")
Back to Journals » Infection and Drug Resistance » Volume 15
Detection of Two Copies of a blaNDM-1-Encoding Plasmid in Escherichia coli Isolates from a Pediatric Patient with Diarrhea
Authors Jiang BW, Ji X, Lyu ZQ, Liang B, Li JH , Zhu LW, Guo XJ, Liu J, Sun Y , Liu YJ
Received 5 November 2021
Accepted for publication 11 January 2022
Published 26 January 2022 Volume 2022:15 Pages 223—232
DOI https://doi.org/10.2147/IDR.S346111
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. Héctor Mora-Montes
Bo-Wen Jiang,1,2,* Xue Ji,1,2,* Zhong-Qing Lyu,3 Bing Liang,1,2 Jian-Hang Li,3 Ling-Wei Zhu,1,2 Xue-Jun Guo,1,2 Jun Liu,1,2 Yang Sun,1,2 Yan-Jing Liu3
1Changchun Veterinary Research Institute, Chinese Academy of Agricultural Sciences, Changchun, Jilin, People’s Republic of China; 2Key Laboratory of Jilin Province for Zoonosis Prevention and Control, Changchun, Jilin, People’s Republic of China; 3Third Affiliated Clinical Hospital to Changchun University of Chinese Medicine, Changchun, Jilin, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Yang Sun; Yan-Jing Liu, Tel +86 431-86986933, Email [email protected]; [email protected]
Purpose: To elucidate the contribution of a transferable plasmid harboring the blaNDM-1 gene in an Escherichia coli clinical isolate to the spread of resistance determinants.
Methods: Nine extended-spectrum β-lactamase-producing E. coli were collected from diarrhea samples from a pediatric patient and genetic linkage was investigated through enterobacteriaceae repetitive intragenic consensus polymerase chain reaction (PCR). Bacterial species were identified by 16s rRNA sequencing, susceptibility testing with the use of a BD PhoenixTM-100 Automated Microbiology System, and assessment of virulence genes by PCR. The transferability of blaNDM-1 in E. coli strain TCM3e1 was confirmed by conjugation experiments. Complete sequencing of E. coli strain TCM3e1 was determined with the PacBio and Illumina NovaSeq platforms and the characteristics were analyzed with bioinformatics software.
Results: The results showed that all nine E. coli strains were the same clone. E. coli strain TCM3e1 was resistant to 12 antimicrobial agents and carried the virulence gene EAST-1. Conjugation transfer analysis showed that blaNDM-1 was carried on a self-transmissible plasmid. Two copies of the blaNDM-1 gene were present on an IncC plasmid and some resistance genes with two or three copies were located downstream of the blaNDM-1 gene and formed a tandem repeat fragment (blaDNM-1-bleo-sul1- aadA17- dfrA12).
Conclusion: A transmissible plasmid harboring two copies of the blaNDM-1 gene, including clonal dispersions of the blaNDM-1 gene, was identified in clinical isolates. These findings emphasized the necessity of surveillance of the plasmid-borne blaNDM-1 to prevent dissemination.
Keywords: diarrheagenic Escherichia coli, multi-drug-resistant, New Delhi metallo-β-lactamase, blaDNM-1 gene
Introduction
Diarrhea is a severe public health problem and a leading cause of morbidity and mortality in infants and young children.1,2 Severe diarrhea is among the most common reasons for hospital admissions of children in low- and middle-income countries in Africa, Asia, and Latin America. Over the past decade, there has been a substantial reduction in deaths due to diarrhea, yet diarrheal diseases account for nearly 10% of the 7.6 million deaths of children younger than 5 years annually.3 Diarrhea is a multifactorial illness associated with wide spectrum of pathogens, including viruses, bacteria, and parasites. Among the bacterial pathogens, diarrheagenic Escherichia coli is an important cause of endemic and epidemic diarrhea worldwide,4 as strains have evolved by horizontal gene transfer of a particular set of characteristics that have successfully persisted in the host.5,6 Importantly, diarrheal patients are more prone to potentially fatal infections of carbapenem-resistant E. coli.
Carbapenems are important therapeutic agents for the treatment of infections of extended-spectrum β-lactamase (ESBL)-producing isolates.7 Bacteria that possess the New Delhi metallo-β-lactamase (NDM) enzyme, the main carbapenemase detected in children,8 are resistant to almost all β-lactams antibiotics.9 NDM-1, which first emerged in 2009,10 has become more of a global threat than other metallo-β-lactamases. Mobile-resistance elements, such as plasmids, are the main vehicles carrying the blaNDM-1 gene and regarded as major mechanisms driving the dramatically increased prevalence of carbapenemase-resistant Enterobacteriaceae isolates.11,12
In this study, we reported the discovery of an E. coli strain isolated from a diarrhea sample from a pediatric patient carrying a transferable IncC-type plasmid of pNDM-TCM3e1 with two copies of blaNDM-1 and blaCTX-M-61.
Methods
Strain Information and Case History
A 4-month-old boy presented to the Affiliated Hospital of Changchun University of Traditional Chinese Medicine with abdominal pain, diarrhea, coughing, and wheezing accompanied by fever (the highest temperature reaching 38.5°C) for 3 days in June 2019. He received nonstandard antibiotic and antiviral treatment, which included injection of cefixime and the antiviral drug vidarabine monophosphate in another hospital 2 days before admission. On admission, his temperature was 38°C with a white blood cell count of 21.74×109 /L, platelet count of 735×109 /L, 0.34% neutrophils, and 0.52% lymphocytes. The E. coli isolates from fecal samples were extremely resistant to multiple antimicrobial agents. Subsequently, the boy received treatment consisting of an antibiotic (ceftazidime), antiviral (potassium sodium dehydroandrographolide succinate), and traditional Chinese medicine, including an ointment at the acupoint, manipulation, and chiropractic therapy for children for one week. On follow-up, the patient had quickly recovered.
