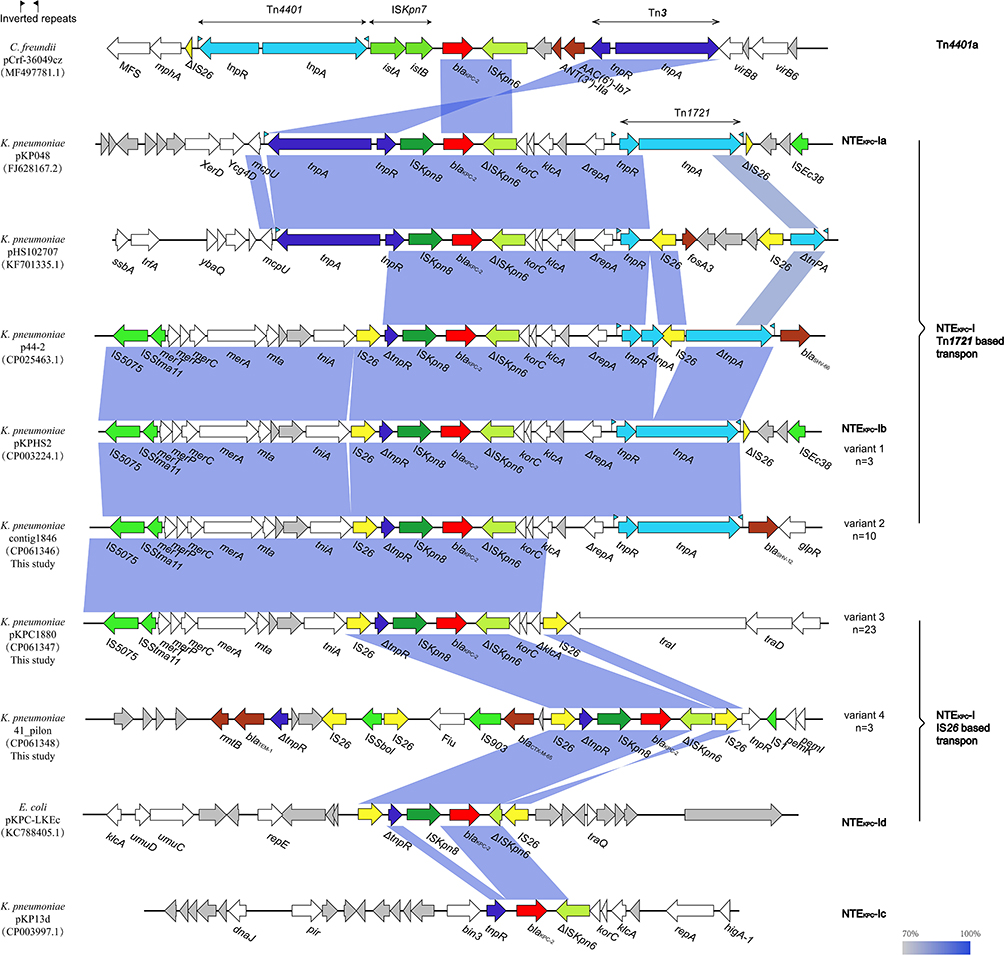 |
Figure 3 Comparison of the blaKPC-2-encoding regions between 10 representative plasmid sequences. The arrows represent the direction of transcription. ORFs are depicted by arrows and are colored based on predicted gene function. Light blue shading denotes homology of regions in each adjacent plasmid. The blaKPC-2 genes are shown in red. |
Discussion
In this study, all KPC-Kp strains were confirmed to be carbapenemase producers and were highly resistant to the most common antimicrobial agents. All isolates showed MDR phenotypes, with 20% of them resistant to agents of all antimicrobial categories tested except for colistin, which could be evaluated as XDR pathogens according to a previous report.15 The data showed that CAZ/AVI displayed potent activity against clinical KPC-Kp isolates with an MIC90 of 2/4 mg/L. Intriguingly, in addition to the high level of carbapenem resistance with a meropenem MIC of 1,024 mg/L, strain KP3034 exhibited a higher MIC level for CAZ/AVI (16/4 mg/L) than the other strains detected. In contrast to the alteration recently reported by Coppi et al26 two-amino-acid duplications (Gly136-Asp137) within transmembrane β-strand loop 3 (L3) of OmpK36 were observed in KP3034, which was previously associated with reduced susceptibility to carbapenems.27 Since there is no history of previous antimicrobial exposure to CAZ/AVI treatment or specific mutations were observed in the blaKPC-2 gene, hyperexpression of the blaKPC-2 gene associated with alteration in the outer membrane or other unknown mechanisms might account for this low level of CAZ/AVI resistance.28 The transconjugants exhibited reduced resistance levels to carbapenems compared with clinical isolates. The copy number of genes and selective expression of promoters in strains of different species or genera,29 restoration of deficient Omp proteins, and antibiotic selection pressure could influence the MIC level of the transconjugants.30
In addition to the blaKPC-2 gene, one or more other kinds of β-lactamase genes (such as blaCTX-M, blaSHV and blaTEM) were identified in these KPC-producing K. pneumoniae strains, with 90% (36/40) of the strains carrying the ESBL gene blaCTX-M. blaCTX-M-65 identified in this work belongs to the group 9 CTX-M β-lactamase gene, which mediates cephalosporin resistance,31 while blaSHV, a core chromosomal gene in K. pneumoniae, mainly mediates inherent ampicillin resistance.32 CTX-M-65 is a variant of CTX-M-14 that differs by 2 amino acid substitutions (Ala78Val and Ser273Arg),33 and it was identified in the variable regions of F33:A-:B- plasmids from food-producing animals in China.34 In contrast to blaCTX-M-14 reported by Yang et al14 blaSHV-11, blaCTX-M-65, and blaTEM-1 were shown to be the predominant genotypes. The coexistence of these multiple broad-spectrum β-lactamase genes simultaneously in KPC-Kp confers resistance to 1st-, 2nd- and 3rd-generation cephalosporins, leading to difficulties in treating the corresponding bacterial infections.
