Back to Journals » Journal of Pain Research » Volume 18
Decoding Fibromyalgia: Genetic Insights into Gut and Immune System Interactions
Authors Niu M, Li J, Sarafian V ![]() , Maes M
, Maes M ![]()
Received 31 March 2025
Accepted for publication 10 September 2025
Published 31 October 2025 Volume 2025:18 Pages 5685—5699
DOI https://doi.org/10.2147/JPR.S525610
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Michael Überall
Mengqi Niu,1,2 Jing Li,1,2 Victoria Sarafian,3,4 Michael Maes1– 7
1Sichuan Provincial Center for Mental Health, Sichuan Provincial People’s Hospital, School of Medicine, University of Electronic Science and Technology of China, Chengdu, 610072, People’s Republic of China; 2Key Laboratory of Psychosomatic Medicine, Chinese Academy of Medical Sciences, Chengdu, 610072, People’s Republic of China; 3Department of Psychiatry, Medical University of Plovdiv, Plovdiv, Bulgaria; 4Research Institute at Medical University of Plovdiv, Plovdiv, Bulgaria; 5Research and Innovation Program for the Development of MU – Plovdiv (SRIPD-MUP), Network of Research Higher Schools, Medical University of Plovdiv, Plovdiv, Bulgaria; 6Kyung Hee University, Seoul, Korea; 7Department of Psychiatry, Faculty of Medicine, Chulalongkorn University, and King Chulalongkorn Memorial Hospital, the Thai Red Cross Society, Bangkok, Thailand
Correspondence: Michael Maes, Sichuan Provincial Center for Mental Health, Sichuan Provincial People’s Hospital, School of Medicine, University of Electronic Science and Technology of China, Chengdu, 610072, People’s Republic of China, Tel +86 028 8102 0064, Email [email protected]
Background: Fibromyalgia (FM) is a chronic disorder characterized by widespread pain and immune dysregulation. Emerging evidence suggests that gut microbiota and inflammatory proteins may contribute to the development of FM. The aim of this study was to investigate the causal relationships between gut microbiota, inflammatory proteins (cytokines/chemokines), and FM using bidirectional Mendelian randomization (MR) and meta-analysis approaches.
Methods: MR analyses were conducted using genetic data from European populations, employing methods such as MR-IVW, MR-Egger, and MR-weighted median. Reverse MR was also performed, with FM treated as the exposure. A meta-analysis was conducted to consolidate the findings.
Results: Ruminococcus gauvreauii was identified as a risk factor for FM, while Enterorhabdus, Parabacteroides, Butyricicoccus, and Prevotella 9 were found to be protective. Five inflammatory proteins—C-X-C motif chemokine 5 (CXCL5), S100-A12, Leukemia inhibitory factor receptor (LIFR), Monocyte chemoattractant protein 2 (MCP-2/CCL8), and Tumor necrosis factor (TNF-α)—exhibited protective associations, while Natural killer cell receptor 2B4 (NKCR-2B4/CD244) and Interleukin-12 subunit beta (IL-12β) were associated with an increased risk of FM.
Conclusion: This study highlights the role of gut microbiota and inflammatory proteins (cytokines/chemokines) in the pathogenesis of FM. Through Gene Ontology (GO) functional enrichment and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analyses, the findings suggest their involvement in immune regulation, inflammatory responses, and viral pathways. These findings provide new insights into potential therapeutic targets for modulating gut health and immune responses, opening new avenues for future research and clinical interventions.
Keywords: fibromyalgia, gut microbiota, inflammatory proteins, mendelian randomization study, meta analysis, multi-omics
Introduction
Fibromyalgia (FM) is a condition characterized by widespread chronic musculoskeletal pain, stiffness in specific areas, and a heightened sensitivity to pressure, known as tender points in soft tissue.1 Despite pain being its primary and prominent feature, FM is characterized by a complex array of symptoms including fatigue, sleep disturbances, and functional symptoms (medical symptoms that cannot be explained by structural or pathological definitions).2 FM also exerts a significant impact on mental health, with common associated symptoms including depression, anxiety, cognitive impairment (often referred to as “fibro fog”), and emotional distress.3 Globally, FM is estimated to affect 2.7% of the population, with higher prevalence rates observed in women, individuals over 50 years old, those with lower socioeconomic status, and obese individuals.4 Central sensitization refers to a pathological state in which the central nervous system exhibits an exaggerated response to external stimuli. It is currently regarded as one of the key mechanisms underlying FM, contributing to heightened pain perception and the persistence of symptoms.5 However, due to the lack of consensus on the pathogenic mechanisms underpinning FM, there is currently no universally effective treatment.2
A recent meta-analysis shows that FM is characterized by indicants of peripheral immune activation with changes in immunoregulatory cytokines and pro-inflammatory cytokines.6 FM patients exhibit enhanced circulating inflammatory cytokines and chemokines released by peripheral blood mononuclear cells (PBMCs), indicating activation of both innate and adaptive immunity.7–9 Current research revealed elevated concentrations of M1 macrophage cytokines, such as IL-6, IL-8, IL-1β, and TNF-α, in the serum of FM patients.6,7,10–12 Immune cells, including mast cells, monocytes, and neutrophils, which mediate the inflammatory process, may also contribute to the immune pathogenesis of FM.13 In patients with major depression, acute COVID-19 infection, Long-COVID, schizophrenia and end-stage renal disease, FM symptoms are associated with immune activation as indicated by increased levels of pro-inflammatory mediators.14–18 Interestingly, plasma levels of soluble gp130 (sgp130), the natural inhibitor of IL-6 transsignalling, is significantly decreased in FM,19 which together with increased IL-6 levels suggest that the immune activation in FM is mainly driven by increased IL-6 signaling.6
Additionally, in animal studies, resident macrophages in muscle tissue have been shown to play a role in the development of chronic widespread muscle pain.20 Increasing evidence suggests that neurogenic inflammatory processes occurring in peripheral tissues, the spinal cord, and the brain contribute to the pathophysiology of FM.7,21,22 All of these translate into the clinical symptoms reported by FM patients, such as swelling and sensory dullness, which also impact central symptoms including cognitive changes and fatigue. Furthermore, physiological mechanisms related to stress and emotions are considered upstream driving factors of neurogenic inflammation in FM.9,23 Psychological factors such as chronic stress, anxiety, and depression have been shown to modulate neurogenic inflammation by altering neuroimmune interactions, thereby exacerbating the symptoms of FM.23,24
In 2007, it was published that FM symptoms of chronic fatigue syndrome are associated with increased translocation of Gram-negative gut commensal bacteria, indicating leaky gut.25 In schizophrenia patients, FM symptoms are strongly associated with breakdown of the paracellular pathways (leaky gut) and increased bacterial translocation.15 Other findings suggested that gut dysbiosis may play a role in FM. Thus, studies by Minerbi et al26 and Minerbi and Fitzcharles et al27 reported associations between gut microbiota and FM. In addition, a systematic review showed that intestinal microbiota is involved in the pathophysiology of FM.28
However, until now it has remained unclear whether there are causal relationships between immune activation, the gut-immune axis, and FM. Recently developed Mendelian randomization (MR) designs have been used to study whether specific exposures have causal effects on specific outcomes.29 As an analytical method, MR designs utilize lineage genetic variations (single nucleotide polymorphisms, SNPs) as instrumental variables for the target exposure.30 Based on the random assortment during meiosis and the fixed allocation of genetic variations at conception, MR studies are less susceptible to confounding factors and reverse causation biases. Additionally, the use of summary statistics from genome-wide association studies (GWAS) in two-sample MR designs significantly enhances the statistical power of causal inference.31 Therefore, in this study, we use summary statistics from GWAS to conduct MR research. The aim is to investigate whether immune activation and alterations in the gut microbiota play a causal role in the pathogenesis of FM. The a priori hypothesis is to discover clear multi-omics causal relationships between FM and immune activation as assessed with cytokines/chemokines and changes in the gut microbiota indicating an inflammatory milieu or leaky gut.
