Back to Journals » Infection and Drug Resistance » Volume 13
Cucurbita pepo Seed Oil Induces Microsatellite Instability and Tegumental Damage to Schistosoma mansoni Immature and Adult Worms In vitro
Authors Ammar AI ![]() , Afifi AF, Essa A
, Afifi AF, Essa A ![]() , Galal-Khallaf A
, Galal-Khallaf A ![]() , Mokhtar MM, Shehab-Eldeen S
, Mokhtar MM, Shehab-Eldeen S ![]() , Rady AA
, Rady AA
Received 13 June 2020
Accepted for publication 17 August 2020
Published 7 October 2020 Volume 2020:13 Pages 3469—3484
DOI https://doi.org/10.2147/IDR.S265699
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor Suresh Antony
Amany Ibrahim Ammar, 1 Amira Fathy Afifi, 1 Abdallah Essa, 2, 3 Asmaa Galal-Khallaf, 4 Mostafa Mohamed Mokhtar, 5 Somaia Shehab-Eldeen, 2, 3 Amany Ahmed Rady 1
1Department of Medical Parasitology, Faculty of Medicine, Menoufia University, Shebin El Kom, Menoufia, Egypt; 2Tropical Medicine Department, Faculty of Medicine, Menoufia University, Shebin El Kom, Menoufia, Egypt; 3Internal Medicine Department, College of Medicine, King Faisal University, Al-Ahsaa, Saudi Arabia; 4Molecular Biology and Biotechnology Laboratory, Department of Zoology, Faculty of Science, Menoufia University, Shebin El- Kom, Menoufia, Egypt; 5Department of Zoology and Entomology, Faculty of Science, Al-Azhar University, Nasr City, Egypt
Correspondence: Abdallah Essa
Faculty of Medicine, Menoufia University, Yassen Abd Al Ghafar Street, Shebin Elkom City 32511, Menoufia Governorate, Egypt
Tel +20-10-2242-4341
Email [email protected]
Background: Schistosomiasis is a tropical parasitic disease treated exclusively with praziquantel (PZQ). However, PZQ has low efficacy against schistosomula and juveniles. In addition, the emergence of PZQ resistance has prompted the search for new drugs.
Methods: This study investigated the effects of pumpkin (Cucurbita pepo)-seed oil (PSO) on Schistosoma mansoni adults, juveniles, and newly formed schistosomula in vitro by exposing the parasites to increasing concentrations of PSO (20, 40, 60, 80, and 100 μl/mL) with variable incubation periods (24, 48, and 72 hours). Dose–response effects of PSO on mortality rate, worm activity, and tegumental changes were studied. Also, effect on DNA were assessed with microsatellite analysis.
Results: All tested stages of S. mansoni were susceptible to PSO, which was more effective than PZQ on juvenile worms and schistosomula. Juveniles and schistosomula S. mansoni were more sensitive to the antischistosomal activity of PSO than adult worms. PSO showed evident changes in the integuments of adults, juveniles, and schistosomula. These changes were more evident with increased concentrations. At the genomic level, PSO induced clear qualitative and quantitative changes in the microsatellite loci R95529 and SMD57 of S. mansoni adults and schistosomula. This microsatellite instability is being reported through the current study for S. mansoni in response to PSO for the first time.
Conclusion: This study suggested that PSO possesses effective antischistosomal activity against various stages of S. mansoni. Further investigations are needed to figure out the mechanism of action of PSO on this parasite.
Keywords: Schistosoma mansoni, Cucurbita pepo, praziquantel, tegumental alteration, microsatellite
Introduction
Schistosomiasis is a tropical parasitic disease caused by blood flukes of the genus Schistosoma.1 More than 700 million individuals are in danger of contracting schistosomiasis worldwide and more than 200 million are currently infected, making this an urgent global health concern.2 There is no available vaccine against schistosomiasis, and the infection is treated solely with praziquantel (PZQ), which is advised by the World Health Organization (WHO) for schistosomiasis treatment.3 Regrettably, repeated use of PZQ for long periods, especially in mass drug administration, has increased the potential for drug resistance.4 Low activity against immature schistosomula is another drawback with PZQ.5 It is thus an urgent priority to find a novel drug to substitute or potentiate PZQ.
Herbal extracts are an important source of new drugs, due to their variety of biomedical activities and chemical components.6 The Cucurbitaceae plant family, collectively known as cucurbits, comprises 130 genera and around 800 species.7 Pumpkin (Cucurbita pepo) seed oil (PSO) is a remarkably rich source of various bioactive components with functional properties. The seeds are used for an edible oil and as a nutraceutical product. In addition, PSO has been recognized to have therapeutic properties as an antibacterial, antioxidant and anti-inflammatory agent.8, 9 Traditional described medicinal uses of PS have prompted experimental studies on its anthelmintic properties.10 Studies on the effect of PS on heterophyiasis and taeniasis have shown promising results.11 However, such studies are few in number, and they still leave many questions to be answered.
Microsatellite loci are highly variable DNA markers that are commonly used in schistosomiasis research and allow population-level analysis.12 As such, it could be useful to use these microsatellites in assessing the variability occurring in DNA of the parasites when exposed to drugs. The present work was conducted to study the effect of PSO on S. mansoni adults, juveniles, and newly formed schistosomula in vitro.
Methods
Schistosoma mansoni Maintenance
Ten adult male CD1 Swiss albino mice 6–8 weeks of age weighing 20±2 g were purchased from the Schistosome Biological Supply Centre (SBSC), Theodor Bilharz Research Institute (TBRI), Egypt. They were maintained on a standard commercial pellet diet and kept in an air-conditioned animal house at 20°C–22°C for 1 week before infection, in accordance with university regulations. Each mouse was infected with 70±5 cercariae of an Egyptian S. mansoni strain by using the tail-immersion technique, as described by Oliver and Stirewalt.13 Cercariae had been freshly shed from experimentally infected Biomphalaria alexandrina and used within 1 hour.
Parasite Preparation
Juvenile worms of S. mansoni were obtained from hepatic veins at 21 days postinfection, while adult worms were collected from the portal system and mesenteric veins of infected mice by perfusion 7 weeks later.14 Juvenile and adult worms were washed with PBS two to three times and undamaged worms picked out. They were placed in RPMI 1640 medium (Sigma-Aldrich) and stored in an incubator chamber at 37°C for 6 hours for adaptation before starting the experiment. Schistosomula were prepared in vitro according to the method of Colley and Wikel.15 The newly transformed schistosomula (NTS) were resuspended in preheated RPMI 1640 medium containing 10% heat-inactivated FBS, streptomycin (100 μg/mL), and penicillin (100 IU/mL) and incubated overnight in 5% CO2 and 100% humidity at 37°C.
Praziquantel and Pumpkin-Seed Oil
PZQ (Discocide 600 mg; EIPICO, Egypt) and Cucurbita pepo seed oil (Now PSO 1,000 mg; Now Health Group, USA) was mixed in dimethyl sulfoxide (DMSO; Sigma-Aldrich). A stock solution of 1 mL PSO was dissolved in 1%DMSO and then diluted with RPMI 1640 to prepare 2 mL of working solution (100 µl/mL).
In vitro Assessment of Antischistosomal Effect of PSO
In vitro evaluation was done on different stages of S. mansoni schistosomula, juveniles, and adults.
Mortality Rate, Worm Activity, and Morphological Changes
As per Xiao et al,16 juvenile worms were collected, washed with RPMI 1640, counted, and relocated to 24-well culture plates (TPP, St Louis, MO, USA), with 100 juveniles in each well containing 2 mL RPMI 1640 medium enriched with 2 mM L-glutamine, 20% FBS (Gibco), and antibiotics (penicillin, gentamycin, and streptomycin: 300 IU/mL, 160 μg/mL, and 300 μg/mL, respectively). Then, plates were incubated in 5% CO2 at 37°C. The same was done with adult worms, with equal representation of males and females in each well. Numbers of adults in each well for untreated, PZQ-treated, PSO (100), PSO (80), PSO (60), PSO (40), and PSO (20) samples were 12, 20, 28, 24, 30, 28, and 24, respectively. PSO was tested at concentrations of 20, 40, 60, 80, and 100 µL/mL. Experiments were conducted in triplicate and read at 24, 48, and 72 hours, then repeated at least twice. Two groups of control worms were used. The negative-control group was assayed in RPMI 1640 medium containing 1% DMSO without drugs, and the positive-control group with 10 μg/mL PZQ, the most effective schistosomicidal drug, as reference drug.