Bacterial Isolates and Identification
Nine E. coli strains were isolated from a fecal specimen collected from the patient with the use of MacConkey agar with no antibiotic or containing 4 mg/L of imipenem or 8 mg/L of cefotaxime. Bacterial species identification was performed by 16s rRNA sequencing and further confirmed with a BD PhoenixTM-100 Automated Microbiology System (Becton, Dickinson and Company, Franklin Lakes, NJ, USA). The genetic linkages of the nine E. coli strains were investigated using the ERIC-PCR technique.13
Antimicrobial Susceptibility Testing and blaNDM Location
Antimicrobial susceptibility testing of 12 antimicrobial agents was performed using E-test strips (bioMérieux Sweden AB, Askim, Sweden) in accordance with the 2019 Performance Standards for Antimicrobial Susceptibility Testing of the Clinical and Laboratory Standards Institute (CLSI; https://clsi.org/). Susceptibility to imipenem was determined by the micro-broth dilution and the breakpoints of the other tested antibiotics were interpreted according to the CLSI-2019 guidelines. E. coli ATCC 25922 served as a quality control strain for susceptibility testing. The presence of key carbapenemase-encoding genes (blaNDM, blaIMP, blaVIM) and ESBL genes (blaSHV, blaTEM, blaCTX-M) were established by PCR and confirmed by DNA sequencing.
Previous reports have confirmed that most antibiotic resistance markers (blaNDM-1, blaTEM, blaCTX-M) are located on plasmids.14–16 The transferability of blaNDM-1 in E. coli strain TCM3e1 was determined by filter-mating using azide-resistant E. coli strain J53 as the recipient. Briefly, the donor and recipient strains were grown to the log-phase in Luria-Bertani (LB) broth and then mixed at a ratio of 1:3 and applied to a 0.22-μm filter. After culturing at 37°C for 16 h, the transconjugants putatively possessing the blaNDM-1-bearing plasmid were screened on LB agar supplemented with imipenem (32 mg/L) and sodium azide (100 mg/L). PCR amplification and sequencing of the transconjugants were subsequently performed to confirm whether the plasmid harboring the antibiotic resistance marker was successfully transferred to the recipient. To determine whether the blaNDM-1 gene was expressed in E. coli strains TCM3e1 and J53 AziR and the transconjugant strain J53-pTCM3e1, testing with imipenem was performed using E-test strips to evaluate the resistance to lactam antibiotics.
Screening of Virulence Genes
The presence of virulence genes and virulence-associated genes, including eae, F4, F5, F6, F18, F41, LT, STa, STb, EAST1, Stx1, Stx2, and Stx2e, in E. coli strain TCM3e1 was investigated by PCR with the use of ExTaq DNA polymerase (TaKaRa Biotechnology (Dalian) Co., Ltd., Dalian, China) and gene-specific primers as described previously.17–19
Genome Sequencing and Component Prediction
Genomic DNA of E. coli strain TCM3e1 was extracted using the sodium dodecyl sulfate method.20 The harvested DNA was separated by agarose gel electrophoresis, quantified with a Qubit® 2.0 Fluorometer (Thermo Fisher Scientific, Waltham, MA, USA), and sequenced using the PacBio Sequel platform (Pacific Biosciences, Menlo Park, CA, USA) and NovaSeq 6000 System (Illumina, Inc., San Diego, CA, USA), which was performed by Beijing Novogene Bioinformatics Technology Co., Ltd. (Beijing, China).
A draft genome was assembled using the single molecule real-time (SMRT) Link v.5.0.1 program (Pacific Biosciences) and then corrected with Illumina data. Next, based on the overlap between the head and tail, the chromosomal sequence formed a circle. Then, the initial site was corrected in reference to the DNA database with the Basic Local Alignment Search Tool (BLAST; https://blast.ncbi.nlm.nih.gov/Blast.cgi). Finally, the chromosome and plasmid sequences were screened against the plasmid database using BLAST. The E. coli coding genes were predicted using the GeneMarkS program (http://opal.biology.gatech.edu/GeneMark/) and interspersed repetitive sequences were predicted using the RepeatMasker program (https://www.repeatmasker.org/). The Tandem Repeats Finder database (https://tandem.bu.edu/trf/trf.html) was searched to identify tandem repeats. The tRNAscan-SE web server (http://lowelab.ucsc.edu/tRNAscan-SE/) was used to predict genes coding for tRNA, while the rRNAmmer server (http://www.cbs.dtu.dk/services/RNAmmer/) was used to identify genes coding for rRNA. Small nuclear RNA was predicted with the use of BLAST against the Rfam database. The IslandPath-DIOMB program was used to predict the genomics islands, and the TransposonPSI tool (http://transposonpsi.sourceforge.net/) was used to identify transposons based on the homologous BLAST method. The PHAge Search Tool was used to identify prophages (http://phast.wishartlab.com/) and clustered regularly interspaced short palindromic repeat sequences (CRISPR) were identified with the CRISPRfinder program (https://crispr.i2bc.paris-saclay.fr/Server/). The online server Multi-Locus Sequence Typing (MLST) v 2.0 (http://www.cbs.dtu.dk/services/MLST-1.3/) was used to determine the sequence type of E. coli strain TCM3e1 with the assembled contigs. The serotype was predicted from the whole-genome sequences of E. coli isolates using the SeroTypeFinder database (https://cge.cbs.dtu.dk/services/SerotypeFinder/).