Materials and Methods
Study Design
In our analysis, we used publicly available data from shared repositories. In accordance with Article 32, Items 1 and 2 of the Measures for Ethical Review of Life Science and Medical Research Involving Human Subjects (February 18, 2023, China), no additional ethical approval was required for this analysis. The MR analysis must meet three assumptions: (1) SNPs are associated with exposure; (2) SNPs are independent of confounders in the exposure-outcome relationship (independence assumption); (3) SNPs influence the outcome solely through exposure.32 Figure 1 illustrates the workflow of this study.
 |
Figure 1 The flowchart of bidirectional mendelian randomization study and meta-analysis explore the causal relationship between gut microbiota, inflammatory proteins, and fibromyalgia. PQTL: protein quantitative trait loci. |
Data Sources
This study utilized two types of exposure data and two types of outcome data. To minimize bias due to population heterogeneity, only aggregated data from European populations were used. SNPs related to the human gut microbiome composition were obtained from the latest data from the MiBioGen international consortium (https://mibiogen.gcc.rug.nl/). This GWAS included 211 microbial taxa classified at six levels, from phylum to species.33 We sourced data on 91 inflammatory protein types from the study by Zhao et al.34 Summary statistics for FM outcomes were sourced from the FinnGen consortium (released in 2023, https://r10.finngen.fi/) and the study by Moscati et al.35
Genetic Instrument Selection
(1) Based on previous studies, a significance threshold (p < 1×10−5) was applied for gut microbiota,36 while a stricter p-value (p < 5×10−6) was used for inflammatory proteins37 to ensure a moderate number of SNPs for all traits. (2) To ensure independence of the instruments used for exposure, we excluded SNPs in linkage disequilibrium (LD) (r² < 0.001, clumping window = 10,000 kb).38 (3) Palindromic SNPs with intermediate minor allele frequencies (MAF) were excluded.39 (4) To determine the existence of weak instrument bias, an F-statistic was computed to measure the power of the instrumental variable. An F-statistic above 10 indicated a low likelihood of weak instrument bias.40
Two-Sample Mendelian Randomization
This study primarily relied on random effects inverse variance weighting (IVW) to assess the causal effect of gut microbiota, and inflammatory proteins on FM. The IVW estimates were derived by aggregating the Wald ratios of all genetic variants. IVW is based on the assumption that all SNPs do not exhibit horizontal pleiotropy, thus providing the most accurate causal effect estimate. To ensure robustness, we also employed MR-Egger and weighted median (WM) methods for supplementary analysis.31,41
Sensitivity analyses are essential for examining horizontal pleiotropy and heterogeneity, which can significantly impact MR estimates. In this study, we used three methods to detect and correct these issues: Cochran’s Q test, MR-Egger intercept test, and MR-PRESSO (Pleiotropy RESidual Sum and Outlier) analysis. A p-value < 0.05 from Cochran’s Q test indicated heterogeneity in the results.42 The MR-Egger intercept was used to detect directional pleiotropy and bias due to invalid instruments.41 Finally, MR-PRESSO was used to identify and correct for potential pleiotropic outliers.43 To assess the robustness of the results, we employed the leave-one-out method, sequentially excluding each SNP to determine if any single SNP drove the estimates.41
We stringently screened the gut microbiota and cytokines/chemokines with potential causal relationships to FM based on several criteria: (1) Significant p-value (p < 0.05) from the primary MR IVW analysis; (2) Consistent direction and magnitude across three MR methods; (3) No significant heterogeneity or horizontal pleiotropy in MR results.
Meta-Analysis and Genetic Directionality Assessment
To comprehensively estimate the robustness of gut microbiota and inflammatory proteins identified by the aforementioned criteria, we used the FinnGen consortium’s summary statistics for FM as the preliminary analysis. We replicated the IVW analysis in another FM cohort (study by Moscati et al). The meta-analysis of the results from the two MR analyses identified gut microbiota and immune proteins causally related to FM.
Additionally, to rule out bias from reverse causality and further evaluate the robustness of the causal relationship between gut microbiota and immune proteins, and FM, we conducted reverse MR analysis within the preliminary analysis cohort (FinnGen consortium) for significant results (P < 0.05) from the meta-analysis. This provided final evidence for causal relationships.
Statistical analyses were performed using R software (version 4.3.2) with the packages MendelianRandomization, MRPRESSO, TwoSampleMR, Meta, among others. For binary outcomes, results were reported as odds ratios (OR) with corresponding 95% confidence intervals (CI) to quantify the strength of causal relationships. For continuous outcomes, results were reported as β coefficients, standard errors (SE), and corresponding 95% CI.
Protein-Protein Interaction (PPI) Annotation and Enrichment
To better explore the interactions between proteins and the pathogenesis, we performed PPI annotation and enrichment analysis using STRING (https://cn.string-db.org) and Kyoto Encyclopedia of Genes and Genomes (KEGG) and Gene Ontology (GO) databases.
Results
Causal Relationship Between Gut Microbiota and Fibromyalgia
After excluding unnamed bacteria, we identified 196 gut microbiotas. In the preliminary analysis cohort, six gut microbiota showed statistically significant results (Supplementary Table 1), including family Enterobacteriaceae (OR = 0.68, 95% CI: 0.47–0.98, p = 0.041), genus Butyricicoccus (OR = 0.68, 95% CI: 0.49–0.95, p = 0.023), genus Coprococcus1 (OR = 0.69, 95% CI: 0.52–0.93, p = 0.014), genus Eggerthella (OR = 1.31, 95% CI: 1.08–1.59, p = 0.006), genus Ruminococcaceae UCG005 (OR = 1.37, 95% CI: 1.04–1.81, p = 0.025), and order Enterobacteriales (OR = 0.68, 95% CI: 0.47–0.98, p = 0.041). Significant gut microbiota also passed sensitivity tests including Cochran’s Q test (p > 0.05), MR-Egger intercept test (p > 0.05), MR-PRESSO analysis (p > 0.05), and leave-one-out analysis (Supplementary Table 2 and Figure S1). We replicated the MR analysis using another GWAS dataset for FM. As expected, similar trends were observed for the candidate gut microbiota in another GWAS dataset (study by Moscati et al), but results were not significant due to large sample size differences.
To strengthen the credibility of the estimates, we performed a meta-analysis of MR results from the preliminary and replication cohorts, identifying six gut microbiota influencing FM (Figure 2). Specifically, higher levels of genus Enterorhabdus (OR = 0.78, 95% CI: 0.61–1.00, p = 0.048), genus Parabacteroides (OR = 0.61, 95% CI: 0.41–0.91, p = 0.016), genus Butyricicoccus (OR = 0.73, 95% CI: 0.55–0.96, p = 0.027), genus Candidatus Soleaferrea (OR = 0.83, 95% CI: 0.70–1.00, p = 0.046), and genus Prevotella 9 (OR = 0.83, 95% CI: 0.69–0.99, p = 0.035) reduced the risk of FM. However, higher levels of genus Ruminococcus gauvreauii group (OR = 1.56, 95% CI: 1.19–2.04, p = 0.001) increased susceptibility to FM. In the reverse MR analysis (Table 1), we found a reverse causal relationship between FM and the genus Candidatus Soleaferrea (p < 0.05), indicating that genus Candidatus Soleaferrea did not pass the reverse causality validation.
