Back to Journals » Infection and Drug Resistance » Volume 15
Detection of NDM-1-Positive Aeromonas caviae from Bacteremia by Using Whole-Genome Sequencing
Authors Xu S, Tu J, Zhang L, Chen Y, Dong X ![]() , Chi X, Xu H
, Chi X, Xu H
Received 9 February 2022
Accepted for publication 23 May 2022
Published 2 June 2022 Volume 2022:15 Pages 2835—2841
DOI https://doi.org/10.2147/IDR.S360353
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Prof. Dr. Héctor Mora-Montes
Shuguang Xu,1 Jinjing Tu,1 Liang Zhang,1 Yinv Chen,1 Xiaoqi Dong,1 Xiaohui Chi,2,* Hao Xu2,*
1Department of Pulmonary and Critical Care Medicine, Ningbo Medical Center Lihuili Hospital, Ningbo, People’s Republic of China; 2Collaborative Innovation Center for Diagnosis and Treatment of Infectious Diseases, State Key Laboratory for Diagnosis and Treatment of Infectious Diseases, The First Affiliated Hospital, College of Medicine, Zhejiang University, Hangzhou, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Hao Xu; Xiaohui Chi, Email [email protected]; [email protected]
Purpose: Nosocomial infections caused by New Delhi metallo-β-lactamase (NDM)-producing bacteria are prevalent worldwide. However, such diseases caused by NDM-producing Aeromonas caviae had never been reported. Our study aimed to elucidate the genomic characteristics of NDM-1-producing A. caviae isolated from hospital patients.
Methods: Bacterial genomic features and possible origins were assessed by whole-genome sequencing (WGS) and phylogenetic analysis. Subsequent investigations include antimicrobial susceptibility testing and multilocus sequence typing (MLST).
Results: We identified here two NDM-1-producing A. caviae isolates from bacteremia. Susceptibility testing showed that two isolates were multi-drug resistant and shared a similar resistance profile and were only sensitive to amikacin and trimethoprim/sulfamethoxazole. Both A. caviae isolates carry the carbapenem resistance gene blaNDM-1 and also have antibiotic resistance genes such as β-lactams, AmpC enzymes, macrolides, aminoglycosides, and quinolones. S1-PFGE and Southern blot analysis were negative. Whole-genome sequencing and comparative analysis revealed that these two isolates shared a close relationship.
Conclusion: To the best of our knowledge, this work describes the first detection of non-plasmid encoded blaNDM-1 in A. caviae. The A. caviae isolated in this study has a broad drug resistance spectrum. Phenotypic and molecular analysis indicated the two isolates belong to the same clone. Routine genomic surveillance of this species is now necessary to effectively curb the further dissemination of carbapenem-resistant bacteria in the region.
Keywords: Aeromonas caviae, New Delhi metallo-β-lactamase, whole-genome sequencing, SNP, phylogenetic analysis
Introduction
The Genus Aeromonas belongs to the Aeromonadaceae, and it is a group of Gram-negative bacteria widely distributed in the aquatic environment.1,2 However, Aeromonas can cause a wide range of diseases in humans and animals, and the main pathogenic species of clinical relevance are Aeromonas hydrophila and Aeromonas caviae.3 They are emerging opportunistic human pathogens that can cause host wound infections, gastrointestinal infections, and even bacteremia.4,5
The prevalence of carbapenemase-resistant Enterobacteriaceae (CRE) has risen since the 2000s.6 New Delhi metallo-β-lactamase (NDM) is a type of metallo-β-lactamase (MBL) able to hydrolyze most β-lactams (including carbapenems).7,8 Since the isolation of NDM-1-producing Klebsiella pneumoniae strains in India in 2008, NDM-1 has been found in various species of Enterobacteriaceae, Acinetobacter, and Pseudomonas.9,10 Since then, NDM-1 has become widely popular around the world. Successful dissemination of carbapenemase-producing bacteria poses an enormous global public health challenge.11,12
However, there is no detailed genomic information on A. caviae carrying blaNDM-1 has been reported. In this study, two A. caviae strains carrying blaNDM-1 were isolated from patients with sepsis. The two clinically derived strains were further characterized by whole-genome sequencing and phylogenetic analysis. To the best of our knowledge, this is the first report that describes the detection of NDM-1-producing A. caviae.