Gene Function Analysis
Seven databases were used to predict gene functions in this study, which included Gene Ontology (http://geneontology.org/), Kyoto Encyclopedia of Genes and Genomes (http://www.genome.jp/kegg/), Clusters of Orthologous Groups (http://www.ncbi.nlm.nih.gov/COG/), Non-Redundant Protein Database (https://www.ncbi.nlm.nih.gov/refseq/about/nonredundantproteins/), Transporter Classification Database (http://tcdb.org/), and Swiss-Prot (http://www.ebi.ac.uk/uniprot/). A whole genome search was performed against the above seven databases using BLAST. Virulence genes were identified using VirulenceFinder v2.0 software of the Center for Genomic Epidemiology (https://cge.cbs.dtu.dk/services/VirulenceFinder/). Analysis of antimicrobial resistance genes was performed using the Antibiotic Resistance Genes Database (https://ardb.cbcb.umd.edu/) and Comprehensive Antibiotic Research Database (https://card.mcmaster.ca/).
Plasmid Analysis
Plasmid DNA was extracted using a HiSpeed Plasmid Midi kit (Qiagen, Valencia, CA, USA) and sequenced using the NovaSeq 6000 System with 350-bp paired-end reads, which was performed by Beijing Novogene Bioinformatics Technology Co., Ltd. Identification of plasmid incompatibility types was performed with the use of complete plasmid sequences via the PlasmidFinder v2.0 tool (https://cge.cbs.dtu.dk/services/PlasmidFinder/). The plasmid sequences were annotated using the Rapid Annotations using Subsystems Technology server (https://rast.nmpdr.org/) and edited manually. Sequence alignment of blaNDM-1-carrying plasmids was performed using BLASTx (https://blast.ncbi.nlm.nih.gov/Blast.cgi) or the conserved domain database Domain (https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi) and visualized with the application Easyfig v 2.2.3.21 Highly homologous complete plasmid sequences available in the National Center for Biotechnology Information database (https://www.ncbi.nlm.nih.gov/) were aligned using the BLAST Ring Image Generator (BRIG) tool. Circular maps of chromosomes and plasmids were also generated using the BRIG tool.22
Results
Antimicrobial Susceptibility Analysis
The results of the BD PhoenixTM-100 Automated Microbiology System and 16s rRNA sequencing confirmed that nine ESBL-producing E. coli strains shared the same biochemical phenotype and were resistant to 15 classes of antibiotics, thus, considered as multi-drug resistant. The ERIC-PCR results revealed that all nine E. coli strains had the same ERIC pattern (ERIC-types), as shown in Figure S1. Therefore, these strains were uniformly named E. coli strain TCM3e1 in this article. The antimicrobial susceptibility test results determined that E. coli strain TCM3e1 was resistant to multiple antimicrobial agents, including cephalosporins, carbapenems, penicillins, sulfonamides and tetracyclines, and susceptible to quinolones, chloramphenicols, aminoglycosides, and polymyxin according to the CLSI breakpoints (M100-S29) (Table 1).
 |
Table 1 Antimicrobial Susceptibility Testing of E. coli Strain TCM3e1 Isolates and Associated Transconjugants |
Detection of Antibiotic Resistance Genes and Conjugation Experiment
PCR amplification and sequencing confirmed that E. coli strain TCM3e1 harbored the blaNDM-1 and blaTEM genes. Furthermore, the results of the conjugation assays showed that blaNDM-1 was successfully transferred from E. coli strain TCM3e1 to laboratory E. coli strain J53 at a frequency of 10−6, suggesting that blaNDM-1 was located on a self-transmissible plasmid, while transfer of blaTEM-1 was unsuccessful (Figure S2). The transconjugant (designated J53-pTCM3e1) was resistant to both of the tested β-lactams (imipenem and meropenem). Interestingly, strain J53-pTCM3e1 exhibited a resistance profile similar to that of the parental isolate E. coli strain TCM3e1, with the exception of tetracycline (Table 1).
Genomic Sequence Analysis
Assembly of whole-genome sequencing data revealed that E. coli strain TCM3e1 consisted of a 4,994,807-bp circular chromosome and two circular plasmids (Table 2). The chromosome had a GC content of 50.51% and coded for 22 rRNA operons, 88 tRNAs, 61 nRNAs, and 5052 predicted protein coding sequences (CDSs) (Table 2 and Figure 1). MLST analysis showed that E. coli strain TCM3e1 belonged to sequence type 1722 (ST1722) (adk-fumC-gyrB-icd-mdh-purA-recA). Further analysis determined that strain TCM3e1 had three replicon types, IncFIB, IncC, and IncQ, which is uncommon for plasmids associated with blaNDM-1 (Table 2).