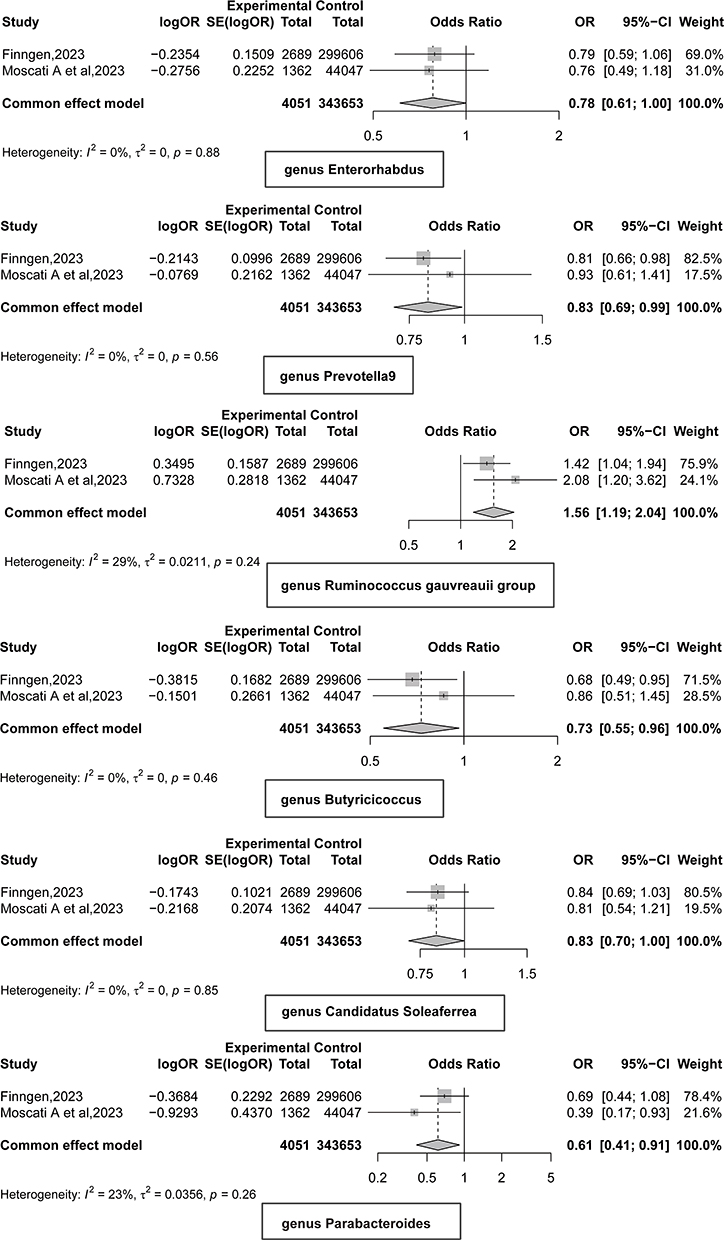 |
Figure 2 Meta results of gut microbiota and fibromyalgia mendelian randomization analysis. OR < 1, protective factor; OR > 1, risk factors. Summary statistics for fibromyalgia were sourced from the FinnGen consortium (released in 2023, https://r10.finngen.fi/) and the study by Moscati et al.35 |
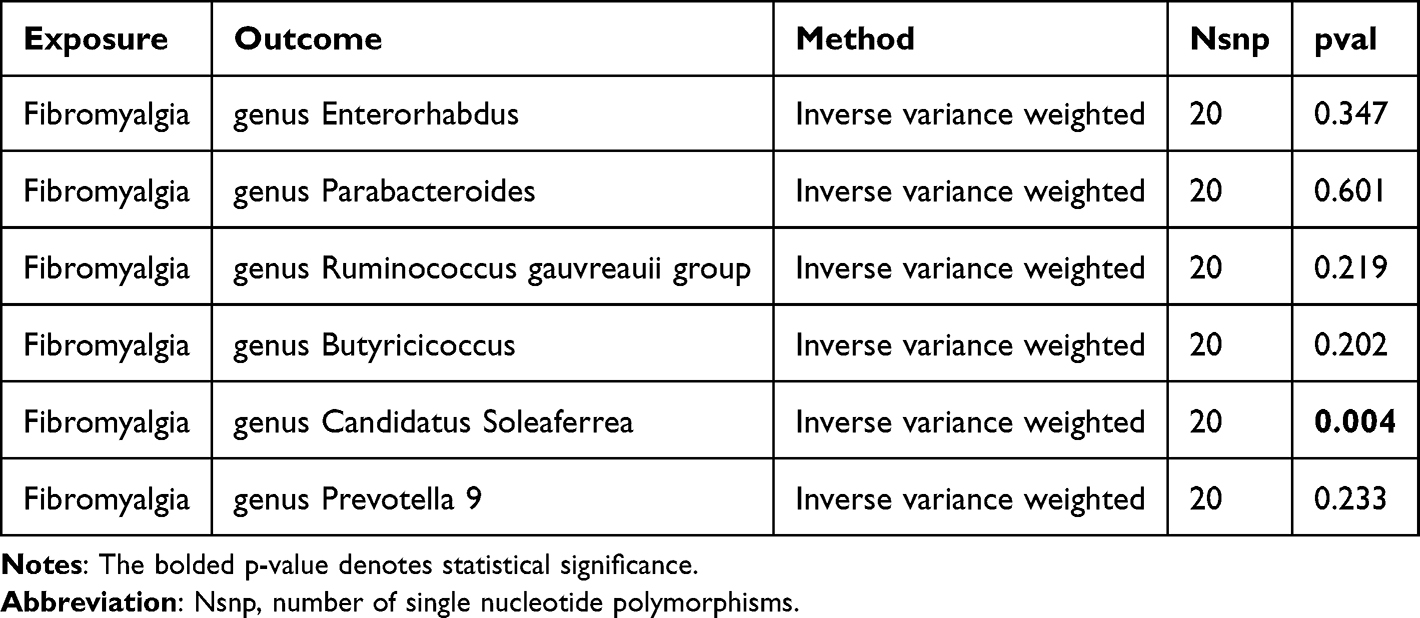 |
Table 1 Inverse Mendelian Randomization (MR) Results of Fibromyalgia and Significant Gut Microbiota After Meta-Analysis |
Causal Relationship Between 91 Inflammatory Proteins and Fibromyalgia
In the preliminary analysis cohort, six inflammatory proteins showed statistically significant results (Supplementary Table 3), including Natural killer cell receptor 2B4 (NKCR-2B4/CD244; OR = 1.18, 95% CI: 1.02–1.38, p = 0.031), CUB domain-containing protein 1 (OR = 0.78, 95% CI: 0.61–0.99, p = 0.037), S100-A12 (OR = 0.78, 95% CI: 0.61–0.99, p = 0.037), Interleukin-12 subunit beta (IL-12β; OR = 1.11, 95% CI: 1.02–1.21, p = 0.016), Monocyte chemoattractant protein 2 (MCP-2,/CCL8; OR = 0.90, 95% CI: 0.84–0.97, p = 0.008), and Tumor necrosis factor (TNF-ɑ; OR = 0.77, 95% CI: 0.61–0.96, p = 0.020). Significant immune proteins also passed sensitivity tests including Cochran’s Q test (p > 0.05), MR-Egger intercept test (p > 0.05), MR-PRESSO analysis (p > 0.05), and leave-one-out analysis (Supplementary Table 4 and Figure S2). MR analysis in the replication cohort showed similar trends for candidate immune proteins, but results were not significant due to large sample size differences.
We performed a meta-analysis of MR results from the preliminary and replication cohorts, ultimately identifying seven immune proteins influencing FM (Figure 3). Specifically, higher levels of C-X-C motif chemokine 5 (CXCL5; OR = 0.89, 95% CI: 0.80–1.00, p = 0.046), S100-A12 (OR = 0.76, 95% CI: 0.62–0.93, p = 0.007), Leukemia inhibitory factor receptor (LIFR; OR = 0.85, 95% CI: 0.73–0.98, p = 0.025), MCP-2/CCL8 (OR = 0.93, 95% CI: 0.87–0.99, p = 0.025), and TNF-α (OR = 0.79, 95% CI: 0.65–0.95, p = 0.013) reduced the risk of FM.
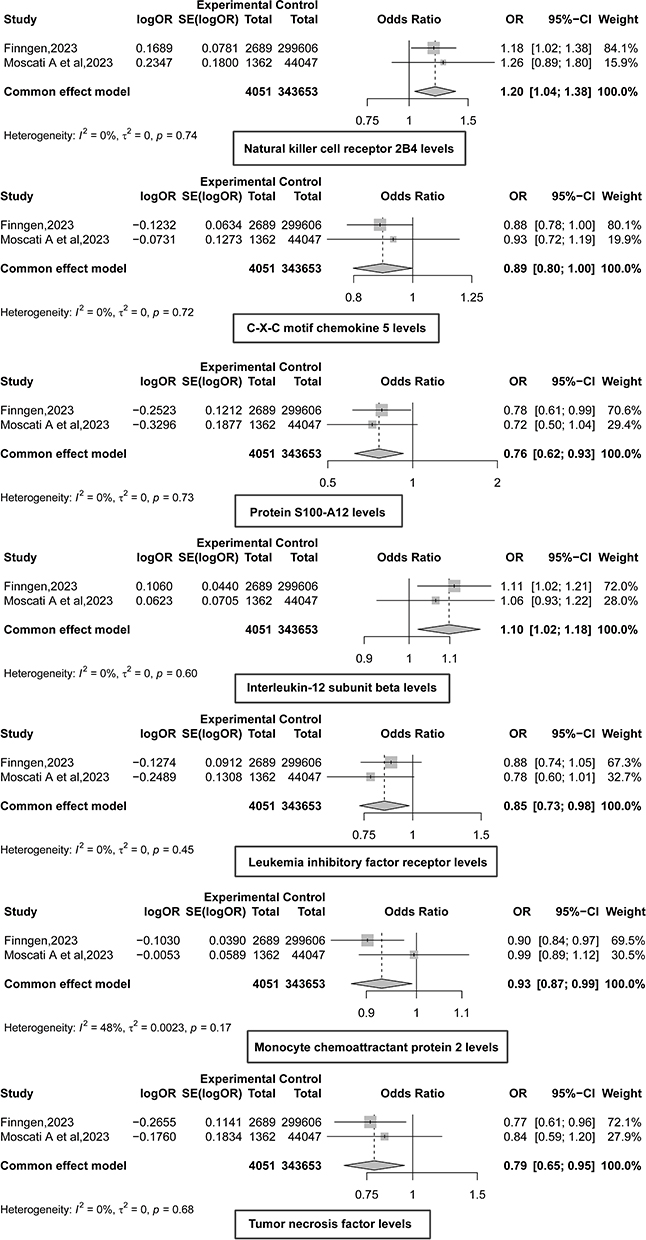 |
Figure 3 Meta results of inflammatory proteins and fibromyalgia mendelian randomization analysis. OR < 1, protective factor; OR > 1, risk factors. Summary statistics for fibromyalgia were sourced from the FinnGen consortium (released in 2023, https://r10.finngen.fi/) and the study by Moscati et al.35 |
However, higher levels of NKCR-2B4/CD244 (OR = 1.20, 95% CI: 1.04–1.38, p = 0.012) and IL-12β (OR = 1.10, 95% CI: 1.02–1.18, p = 0.012) increased susceptibility to FM. All significant results in the meta-analysis passed the reverse causality validation in subsequent analyses (Table 2).