Schistosomula obtained by mechanical transformation were carefully counted and adjusted to a concentration of 100 NTS/50 µL,17 then placed in RPMI 1640 medium, as previously described for juvenile worms. The plates were incubated in 5% CO2 at 37°C for a minimum of 12–24 hours to achieve complete transformation into schistosomula. Then, PSO was tested in the same concentrations tested in adult worms and read at the same time points and the procedure repeated twice. Control NTS were assayed in RPMI 1640 medium containing 1% DMSO without drugs as the negative-control group and with 10 μg/mL PZQ as the positive-control group.16
Assessment of the schistosomicidal effect of PSO was carried out by determining worm motor activity and mortality rate using inverted microscopy at 40× and 100× magnification (IX70; Olympus, Tokyo, Japan).18 Worms that did not show any sign of motility for 2 minutes were counted as dead. To quantify the degree of motility and morphological changes, values assigned were 3 = normal motility, no morphological changes; 2 = reduced motility and/or moderate damage to integument noted; 1 = severe reduction in motility and/or severe morphological/tegumental changes; 0 = death of all worms.19
The same experiment was repeated six times to determine lethal concentrations (LC50 and LC90). Worm mortality was counted in each case, and LC50 and LC90 values of PSO were calculated using probit analysis and SPSS version 20. For each concentration, the worm-mortality rate was recorded by computing the number of dead worms divided by the total.20,21 Since each experiment was repeated three times, means and standard deviations were calculated for each concentration at different times.
Scanning Electron Microscopy
Morphological changes in the integument of NTS, juveniles, and male adults were observed by scanning electron microscopy (SEM) as per Hassan et al.22 SEM was performed on worms that had been exposed to PSO at 60 and 100 µL/mL for 72hours. Briefly, worms were washed in normal saline several times, fixed with 2.5% glutaraldehyde in 0.2 M sodium cacodylate buffer (pH 7.3) for 4hours, then fixed with osmium tetraoxide (OSO4) for 2 hours. That was followed by three washes in 0.2 M sodium cacodylate buffer and dehydration in gradual concentrations of ethanol (10%–100%, 10 minutes for each concentration). Further dehydration was performed in liquid carbon dioxide. Mounting of worms was done on copper stubs with double-sided adhesive tape. They were gold-coated using an S150A sputter coater, then viewed by SEM (JEOL JSM-5400) at the National Center for Research, Radiation, and Technology, Cairo, Egypt.
DNA-Microsatellite Analysis
Frozen pools of Schistosoma subjected to the different treatments were transferred to the Molecular Biology Laboratory, Zoology Department, Faculty of Science, Menoufia University. Each pool of Schistosoma specimens, placed in 1.5 μL Eppendorf tubes, was cleaned to remove traces of oils using the protocol applied by Galal-Khallaf et al23 and Ardura et al.24 In brief, each pool was rinsed with a mixture of water, methanol, and chloroform (0.8:2:1) for 2 hours, followed by another wash in distilled water and a final one in PBS. Three pools of worms per treatment, 30 mg each, were each subjected to total genomic DNA extraction.
DNA extraction was performed using the protocol described by Wasko et al25 and modified by Mohammed-Geba et al.26 Briefly, 2.4 U/mL proteinase K solution (Thermo Fisher Scientific) and 100 μL TNES–urea buffer (10 mM Tris HCl pH 8, 125 mM NaCl, 10 mM EDTA pH 8, 0.5% sodium dodecyl sulfate, and 4 M urea) was added to each sample. The mixture was incubated at 55°C for 60 minutes with occasional vortexing until completely lysed. Later, 5 M NaCl (27 μL) was added, with mixing of the tubes and centrifugation at 4,000 g for 10 minutes. The supernatant was placed into a clean 1.5 mL Eppendorf tube. DNA was precipitated by adding 100 µL freezer-cold isopropanol with shaking. Centrifugation of the tubes was done at 11,000 g for 10 minutes, with complete removal of the supernatant. Washing of DNA pellets was done in 70% ethanol (200 µL), followed by centrifugation at 11,000 g for 5 minutes, then after the ethanol had dried, 30 µL of Tris–EDTA buffer (10 mM Tris HCl pH 8, 2 mM EDTA pH 8) were added for pellet resuspendion. To check the quality of the DNA, 5 µL extracted DNA with 1 µL of 6x DNA loading buffer (bromophenol blue 0.25% w/v, sucrose 40% w/v) was run on agarose gels (1%) stained with ethidium bromide (0.5 μg/mL). Purity and quantity of the DNA were detected by spectrophotometric measurement for light absorbance at 260 nm and 280 nm.
For detecting different microsatellite loci, the primers identified by Durand et al27 were used, namely SMD57, R95529, SCMSMOXII, SMU31768, and SMIMP25. Primer sequences used are shown in Table 1. Amplification reactions were set up in a total volume of 15 μL. The reaction mixture was composed of 0.5 μL DNA template (100 ng), 0.25 μM each primer, 1X MyTaq red master mix (Bioline), and 0.2 mg/mL BSA, and reached 15 μL with PCR-grade water. PCR amplification was done in a TPersonal 48 thermal cycler (Biometra). Thermocycler conditions followed Durand et al: initial denaturation at 94°C for 4 minutes, followed by 35 cycles of 94°C for 30 seconds, annealing for 30 seconds, 72°C for 30 seconds, and an extension of 72°Cfor 10 minutes as a final step. Electrophoresis of the PCR products was performed in 2% agarose gel.
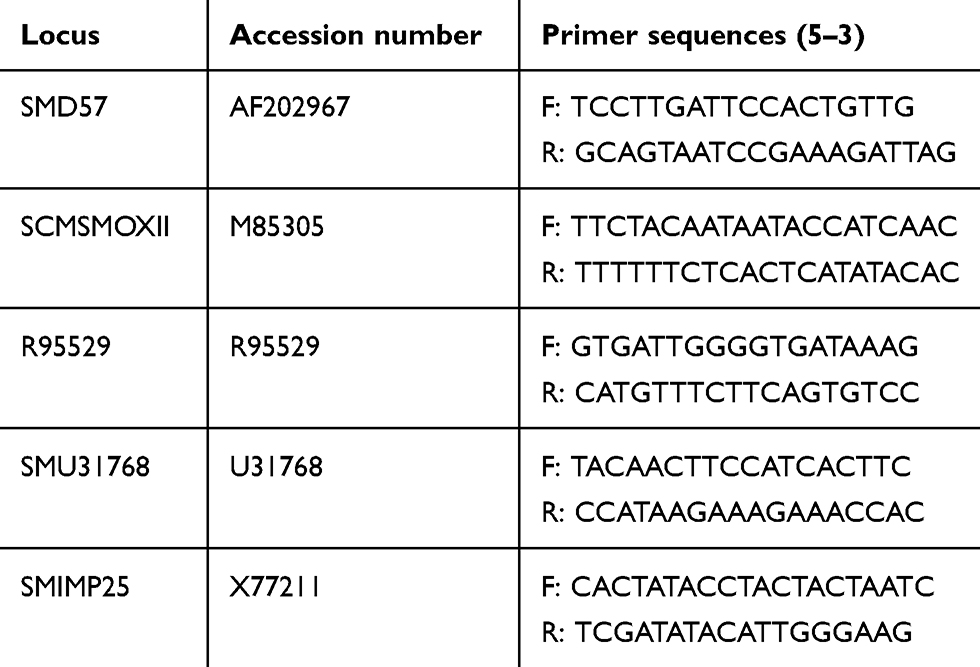 |
Table 1 Sequences of primers of S. mansoni applied in the current study (Durand et al 2000). GenBank accession numbers for these loci are also shown |
Ethical Considerations
The study was approved by the ethics committee of the Faculty of Medicine, Menoufia University, Egypt (approval protocol FWA00005112), and was in compliance with guidelines for the care and use of laboratory animals.28 Experiments were carried out at SBSC/TBRI, while DNA extraction and PCR procedures were carried out at the Molecular Biology Laboratory, Zoology Department, Faculty of Science, Menoufia University.
Statistical Analysis
Data were analyzed with SPSS 23.0 (IBM, Armonk, NY, USA). Descriptive statistics are expressed as numbers and percentages for nonnumeric data. For analytical statistics, χ2 was used to determine the association between qualitative variables and Z-tests to compare two proportions in two groups. Data were considered statistically significant at 0.05. These results were used to define LC50 and LC90 values of extract using probit analysis and SPSS.
Results
In vitro Antischistosomal Effects of PSO on Mortality Rate and Worm Activity
Adult Worms
Adult worms exposed to PZQ 10 μg/mL showed 100% mortality, with contraction and paralysis after 24 hours’ exposure. On exposure to PSO, no worm mortality was observed at any concentration after 24 hours’ exposure. The mortality rate increased with increased duration of exposure, and occurred in a dose-dependent manner. At 100 µL/mL, the mortality rate was 100% after 48 and 72 hours’ exposure. The lowest mortality rate was seen at 20 µL/mL: 25% and 37.5% at 48 and 72 hours, respectively, whereas no mortality was noticed in worms of the negative-control group. There was a highly significant difference in mortality rate among the PSO concentrations (80, 60, 40, 20 µL/mL) and PZQ (P2–5, all <0.001; Table 2). Concentrations of PSO required for LC50 and LC90 in adult worms in vitro after 72 hours were 36.94 µl/mL and 66.73 µL/mL, respectively.