 |
Table 2 Genome and Plasmids of E. coli Strain TCM3e1 |
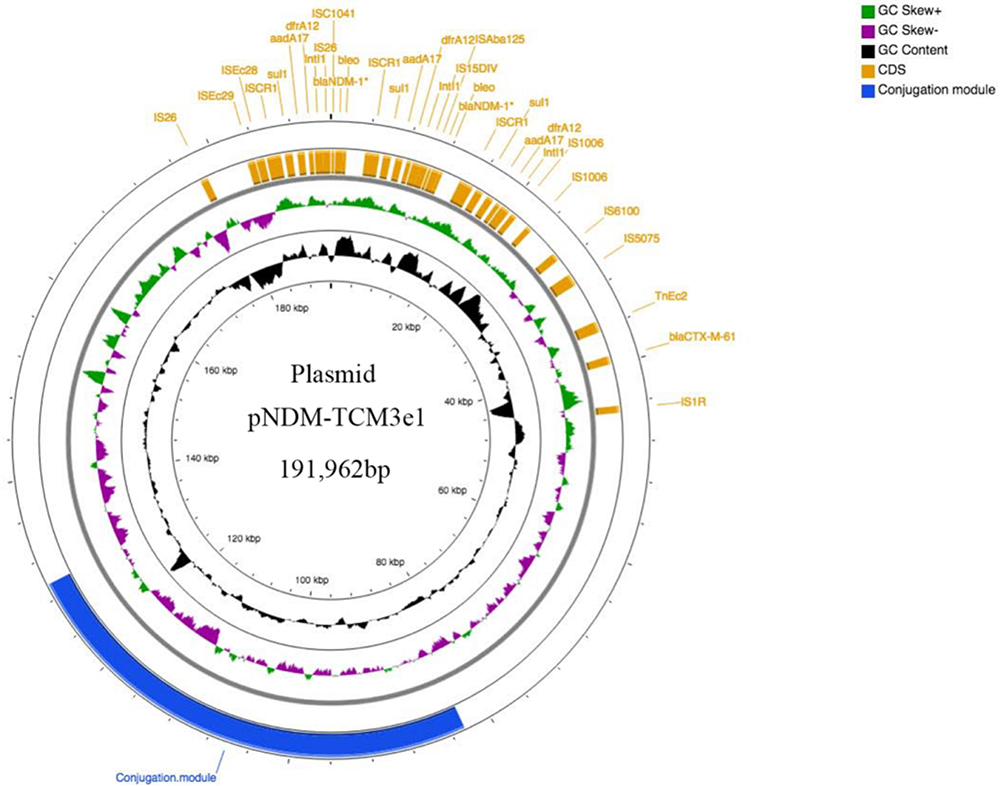 |
Figure 1 Genetic structure of plasmid pNDM-TCM3e1. This circular graph was created to illustrate the backbone and the location of the genetic load region of pNDM-TCM3e1. Genes are denoted by yellow rectangles. The GC skew is depicted as the inner circle in black and the GC content as the outer circle in green and purple. The blue region represents conjugation. |
Plasmid pTCM3e1-1 contained 182 CDSs and sequence analysis revealed query coverages of 47.02–66.97% and sequence identities of 96.59– 98.59% with five plasmids listed in Table 3 at the nucleotide level. The E. coli species harbored all five of the plasmids (Figure S4). Similarly, complete sequence analysis revealed that the 191,962-bp plasmid pTCM3e1-2 contained 254 CDSs. Plasmid pTCM3e1-2 had query coverages of 65.98–87.47% and sequence similarities of 99.22–99.99% with another five plasmids in Table 3. Surprisingly, none of these plasmids, as opposed to plasmid pTCM3e1-2, were derived from E. coli, but rather other species, including Klebsiella pneumoniae, Proteus mirabilis, and Salmonella enterica (Figure S5).
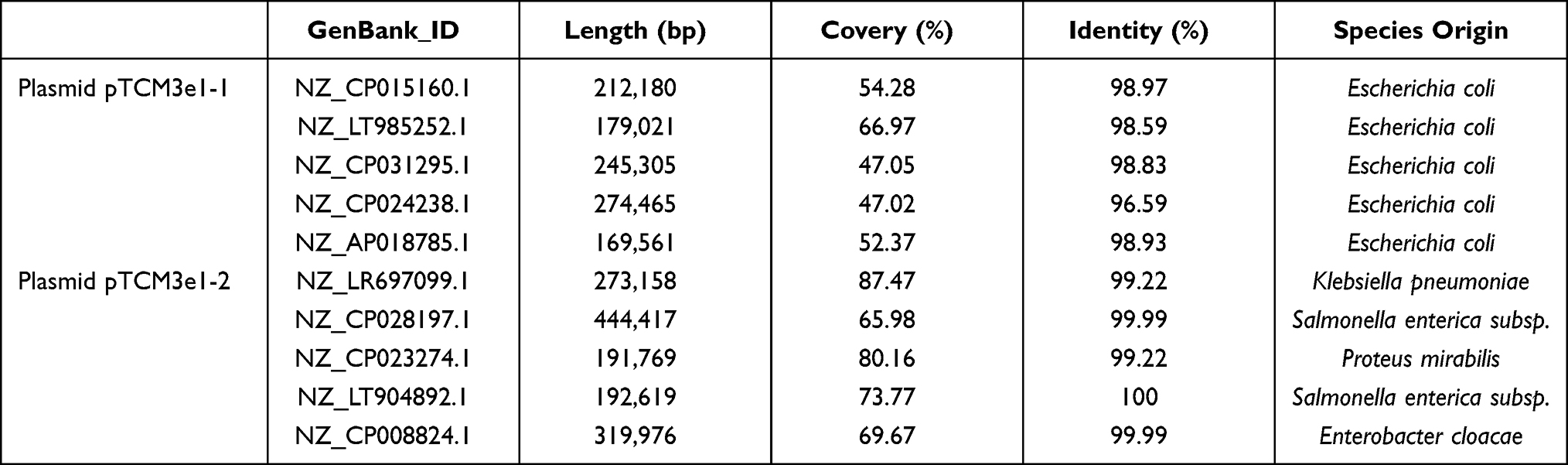 |
Table 3 Match in GenBank of the E. coli Strain TCM3e1 Plasmid |
Resistance Gene Analysis
Multiple resistance genes responsible for resistance to β-lactams, aminoglycosides, polymyxin, fluoroquinolones, sulfonamide, and tetracycline were identified in E. coli strain TCM3e1. Specifically, 13 antimicrobial resistance genes mediating resistance to β-lactams (blaTEM-1, blaCTX-M-105 and blaCMY-62), aminoglycosides [kdpE, aph (3”)-Ib and aph (6)-Id], polymyxin (arnA, PmrB, PmrC and PmrF), fluoroquinolones (mfd), sulfonamide (sul2), and tetracycline (fyuA) were coded by the genomic chromosome, while nine (blaNDM-1, sul1, aadA17, blaCTX-M-61, dfrA12, mphA, Mrx, NmcR, and mphG) were coded by plasmid pTCM3e1-2, and only one β-lactams resistance gene (blaTEM-1) by plasmid pTCM3e1-1 (Table 2). Therefore, the two plasmids were designated as pTEM-TCM3e1 and pNDM-TCM3e1, respectively. Surprisingly, the whole-genome sequencing results revealed that plasmid pNDM-TCM3e1 carried two copies of the blaNDM-1 gene (Figure 1). Additionally, there were two or three copies of the sulfonamide resistance gene sul1, the aminoglycoside resistance gene aadA17, the diaminopyrimidine resistance genes dfrA12, and the bleomycin resistance protein located downstream of the blaDNM-1 gene, which formed a tandem repeat fragment (bla DNM-1-bleo-sul1- aadA17- dfrA12), as shown in Figure 1.