 |
Table 2 Inverse Mendelian Randomization (MR) Results of Fibromyalgia and Significant Inflammatory Protein After Meta-Analysis |
For the genes CXCL5, S100-A12, LIFR, CCL8, and TNF, the functional enrichment analyses using GO biological processes and KEGG pathways are shown in Supplementary Table 5. According to results after False Discovery Rate (FDR), the GO analysis shows significant enrichment mainly in the “neutrophil chemotaxis”. KEGG pathway analysis indicates notable enrichment in “viral protein interaction with cytokine and cytokine receptor”. For the genes CD244 and IL-12β, the GO biological process and KEGG functional enrichment analyses are presented in Supplementary Table 6. Post-FDR analysis reveals significant enrichment primarily in “positive regulation of interferon-gamma production” for GO biological processes, while KEGG analysis highlights significant enrichment in the “JAK-STAT signaling pathway”.
Discussion
Gut Microbiota and Fibromyalgia
Increasing evidence suggests that symbiotic gut bacteria play a significant role in FM.25–28 In our study, we identified Enterorhabdus, Parabacteroides, Butyricicoccus, and Prevotella 9 as protective factors against FM, while the R. gauvreauii group was identified as a risk factor.
Research on Enterorhabdus is currently limited. A study investigating the antidepressant effects of Ziyan Green Tea on mildly stressed mice through fecal metabolomics revealed a significant negative correlation between Enterorhabdus and the pro-inflammatory cytokine IL-6.44 This suggests that Enterorhabdus may help inhibit inflammatory responses, thereby acting as a protective factor. Parabacteroides has been shown to produce short-chain fatty acids (SCFAs), such as butyrate, which play critical roles in immune regulation.28,45,46 Parabacteroides may also help repair the intestinal barrier, reduce gut inflammation, and improve metabolic abnormalities like insulin resistance,47 highlighting its potential as a protective factor in FM. Butyricicoccus is one of the main producers of butyrate, a key immunomodulatory SCFA that inhibits dendritic cell differentiation and promotes the development of anti-inflammatory regulatory T cells, underscoring the potential role of Butyricicoccus in FM.26,28,48 Research on Prevotella 9 in FM is scarce, but studies have suggested that Prevotella may act as a potential probiotic.49,50 For example, in multiple sclerosis, treatment with Prevotella histicola increased the frequency and number of CD4FoxP3 regulatory T cells in the periphery and gut, while decreasing pro-inflammatory IFN-γ and IL17-producing CD4 T cells in the central nervous system, suggesting that Prevotella histicola suppresses disease progression by enhancing anti-inflammatory immune responses and inhibiting pro-inflammatory ones.50 Similarly, Prevotella may have protective properties through its anti-inflammatory activities.
Ruminococcus, on the other hand, may activate inflammatory signaling pathways through lipid dysregulation (especially polyether amine and glycerophosphocholine) and NOD-like receptor family pyrin domain containing 3 (NLRP3) inflammasome activation (45). Importantly, R. gauvreauii may induce a systemic immune response by activating IL-6 and TNF-α signaling.51,52 Moreover, R. gauvreauii may be associated with an increased risk for gastro-duodenal ulcers.53 Studies on Ruminococcus in depression have shown conflicting results. Zhao et al found higher levels of Ruminococcus in major depressive disorder (MDD) patients, positively correlating with IL-1β, suggesting a pro-inflammatory role that may be important to the pathophysiology of FM.54 Conversely, other studies have shown higher relative abundance of Ruminococcus in healthy controls compared to MDD patients.55
In 2007 (Maes et al, 2007), it was reported that the FM symptoms of chronic fatigue syndrome are associated with increased translocation of Gram-negative gut commensal bacteria and gut barrier dysfunction.25 Dysbiosis, particularly the imbalance between pathobionts (which promote inflammation and damage to epithelial cells and tight junctions) and symbionts (health-promoting microbiota, which engage in anti-inflammatory activities, support gut homeostasis and tight junctions, and produce SCFAs and vitamins), is a potential cause of gut barrier dysfunction and bacterial translocation, playing a crucial role in FM.56–59 Consequently, our results may suggest that FM is associated with a specific enterotype, namely with R. gauvreauii as a risk factor and Enterorhabdus, Parabacteroides, Butyricicoccus, and Prevotella 9 as protective factors. These results deserve replication in new cohorts of patients with FM.
Cytokines/Chemokines in Fibromyalgia
In this study, five inflammatory proteins—CXCL5, S100-A12, LIFR, MCP-2/CCL8, and TNF-α—were identified as protective factors against FM. Conversely, NKCR-2B4/CD244 and IL-12β were identified as risk factors.
Functional enrichment analysis of CXCL5, S100-A12, LIFR, CCL8, and TNF genes revealed significant enrichment in the GO biological process “Cellular response to chemical stimulus”, with other genes also enriched in “Neutrophil chemotaxis”, “Leukocyte migration”, and “Cytokine-mediated signaling pathway”. KEGG pathway analysis indicated that CXCL5, CCL8, LIFR, and TNF genes were enriched in pathways such as “Viral protein interaction with cytokine and cytokine receptor”, “Cytokine-cytokine receptor interaction”, and “Chemokine signaling pathway”. For CD244 and IL-12β, GO biological process analysis showed enrichment in “Positive regulation of interferon-gamma production”; CD244 was also enriched in “Positive regulation of natural killer cell activation” and “Positive regulation of natural killer cell mediated immunity”. KEGG analysis revealed that CD244 was enriched in pathways like “JAK-STAT signaling pathway”, “Inflammatory bowel disease”, and “Th1 and Th2 cell differentiation”.