Materials and Methods
Sample Collection
Two carbapenem-resistant strains, HZ574 and HZ578, were isolated from two female patients with sepsis hospitalized in a tertiary hospital in Huzhou, China. Bacterial species were identified using matrix-assisted laser desorption/ionization time-of-flight mass spectrometry (MALDI-TOF/MS) (Bruker, Bremen, Germany). In addition, High throughput ANI analysis is used to compare the whole genome sequencing results to reveal distinct species of Aeromonadaceae (Figure S1). The carbapenemase genes (blaKPC, blaNDM, blaOXA-48, blaVIM and blaIMP) were identified by polymerase chain reaction (PCR)13 (Table S1).
Antibiotic Susceptibility Testing
Minimum inhibitory concentrations (MICs) were determined by VITEK 2 system with AST-GN16 panel.14 The results were interpreted using European Committee on Antimicrobial Susceptibility Testing breakpoints, version 6.0, and Clinical and Laboratory Standards Institute document M100-S25.
Plasmid Characterization and Conjugation Assay
The number and size of the plasmid of the two isolates were characterized by the S1 Nuclease-Pulsed Field Gel Electrophoresis (S1-PFGE).15 Briefly, DNA plugs were digested using S1 Nase restriction enzyme (Takara Bio Inc., Kyoto, Japan) for 30 minutes. S1-PFGE was undertaken on a CHEF-DR III (Bio-Rad, Hercules, CA, USA) using the following parameters: running time 16 hours, temperature 14 °C, field strength 6 V/cm2, angles 120°, initial pulse time 2.2 s, final pulse time 63.8 s. The location of blaNDM-1 was confirmed by Southern blotting and hybridization with a digoxigenin-labelled blaNDM-1 probe using DIG-High Prime DNA Labeling and Detection Starter Kit II (Roche Diagnostics). Plasmid conjugation experiments were performed by mating E. coli J53/EC600 as the recipient strain. Transconjugants were selected on agar (OXOID, Hampshire, UK) medium at a concentration of 200 mg/L sodium azide and two mg/L meropenem. Finally, MALDI-TOF-MS was used to identify transconjugants, and the target gene was verified by PCR. The location and size of plasmids were characterized by S1-PFGE as described previously.
Whole-Genome Sequencing
Whole-genome sequencing (WGS) was performed by Novo Gene Co., Ltd. Beijing, China, using the Illumina HiSeq (Illumina, San Diego, California) platform for all isolates. Alignment of antimicrobial resistance genes was performed through the ResFinder platform (https://cge.cbs.dtu.dk/services/ResFinder/). Multilocus sequence typing (MLST) was performed on bacteria using the website (https://cge.cbs.dtu.dk/services/MLST/). After annotating the strains using Prokka (rapid prokaryotic genome annotation), the genetic environment of carbapenemase encoding genes was characterized using Easyfig 2.2.3. Virulence genes were identified by blasting the VFDB database (http://www.mgc.ac.cn/VFs/main.htm).
Phylogenetic Reconstruction and Analysis
Thirty complete A. caviae genomes were downloaded from the National Center for Biotechnology Information (NCBI) for phylogenetic analysis (Table S2). Snippy (rapid haploid variant calling and core genome alignment) was used to compare genomic differences between strains.16 The alignment file was filtered from variants with elevated densities of base substitutions as putative recombination events by Gubbins version 2.4.1.17 The filtered core-genome alignment file was used to construct a maximum likelihood tree using FastTree with the GTR+CAT model.