Virulence of E. coli Strain TCM3e1 Isolates
The presence of enteroaggregative E. coli heat-stable enterotoxin 1 (EAST-1) in E. coli strain TCM3e1 was confirmed by PCR and sequencing (Figure S3). Furthermore, the virulence and virulence-related genes were identified with the use of the VirulenceFinder database. The results indicated that E. coli strain TCM3e1 harbored the EAST-1 heat-stable toxin gene astA, which was consistent with the PCR results. In addition, strain TCM3e1 harbored genes encoding regulators of the enteroaggregative immunoglobulin repeat protein (air), afimbrial adhesion (afaD), siderophore receptor (fyuA), heat-resistant agglutinin (bra), outer membrane protein complement resistance (traT), outer membrane hemin receptor (chuA), and long polar fimbriae (lpfA). The plasmid pTEM-TCM3e1 did not code for the afaD and traT genes. Strain TCM3e1 belonged to the serotypes O11:H25, as determined with the SeroTypeFinder database.
Discussion
Globally, diarrheal diseases remain the third leading cause of morbidity and mortality among children under 5-years-old, after pneumonia and preterm birth complications.23 Every day, approximately 2195 children die due to diarrheal diseases, which accounts for one in nine child deaths worldwide.24 According to the “Chinese Clinical Practice Guidelines for Acute Infectious Diarrhea in Children,” the incidence of acute infectious diarrhea in Chinese children under 5-years-old is 2.50–3.38 per person-year. While many etiological agents can cause diarrhea, diarrheagenic E. coli is a major contributor. Importantly, >40% of diarrheal episodes among children are caused by diarrheagenic E. coli pathotypes.25
Nine E. coli strains were isolated from clinical fecal specimens of the patient and were identified with the BD PhoenixTM-100 Automated Microbiology System. The results showed that all of the E. coli isolates had the same biochemical and drug-resistant phenotype, the same ERIC patterns, and carried the same virulence gene. Significantly, all nine E. coli strains harbored the blaNDM-1 and blaTEM-1 genes, as confirmed by PCR amplification. The cloned strains were named E. coli TCM3e1 for use in future studies.
The O11:H25 serotype of E. coli strain TCM3e1 was identified with the use of the SeroTypeFinder database. The results showed that O11:H25 was a non-classical serotype and was difficult to classify as a particular pathotype of diarrheal E. coli. Nevertheless, EAST-1, which was detected in E. coli strain TCM3e1 by PCR and sequencing, is the first toxin characterized for the enteroaggregative E. coli (EAEC) pathotype.26 EAEC has been systematically identified as an emerging enteropathogen that is strongly associated with acute and persistent diarrhea among children younger than 5 years living in developing countries.27 Considering the significant health burden of diarrheagenic E. coli in children, an in-depth study of the dominant strain E. coli TCM3e1 was conducted.
The BD PhoenixTM-100 Automated Microbiology System determined that strain TCM3e1 produced ESBL and was resistant to 15 classes of antibiotics, and thus was considered as multi-drug resistant. Importantly, the E. coli strain TCM3e1 was resistant to all of the tested β-lactam antibiotics (Table 1). Additionally, PCR amplification and genome sequencing confirmed that E. coli strain TCM3e1 harbored the blaNDM-1 and blaTEM-1 genes.
NDM-1 is a broad-spectrum β-lactamase that is capable of hydrolyzing almost all β-lactam antimicrobials. NDM-1-producing Enterobacteriaceae isolates have swept across the globe since first reported in 200910 and the prevalence has dramatically increased recently not only in the United States and European Union, but also Asian countries.12 Many provinces of China, including Beijing,11 Zhejiang,15 and Henan,28 have reported the emergence of Enterobacteriaceae positive for the blaNDM-1 gene. Enterobacteriaceae isolates producing NDM-1-type carbapenemase are often called “super bugs” due to extreme drug resistance.28 Studies have confirmed that this super resistance gene contains genetic determinants that widely exist on mobile genetic elements, such as plasmids, and thus are easily transferred to other organisms, which is also considered to be the main reason for the rapid dissemination of blaNDM-1 into various Enterobacteriaceae.29 Conjugation experiments with azide-resistant E. coli strain J53 were conducted with the use of broth and filters. The result showed that strain TCM3e1 successfully transferred a plasmid carrying blaNDM-1 into recipient E. coli strain J53 at a frequency of 10−6 (transconjugant/recipient) by mating, suggesting that blaNDM-1 was carried on a self-transmissible plasmid. Compared to E. coli strain J53, the transconjugants exhibited significantly increased resistance to carbapenems (Table 1). In addition to blaNDM-1, strain TCM3e1 had other resistance genes, including the ESBL genes blaCTX-M-61 and blaTEM-1, but neither was capable of transfer by conjugation.