CXCL5, a CXCR2 ligand chemokine, is involved in recruiting leukocytes to tissues, contributing to neuroinflammation, blood-brain barrier breakdown, and neutrophil infiltration into the brain.60 Previous studies have shown that regulating the IL-6 pathway can alleviate pain-like behaviors in FM by reducing pro-inflammatory cytokines such as TNF-α and CXCL5. These observations may seem contradictory to our findings but reflect the complex interplay between immune activation and imbalance in FM, where both immune-inflammatory response system (IRS) and compensatory immune-regulatory response system (CIRS) are activated, leading to imbalances in regulatory and pro-inflammatory cytokines.6
S100A12, a member of the S100 calcium-binding protein family, is mainly secreted by neutrophils, macrophages, and smooth muscle cells. Under inflammatory conditions, S100A12 binds to receptor for advanced glycation end-products (RAGE), signaling through JNK, ERK, and p38 to activate transcription factors AP-1 and nuclear factor (NF)-κB, leading to the release of pro-inflammatory cytokines.61 Elevated S100A12 levels have been observed in the saliva of FM patients.62 Previous studies also identified S100A12 as a diagnostic biomarker for rheumatoid arthritis and depression.63
In a 2002 study by Maes et al, it was found that patients with depressive and anxiety symptoms during the early postpartum period exhibited reduced serum LIFR levels, which may contribute to immune activation.64 Another study indicated that long-term treatment with atypical antipsychotics could increase the anti-inflammatory capacity of serum in schizophrenia patients by elevating serum LIFR levels.65
MCP-2/CCL8, a member of the CC chemokine subfamily, has been reported as an agonist for C-C chemokine receptor types 2 and 4. It plays a crucial role in regulating leukocyte chemotaxis and inflammatory diseases.57 A study by Bäckryd et al found high levels of the chemokine CCL8 in the cerebrospinal fluid of patients with peripheral neuropathic pain compared to healthy controls.66 In a mouse study, MCP-2/CCL8 levels were increased in oxidative stress-induced allergic inflammation, and anti-inflammatory treatment was shown to reduce MCP-2/CCL8 levels.67 Another study revealed that MCP-2/CCL8 levels were higher in MDD patients with suicidal ideation (SI) compared to those without SI and healthy controls.68 MCP-2/CCL8 may contribute to the neurobiology of suicidality in depressed elderly individuals by enhancing neuroinflammatory responses, oxidative stress, apoptotic pathways, impairing neurogenesis, and disrupting serotonergic neurotransmission.68
TNF-α is one of the most critical cytokines released by macrophages and microglia and is active in neuropathic pain pathways.69 Although our MR analysis revealed a protective association between genetically predicted TNF-α levels and the risk of FM, TNF-α may exert dual effects depending on its concentration, receptor distribution, the immune microenvironment, and temporal dynamics. At low levels or with transient increases, membrane-bound TNF-α signals through TNF receptor 2 to promote tissue repair and maintain immune homeostasis. In contrast, sustained high levels of soluble TNF-α binding to TNF receptor 1 can trigger chronic inflammation and heightened pain sensitivity.69 Thus, the overgrowth of Ruminococcus gauvreauii may push TNF-α/IL-6 signaling beyond a physiological threshold, shifting the balance between the IRS and CIRS toward a pro-inflammatory state, thereby contributing to FM symptoms.51,52 Research on TNF levels in FM patients has produced mixed results, with some studies reporting lower TNF levels10,70,71 and others finding elevated TNF levels.72,73
CD244, also known as natural killer cell receptor 2B4, is a transmembrane protein belonging to the immunoglobulin superfamily and plays a vital role in immune regulation. KEGG analysis revealed that CD244 is primarily enriched in the JAK-STAT signaling pathway. Previous studies have shown that this pathway plays a significant role in FM pathogenesis, leading to chemokine overexpression and glial cell activation.74
IL-12β is a pro-inflammatory cytokine and a component of IL-12, involved in activating Th1 immune responses. GO biological process analysis revealed that CD244, along with IL-12β, enhances inflammatory responses through the positive regulation of interferon-gamma production, which may be a risk factor for FM. Previous studies have found elevated IL-12 in patients with FM syndrome.74,75 Recently, it has been shown that IL-12 also induces inflammatory hypernociception.76 A study demonstrated mechanical hypernociception in rats injected with IL-12, mediated by endothelin-1 and endothelin-B receptor interaction. This finding is significant as pain at the IL-12 injection site has been reported in human treatment.77 IL-12 has also been identified as an essential pro-inflammatory factor in other diseases, such as COVID-19, major depression, psoriasis, and systemic lupus erythematosus.78–82
To summarize, the immunological characteristics observed in the present investigation of FM cannot be solely attributed to a simple “inflammation”. The development of FM may be assigned to an intricate imbalance between the IRS and the CIRS. A recent meta-analysis has substantiated that FM is distinguished by alterations and disparities in immunoregulatory and pro-inflammatory cytokines, resulting in peripheral immunological activation.6 Based on the analysis of enrichment and annotation, the present study indicates that FM may be associated with elevated production of IFN-γ and activation of natural killer cells, as well as reduced chemotaxis and diminished defense against viruses. Indeed, CXCL5, CCL8, and TNF participate in the hsa04061 KEGG pathway (KEGG PATHWAY: hsa04061 (genome.jp)), specifically referred to as “viral protein interaction with cytokine and cytokine receptor”. DNA viruses could undermine the host immune system network in this pathway. A recent study by Maes et al (2024) demonstrated a significant association between the replication of human herpes type 6 (HHV-6) and the manifestation of FM symptoms in individuals with Long COVID. In addition, reactivation of HHV-6 can potentially impact the integrity of the gut tight junctions, resulting in immune-inflammatory reactions.83 Another report indicated that individuals suffering from FM may exhibit HHV-6 infection.84 Consequently, the replication of HHV-6 may play a role in the immune-inflammatory mechanisms associated with FM by activation the immune-inflammatory response system,85 impairing chemotaxis and inducing antiviral responses via elevated production of IFN-γ.86 Consequently, the elevation of IFN-γ production has the potential to elicit peripheral pain responses via interactions between neurons and microglia within the spinal cord.86,87 The JAK-STAT signaling pathway is a key route for IFN-γ signal transduction; its activation can induce microglial release of pro-inflammatory mediators and enhance central sensitization, thereby potentially modulating chronic pain mechanisms in FM.88 This is consistent with our enrichment analysis, which shows significant enrichment of “positive regulation of interferon-gamma production” and the “JAK-STAT signaling pathway”, suggesting a state of immune activation in FM that may drive pain sensitization and chronic inflammation. The immune response in FM may be exacerbated by the heightened pro-inflammatory effects of R. gauvreauii, alongside a diminished presence of gut bacteria that typically provide a protective barrier against inflammation and the phenomenon of leaky gut. This includes notable genera such as Enterorhabdus, Parabacteroides, Butyricicoccus, and Prevotella 9.
Limitations
When interpreting the results of this study, certain limitations must be acknowledged. First, the study population was limited to individuals of European ancestry. Given the differences in allele frequencies across ethnic groups, this homogeneity restricts the generalizability of the causal inferences to other populations. Second, the dataset lacked essential demographic variables such as age and sex, which hindered more detailed subgroup analyses. Third, although MR is a robust method for inferring causal relationships between exposures and outcomes, the current analysis relied solely on cross-sectional data and could not capture temporal changes within the causal pathway. Future research should aim to replicate these findings in large, multi-ethnic, multicentre cohorts, incorporating variables such as age and sex to enable more refined stratified analyses. In addition, prospective longitudinal or interventional studies, combined with animal models and in vitro functional assays, are recommended to dynamically track the biological roles of key microbial taxa and their associated proteins over time. This integrative approach could provide a more comprehensive and robust evidence base for understanding the pathophysiology of FM.
Conclusion
This study presents a comprehensive bidirectional MR analysis and meta-analysis investigating the causal relationships between gut microbiota, inflammatory proteins, and FM. Our findings identify R. gauvreauii as a potential risk factor for FM, while Enterorhabdus, Parabacteroides, Butyricicoccus, and Prevotella 9 appear to act as protective factors. Additionally, five inflammatory proteins—CXCL5, S100-A12, LIFR, MCP-2/CCL8, and TNF-α —were associated with protective roles, whereas NKCR-2B4/CD244 and IL-12β were identified as risk factors. The study provides new insights into the complex interactions between gut microbiota, immune responses, and FM pathophysiology. The dysregulation of immune-inflammatory and compensatory immune-regulatory systems appears central to FM development, with potential links to viral pathways such as HHV-6 reactivation. In conclusion, this study underscores the importance of gut microbiota and immune modulation in FM, offering promising avenues for future therapeutic interventions.
Author Information
Prof. Dr. Michael Maes, M.D., Ph.D.
https://scholar.google.co.th/citations?user=1wzMZ7UAAAAJandhl=thandoi=ao
Highly cited author: 2003–2023 (ISI, Clarivate)
ScholarGPS: Worldwide #1 in molecular neuroscience; #1/4 in pathophysiology
Expert worldwide medical expertise ranking, Expertscape (December 2022), worldwide:
#1 in CFS, #1 in oxidative stress, #1 in encephalomyelitis, #1 in nitrosative stress, #1 in nitrosation,
#1 in tryptophan, #1 in aromatic amino acids, #1 in stress (physiological), #1 in neuroimmune;
#2 in bacterial translocation; #3 in inflammation, #4-5: in depression, fatigue and psychiatry.