Data Availability
The whole-genome sequences of the A. caviae were submitted to GenBank under the following BioProject numbers: PRJNA799930.
Results
Antibiotic Resistance Signature of A. caviae
The results of the MIC of antibiotics are shown in Table 1. The two A. caviae strains in this study have a broad drug resistance spectrum and belong to multidrug-resistant bacteria. Antibiotics to which bacteria are resistant include amoxicillin/clavulanic acid, piperacillin/tazobactam, ceftazidime, ceftriaxone, cefepime, ertapenem, imipenem, levofloxacin, tetracycline, tigecycline. HZ574 and HZ578 are only sensitive to amikacin and trimethoprim/sulfamethoxazole. Two isolates shared similar antimicrobial profiles.
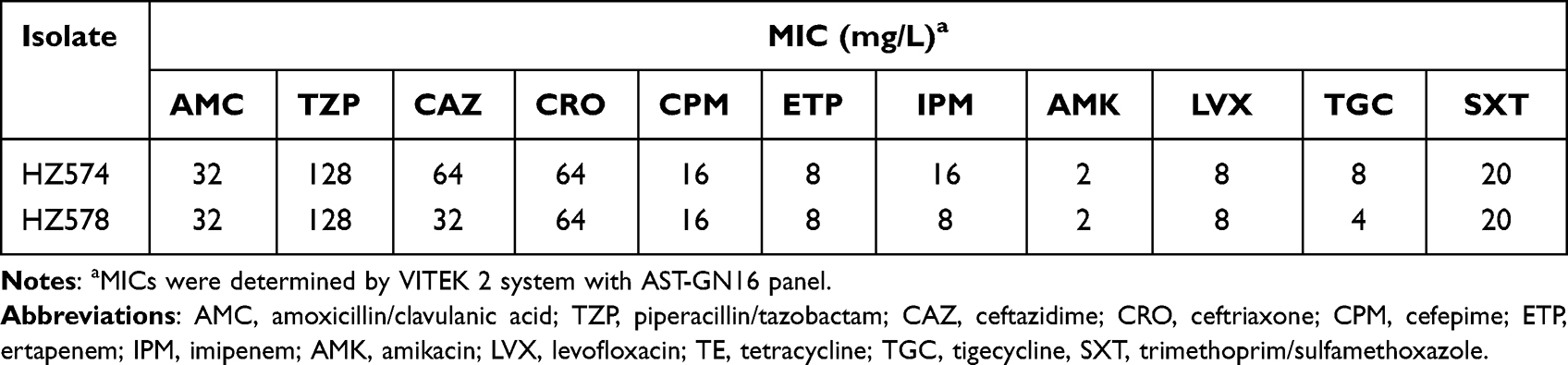 |
Table 1 Antimicrobial Susceptibility Profiles of NDM-Producing Aeromonas caviae of Isolates |
Antimicrobial Resistance Genes
We provide data on antimicrobial resistance genes in Figure 1. Both HZ574 and HZ578 carry the carbapenem resistance gene blaNDM-1, which is the first reported A. caviae carrying blaNDM-1 so far. In addition, bacteria also carry antibiotic resistance genes such as β-lactams, AmpC enzymes, macrolides, aminoglycosides, quinolones, etc. (Figure 1). We also compared the resistance genes carried by 30 A. caviae strains downloaded from NCBI, and the results showed that 30 A. caviae strains mainly had AmpC enzyme (86.21%), followed by β-lactamase (24.14%) (Figure 1).