To determine the molecular characterization of carbapenemase-resistant NDM-1-producing E. coli strains and the genetic location of blaNDM-1, strain TCM3e1 was subjected to sequencing using the Illumina HiSeq X10 platform. The assembly of whole-genome sequencing data revealed that the blaNDM gene was detected in a 191,962-bp circular plasmid (designated pNDM-TCM3e1) containing an IncC replicon. Studies have confirmed that the IncC plasmid in an important vector for the transmission of drug-resistance genes. In Guangdong, Fujian, and other provinces of China, there have been reports of IncC-type plasmids from E. coli, Salmonella, Klebsiella pneumoniae, and other bacterial isolates in humans and various animals, including pigs, dogs, cattle, chickens, fish, and shrimp. This type of plasmid is the main type of conjugation plasmid carrying the blaNDM and blaCMT genes in China.30–32 Surprisingly, strain TCM3e1 harbored two copies of blaNDM-1, both of which were present on pNDM-TCM3e1. The two blaNDM genes were identical in sequence, but present at different locations on pNDM-TCM3e1. Importantly, there were two or three copies of the class 1 integrin integrase IntI1 with the aadA17–dfrA12 cassette array, sul1 (encoding sulfonamide resistance protein), and bleomycin resistance protein that were all located downstream of the blaDNM-1 gene and formed a tandem repeat fragment (blaDNM-1-bleo-sul1-aadA17-dfrA12), suggesting that this duplicated region may be transferred horizontally among different strains as a whole (Figure 1). Most previous studies have reported only one copy of the blaDNM-1 gene coded by the chromosome or plasmid, as the coexistence of two blaDNM-1 genes has only been described in the chromosomes of an ST167 E. coli strain Y5 in China (GenBank accession no. CP013483)12 and Pseudomonas aeruginosa strain MMA83 (GenBank accession no. HF546976) in Serbia.33 To our knowledge, this is the first report of two blaNDM-1 gene copies coded by a single plasmid. The genetic environment of the blaNDM-1 gene in the plasmid of strain TCM3e1 showed certain similarities to, but was general different from, the nucleotide sequences of the two strains analyzed. As observed in other blaNDM-1 gene environments, ISAba125 insertion sequences were identified immediately upstream of the blaNDM-1 genes (Figure 2), which provides the −35 promoter sequence for the blaNDM-1 gene.34 The genetic organization downstream of the blaNDM-1 genes in strain TCM3e1 is the same as P. aeruginosa strain MMA83, encompassing a bleomycin resistance gene followed by the sul1 gene. In addition, other antibiotic-resistance genes, such as the aminoglycoside resistance gene aadA17 and the diaminopyrimidine resistance gene dfrA12, appeared after the sul1 and the integrin-1 (intl1) genes, which were downstream of the dfrA1 gene. It has been reported that the ISCR1 mobile element was usually associated with intl1, at a point where cassette integration usually occurred.35 ISCR1, which was first reported by Wang et al, is usually neighbored by a wide array of antibiotic resistance genes and exhibits potential for mobilizing adjacent antibiotic-resistance genes.36,37 Considering the important role of the ISCR1 element in the transposition of antibiotic resistance genes, further analysis of the insertion sequence revealed three tandem copies of the ISCR1 element surrounding blaNDM-1, suggesting that blaNDM-1 in E. coli strain TCM3e1 was carried in a similar ISCR1 element in E. coli strain Y5 and P. aeruginosa strain MMA83, although the sequences within the element were significantly different (Figure 2). Additionally, E. coli strain TCM3e1a harbored a composite Tn structure consisting of two insertion sequences (ISCR1 and ISAba125). Given that both ISCR1 and ISAba125 are capable of transfer and the important role played by the ISCR1 element in the genetic transmission of blaNDM-1, future studies are warranted to explore the effect of this insertion region on the transfer efficiency of the pNDM-TCM3e1 plasmid.
 |
Figure 2 Alignment of sequences surrounding the region of blaNDM-1. Genes are denoted by arrowheads and colored based on gene function classification. Red letters are for blaNDM-1. Resistant genes are indicated with red arrowheads. IS elements are indicated in blue. White arrows indicate integron IntI1. The tandem repeat fragment surrounding the blaNDM-1 gene is highlighted with a light grey frame. Abbreviations: blaNDM-1, New Delhi metallo-β-lactamase gene; bleo, bleomycin resistance gene; sul1, sulfonamide resistance gene; aadA17, aminoglycoside 3-N-acetyltransferase ANT-(3”); dfrA1, dihydrofolate reductase; IS, insertion sequence; intl1, integron gene. |
Importantly, the copy number of the blaNDM-1 gene in this strain had no effect on carbapenemase resistance. The minimum inhibitory concentration of imipenem against E. coli strain TCM3e1 carrying two copies of the blaNDM-1 gene was 8 μg/mL, which was equal to or lower than that of strains with a single copy of blaNDM-1.38 Our presumption is that the increased copy number of blaNDM-1 gene in pNDM-TCM3e1 of strain TCM3e1 did not convey higher resistance to carbapenemase as compared with that of a plasmid with a single copy as the result of one or even both of the following reasons (i) two copies of the blaNDM-1 gene were transferred as a block from an ancestor strain and as a nonfunctional multiplication mechanism, and (ii) as a hydrolase, NDM-1 is able to hydrolyze all β-lactam antimicrobials, with the exception of aztreonam, as its activity is not only affected by the expression of the blaNDM-1 genes, but also by substrate or/and enzyme inhibitors, such as metal chelators and Ryukyu compounds. Hence, future studies are needed to elucidate the effects of carbapenemase-resistance and the exact mechanism for the repeated fragment.