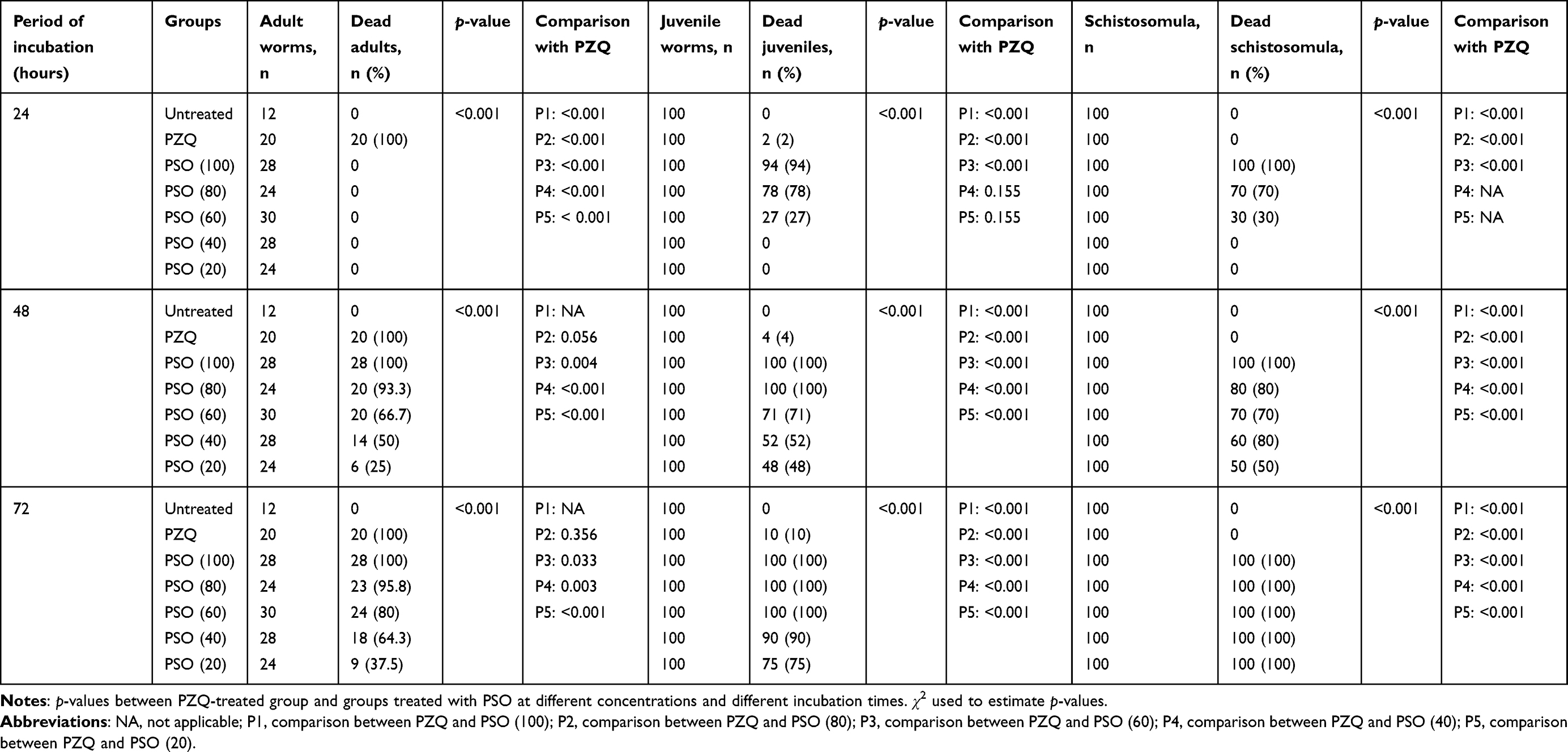 |
Table 2 In vitro effects of PSO and PZQ on mortality of adult worms, juveniles and schistosomula of S. mansoni after different incubation periods |
After 72 hours’ exposure to PSO, the drug had a deadlier effect on females than on males. At 80 and 100 µL/mL, all female worms died, while in male worms mortality rates were 91.7% and 100%, respectively, with no significant difference between males and females (Figure 1).Adult S. mansoni motility was remarkably affected by rising concentrations of PSO. Significant reductions in motility and severe morphological changes (contraction and shrinkage) were observed after 48 and 72 hours’ exposure with 60, 80, and 100 µL/mL PSO (Figure 2). Bodies of adult worms were poorly flexible, and suckers became less contractile. With lower concentrations (20 and 40 µL/mL), all worms showed normal motor activity at 24 hours’ exposure, while reduced motility and minimal alteration in morphology, such as stretched head end, slight elongation, and focal swelling of the worm body, were seen at 48 and 72 hours’ exposure. The positive-control group exposed to PZQ 10 μg/mL showed significant reductions in worm motility and marked changes in morphology. All worms acquired a coiled appearance after 24 hours’ exposure, along with spasmodic contractions and feeble movement of the tail portion and oral suckers and vesiculation along the surface of the worm. All S. mansoni adult worms in the negative-control group showed characteristic wavy and peristaltic movement along the body axis without morphological alterations (Table 3).
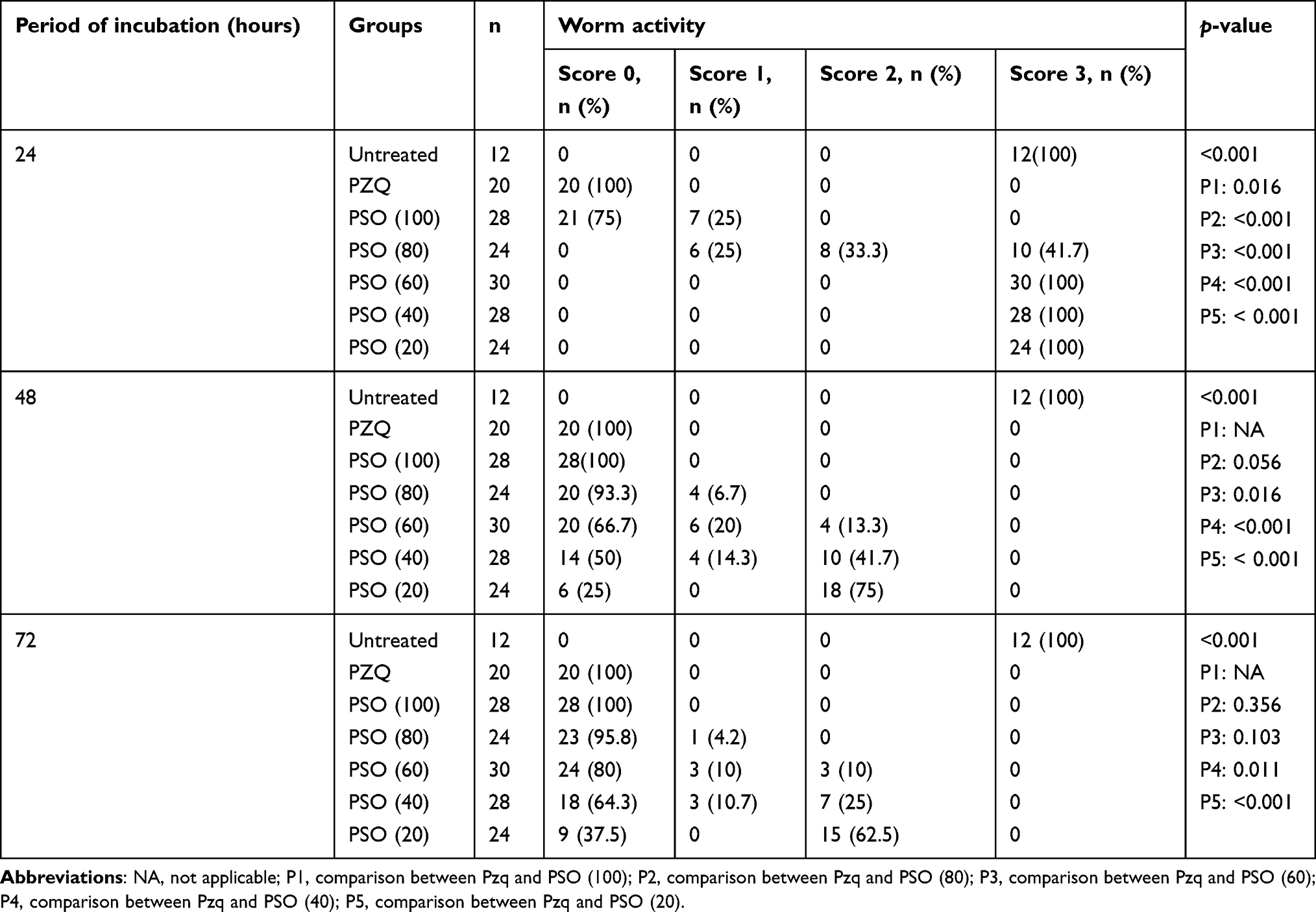 |
Table 3 Activity of adult Schistosoma mansoni in PSO- and PZQ-treated groups after different incubation periods |
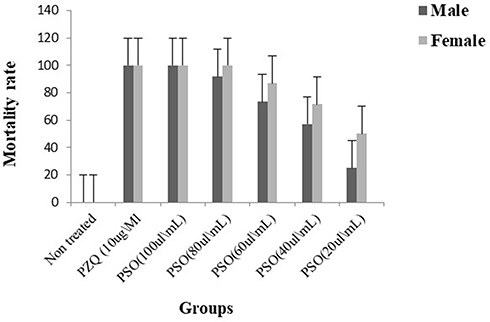 |
Figure 1 In vitro mortality rate of adult male and female Schistosoma mansoni of differently treated groups at 72 hours’ exposure. |
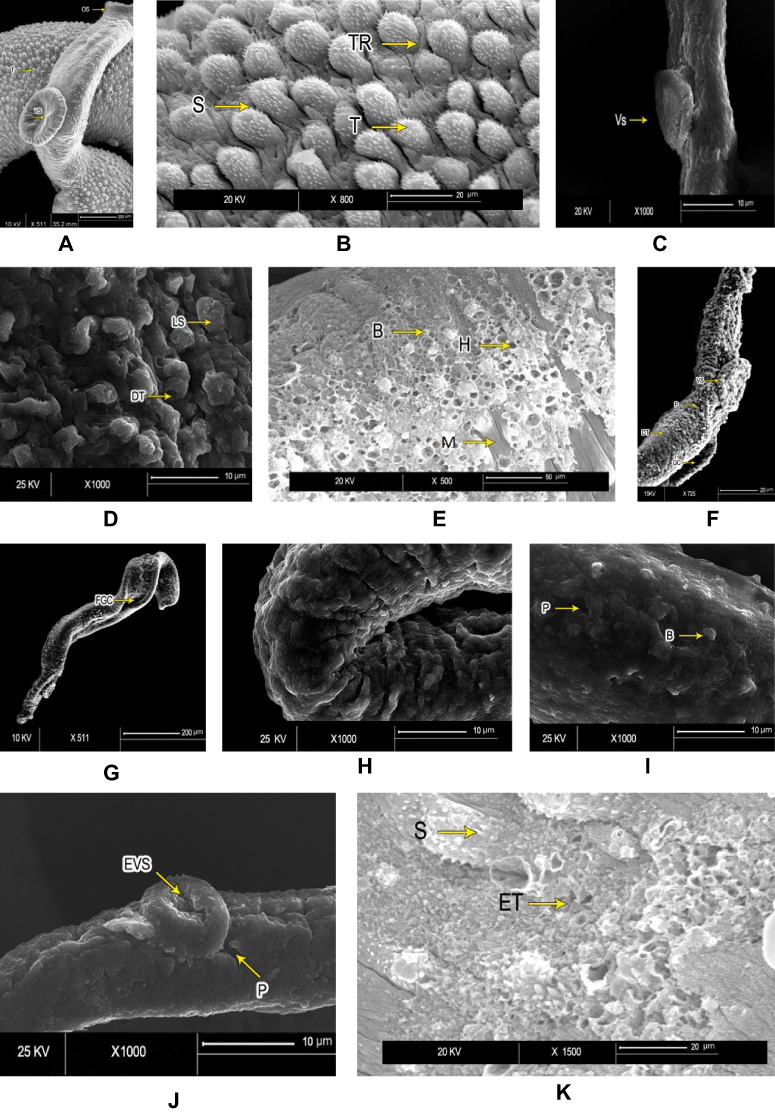 |
Figure 2 Scanning electron microscopy of Schistosoma mansoni adult male worminteguments. After in vitro incubation for 72 hours in medium with 1% DMSO (negative control group): (A) intact oral suckers (OS), ventral suckers (VS), and well-developed tubercles (T). Bar 200 µm; (B) higher magnification showing well-developed tubercles (T), spines (S), and tegumental ridges (TR) on the dorsal surface (bar 20 µm). PZQ-treated worms after 72 hours’ incubation: (C) anterior ends exhibited deformity of ventral suckers (VS) (bar 10 µm); (D) distorted tubercles (DT) with loss of spines (LS) and areas of surface peeling (bar 10 µm) (E); and severe erosion of tubercles with multiple holes (H), bleb formation (B), and exposed muscle layer (M) in some (bar 50 µm). Worms exposed to PSO for 72 hours at 60 µg/mL concentration: (F) destruction of integument (DT) and loss of normal tubercular pattern, with peeling areas (P) and deformity of ventral suckers (VS) (bar 20 µm); (G) and distortion and flattening of gynecophoric canal (FGC). bar 200 µm). At higher PSO concentration (100 µg/mL): (H) worms showed edematous integuments with more damage (bar 10 µm); (I) deformity of tubercles, loss of spines, and tegumental ridges, with formation of blebs (B) and peeling (P; bar 10 µm); (J) deformity and edematous ventral suckers (EVS) with tegumental peeling (P), laceration, complete loss of spines, and destruction of the integument (bar 10 µm); and (K) erosion, marked laceration, loss of spines (S), and extermination of tubercles (ET) with bleb formation (bar 20 µm). |
Immature Juvenile Worms
After incubation with PSO at different concentrations, the mortality rate of juvenile S. mansoni worms depended directly on the concentration evaluated and the incubation period. Concentrations of 80 and 100 µL/mL of PSO had proved lethal to 100% of juvenile worms at 48 hours, while at 20, 40, and 60 μL/mL the mortality rate in juvenile worms was 48%, 52%, and 71%, respectively after 48 hours. At the end of the experiment (72 hours), the mortality rate had increased to 100% at 60, 80, and 100 µL/mL PSO. The positive-control group exposed to PZQ 10 μg/mL showed slow motility and had a mortality rate of 4% and 10% after 24 and 72 hours, respectively, in juvenile worms (Table 2). These results were all highly significant (p<<0.001).
Regarding changes in worm activity, the motility of juvenile S. mansoni was remarkably affected by increasing concentrations of PSO. Significant reductions in motility and severe morphological changes in the form of abnormal body posture, elongation, and shrunken suckers were detected at higher concentrations of PSO (60, 80, and 100 µL/mL), followed by death of all worms after 72 hours’ exposure (p<0.001). At the lower concentration of 40 µL/mL, slight morphological changes and reduced motility were detected in 91% (score 2) after 24 hours’ exposure. At the end of the experiment (72 hours), marked reductions in motility and morphological changes were seen even at lower concentrations (p<0.001, Table 4).
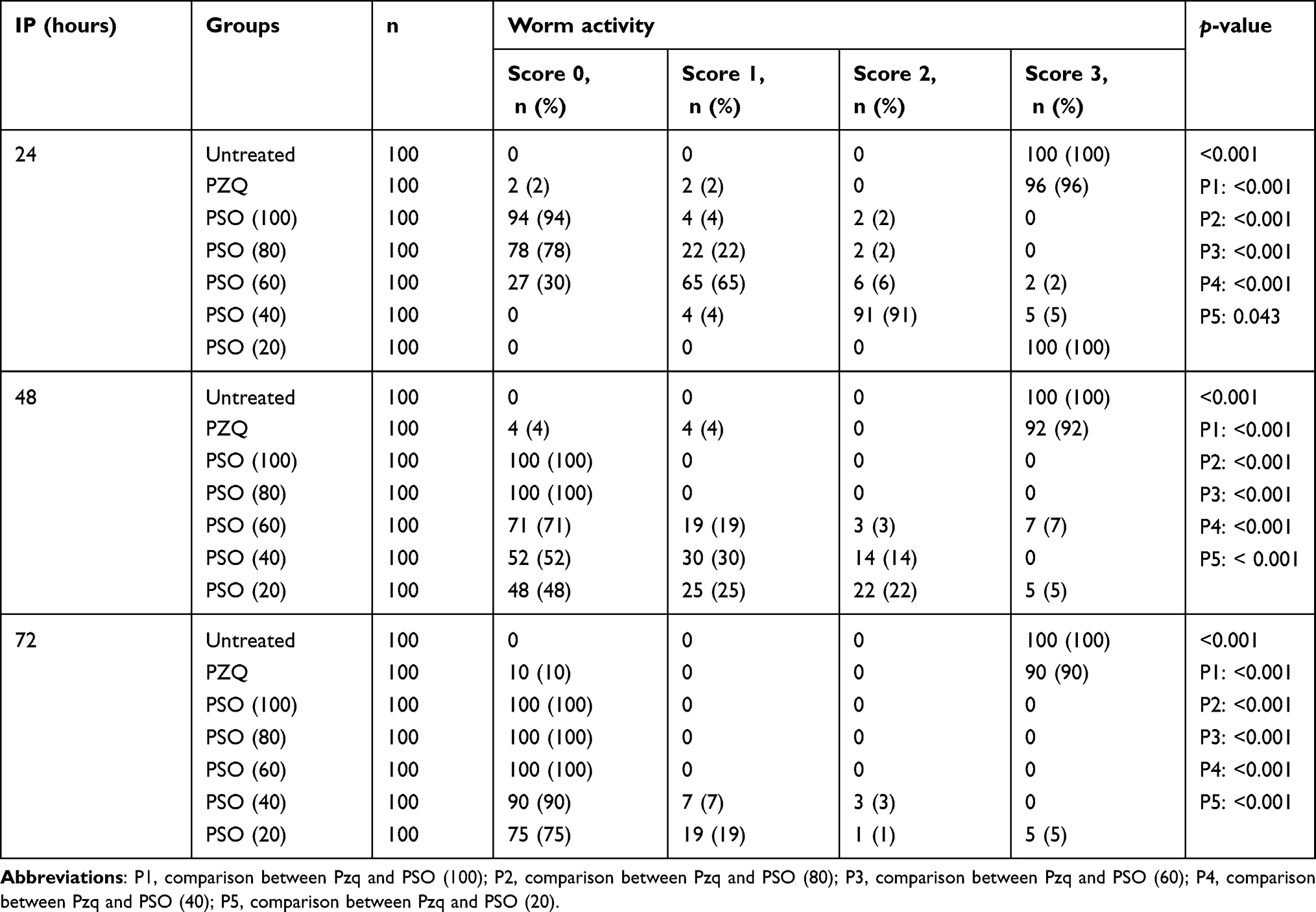 |
Table 4 Activity of immature juvenile Schistosoma mansoni in PSO- and PZQ-treated groups after different incubation periods (IPs) |
Schistosomula
All schistosomula died at concentrations of 100 µL/mL PSO, regardless of length of exposure. Worms in the control group remained viable until the end of the experiment. The lethal effect of PSO was dose-dependent and related to the duration of exposure. At lower concentrations (20 and 40 µL/mL), the viability of schistosomula was not affected (zero mortality rate) after 24hours while at 72 hours all schistosomula had died (100%), regardless of PSO concentration. No death was detected after treatment of schistosomula with PZQ at concentrations as high as 10 μg/mL (p<0.001, Table 2).