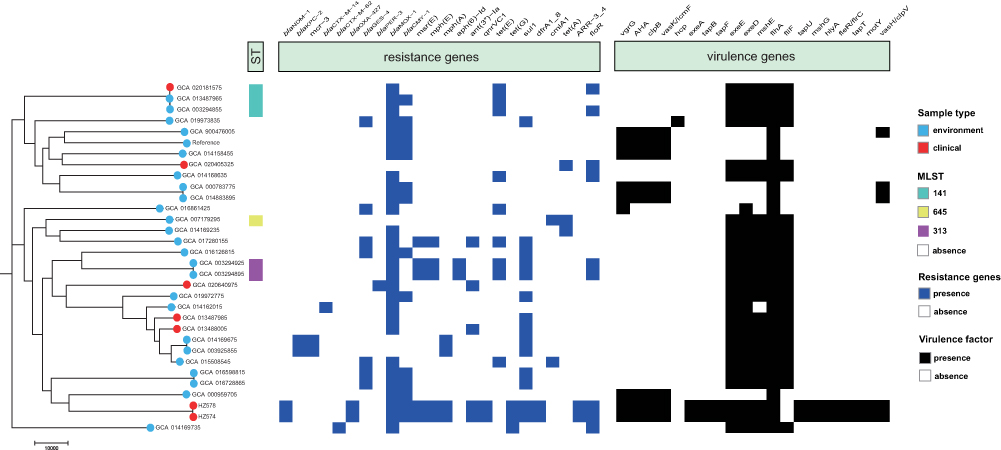 |
Figure 1 Construction of phylogenetic trees of A. caviae. The figure includes the sample source of the isolate, MLST, antibiotic resistance gene, and the comparison result of the virulence gene. |
Virulence Genes
The results of the alignment of virulence genes are shown in Figure 1. The virulence genes carried by HZ574 and HZ578 are related genes of the Type VI secretion system, Tap type IV pili, Polar flagella, twitching ATPase, and Hemolysin A. The strains downloaded from NCBI mainly carried secretion pathway protein (83.33%) and related genes of Polar flagella (100%).
Molecular Characteristics of A. caviae
MLST typing analysis of 32 A. caviae strains showed that only six strains had a known ST type, including ST141, ST313, and ST645 (Figure 1). S1-PFGE and Southern blot demonstrated that the blaNDM-1 gene is not carried by plasmid in isolates HZ574 and HZ578 (Figure 2). Moreover, two isolates were both negative for genes of plasmid typing replicon. These results indicated that blaNDM-1 gene is chromosome encoded in these two isolates. Phylogenetic analysis of all the strains showed that HZ574 and HZ578 were located in the same clade and had a close relationship with GCA_000959705 from Brazil (Figure 1). Moreover, the blaNDM-1 genes of HZ574 and HZ578 share the same genetic environment (Figure 3).
 |
Figure 2 S1 Nuclease-pulsed field gel electrophoresis (S1-PFGE) and southern blot. The number and size of the plasmid of the two isolates were characterized by the S1 Nuclease-pulsed field gel electrophoresis (S1-PFGE). |
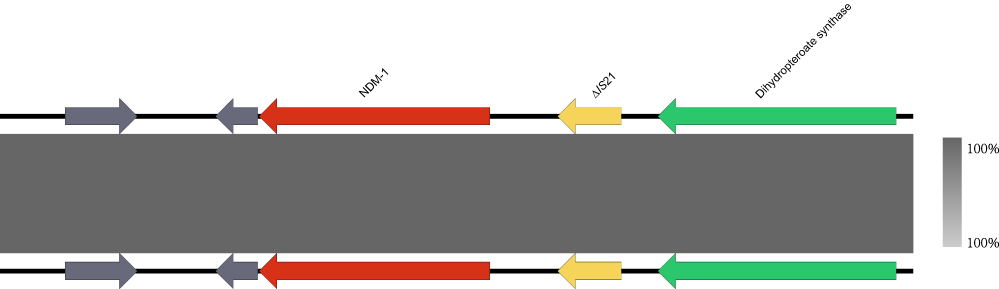 |
Figure 3 The genetic environment of the blaNDM-1 gene in A. caviae was isolated from clinical sources. The arrows represent the direction of transcription. The red open reading frame (ORF) indicates the blaNDM-1 gene, the yellow ORF indicates the mobile element, the green ORF indicates enzymes, and the grey ORF indicates other genes or genes of unknown function. |
Discussion
With the widespread of carbapenemase-producing bacteria, blaNDM-1 has been detected in various bacteria worldwide, but there is no detailed genomic information on A. caviae carrying blaNDM-1 has been reported.10,18 In this study, two strains of A. caviae carrying NDM-1 were isolated from patients. The two clinically derived strains were characterized by genome and phylogenetic analysis using whole-genome sequencing technology.