MLST results revealed that these 40 KPC-producing K. pneumoniae strains were composed of three ST types (ST11, ST15 and ST656), and most of these strains were ST11 (92.5%, 37/40). Consistent with previous reports, ST11 was the major ST type attributed to the spread of carbapenem resistance in CRKP isolates,12 while ST15 and ST656 appeared sporadically in this study. Different blaSHV variants were found in CRKP strains of different ST types. blaSHV-11 was the predominant subtype in ST11 K. pneumoniae, while blaSHV-2 and blaSHV-28 were restricted to ST656 and ST15 K. pneumoniae in this work. blaSHV-28 was previously found in ST15 K. pneumoniae strains isolated from dogs in a veterinary clinic but produced OXA-48 instead of KPC-type carbapenemase.35 We also found a substantial proportion of ST11 K. pneumoniae from various samples sharing identical PFGE profiles and similar MIC values, indicating nosocomial transmission and outbreaks of ST11-type KPC-Kp in the hospital. ST11 was proven to be a successful clone that contributed to the outbreak of KPC-Kp, leading to severe nosocomial infections.7,36
Our results are consistent with those from previous studies in which the blaKPC-2 gene is associated with the core blaKPC platform (ΔISKpn6-blaKPC-2-ISKpn8) and can be broadly classified as the NTEKPC-II (blaKPC-bearing non-Tn4401 elements type II) group due to the absence of blaTEM between ISKpn8 and blaKPC-2.32,37 Sequencing data revealed that the blaKPC-2 gene was embedded in divergent IncFII-like plasmids, which had a superiority to capture blaKPC-2 by mobile gene elements, resulting in gradual acquisition or accumulation of carbapenem resistance in ST11 K. pneumoniae.38 The Tn1721 transposon was located adjacent to blaKPC-2 in the plasmid pKPC3020-124. Tn1721 was a result of an integration of the Tn3-based transposon and the partial Tn4401 segment reported in the USA and European countries and was confirmed to transport the carbapenem resistance gene blaKPC-2 in China.39 Interestingly, the Tn1721-based transposons were more flexible than expected when IS26 was inserted into Tn1721-tnpA in p44-2,10 and the insertion of the fosA3 gene sandwiched by two IS26 sequences was found in the transferable blaKPC-2-carrying plasmid pHS102707 at the same position,40 resulting in additional fosfomycin resistance (Figure 3). In this study, Tn1721-related blaKPC-2 and blaSHV-12 cocarriers were found in ten KPC-bearing plasmids in strains of PFGE plusotype C (Figure 3, variant 2), and these strains exhibited elevated ceftazidime resistance compared with those with a single blaKPC-2 gene (Table S1). An identical blaKPC-2-bearing region with a Tn1721-like transposon was found in 15 sequences investigated in comparative genomics analysis, suggesting that the blaKPC-2 gene could transfer across different Enterobacteriaceae species, such as E. coli and S. marcescens, through horizontal transfer of mobile genetic elements or conjugative plasmids. Moreover, an identical Tn1721-ISKpn6-blaKPC-2-ISKpn8-ΔTn3-IS26 configuration was observed in conjugative plasmids within ST656 K. pneumoniae strain KP2159 and ST15 strain KP3020. Congruent with previous reports,39 the blaKPC-2-carrying plasmid was transferable, even if not embedded in Tn1721 transposons and was not restricted to a certain ST or PFGE subtype.