Abbreviations
FM, Fibromyalgia; PBMCs, Peripheral Blood Mononuclear Cells; MR, Mendelian Randomization; SNPs, Single Nucleotide Polymorphisms; GWAS, Genome-Wide Association Studies; MAF, Minor Allele Frequencies; IVW, Inverse Variance Weighting; WM, Weighted Median; PPI, Protein-Protein Interaction; KEGG, Kyoto Encyclopedia of Genes and Genomes; GO, Gene Ontology; NKCR-2B4/CD244, Natural Killer Cell Receptor 2B4; IL-12β, Interleukin-12 Subunit Beta; MCP-2/CCL8, Monocyte Chemoattractant Protein 2; TNF-ɑ, Tumor Necrosis Factor Alpha; CXCL5, C-X-C Motif Chemokine 5; LIFR, Leukemia Inhibitory Factor Receptor; FDR, False Discovery Rate; SCFAs, Short-Chain Fatty Acids; NLRP3, NOD-Like Receptor Family Pyrin Domain Containing 3; MDD, Major Depressive Disorder; IRS, Immune-Inflammatory Response System; CIRS, Compensatory Immune-Regulatory Response System; RAGE, Receptor for Advanced Glycation End-Products; NF, Nuclear Factor.
Data Access Statement
The datasets used and/or analyzed during the current study are presented in the manuscript. Summary statistics for GWAS are publicly available.
Ethics approval and consent to participate
Our analyses were based on publicly available data that had been approved by relevant review boards. Therefore, no additional ethical approval is required.
Acknowledgment
The research community is thanked for making summary statistics from genome-wide association studies publicly available. The paper was pre-published as a preprint by MedrXiv (https://www.medrxiv.org/content/10.1101/2024.09.13.24313599v1).
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
There was no specific funding for this specific study.
Disclosure
The authors have no conflicts of interest with any commercial or other association in connection with the submitted article.
References
1. Wolfe F, Smythe HA, Yunus MB, et al. The American College of Rheumatology 1990 criteria for the classification of fibromyalgia. Report of the Multicenter Criteria Committee. Arthritis Rheum. 1990;33(2):160–172. doi:10.1002/art.1780330203
2. Sarzi-Puttini P, Giorgi V, Marotto D, Atzeni F. Fibromyalgia: an update on clinical characteristics, aetiopathogenesis and treatment. Nat Rev Rheumatol. 2020;16(11):645–660. doi:10.1038/s41584-020-00506-w
3. Galvez-Sánchez CM, Duschek S, Reyes del paso GA. Psychological impact of fibromyalgia: current perspectives. Psychol Res Behav Manage. 2019;12:117–127. doi:10.2147/PRBM.S178240
4. Favretti M, Iannuccelli C, Di Franco M. Pain biomarkers in fibromyalgia syndrome: current understanding and future directions. Int J Mol Sci. 2023;24(13):10443. doi:10.3390/ijms241310443
5. Hazra S, Venkataraman S, Handa G, et al. A cross-sectional study on central sensitization and autonomic changes in fibromyalgia. Front Neurosci. 2020;14:788. doi:10.3389/fnins.2020.00788
6. Andrés-Rodríguez L, Borràs X, Feliu-Soler A, et al. Peripheral immune aberrations in fibromyalgia: a systematic review, meta-analysis and meta-regression. Brain Behav Immun. 2020;87:881–889. doi:10.1016/j.bbi.2019.12.020
7. Mendieta D, De la Cruz-Aguilera DL, Barrera-Villalpando MI, et al. IL-8 and IL-6 primarily mediate the inflammatory response in fibromyalgia patients. J Neuroimmunol. 2016;290:22–25. doi:10.1016/j.jneuroim.2015.11.011
8. Bote ME, García JJ, Hinchado MD, Ortega E. An exploratory study of the effect of regular aquatic exercise on the function of neutrophils from women with fibromyalgia: role of IL-8 and noradrenaline. Brain Behav Immun. 2014;39:107–112. doi:10.1016/j.bbi.2013.11.009
9. Siracusa R, Paola RD, Cuzzocrea S, Impellizzeri D. Fibromyalgia: pathogenesis, mechanisms, diagnosis and treatment options update. Int J Mol Sci. 2021;22(8).
10. Kosek E, Altawil R, Kadetoff D, et al. Evidence of different mediators of central inflammation in dysfunctional and inflammatory pain--interleukin-8 in fibromyalgia and interleukin-1 β in rheumatoid arthritis. J Neuroimmunol. 2015;280:49–55. doi:10.1016/j.jneuroim.2015.02.002
11. Bote ME, García JJ, Hinchado MD, Ortega E. Inflammatory/stress feedback dysregulation in women with fibromyalgia. Neuroimmunomodulation. 2012;19(6):343–351. doi:10.1159/000341664
12. Imamura M, Targino RA, Hsing WT, et al. Concentration of cytokines in patients with osteoarthritis of the knee and fibromyalgia. Clin Interventions Aging. 2014;9:939–944. doi:10.2147/CIA.S60330
13. Mastrangelo F, Frydas I, Ronconi G, et al. Low-grade chronic inflammation mediated by mast cells in fibromyalgia: role of IL-37. J Biol Regul Homeost Agents. 2018;32(2):195–198.
14. Maes M, Almulla AF, Zhou B, Algon AAA, Sodsai P. In major dysmood disorder, physiosomatic, chronic fatigue and fibromyalgia symptoms are driven by immune activation and increased immune-associated neurotoxicity. Sci Rep. 2024;14(1):7344. doi:10.1038/s41598-024-57350-1
15. Maes M, Andrés-Rodríguez L, Vojdani A, Sirivichayakul S, Barbosa DS, Kanchanatawan B. In schizophrenia, chronic fatigue syndrome- and fibromyalgia-like symptoms are driven by breakdown of the paracellular pathway with increased zonulin and immune activation-associated neurotoxicity. CNS Neurol Disord Drug Targets. 2023;22(2):215–225. doi:10.2174/1871527321666220806100600
16. Asad HN, Al-Hakeim HK, Moustafa SR, Maes M. A causal-pathway phenotype of chronic fatigue syndrome due to hemodialysis in patients with end-stage renal disease. CNS Neurol Disord Drug Targets. 2023;22(2):191–206. doi:10.2174/1871527321666220401140747
17. Al-Hakeim HK, Khairi Abed A, Rouf Moustafa S, Almulla AF, Maes M. Tryptophan catabolites, inflammation, and insulin resistance as determinants of chronic fatigue syndrome and affective symptoms in long COVID. Front Mol Neurosci. 2023;16:1194769. doi:10.3389/fnmol.2023.1194769
18. Al-Jassas HK, Al-Hakeim HK, Maes M. Intersections between pneumonia, lowered oxygen saturation percentage and immune activation mediate depression, anxiety, and chronic fatigue syndrome-like symptoms due to COVID-19: a nomothetic network approach. J Affective Disorders. 2022;297:233–245. doi:10.1016/j.jad.2021.10.039
19. Maes M, Libbrecht I, Van Hunsel F, et al. The immune-inflammatory pathophysiology of fibromyalgia: increased serum soluble gp130, the common signal transducer protein of various neurotrophic cytokines. Psychoneuroendocrinology. 1999;24(4):371–383. doi:10.1016/S0306-4530(98)00087-0
20. Gregory NS, Brito RG, Fusaro M, Sluka KA. ASIC3 is required for development of fatigue-induced hyperalgesia. Mol Neurobiol. 2016;53(2):1020–1030. doi:10.1007/s12035-014-9055-4
21. Slade GD, Conrad MS, Diatchenko L, et al. Cytokine biomarkers and chronic pain: association of genes, transcription, and circulating proteins with temporomandibular disorders and widespread palpation tenderness. Pain. 2011;152(12):2802–2812. doi:10.1016/j.pain.2011.09.005
22. Sturgill J, McGee E, Menzies V. Unique cytokine signature in the plasma of patients with fibromyalgia. J Immunol Res. 2014;2014:938576. doi:10.1155/2014/938576
23. Littlejohn G, Guymer E. Neurogenic inflammation in fibromyalgia. Semin Immunopathol. 2018;40(3):291–300. doi:10.1007/s00281-018-0672-2
24. Sarapultsev A, Gusev E, Chereshnev V, Komelkova M, Hu D. Neuroimmune interactions in stress and depression: exploring the molecular and cellular mechanisms within the neuroinflammation-depression nexus. Curr Med Chem. 2024. doi:10.2174/0109298673320710240920055041
25. Maes M, Mihaylova I, Leunis JC. Increased serum IgA and IgM against LPS of enterobacteria in chronic fatigue syndrome (CFS): indication for the involvement of gram-negative enterobacteria in the etiology of CFS and for the presence of an increased gut-intestinal permeability. J Affective Disorders. 2007;99(1–3):237–240. doi:10.1016/j.jad.2006.08.021
26. Minerbi A, Gonzalez E, Brereton NJB, et al. Altered microbiome composition in individuals with fibromyalgia. Pain. 2019;160(11):2589–2602. doi:10.1097/j.pain.0000000000001640
27. Minerbi A, Fitzcharles MA. Gut microbiome: pertinence in fibromyalgia. Clin Exp Rheumatol. 2020;38 Suppl 123(1):99–104.