Conclusion
This is the first report of an E. coli clinical isolate carrying two copies of the blaNDM-1 gene on a IncC transferable plasmid, although the coexistence of two blaNDM-1 genes on the chromosome was reported and a plasmid coding for one blaNDM-1 gene is already widespread. This study provided new data on the complexity and diversity of genetic features associated with the blaNDM-1 gene, which are of great importance for further analyses of the dissemination routes and mechanisms, including clonal dispersions of the blaNDM-1 gene within clinical isolates.
Accession Number(s)
The complete sequences of the chromosome and two plasmids of E. coli strain TCM3e1 have been deposited in the GenBank database under accession numbers CP066806, CP066807, and CP066808, respectively.
Ethics Statement
The study protocols were approved by the Ethics Committee of Changchun Children’s Hospital (Changchun Traditional Chinese Medicine University, Changchun, China) (Review No.: CCZYFYLL9) and signed informed consent was obtained from the guardians of the patient, which included consent to publish the case details and any accompanying images.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This study was facilitated by special funds from the National Key Research and Development Project of China (Grant agreement 2017YFC1703206).
Disclosure
The authors declare that none have any financial or non-financial conflicts of interest associated with this study.
References
1. Keusch GT, Walker CF, Das JK, Horton S, Habte D. Diarrheal diseases. In: Black RE, Laxminarayan R, Temmerman M, Walker N, editors. Reproductive, Maternal, Newborn, and Child Health: Disease Control Priorities, Third Edition (Volume 2). Washington, DC; 2016.
2. Levine GA, Walson JL, Atlas HE, Lamberti LM, Pavlinac PB. Defining pediatric diarrhea in low-resource settings. J Pediatric Infect Dis Soc. 2017;6:289–293. doi:10.1093/jpids/pix024
3. Walker CLF, Rudan I, Liu L, et al. Global burden of childhood pneumonia and diarrhoea. Lancet. 2013;381:1405–1416. doi:10.1016/S0140-6736(13)60222-6
4. Javadi K, Mohebi S, Motamedifar M, Hadi N. Characterization and antibiotic resistance pattern of diffusely adherent Escherichia coli (DAEC), isolated from paediatric diarrhoea in Shiraz, southern Iran. New Microbes New Infect. 2020;38:100780. doi:10.1016/j.nmni.2020.100780
5. Croxen MA, Law RJ, Scholz R, et al. Recent advances in understanding enteric pathogenic Escherichia coli. Clin Microbiol Rev. 2013;26:822–880. doi:10.1128/CMR.00022-13
6. Gomes TA, Elias WP, Scaletsky IC, et al. Diarrheagenic Escherichia coli. Braz J Microbiol. 2016;47(Suppl 1):3–30. doi:10.1016/j.bjm.2016.10.015
7. Williamson DA, Sidjabat HE, Freeman JT, et al. Identification and molecular characterisation of New Delhi metallo-beta-lactamase-1 (NDM-1)- and NDM-6-producing Enterobacteriaceae from New Zealand hospitals. Int J Antimicrob Agents. 2012;39:529–533. doi:10.1016/j.ijantimicag.2012.02.017
8. Tian D, Pan F, Wang C, Sun Y, Zhang H. Resistance phenotype and clinical molecular epidemiology of carbapenem-resistant Klebsiella pneumoniae among pediatric patients in Shanghai. Infect Drug Resist. 2018;11:1935–1943. doi:10.2147/IDR.S175584
9. Johnson AP, Woodford N. Global spread of antibiotic resistance: the example of New Delhi metallo-beta-lactamase (NDM)-mediated carbapenem resistance. J Med Microbiol. 2013;62:499–513. doi:10.1099/jmm.0.052555-0
10. Yong D, Toleman MA, Giske CG, et al. Characterization of a new metallo-beta-lactamase gene, bla (NDM-1), and a novel erythromycin esterase gene carried on a unique genetic structure in Klebsiella pneumoniae sequence type 14 from India. Antimicrob Agents Chemother. 2009;53:5046–5054. doi:10.1128/AAC.00774-09
11. Feng J, Qiu Y, Yin Z, et al. Coexistence of a novel KPC-2-encoding MDR plasmid and an NDM-1-encoding pNDM-HN380-like plasmid in a clinical isolate of Citrobacter freundii. J Antimicrob Chemother. 2015;70:2987–2991. doi:10.1093/jac/dkv232
12. Shen P, Yi M, Fu Y, et al. Detection of an Escherichia coli sequence type 167 strain with two tandem copies of blaNDM-1 in the chromosome. J Clin Microbiol. 2017;55:199–205. doi:10.1128/JCM.01581-16
13. Sun Y, Song Y, Song H, et al. Complete genome sequence of an Acinetobacter strain harboring the NDM-1 gene. Genome Announc. 2013;1:e0002312. doi:10.1128/genomeA.00023-12
14. Falah F, Shokoohizadeh L, Adabi M. Molecular identification and genotyping of Acinetobacter baumannii isolated from burn patients by PCR and ERIC-PCR. Scars Burn Health. 2019;5:2059513119831369. doi:10.1177/2059513119831369
15. Du XX, Wang JF, Fu Y, et al. Genetic characteristics of blaNDM-1-positive plasmid in Citrobacter freundii isolate separated from a clinical infectious patient. J Med Microbiol. 2013;62:1332–1337. doi:10.1099/jmm.0.057091-0
16. Fu Y, Du X, Ji J, et al. Epidemiological characteristics and genetic structure of blaNDM-1 in non-baumannii Acinetobacter spp. in China. J Antimicrob Chemother. 2012;67:2114–2122.