As seen in Table 5, schistosomula of control groups showed normal activity without any morphological changes till the end of the experiment (72 hours). Following incubation with 100 µL/mL PSO, schistosomula died rapidly: no movement was noticed at 3–4 minutes after incubation. Morphological changes like dark granular, crescent-shaped bodies and reduction in body length were seen regardless of exposure length. At lower concentrations (20 and 40 µL/mL), slight morphological changes, such as shorter bodies and reduced activity were observed after 24 hours’ incubation. At this time point, schistosomula incubated with 80 and 60 µL/mL showed reduced activity, with morphological alteration in 30% and 70%, respectively, which become marked with increased incubation time. In groups exposed to PZQ, no effect was observed at 10 µL/mL throughout the experiment. These results were statistically significant in comparison to PZQ as drug control (p<0.001).
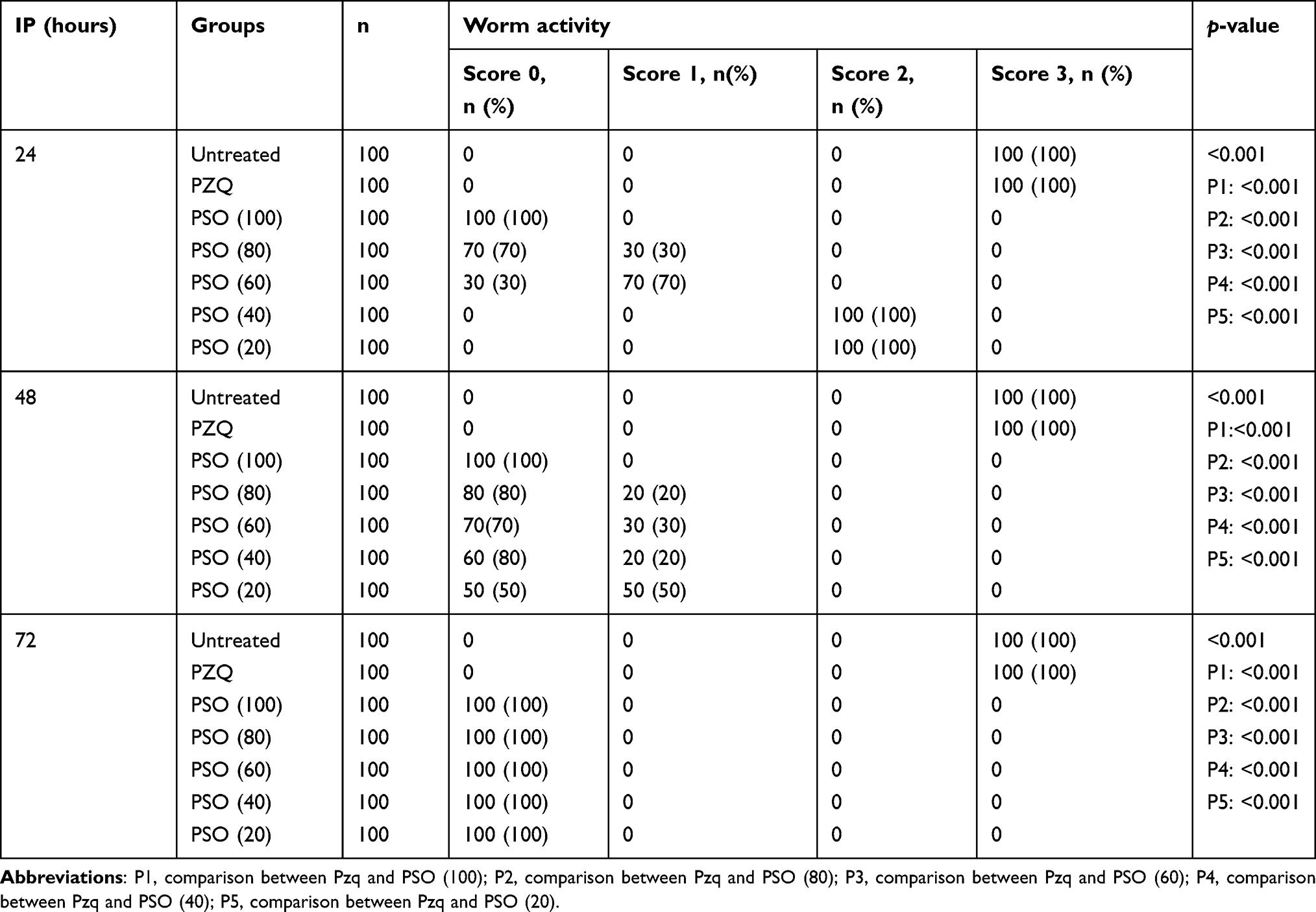 |
Table 5 Activity of Schistosoma mansoni schistosomula in PSO- and PZQ treated groups after different incubation periods (IPs) |
Ultrastructural Observations
All worms examined except those in the untreated control groups, showed changes of variable degrees. SEM of control adult male S. mansoni showed intact integuments with preserved surface architecture (Figure 2A and B). Worms exposed to PZQ for 72 hours revealed deformities of suckers, distorted tubercles with erosion, and peeling of the surface (Figure 2C–E). Adult worms exhibited destruction of integuments with loss of normal pattern and distortion of the gynecophoric canal after 72 hours’ exposure to PSO 60 µl/mL (Figure 2F, and G). At the higher concentration of 100 µg/mL, there was more damage to the integuments, with edema, lacerations, and formation of blebs (Figure 2H–K). Juveniles from control groups showed intact, well developed oral and ventral suckers with primitive ill-developed gynecophoric canals. The dorsal surface was covered with tegumental folds (Figure 3A). PSO-exposed worms (100 µl/mL for 72 hours) showed destruction of the integument in the form of lacerations and peeling with appearance of multiple vesicles and blebs. Some worms were dorsally bended (Figure 3B–D). Schistosomula incubated in medium with 1% DMSO (control group) exhibited intact integuments with relatively flat surfaces and slight tegumentary ridges. There were apical constrictions between the head and the body region. At the posterior end of the body, there were prominent tail-socket wounds left by breakage of the tail (Figure 4A and B). Schistosomula swelled with multiple blisters and blebs on the surface upon exposure to PSO for 72 hours (Figure 4C and D).