Conclusion
In summary, although carbapenemase-producing K. pneumoniae isolates in this study differed in clonal backgrounds, similarities were found in the phenotypic and genotypic profiles of antibiotic resistance, as well as the identical core configuration of the ΔISKpn6-blaKPC-2-ISKpn8 region. Transmission of the blaKPC-2 gene with the assistance of Tn1721- and IS26-based transposons increases the difficulty of clinical use of carbapenems. Effective prevention and control measures should be adopted to control nosocomial infection caused by the spread of CRKP strains in hospitals.
Data Sharing Statement
The datasets used and analyzed during the current study are included in this published article.
Ethics Approval and Consent to Participate
Individual patient data was not involved, and only anonymous clinical residual samples during routine hospital laboratory procedures were used in this study. This study followed the principles stated in the Declaration of Helsinki (https://www.wma.net/policies-post/wma-declaration-of-helsinki-ethical-principles-for-medical-research-involving-human-subjects/) and was approved by the Second Affiliated Hospital and Yuying Children’s Hospital of Wenzhou Medical University.
Acknowledgments
The authors thank all the colleagues and the reviewers who helped with this work.
Author Contributions
All authors made substantial contributions to conception and design, acquisition of data, or analysis and interpretation of data; took part in drafting the article or revising it critically for important intellectual content; agreed to submit to the current journal; gave final approval of the version to be published; and agree to be accountable for all aspects of the work.
Funding
This study was supported by the National Natural Science Foundation of China (81973382, 81960381 and 81700011); the Natural Science Foundation of Zhejiang Province, China (LY19C060002, LQ17H190001 and LQ17H010003); the Special Project for Significant New Drug Research and Development in the Major National Science and Technology Projects of China (2020ZX09201002); the Science & Technology Project of Wenzhou City, China (Y2020112, Y20170205 and 2019Y0358) and the Science & Technology Project of Inner Mongolia Autonomous Region, China (201802125).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Kitchel B, Rasheed JK, Endimiani A, et al. Genetic factors associated with elevated carbapenem resistance in KPC-producing Klebsiella pneumoniae. Antimicrob Agents Chemother. 2010;54(10):4201–4207. doi:10.1128/aac.00008-10
2. Brandt C, Viehweger A, Singh A, et al. Assessing genetic diversity and similarity of 435 KPC-carrying plasmids. Sci Rep. 2019;9(1):11223. doi:10.1038/s41598-019-47758-5
3. Nordmann P, Cuzon G, Naas T. The real threat of Klebsiella pneumoniae carbapenemase-producing bacteria. Lancet Infect Dis. 2009;9(4):228–236. doi:10.1016/s1473-3099(09)70054-4
4. Nishida S, Ono Y. Genomic analysis of a panresistant Klebsiella pneumoniae sequence type 11 identified in Japan in 2016. Int J Antimicrob Agents. 2020;55(4):105854. doi:10.1016/j.ijantimicag.2019.11.011
5. Peirano G, Bradford PA, Kazmierczak KM, et al. Importance of clonal complex 258 and IncF(K2-like) plasmids among a global collection of Klebsiella pneumoniae with bla(KPC). Antimicrob Agents Chemother. 2017;61(4). doi:10.1128/aac.02610-16
6. Chen L, Mathema B, Pitout JD, et al. Epidemic Klebsiella pneumoniae ST258 is a hybrid strain. mBio. 2014;5(3):e01355–14. doi:10.1128/mBio.01355-14
7. Gu D, Dong N, Zheng Z, et al. A fatal outbreak of ST11 carbapenem-resistant hypervirulent Klebsiella pneumoniae in a Chinese hospital: a molecular epidemiological study. Lancet Infect Dis. 2018;18(1):37–46. doi:10.1016/s1473-3099(17)30489-9
8. Cuzon G, Naas T, Nordmann P. Functional characterization of Tn4401, a Tn3-based transposon involved in blaKPC gene mobilization. Antimicrob Agents Chemother. 2011;55(11):5370–5373. doi:10.1128/aac.05202-11
9. Mathers AJ, Cox HL, Kitchel B, et al. Molecular dissection of an outbreak of carbapenem-resistant enterobacteriaceae reveals Intergenus KPC carbapenemase transmission through a promiscuous plasmid. mBio. 2011;2(6):e00204–11. doi:10.1128/mBio.00204-11
10. Fu P, Tang Y, Li G, et al. Pandemic spread of bla((KPC-2)) among Klebsiella pneumoniae ST11 in China is associated with horizontal transfer mediated by IncFII-like plasmids. Int J Antimicrob Agents. 2019;54(2):117–124. doi:10.1016/j.ijantimicag.2019.03.014
11. CLSI. Performance Standards for Antimicrobial Susceptibility Testing. M100-S28. Wayne, PA: Clinical and Laboratory Standards Institute; 2018.