The study results on the resistance of A. caviae showed that A. caviae is resistant to penicillin and first-generation cephalosporins.19,20 However, it is susceptible to monobactams, carbapenems, third- and fourth-generation cephalosporins, aminoglycosides, and fluoroquinolones.21 As A. caviae has been isolated, its resistance spectrum has changed, most notably due to the presence of genes encoding the production of β-lactamases, resulting in increased resistance to β-lactam antimicrobials.22 The HZ574 and HZ578 isolates in this study were both resistant to ertapenem and imipenem. The results of whole-genome sequencing showed that HZ574 and HZ578 carried blaNDM-1, indicating that the resistant phenotype was consistent with the genotype. In addition, HZ574 and HZ578 also carry antibiotic resistance genes such as β-lactams, AmpC enzymes, macrolides, aminoglycosides, quinolones and have multidrug-resistant phenotypes.23 Notably, A. caviae in this study had a broader spectrum of resistance compared with A. caviae available at NCBI (Figure 1).
The virulence potential and pathogenic mechanism of A. caviae remain unclear.24 Through the analysis of virulence genes, we found that HZ574 and HZ578 in this study contained many kinds of virulence genes, such as Type VI secretion system, Tap type IV pili, Polar flagella, twitching ATPase, Hemolysin A. These virulence factors can help bacteria attach to the host surface, cause host cell and tissue damage, evade the host immune response, and allow bacteria adhere to the cell surface to form biofilms.25,26 The strains downloaded from NCBI mainly carried secretion pathway protein (83.33%) and related genes of Polar flagella (100%), suggesting that A. caviae in this study may have a more substantial virulence potential (Figure 1).27
The phylogenetic analysis of HZ574 and HZ578 and the complete A. caviae genome downloaded from NCBI showed three evolutionary clones of A. caviae. HZ574 and HZ578 were related in the second clone (Figure 1). This is probably due to the isolation of these two strains being in the same period of time, and both patients were hospitalized in the same hospital. Phylogenetic analysis showed that HZ574 and HZ578 were located in the same branch as GCA_000959705 from Brazil. We speculate that HZ574 and HZ578 belong to the same clone, although two isolates were recovered from different patients. Analysis of the gene-environment of blaNDM-1 found that there are mobile transfer elements around blaNDM-1, which increases the risk of spreading the drug resistance gene, and also suggests that we should carry out routine testing and appropriately treat patients (Figure 3).28 This work may have important implications on the transmission of blaNDM-1 gene in the hospital. We emphasize the importance of improved multisectoral surveillance for carbapenemase-producing isolates, which may contribute to the spread of antimicrobial resistance genes in clinical settings and communities.
Conclusions
We report the A. caviae carrying blaNDM-1 from bacteremia. The study demonstrated the genomic characteristics of A. caviae carrying blaNDM-1 by whole-genome sequencing. The A. caviae isolated in this study has a broad drug resistance spectrum, and the virulence genes carried by the strains pose challenges to clinical treatment. Routine genomic surveillance of this clone is now necessary to effectively curb the dissemination of drug-resistant bacteria in this region.
Ethical Statement
The study protocol was reviewed, approved by and carried out following the recommendations of the Ethics Committee of the Ningbo Medical Center Lihuili Hospital. Written informed consents were obtained from patients. The study complies with the Declaration of Helsinki.