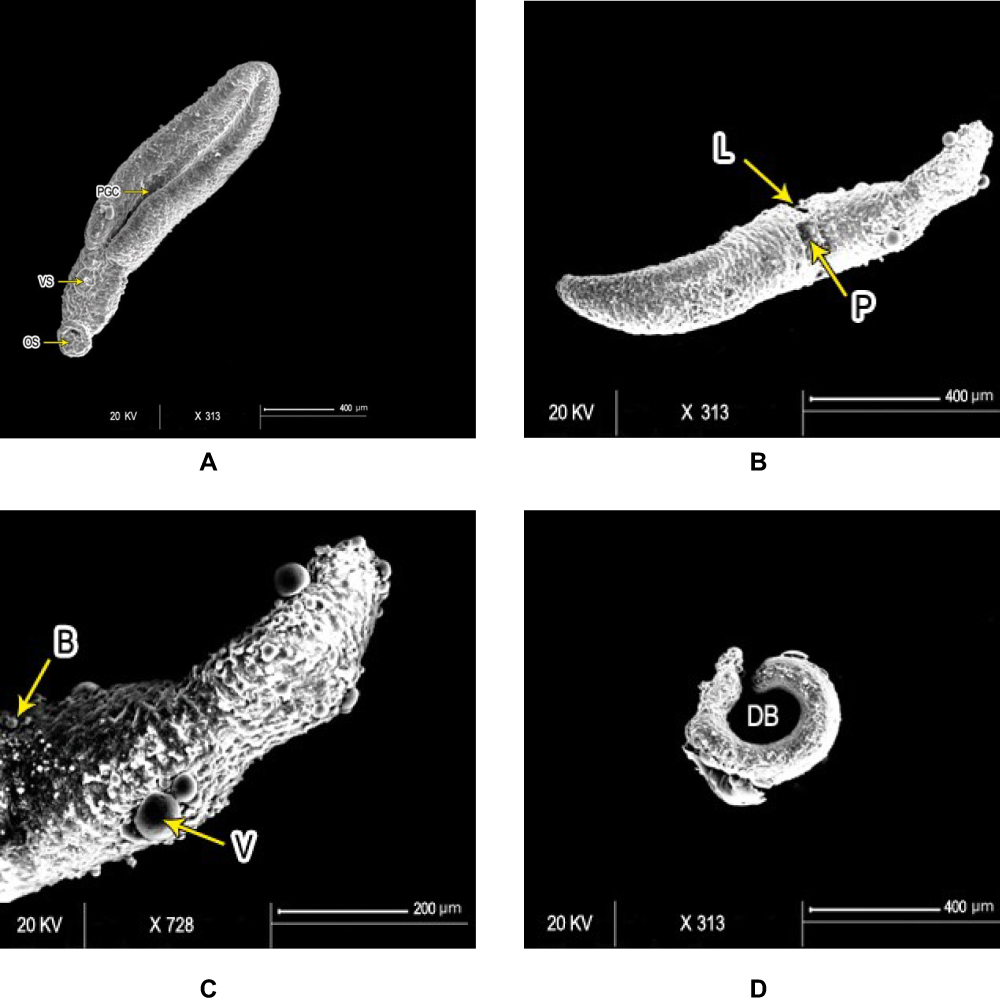 |
Figure 3 Scanning electron micrography of Schistosoma mansoni juvenile wormsafter in vitro incubation for 72 hours in medium with DMSO (negative-control group) showed (A) intact oral (OS) and ventral suckers (VS) and primitive ill-developed gynecephoric canal (PGC), whereas the dorsal surface was covered with rows of tegumental folds (TF; bar 400 µm). At 72 hours after exposure to PSO at 100 µg/mL concentration, worms exhibited (B) areas of laceration (L) and peeling (P), (C) multiple vesicles (V) and blebs (B) (bars 400 µm and 200 µm, respectively), (D) and some showed dorsal bending (DB) of the worm body (bar 400 µm). |
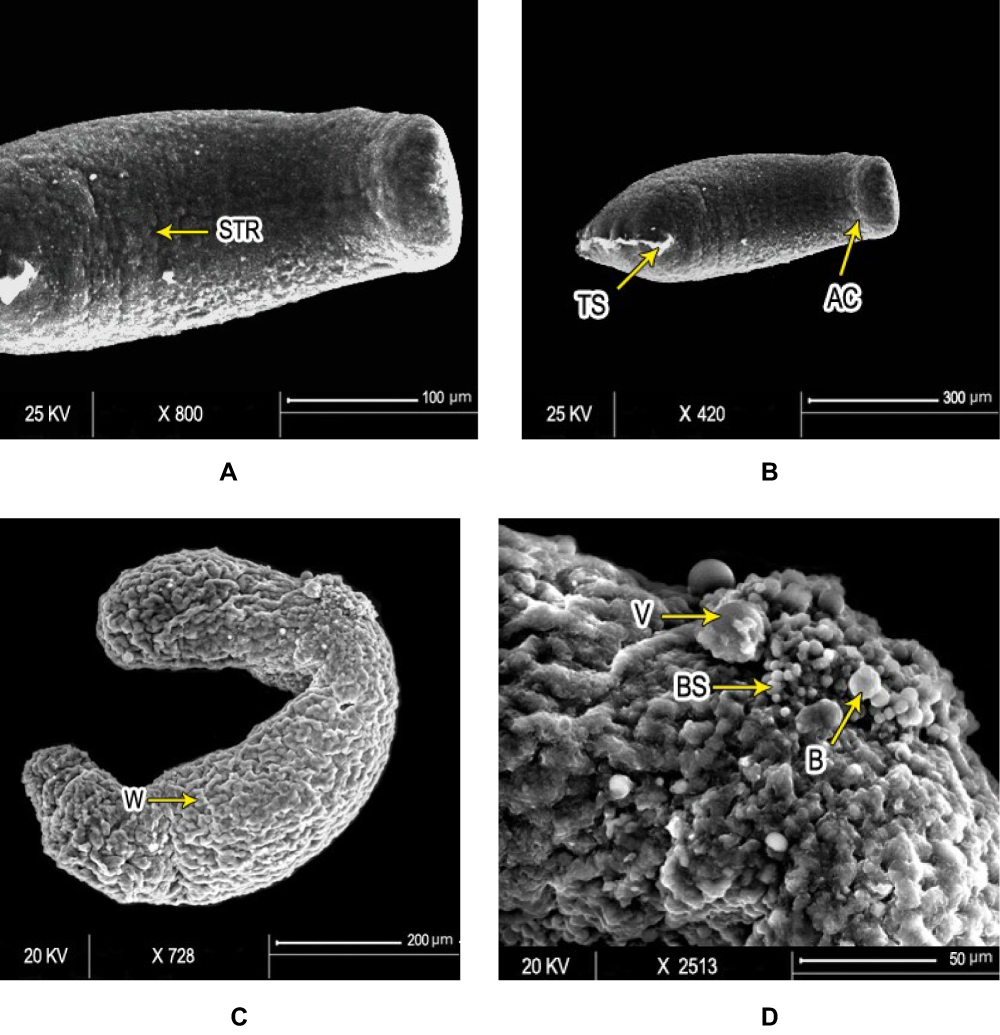 |
Figure 4 Scanning electron micrography of S. mansoni schistosomula. Ultrastructure of Sschsiistosomula incubated in medium with 1% DMSO (control group) exhibited (A) intact integument with relatively flat surface and slight tegumentary ridges (STR; bar 100 µm) and (B) an apical constriction (AC) between the head and the body region. At the posterior end of the body, there was a prominent tail-socket (TS) wound left by the breakage of the tail (bar 300 µm). After 72 hours of exposure to PSO, Sschsiistosomula showed (C) swelling of body with obvious wrinkling (W) and focal lesions (FL). There was an elongation of the worm, which may indicate flaccid paralysis (bar 200 µm). (D) Blisters (BL), blebs (B), and vesicles scattered on the surface (bar 50 µm) at high concentration (100 µL/mL) of PSO. |
Microsatellite Analysis
All tested microsatellite loci showed positive bands in the agarose gel. However, only two showed significant variations among the treatments: R95529 and SMD57. For the R95529 locus, four alleles were found of size 250–1,000 bp (Figure 5A). For SMD57, three alleles were detected in the same size range (Figure 5B). For the R95529 locus, immersion of schistosomula in 100% PSO resulted in significant elevation of abundance in all four alleles found; however, the effect was more modest than in adult worms. For the adult worms, immersion in PSO and PZQ led to complete vanishing of the small allele (allele 4). Immersion in PZQ led to three- to fivefold increases in fluorescence levels of all three alleles that did not vanish. Allele 1 showed the same fluorescence level as the control in the group treated with 20% PSO. However, both alleles 2 and 3 showed three- to fivefold increases. All these increases were significantompared the control upon treatment with 40% PSO. Upon treatment with 100% PSO, allele 1 dropped to about fivefold that of the control, while alleles 2 and 3 were only two- to threefold higher than the control (Figure 6). For SMD57, schistosomula response to PSO was less pronounced. However, a dose-dependent tendency in allele variations was found in all immersion groups. The large allele 1 vanished completely in response to PZQ and all PSO concentrations. Allele 2 was significantly higher (about 1.5-fold) than the control, but decreased with increasing PSO concentrations to below less than double its fluorescence level in the control group. Allele 3 showed slight but significant increases in the PZQ and 20% PSO groups, but a 1.5- to twofold drop in the 40% and 20% PSO groups (Figure 7).