12. Liu L, Feng Y, Tang G, et al. Carbapenem-resistant isolates of the Klebsiella pneumoniae complex in Western China: the common ST11 and the surprising Hospital-specific types. Clin Infect Dis. 2018;67(suppl_2):S263–s265. doi:10.1093/cid/ciy662
13. Ding B, Shen Z, Hu F, et al. In vivo acquisition of Carbapenemase Gene bla(KPC-2) in multiple species of Enterobacteriaceae through horizontal transfer of insertion sequence or plasmid. Front Microbiol. 2016;7:1651. doi:10.3389/fmicb.2016.01651
14. Yang Y, Chen J, Lin D, et al. Prevalence and drug resistance characteristics of carbapenem-resistant Enterobacteriaceae in Hangzhou, China. Front Med. 2018;12(2):182–188. doi:10.1007/s11684-017-0529-4
15. Magiorakos AP, Srinivasan A, Carey RB, et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: an international expert proposal for interim standard definitions for acquired resistance. Clin Microbiol Infect. 2012;18(3):268–281. doi:10.1111/j.1469-0691.2011.03570.x
16. Dallenne C, Da Costa A, Decré D, et al. Development of a set of multiplex PCR assays for the detection of genes encoding important β-lactamases in Enterobacteriaceae. J Antimicrob Chemother. 2010;65(3):490–495. doi:10.1093/jac/dkp498
17. Doumith M, Ellington MJ, Livermore DM, et al. Molecular mechanisms disrupting porin expression in ertapenem-resistant Klebsiella and Enterobacter spp. clinical isolates from the UK. J Antimicrob Chemother. 2009;63(4):659–667. doi:10.1093/jac/dkp029
18. Marques C, Belas A, Aboim C, et al. Evidence of sharing of Klebsiella pneumoniae strains between healthy companion animals and cohabiting humans. J Clin Microbiol. 2019;57(6). doi:10.1128/jcm.01537-18
19. Xu T, Wang J, Ying J, et al. CharacterizationCharacterisation of a class 1 integron associated with the formation of quadruple bla(GES-5) cassettes from an IncP-1β group plasmid in Pseudomonas aeruginosa. Int J Antimicrob Agents. 2018;52(4):485–491. doi:10.1016/j.ijantimicag.2018.07.002
20. Villa L, García-Fernández A, Fortini D, et al. Replicon sequence typing of IncF plasmids carrying virulence and resistance determinants. J Antimicrob Chemother. 2010;65(12):2518–2529. doi:10.1093/jac/dkq347
21. Shen P, Zhang Y, Li G, et al. Characterization of the genetic environment of the blaKPC-2 gene among Klebsiella pneumoniae isolates from a Chinese Hospital. Braz J Infect Dis. 2016;20(4):384–388. doi:10.1016/j.bjid.2016.04.003
22. Koren S, Walenz BP, Berlin K, et al. Canu: scalable and accurate long-read assembly via adaptive k-mer weighting and repeat separation. Genome Res. 2017;27(5):722–736. doi:10.1101/gr.215087.116
23. Chen Q, Zhou W, Qian C, et al. OXA-830, a novel chromosomally encoded extended-spectrum class D β-lactamase in aeromonas simiae. Front Microbiol. 2019;10:2732. doi:10.3389/fmicb.2019.02732
24. Remm M, Storm CE, Sonnhammer EL. Automatic clustering of orthologs and in-paralogs from pairwise species comparisons. J Mol Biol. 2001;314(5):1041–1052. doi:10.1006/jmbi.2000.5197
25. Liu Y, Wu F, Chen Q, et al. Comparative genomics analysis of Raoultella planticola S25 isolated from duck in China, with florfenicol resistance. Comp Immunol Microbiol Infect Dis. 2020;68:101398. doi:10.1016/j.cimid.2019.101398
26. Coppi M, Di Pilato V, Monaco F, et al. Ceftazidime-avibactam resistance associated with increased bla (KPC-3) gene copy number mediated by pKpQIL plasmid derivatives in sequence type 258 Klebsiella pneumoniae. Antimicrob Agents Chemother. 2020;64(4). doi:10.1128/aac.01816-19