Funding
This project was supported by Zhejiang Medical and Health Science and Technology Project (No. 2022KY1090) and the National Natural Science Foundation of China (No. 82072314).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Fernandez-Bravo A, Figueras MJ. An update on the genus Aeromonas: taxonomy, epidemiology, and pathogenicity. Microorganisms. 2020;8(1):129. doi:10.3390/microorganisms8010129
2. Castelo-Branco DS, Silva AL, Monteiro FO, et al. Aeromonas and Plesiomonas species from scarlet ibis (Eudocimus ruber) and their environment: monitoring antimicrobial susceptibility and virulence. Antonie Leeuwenhoek. 2017;110(1):33–43. doi:10.1007/s10482-016-0771-9
3. Rosso F, Cedano JA, Parra-Lara LG, et al. Emerging carbapenem-resistant Aeromonas spp. infections in Cali, Colombia. Braz J Infect Dis. 2019;23(5):336–342. doi:10.1016/j.bjid.2019.08.005
4. Kitagawa H, Ohge H, Yu L, et al. Aeromonas dhakensis is not a rare cause of Aeromonas bacteremia in Hiroshima, Japan. J Infect Chemother. 2020;26(2):316–320. doi:10.1016/j.jiac.2019.08.020
5. De Silva L, Wickramanayake M, Heo GJ. Virulence and antimicrobial resistance potential of Aeromonas spp. associated with shellfish. Lett Appl Microbiol. 2021;73(2):176–186. doi:10.1111/lam.13489
6. Munoz-Price LS, Poirel L, Bonomo RA, et al. Clinical epidemiology of the global expansion of Klebsiella pneumoniae carbapenemases. Lancet Infect Dis. 2013;13(9):785–796. doi:10.1016/S1473-3099(13)70190-7
7. Nordmann P, Poirel L, Walsh TR, Livermore DM. The emerging NDM carbapenemases. Trends Microbiol. 2011;19(12):588–595. doi:10.1016/j.tim.2011.09.005
8. Han R, Shi Q, Wu S, et al. Dissemination of Carbapenemases (KPC, NDM, OXA-48, IMP, and VIM) among Carbapenem-resistant Enterobacteriaceae isolated from adult and children patients in China. Front Cell Infect Microbiol. 2020;10:314. doi:10.3389/fcimb.2020.00314
9. Kumarasamy KK, Toleman MA, Walsh TR, et al. Emergence of a new antibiotic resistance mechanism in India, Pakistan, and the UK: a molecular, biological, and epidemiological study. Lancet Infect Dis. 2010;10(9):597–602. doi:10.1016/S1473-3099(10)70143-2
10. Villacis JE, Bovera M, Romero-Alvarez D, et al. NDM-1 carbapenemase in Acinetobacter baumannii sequence type 32 in Ecuador. New Microbes New Infect. 2019;29:100526. doi:10.1016/j.nmni.2019.100526
11. Bassetti M, Giacobbe DR, Giamarellou H, et al. Management of KPC-producing Klebsiella pneumoniae infections. Clin Microbiol Infect. 2018;24(2):133–144. doi:10.1016/j.cmi.2017.08.030
12. Falcone M, Daikos GL, Tiseo G, et al. Efficacy of Ceftazidime-avibactam plus Aztreonam in patients with bloodstream infections caused by metallo-beta-lactamase-producing Enterobacterales. Clin Infect Dis. 2021;72(11):1871–1878. doi:10.1093/cid/ciaa586
13. Zheng B, Zhang J, Ji J, et al. Emergence of Raoultella ornithinolytica coproducing IMP-4 and KPC-2 Carbapenemases in China. Antimicrob Agents Chemother. 2015;59(11):7086–7089. doi:10.1128/AAC.01363-15