17. Vidal M, Kruger E, Duran C, et al. Single multiplex PCR assay to identify simultaneously the six categories of diarrheagenic Escherichia coli associated with enteric infections. J Clin Microbiol. 2005;43:5362–5365. doi:10.1128/JCM.43.10.5362-5365.2005
18. Kim YJ, Kim JH, Hur J, Lee JH. Isolation of Escherichia coli from piglets in South Korea with diarrhea and characteristics of the virulence genes. Can J Vet Res. 2010;74:59–64.
19. Jeong YW, Kim TE, Kim JH, Kwon HJ. Pathotyping avian pathogenic Escherichia coli strains in Korea. J Vet Sci. 2012;13:145–152. doi:10.4142/jvs.2012.13.2.145
20. Lim HJ, Lee EH, Yoon Y, Chua B, Son A. Portable lysis apparatus for rapid single-step DNA extraction of Bacillus subtilis. J Appl Microbiol. 2016;120:379–387. doi:10.1111/jam.13011
21. Sullivan MJ, Petty NK, Beatson SA. Easyfig: a genome comparison visualizer. Bioinformatics. 2011;27:1009–1010. doi:10.1093/bioinformatics/btr039
22. Alikhan NF, Petty NK, Zakour NLB, Beatson SA. BLAST Ring Image Generator (BRIG): simple prokaryote genome comparisons. BMC Genomics. 2011;12:402. doi:10.1186/1471-2164-12-402
23. Liu L, Oza S, Hogan D, et al. Global, regional, and national causes of child mortality in 2000-13, with projections to inform post-2015 priorities: an updated systematic analysis. Lancet. 2015;385:430–440. doi:10.1016/S0140-6736(14)61698-6
24. Das JK, Salam RA, Bhutta ZA. Global burden of childhood diarrhea and interventions. Curr Opin Infect Dis. 2014;27:451–458. doi:10.1097/QCO.0000000000000096
25. Thakur N, Jain S, Changotra H, et al. Molecular characterization of diarrheagenic Escherichia coli pathotypes: association of virulent genes, serogroups, and antibiotic resistance among moderate-to-severe diarrhea patients. J Clin Lab Anal. 2018;32:e22388.
26. Savarino SJ, Fasano A, Robertson DC, Levine MM. Enteroaggregative Escherichia coli elaborate a heat-stable enterotoxin demonstrable in an in vitro rabbit intestinal model. J Clin Invest. 1991;87:1450–1455. doi:10.1172/JCI115151
27. Hebbelstrup Jensen B, Olsen KE, Struve C, Krogfelt KA, Petersen AM. Epidemiology and clinical manifestations of enteroaggregative Escherichia coli. Clin Microbiol Rev. 2014;27:614–630. doi:10.1128/CMR.00112-13
28. Qin S, Qi H, Zhang Q, et al. Emergence of extensively drug-resistant Proteus mirabilis harboring a Conjugative NDM-1 plasmid and a novel Salmonella genomic island 1 variant, SGI1-Z. Antimicrob Agents Chemother. 2015;59:6601–6604. doi:10.1128/AAC.00292-15
29. Walsh TR, Weeks J, Livermore DM, Toleman MA. Dissemination of NDM-1 positive bacteria in the New Delhi environment and its implications for human health: an environmental point prevalence study. Lancet Infect Dis. 2011;11:355–362. doi:10.1016/S1473-3099(11)70059-7
30. Yang J, Zhang Z, Zhou X, et al. Prevalence and characterization of antimicrobial resistance in salmonella enterica isolates from retail foods in Shanghai, China. Foodborne Pathog Dis. 2020;17:35–43. doi:10.1089/fpd.2019.2671
31. Li R, Xie M, Liu L, et al. Characterisation of a cointegrate plasmid harbouring blaNDM-1 in a clinical Salmonella Lomita strain. Int J Antimicrob Agents. 2020;55:105817. doi:10.1016/j.ijantimicag.2019.09.021
32. Zhang WJ, Xu XR, Schwarz S, et al. Characterization of the IncA/C plasmid pSCEC2 from Escherichia coli of swine origin that harbours the multiresistance gene cfr. J Antimicrob Chemother. 2014;69:385–389. doi:10.1093/jac/dkt355
33. Jovcic B, Lepsanovic Z, Begovic J, et al. The clinical isolate pseudomonas aeruginosa MMA83 carries two copies of the blaNDM-1 gene in a novel genetic context. Antimicrob Agents Chemother. 2013;57:3405–3407. doi:10.1128/AAC.02312-12
34. Poirel L, Dortet L, Bernabeu S, Nordmann P. Genetic features of blaNDM-1-positive Enterobacteriaceae. Antimicrob Agents Chemother. 2011;55:5403–5407. doi:10.1128/AAC.00585-11
35. Toleman MA, Bennett PM, Walsh TR. ISCR elements: novel gene-capturing systems of the 21st century? Microbiol Mol Biol Rev. 2006;70:296–316. doi:10.1128/MMBR.00048-05
36. Wang F, Wu K, Sun J, et al. Novel ISCR1-linked resistance genes found in multidrug-resistant gram-negative bacteria in southern China. Int J Antimicrob Agents. 2012;40:404–408. doi:10.1016/j.ijantimicag.2012.06.016
37. Wu J, Xie L, Zhang F, Ni Y, Sun J. Molecular characterization of ISCR1-mediated blaPER-1 in a non-O1, non-O139 vibrio cholerae strain from China. Antimicrob Agents Chemother. 2015;59:4293–4295. doi:10.1128/AAC.00166-15
38. Sun Y, Liu Q, Chen S, et al. Characterization and plasmid elimination of NDM-1-producing Acinetobacter calcoaceticus from China. PLoS One. 2014;9:e106555. doi:10.1371/journal.pone.0106555
 © 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.