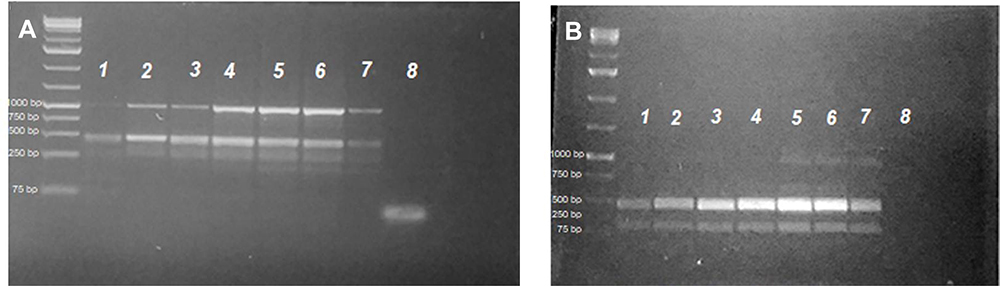 |
Figure 5 Lanes 2–7 after the 1 kb plus DNA-ladder PCR profiles for S. mansoni microsatellite loci R95529 (A) and SMD57 (B) in adult worms treated with 100 µL/mL PSO (lane 1), 40 µL/mL PSO (lane 2), 20 µL/mL PSO (lane 3), PZQ (Lane 4), schistosomula treated with 100 µL/mL PSO (lane 5), schistosomula sham control (lane 6), and adult schistosomula sham control (lane 7). A negative PCR control was loaded in lane 8 in both gels. |
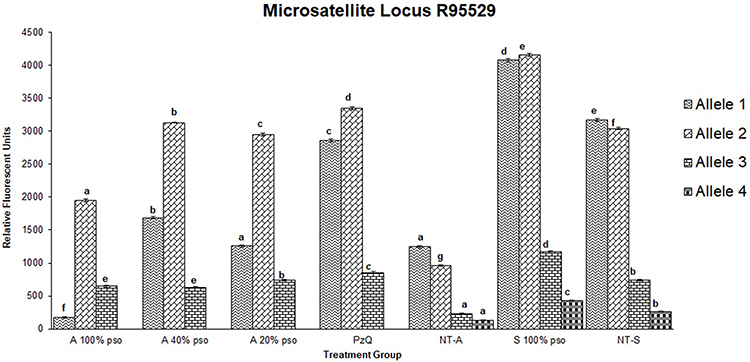 |
Figure 6 Densitometric measurements of alleles-band intensity for the microsatellite locus R95529 in adult worms treated with 100 µL/mL PSO (100% PSO), 40 µL/mL PSO (40% PSO), 20 µL/mL PSO (20% PSO), praziquantel (PZQ), untreated adult (NT-A) schistosomula treated with 100 µL/mL PSO (S100% PSO), and schistosomula untreated control (NT-S). Letters a–d refer to significant differences (p<0.05) in comparison to the control of each tested maturation (adult schistosomula) group. |
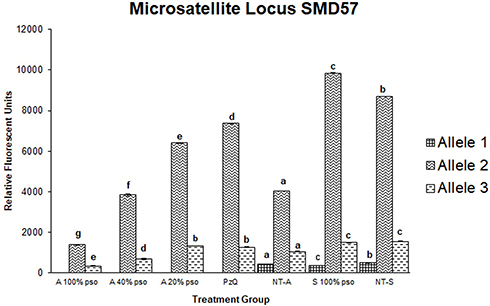 |
Figure 7 Densitometric measurements of alleles-bands intensities for the microsatellite locus SMD57 in adult worms treated with 100 µL/mL PSO (100% group), 40 µL/mL PSO (40% group), 20 µL/mL PSO (20% PSO), praziquantel (PZQ), untreated adult (NT-A), schistosomula treated with 100 µL/mL (S100% PSO), and schistosomula control (NT-S). Letters a–d refer to significant differences (p<0.05) in comparison to the control of each tested maturation (adult schistosomula) group. |
Discussion
Praziquantel (PZQ) is effective against all Schistosoma spp., but shows relatively lower efficacy against immature stages of the parasite.29 Continued use of PZQ as a sole drug can contribute to the development of drug resistance.30 These pitfalls have driven researchers to seek novel and alternative antischistosomal compounds. The last decade has seen renewed interest in the use of natural compounds as a base for drug development.31 Among the natural plants studied as possible anthelmintic agents are the family Cucurbitaceae, which are administered in traditional medicine as antiparasitic agents and have been reported to have anthelmintic properties in both humans and animals.32,33
The challenging part of new-drug discovery is to find a substance that can efficiently destroy the parasite without harming the host.34 PSO is tolerable, safe, and has been found to play a significant role in guarding against alcohol-related hepatotoxicity and oxidative stress by modulating inflammation and decreasing oxidative stress.35 Additionally, it has been found to have a defensive impact against sodium nitrate–induced oxidative damage.36 This protective role could be attributed to phenolics, tocopherols, and sterols that play an antioxidant role.37
In previous research by Beshay et al,38 PSO was tested in vivo in S. mansoni–infected mice at 50 mg/kg/day for 4 weeks, starting in the fourth week postinfection. The results showed a significant reduction in worm burden and liver and intestinal egg load, with a significant increase in the percentage of dead eggs in the oogram pattern. However, to our knowledge, no previous study on the in vitro effect of PSO on larval stages (newly formed schistosomula and juvenile worms) has been conducted. To develop our comprehension of the effect of this compound against schistosomula, we studied the in vitro impact of different concentrations of PSO on various stages of S. mansoni and estimated its LC50 and LC90. In studies on schistosomula, several parameters are often assessed as indicators of biological activity and toxicity, such as mortality rate, motor function, and tegumental changes.39 Here, besides the previous parameters, we assessed DNA changes through microsatellite analysis.
Our results showed that all tested stages of S. mansoni were susceptible to PSO. PZQ gave better results compared to PSO in adult worms, but PSO was more effective than PZQ in juvenile worms and schistosomula. Juvenile worms and schistosomula appeared to be more sensitive to the schistosomicidal activity of PSO than adult ones. It was observed that on exposure to PSO, mortality rates of adults, juveniles, and schistosomula increased with duration of exposure and occurred in a dose-dependent manner. PSO had caused 100% mortality in adults, juveniles and schistosomula at 100 µl/mL after 72 hours’ incubation, and it affected females more than males. All schistosomula died at concentrations of 100 µl/mL PSO, regardless of the duration of exposure. It seems that the stage most affected by PSO was schistosomula, followed by juveniles and lastly adults. Being more susceptible than the adults, the juveniles and schistosomula also seen after exposure to mefloquine40 and to artemisinin derivatives.41 This point needs to be researched further.
As previously mentioned by Utzinger et al42 and Xiao et al,40 schistosomula and juveniles are not susceptible to PZQ. We assessed the effect of PZQ on schistosomula and juveniles in vitro, but observed no death at concentrations as high as 10 μg/mL. In another in vitro study by de Moraes et al,34 which studied the impact of PZQ on schistosomula of different ages (3 hours and 1, 3, 5, and 7 days old), no death was observed after treatment at concentrations as high as 20 µM. It was observed that changes in S. mansoni motility at different stages varied according to PSO concentration and incubation period. Motility and morphological changes started after 48 hours’ incubation, especially at 60, 80, and 100 µl/mL concentrations. De Oliveira et al39 suggested that effects on adult S. mansoni motility exposed to treatment are associated with alterations in neurotransmitters or the parasite-neuromodulator system, since parasite motility is related to important neurotransmitters or neuromodulators like serotonin, dopamine, epinephrine, glutamate, acetylcholine, and neuropeptides.
The positive control group exposed to PZQ, at a concentration of 10 μg/mL, demonstrated significant reduction in worm motility and marked morphological changes. After 24h of exposure, adult worms showed spasmodic contractions with weak movement of the oral sucker and tail portion vesiculation along the surface of the worm. All worms acquired a coiled appearance. Conversely, no significant effect of PZQ on juveniles and schistosomula was observed. These findings were consistent with those reported by Pica-Mattoccia and Cioli43 who demonstrated that PZQ, in adult S. mansoni worms triggered both contraction and paralysis with little effect on juvenile worms. On comparison of the two drugs used in the study, PSO showed greater effects on the developmental stages of Schistosoma worms than PZQ. It would be interesting to study the effect of a combination of the two compounds on various stages of schistosomula. Beshay et al38 performed an in vivo experiment on S. mansoni, and showed that the most significant results were obtained when combining PZQ and PSO.
The integument of schistosomula appears to be an important focus in the schistosomicidal action of the therapeutic compounds. SEM is a valuable approach for determining whether a tested substance can cause damage to the integument of schistosomula. Disruption to the integument of the parasite exposes the worms to the immune system of the host and thus interferes with their survival.44 Many schistosomulum medications currently in use, such as PZQ,45 mefloquine,46 and artemether,47 cause damage to the schistosomal integument.