14. Lellouche J, Schwartz D, Elmalech N, et al. Combining VITEK((R)) 2 with colistin agar dilution screening assist timely reporting of colistin susceptibility. Clin Microbiol Infect. 2019;25(6):711–716. doi:10.1016/j.cmi.2018.09.014
15. Xu H, Wang X, Yu X, et al. First detection and genomics analysis of KPC-2-producing Citrobacter isolates from river sediments. Environ Pollut. 2018;235:931–937. doi:10.1016/j.envpol.2017.12.084
16. Stohr J, Kluytmans-van den Bergh MFQ, Weterings V, Rossen JWA, Kluytmans J. Distinguishing bla KPC gene-containing IncF plasmids from epidemiologically related and unrelated Enterobacteriaceae based on short- and long-read sequence data. Antimicrob Agents Chemother. 2021;65(6). doi:10.1128/AAC.00147-21
17. Croucher NJ, Page AJ, Connor TR, et al. Rapid phylogenetic analysis of large samples of recombinant bacterial whole genome sequences using Gubbins. Nucleic Acids Res. 2015;43(3):e15. doi:10.1093/nar/gku1196
18. Liang Z, Pang J, Hu X, et al. Low prevalence of mcr-1 among clinical Enterobacteriaceae isolates and co-transfer of mcr-1 and blaNDM-1 from separate donors. Microb Drug Resist. 2021;27(4):476–484. doi:10.1089/mdr.2020.0212
19. Janda JM, Abbott SL. The genus Aeromonas: taxonomy, pathogenicity, and infection. Clin Microbiol Rev. 2010;23(1):35–73. doi:10.1128/CMR.00039-09
20. Vila J, Ruiz J, Gallardo F, et al. Aeromonas spp. and traveler’s diarrhea: clinical features and antimicrobial resistance. Emerg Infect Dis. 2003;9(5):552–555. doi:10.3201/eid0905.020451
21. Codjoe FS, Donkor ES. Carbapenem resistance: a review. Med Sci. 2017;6(1):1.
22. Dias C, Serra CR, Simoes LC, Simoes M, Martinez-Murcia A, Saavedra MJ. Extended-spectrum beta-lactamase and carbapenemase-producing Aeromonas species in wild animals from Portugal. Vet Rec. 2014;174(21):532. doi:10.1136/vr.101803
23. Chi X, Berglund B, Zou H, et al. Characterization of clinically relevant strains of extended-spectrum beta-lactamase-producing Klebsiella pneumoniae occurring in environmental sources in a rural area of China by using whole-genome sequencing. Front Microbiol. 2019;10. doi:10.3389/fmicb.2019.00211
24. Silva L, Leal-Balbino TC, Melo BST, et al. Genetic diversity and virulence potential of clinical and environmental Aeromonas spp. isolates from a diarrhea outbreak. BMC Microbiol. 2017;17(1):179. doi:10.1186/s12866-017-1089-0
25. Tomas JM. The main Aeromonas pathogenic factors. ISRN Microbiol. 2012;2012:256261. doi:10.5402/2012/256261
26. Batra P, Mathur P, Misra MC. Aeromonas spp.: an emerging nosocomial pathogen. J Lab Physicians. 2016;8(1):1–4. doi:10.4103/0974-2727.176234
27. Hofer E, Reis CM, Theophilo GN, Cavalcanti VO, Lima NV, Henriques Mde F. [Aeromonas associated with an acute diarrhea outbreak in Sao Bento do Una, Pernambuco]. Rev Soc Bras Med Trop. 2006;39(2):217–220. Swedish. doi:10.1590/S0037-86822006000200016
28. Kim JS, Jin YH, Park SH, et al. Horizontal transfer of blaNDM-1-carrying IncX3 plasmid between carbapenem-resistant Enterobacteriaceae in a single patient. J Infect. 2020;81(5):816–846. doi:10.1016/j.jinf.2020.07.013
 © 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.