27. García-Fernández A, Miriagou V, Papagiannitsis CC, et al. An ertapenem-resistant extended-spectrum-beta-lactamase-producing Klebsiella pneumoniae clone carries a novel OmpK36 porin variant. Antimicrob Agents Chemother. 2010;54(10):4178–4184. doi:10.1128/aac.01301-09
28. Gaibani P, Re MC, Campoli C, et al. Bloodstream infection caused by KPC-producing Klebsiella pneumoniae resistant to ceftazidime/avibactam: epidemiology and genomic characterization. Clin Microbiol Infect. 2020;26(4):
29. Wang L, Fang H, Feng J, et al. Complete sequences of KPC-2-encoding plasmid p628-KPC and CTX-M-55-encoding p628-CTXM coexisted in Klebsiella pneumoniae. Front Microbiol. 2015;6:838. doi:10.3389/fmicb.2015.00838
30. Kong HK, Pan Q, Lo WU, et al. Fine-tuning carbapenem resistance by reducing porin permeability of bacteria activated in the selection process of conjugation. Sci Rep. 2018;8(1):15248. doi:10.1038/s41598-018-33568-8
31. Peirano G, Pitout JDD. Extended-spectrum β-lactamase-producing Enterobacteriaceae: update on molecular epidemiology and treatment options. Drugs. 2019;79(14):1529–1541. doi:10.1007/s40265-019-01180-3
32. Octavia S, Kalisvar M, Venkatachalam I, et al. Klebsiella pneumoniae and Klebsiella quasipneumoniae define the population structure of blaKPC-2 Klebsiella: a 5 year retrospective genomic study in Singapore. J Antimicrob Chemother. 2019;74(11):3205–3210. doi:10.1093/jac/dkz332
33. Lee SG, Jeong SH, Lee H, et al. Spread of CTX-M-type extended-spectrum beta-lactamases among bloodstream isolates of Escherichia coli and Klebsiella pneumoniae from a Korean hospital. Diagn Microbiol Infect Dis. 2009;63(1):76–80. doi:10.1016/j.diagmicrobio.2008.09.002
34. Wang J, Zeng ZL, Huang XY, et al. Evolution and comparative genomics of F33: a-:B-plasmids carrying bla(CTX-M-55) or bla(CTX-M-65) in Escherichia coli and Klebsiella pneumoniae isolated from animals, food products, and humans in China. mSphere. 2018;3(4). doi:10.1128/mSphere.00137-18
35. Stolle I, Prenger-Berninghoff E, Stamm I, et al. Emergence of OXA-48 carbapenemase-producing Escherichia coli and Klebsiella pneumoniae in dogs. J Antimicrob Chemother. 2013;68(12):2802–2808. doi:10.1093/jac/dkt259
36. Yang J, Ye L, Guo L, et al. A nosocomial outbreak of KPC-2-producing Klebsiella pneumoniae in a Chinese hospital: dissemination of ST11 and emergence of ST37, ST392 and ST395. Clin Microbiol Infect. 2013;19(11):E509–15. doi:10.1111/1469-0691.12275
37. Chen L, Mathema B, Chavda KD, et al. Carbapenemase-producing Klebsiella pneumoniae: molecular and genetic decoding. Trends Microbiol. 2014;22(12):686–696. doi:10.1016/j.tim.2014.09.003
38. Bi D, Jiang X, Sheng ZK, et al. Mapping the resistance-associated mobilome of a carbapenem-resistant Klebsiella pneumoniae strain reveals insights into factors shaping these regions and facilitates generation of a ‘resistance-disarmed’ model organism. J Antimicrob Chemother. 2015;70(10):2770–2774. doi:10.1093/jac/dkv204
39. Tang Y, Li G, Liang W, et al. Translocation of Carbapenemase Gene bla(KPC-2) both internal and external to transposons occurs via novel structures of Tn1721 and exhibits distinct movement patterns. Antimicrob Agents Chemother. 2017;61(10). doi:10.1128/aac.01151-17
40. Li G, Zhang Y, Bi D, et al. First report of a clinical, multidrug-resistant Enterobacteriaceae isolate coharboring fosfomycin resistance gene fosA3 and carbapenemase gene blaKPC-2 on the same transposon, Tn1721. Antimicrob Agents Chemother. 2015;59(1):338–343. doi:10.1128/aac.03061-14
 © 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.