28. Palma-Ordóñez JF, Moreno-Fernández AM, Ramírez-Tejero JA, Durán-González E, Martínez-Lara A, Cotán D. Implication of intestinal microbiota in the etiopathogenesis of fibromyalgia: a systematic review. Int J Rheumatic Dis. 2024;27(1):e15021. doi:10.1111/1756-185X.15021
29. Smith GD, Ebrahim S. ‘Mendelian randomization’: can genetic epidemiology contribute to understanding environmental determinants of disease? Int J Epidemiol. 2003;32(1):1–22. doi:10.1093/ije/dyg070
30. Burgess S, Butterworth A, Thompson SG. Mendelian randomization analysis with multiple genetic variants using summarized data. Genetic Epidemiol. 2013;37(7):658–665. doi:10.1002/gepi.21758
31. Pierce BL, Burgess S. Efficient design for Mendelian randomization studies: subsample and 2-sample instrumental variable estimators. Am J epidemiol. 2013;178(7):1177–1184. doi:10.1093/aje/kwt084
32. Lawlor DA, Harbord RM, Sterne JA, Timpson N, Davey Smith G. Mendelian randomization: using genes as instruments for making causal inferences in epidemiology. Stat Med. 2008;27(8):1133–1163. doi:10.1002/sim.3034
33. Kurilshikov A, Medina-Gomez C, Bacigalupe R, et al. Large-scale association analyses identify host factors influencing human gut microbiome composition. Nat Genet. 2021;53(2):156–165. doi:10.1038/s41588-020-00763-1
34. Zhao JH, Stacey D, Eriksson N, et al. Genetics of circulating inflammatory proteins identifies drivers of immune-mediated disease risk and therapeutic targets. Nat Immunol. 2023;24(9):1540–1551. doi:10.1038/s41590-023-01588-w
35. Moscati A, Faucon AB, Arnaiz-Yépez C, et al. Life is pain: fibromyalgia as a nexus of multiple liability distributions. Am J Med Genet B Neuropsychiatr Genet. 2023;192(7–8):171–182. doi:10.1002/ajmg.b.32949
36. Li P, Wang H, Guo L, et al. Association between gut microbiota and preeclampsia-eclampsia: a two-sample Mendelian randomization study. BMC Med. 2022;20(1):443. doi:10.1186/s12916-022-02657-x
37. Lin X, Gao W, Huang C, Wu M, She X. Causal relationship between inflammatory proteins and glioblastoma: a two-sample bi‑directional mendelian randomization study. Front Genetics. 2024;15:1391921. doi:10.3389/fgene.2024.1391921
38. Sun W, Wu X, Yang H, et al. Identifying causal associations between women’s reproductive traits and risk of schizophrenia: a multivariate validated two-sample Mendelian randomization analysis. BMC Psychiatry. 2024;24(1):161. doi:10.1186/s12888-024-05614-5
39. Li YS, Xia YG, Liu YL, et al. Metabolic-dysfunction associated steatotic liver disease-related diseases, cognition and dementia: a two-sample mendelian randomization study. PLoS One. 2024;19(2):e0297883. doi:10.1371/journal.pone.0297883
40. Davies NM, Holmes MV, Davey Smith G. Reading Mendelian randomisation studies: a guide, glossary, and checklist for clinicians. BMJ. 2018;362:k601.
41. Burgess S, Thompson SG. Interpreting findings from Mendelian randomization using the MR-Egger method. Eur J Epidemiol. 2017;32(5):377–389. doi:10.1007/s10654-017-0255-x
42. Cohen JF, Chalumeau M, Cohen R, Korevaar DA, Khoshnood B, Bossuyt PM. Cochran’s Q test was useful to assess heterogeneity in likelihood ratios in studies of diagnostic accuracy. J Clin Epidemiol. 2015;68(3):299–306. doi:10.1016/j.jclinepi.2014.09.005
43. Verbanck M, Chen CY, Neale B, Do R. Detection of widespread horizontal pleiotropy in causal relationships inferred from Mendelian randomization between complex traits and diseases. Nat Genet. 2018;50(5):693–698. doi:10.1038/s41588-018-0099-7
44. Jia W, Tang Q, Zou Y, Yang Y, Wu W, Xu W. Investigating the antidepressant effect of Ziyan green tea on chronic unpredictable mild stress mice through fecal metabolomics. Front Microbiol. 2023;14:1256142. doi:10.3389/fmicb.2023.1256142
45. Gudi R, Suber J, Brown R, Johnson BM, Vasu C. Pretreatment with yeast-derived complex dietary polysaccharides suppresses gut inflammation, alters the microbiota composition, and increases immune regulatory short-chain fatty acid production in C57BL/6 mice. J Nutr. 2020;150(5):1291–1302. doi:10.1093/jn/nxz328
46. Li M, Yue H, Wang Y, et al. Intestinal microbes derived butyrate is related to the immunomodulatory activities of Dendrobium officinale polysaccharide. Int J Biol Macromol. 2020;149:717–723. doi:10.1016/j.ijbiomac.2020.01.305
47. Liu D, Zhang S, Li S, et al. Indoleacrylic acid produced by Parabacteroides distasonis alleviates type 2 diabetes via activation of AhR to repair intestinal barrier. BMC Biol. 2023;21(1):90. doi:10.1186/s12915-023-01578-2
48. Jia Z, Liu X, Liao W. Unraveling the association between gut microbiota and chemotherapy efficacy: a two-sample Mendelian randomization study. Microbiol Spectrum. 2024;12(8):e0394823. doi:10.1128/spectrum.03948-23
49. Shahi SK, Jensen SN, Murra AC, et al. Human commensal prevotella histicola ameliorates disease as effectively as interferon-beta in the experimental autoimmune encephalomyelitis. Front Immunol. 2020;11:578648. doi:10.3389/fimmu.2020.578648
50. Shahi SK, Freedman SN, Murra AC, et al. Prevotella histicola, a human gut commensal, is as potent as COPAXONE® in an animal model of multiple sclerosis. Front Immunol. 2019;10:462. doi:10.3389/fimmu.2019.00462
51. Xu T, Ge Y, Du H, et al. Berberis kansuensis extract alleviates type 2 diabetes in rats by regulating gut microbiota composition. J Ethnopharmacol. 2021;273:113995. doi:10.1016/j.jep.2021.113995
52. Crost EH, Coletto E, Bell A, Juge N. Ruminococcus gnavus: friend or foe for human health. FEMS Microbiol Rev. 2023;47(2). doi:10.1093/femsre/fuad014
53. Wu K, Luo Q, Liu Y, Li A, Xia D, Sun X. Causal relationship between gut microbiota and gastrointestinal diseases: a mendelian randomization study. J Transl Med. 2024;22(1):92. doi:10.1186/s12967-024-04894-5
54. Zhao H, Jin K, Jiang C, et al. A pilot exploration of multi-omics research of gut microbiome in major depressive disorders. Transl Psychiatry. 2022;12(1):8. doi:10.1038/s41398-021-01769-x
55. Maes M, Vasupanrajit A, Jirakran K, et al. Exploration of the gut microbiome in Thai patients with major depressive disorder shows a specific bacterial profile with depletion of the ruminococcus genus as a putative biomarker. Cells. 2023;12(9):1240. doi:10.3390/cells12091240
56. Simeonova D, Ivanovska M, Murdjeva M, Carvalho AF, Maes M. Recognizing the leaky gut as a trans-diagnostic target for neuroimmune disorders using clinical chemistry and molecular immunology assays. Curr Top Med Chem. 2018;18(19):1641–1655. doi:10.2174/1568026618666181115100610
57. Rudzki L, Maes M. The Microbiota-Gut-Immune-Glia (MGIG) axis in major depression. Mol Neurobiol. 2020;57(10):4269–4295. doi:10.1007/s12035-020-01961-y
58. Slyepchenko A, Maes M, Jacka FN, et al. Gut microbiota, bacterial translocation, and interactions with diet: pathophysiological links between major depressive disorder and non-communicable medical comorbidities. Psychother Psychosomatics. 2017;86(1):31–46. doi:10.1159/000448957
59. Maes M, Vasupanrajit A, Jirakran K, et al. Adverse childhood experiences and reoccurrence of illness impact the gut microbiome, which affects suicidal behaviours and the phenome of major depression: towards enterotypic phenotypes. Acta Neuropsychiatr. 2023;35(6):328–345. doi:10.1017/neu.2023.21
60. Wang LY, Tu YF, Lin YC, Huang CC. CXCL5 signaling is a shared pathway of neuroinflammation and blood-brain barrier injury contributing to white matter injury in the immature brain. J Neuroinflammation. 2016;13:6. doi:10.1186/s12974-015-0474-6
61. Vega-Ramírez MT, Becerril-Villanueva E, Maldonado-García JL, Pavón L, Pérez-Sánchez G. S100 proteins: a new frontier in fibromyalgia research. Molecular Brain. 2024;17(1):29. doi:10.1186/s13041-024-01102-9
62. Bazzichi L, Ciregia F, Giusti L, et al. Detection of potential markers of primary fibromyalgia syndrome in human saliva. Proteomics Clin Appl. 2009;3(11):1296–1304. doi:10.1002/prca.200900076
63. Wang Z, Meng Z, Chen C. Screening of potential biomarkers in peripheral blood of patients with depression based on weighted gene co-expression network analysis and machine learning algorithms. Front Psychiatry. 2022;13:1009911. doi:10.3389/fpsyt.2022.1009911
64. Maes M, Verkerk R, Bonaccorso S, Ombelet W, Bosmans E, Scharpé S. Depressive and anxiety symptoms in the early puerperium are related to increased degradation of tryptophan into kynurenine, a phenomenon which is related to immune activation. Life Sci. 2002;71(16):1837–1848. doi:10.1016/S0024-3205(02)01853-2
65. Maes M, Bocchio Chiavetto L, Bignotti S, et al. Increased serum interleukin-8 and interleukin-10 in schizophrenic patients resistant to treatment with neuroleptics and the stimulatory effects of clozapine on serum leukemia inhibitory factor receptor. Schizophr Res. 2002;54(3):281–291. doi:10.1016/S0920-9964(00)00094-3
66. Bäckryd E, Lind AL, Thulin M, Larsson A, Gerdle B, Gordh T. High levels of cerebrospinal fluid chemokines point to the presence of neuroinflammation in peripheral neuropathic pain: a cross-sectional study of 2 cohorts of patients compared with healthy controls. Pain. 2017;158(12):2487–2495. doi:10.1097/j.pain.0000000000001061
67. Costa Carvalho JL, de Brito AA, de Oliveira AP, et al. The chemokines secretion and the oxidative stress are targets of low-level laser therapy in allergic lung inflammation. J Biophotonics. 2016;9(11–12):1208–1221. doi:10.1002/jbio.201600061
68. Liu MN, Tsai SJ, Yeh HL, Wu CC, Lin CP. MCP-2/CCL8 level associated with suicidal ideation in elderly men with major depression. Arch Suicide Res. 2020;24(3):467–476. doi:10.1080/13811118.2019.1649772
69. Li Y, Ye R, Dai H, et al. Exploring TNFR1: from discovery to targeted therapy development. J Transl Med. 2025;23(1):71. doi:10.1186/s12967-025-06122-0
70. Hernandez ME, Becerril E, Perez M, et al. Proinflammatory cytokine levels in fibromyalgia patients are independent of body mass index. BMC Res Notes. 2010;3(1):156. doi:10.1186/1756-0500-3-156
71. Blanco I, Janciauskiene S, Nita I, et al. Low plasma levels of monocyte chemoattractant protein-1 (MCP-1), tumor necrosis factor-alpha (TNFalpha), and vascular endothelial growth factor (VEGF) in patients with alpha1-antitrypsin deficiency-related fibromyalgia. Clin Rheumatol. 2010;29(2):189–197. doi:10.1007/s10067-009-1318-5
72. Tsilioni I, Russell IJ, Stewart JM, Gleason RM, Theoharides TC. Neuropeptides CRH, SP, HK-1, and inflammatory cytokines IL-6 and TNF are increased in serum of patients with fibromyalgia syndrome, implicating mast cells. J Pharmacol Exp Ther. 2016;356(3):664–672. doi:10.1124/jpet.115.230060
73. Bazzichi L, Rossi A, Massimetti G, et al. Cytokine patterns in fibromyalgia and their correlation with clinical manifestations. Clin Exp Rheumatol. 2007;25(2):225–230.
74. Ellergezen P, Alp A, Çavun S, Çelebi M, Macunluoğlu AC. Pregabalin inhibits proinflammatory cytokine release in patients with fibromyalgia syndrome. Archi Rheumatol. 2023;38(2):307–314. doi:10.46497/ArchRheumatol.2023.9517
75. Feng J, Zhang Z, Wu X, et al. Discovery of potential new gene variants and inflammatory cytokine associations with fibromyalgia syndrome by whole exome sequencing. PLoS One. 2013;8(6):e65033. doi:10.1371/journal.pone.0065033
76. Verri WA, Cunha TM, Parada CA, et al. IL-15 mediates immune inflammatory hypernociception by triggering a sequential release of IFN-gamma, endothelin, and prostaglandin. Proc Natl Acad Sci USA. 2006;103(25):9721–9725. doi:10.1073/pnas.0603286103
77. Verri WA, Cunha TM, Parada CA, et al. Antigen-induced inflammatory mechanical hypernociception in mice is mediated by IL-18. Brain Behav Immun. 2007;21(5):535–543. doi:10.1016/j.bbi.2006.11.005
78. Sagulkoo P, Plaimas K, Suratanee A, Colado Simão AN, Vissoci Reiche EM, Maes M. Immunopathogenesis and Immunogenetic Variants in COVID-19. Curr Pharm Des. 2022;28(22):1780–1797. doi:10.2174/1381612828666220519150821
79. Brunoni AR, Supasitthumrong T, Teixeira AL, et al. Differences in the immune-inflammatory profiles of unipolar and bipolar depression. J Affective Disorders. 2020;262:8–15. doi:10.1016/j.jad.2019.10.037
80. Cataldi C, Mari NL, Lozovoy MAB, et al. Proinflammatory and anti-inflammatory cytokine profiles in psoriasis: use as laboratory biomarkers and disease predictors. Inflamm Res. 2019;68(7):557–567. doi:10.1007/s00011-019-01238-8
81. Maes M, Ruckoanich P, Chang YS, Mahanonda N, Berk M. Multiple aberrations in shared inflammatory and oxidative & nitrosative stress (IO&NS) pathways explain the co-association of depression and cardiovascular disorder (CVD), and the increased risk for CVD and due mortality in depressed patients. Prog Neuro Psychopharmacol Biol Psychiatry. 2011;35(3):769–783.
82. Guimarães PM, Scavuzzi BM, Stadtlober NP, et al. Cytokines in systemic lupus erythematosus: far beyond Th1/Th2 dualism lupus: cytokine profiles. Immunol Cell Biol. 2017;95(9):824–831. doi:10.1038/icb.2017.53
83. Maes M, Almulla AF, Tang X, Stoyanova K, Vojdani A. From human herpes virus-6 reactivation to autoimmune reactivity against tight junctions and neuronal antigens, to inflammation, depression, and chronic fatigue syndrome due to Long COVID. J Med Virol. 2024;96(8):e29864. doi:10.1002/jmv.29864
84. Krumina A, Chapenko S, Kenina V, et al. The role of HHV-6 and HHV-7 infections in the development of fibromyalgia. J Neurovirol. 2019;25(2):194–207. doi:10.1007/s13365-018-0703-8
85. Vojdani A, Vojdani E, Saidara E, Maes M. Persistent SARS-CoV-2 Infection, EBV, HHV-6 and Other Factors May Contribute to Inflammation and Autoimmunity in Long COVID. Viruses. 2023;15(2):400. doi:10.3390/v15020400
86. Liu CC, Gao YJ, Luo H, et al. Interferon alpha inhibits spinal cord synaptic and nociceptive transmission via neuronal-glial interactions. Sci Rep. 2016;6:34356. doi:10.1038/srep34356
87. Tsuda M, Masuda T, Kitano J, Shimoyama H, Tozaki-Saitoh H, Inoue K. IFN-gamma receptor signaling mediates spinal microglia activation driving neuropathic pain. Proc Natl Acad Sci USA. 2009;106(19):8032–8037. doi:10.1073/pnas.0810420106
88. He H, Zhang X, He H, et al. Microglial priming by IFN-γ involves STAT1-mediated activation of the NLRP3 inflammasome. CNS Neurosci Ther. 2024;30(10):e70061. doi:10.1111/cns.70061
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.