PSO showed evident changes in the integuments of adults, juveniles, and schistosomula in the present study. Changes in adults included destruction of the integument with loss of normal patterns and distortion of the gynecophoric canal. These changes increased in severity with increased concentrations and ended with extreme damage, edema, lacerations, and blebs formation. Furthermore, PSO-exposed juveniles and schistosomula showed lacerations and peeling with vesicles and blebs. These findings were consistent with those found by Beshay et al,38 who studied the in vivo effect of PSO on S. mansoni adults.Microsatellites (slight tegumental ridges) are hypervariable repetitive sequences consisting of two to six bp of DNA that are broadly spread throughout the genome.48 Microsatellite instability is distinguished by the extension, shortening, insertion, or deletion of microsatellites, which is the result of a defective DNA mismatch–repair system, which may result from exposure to some drugs or chemicals.49
Immersion in PSO appeared to induce a state of microsatellite instability, which is to the best of authors' knowledge the first report of such a process in this species. Moreover, after their initial identification by Durand et al,27 this is the first study to report their variation in response to a natural product. The relative abundance of these microsatellite alleles seemed to follow the same tendency in adult and schistosomulum stages, yet they were more abundant in schistosomula than adults for the same initial DNA concentration. However, low and high concentrations of PSO seemed to induce over- or underreplication of such loci in a pattern that was found to be similar to that induced by PZQ. DNA-replication errors leading to instability of microsatellite markers have been identified as a main cause of cancers and aging.50,51 Assessment of genetic instability by comparing normal and affected individuals has been validated and applied at the level of human subjects, and proved to be an accompanying factor in different pathological cases that impact genomes.50 However, in a wide spectrum of organisms, this phenomenon can be identified and directly related to mutations in genes responsible for postreplication DNA mismatch–repair genes.52 Mismatch-repair inefficiency results in either overproduction of nucleotide repeats or deletion of them.53 Such microsatellite instability has been identified as a marker for mismatch-repair loss and an associated reduction in apoptosis in response to some mutations.54 Mismatch repair is crucial in organisms at various taxonomic levels for maintenance of genomic stability and hence normal genetic and physiological performance. The antihelmintic activity of PSO may be due to specific secondary metabolites, such as cucurbitine, saponin sterols, berberine, and palmatine, present in Cucurbita spp. seeds.55 Berberine and palmatine have been identified as antischistosomal agents.56 In this regard, the microsatellite instability observed in this study as a result of different concentrations of PSO may induce genomic instability in adult worms and schistosomula. As a final remark, the resulting microsatellite instability and tegumental damage may lead to loss of the worm’s infectious ability.
Conclusion
We have shown that PSO is an efficient compound against S. mansoni schistosomula, juveniles, and adults in vitro. The mortality rate, motility effect, morphological integument alternations, and PSO-induced DNA instability suggest that this compound is a promising schistosomicidal product. Further studies are needed to clarify the mechanisms of schistosomal PSO toxicity and to assess its adequacy for the development of new antihelmintic agents.
Disclosure
The authors have no competing interests to declare.
References
1. Gryseels B, Polman K, Clerinx J, Kestens L. Human schistosomiasis. Lancet. 2006;368(9541):1106–1118. doi:10.1016/S0140-6736(06)69440-3
2. World Health Organization. “Schistosomiasis and soil-transmitted helminthiases: number of people treated in 2017”. Wkly Epidemiol Rec. 2018;93:681–692.
3. McManus DP, Dunne DW, Sacko M, Utzinger J, Vennervald BJ, Zhou X-N. Schistosomiasis (Primer). Nat Rev Dis Primers. 2018.
4. Abou-El-Naga IF, Amer EI, Boulos LM, El-Faham MH, Seada NMA, Younis SS. Biological and proteomic studies of Schistosoma mansoni with decreased sensitivity to praziquantel. Comp Immunol Microbiol Infect Dis. 2019;66:101341. doi:10.1016/j.cimid.2019.101341
5. Aragon AD, Imani RA, Blackburn VR, et al. Towards an understanding of the mechanism of action of praziquantel. Mol Biochem Parasitol. 2009;164(1):57–65. doi:10.1016/j.molbiopara.2008.11.007
6. Bergquist R, Utzinger J, Keiser J. Controlling schistosomiasis with praziquantel: how much longer without a viable alternative? Infect Dis Poverty. 2017;6(1):74. doi:10.1186/s40249-017-0286-2
7. Kocyan A, Zhang L-B, Schaefer H, Renner SS. A multi-locus chloroplast phylogeny for the Cucurbitaceae and its implications for character evolution and classification. Mol Phylogenet Evol. 2007;44(2):553–577. doi:10.1016/j.ympev.2006.12.022
8. Caili FU, Huan SHI, Quanhong LIA. Review on pharmacological activities and utilization technologies of pumpkin. Plant Foods Human Nutr. 2006;61(2):70–77. doi:10.1007/s11130-006-0016-6
9. Bardaa S, Halima NB, Aloui F, et al. Oil from pumpkin (Cucurbita pepo L.) seeds: evaluation of its functional properties on wound healing in rats. Lipids Health Dis. 2016;15(1):73. doi:10.1186/s12944-016-0237-0
10. Marie-Magdeleine C, Mahieu M, Archimède H. Pumpkin (Cucurbita moschata Duchesne ex Poir.) seeds as an anthelmintic agent? In: Preedy VR, Watson RR, Patel VB, eds. Nuts and Seeds in Health and Disease Prevention. Elsevier; 2011:933–939.
11. Li T, Ito A, Chen X, et al. Usefulness of pumpkin seeds combined with areca nut extract in community-based treatment of human taeniasis in northwest Sichuan Province, China. Acta Trop. 2012;124(2):152–157. doi:10.1016/j.actatropica.2012.08.002
12. Webster B, Rabone M, Pennance T, et al. Development of novel multiplex microsatellite polymerase chain reactions to enable high-throughput population genetic studies of Schistosoma haematobium. Parasit Vectors. 2015;8(1):432. doi:10.1186/s13071-015-1044-6
13. Olivier L, Stirewalt M. An efficient method for exposure of mice to cercariae of Schistosoma mansoni. J Parasitol. 1952;38(1):19–23. doi:10.2307/3274166
14. Duvall RH, DeWitt WB. An improved perfusion technique for recovering adult schistosomes from laboratory animals. Am J Trop Med Hyg. 1967;16(4):483–486. doi:10.4269/ajtmh.1967.16.483
15. Colley DG, Wikel SK. Schistosoma mansoni: simplified method for the production of schistosomules. Exp Parasitol. 1974;35(1):44–51. doi:10.1016/0014-4894(74)90005-8
16. Xiao S-H, Keiser J, Chollet J, et al. In vitro and in vivo activities of synthetic trioxolanes against major human schistosome species. Antimicrob Agents Chemother. 2007;51(4):1440–1445. doi:10.1128/AAC.01537-06
17. Ingram K, Schiaffo CE, Sittiwong W, Benner E, Dussault PH, Keiser J. In vitro and in vivo activity of 3-alkoxy-1, 2-dioxolanes against Schistosoma mansoni. J Antimicrob Chemother. 2012;67(8):1979–1986. doi:10.1093/jac/dks141
18. Silva-Moraes V, Couto FFB, Vasconcelos MM, et al. Antischistosomal activity of a calcium channel antagonist on schistosomula and adult Schistosoma mansoni worms. Mem Inst Oswaldo Cruz. 2013;108(5):600–604. doi:10.1590/0074-0276108052013011
19. Ramirez B, Bickle Q, Yousif F, Fakorede F, Mouries M-A, Nwaka S. Schistosomes: challenges in compound screening. Expert Opin Drug Discov. 2007;2(sup1):S53–S61. doi:10.1517/17460441.2.S1.S53
20. Holtfreter M, Loebermann M, Klammt S, et al. Schistosoma mansoni: schistosomicidal effect of mefloquine and primaquine in vitro. Exp Parasitol. 2011;127(1):270–276. doi:10.1016/j.exppara.2010.08.008
21. El Bardicy S, El Sayed I, Yousif F, et al. Schistosomicidal and molluscicidal activities of aminoalkylamino substituted neo-and norneocryptolepine derivatives. Pharm Biol. 2012;50(2):134–140. doi:10.3109/13880209.2011.578278
22. Hassan M, El-Motaiem M, Afify H, Abaza B, El-Shafei M, Massoud A. In vitro effect of Mirazid on Schistosoma mansoni worms. J Egypt Soc Parasitol. 2003;33(3):999–1008.
23. Galal-Khallaf A, Osman AG, Carleos CE, Garcia-Vazquez E, Borrell YJ. A case study for assessing fish traceability in Egyptian aquafeed formulations using pyrosequencing and metabarcoding. Fish Res. 2016;174:143–150. doi:10.1016/j.fishres.2015.09.009
24. Ardura A, Horreo J, Hernandez E, et al. Forensic DNA analysis reveals use of high trophic level marine fish in commercial aquaculture fish meals. Fish Res. 2012;115–116:115–120. doi:10.1016/j.fishres.2011.08.011
25. Wasko AP, Martins C, Oliveira C, Foresti F. Non‐destructive genetic sampling in fish. An improved method for DNA extraction from fish fins and scales. Hereditas. 2003;138(3):161–165. doi:10.1034/j.1601-5223.2003.01503.x
26. Mohammed-Geba K, El-Nab -SE-SH, Awad E, Nofal AI. DNA barcoding identifies a unique haplotype of nile tilapia Oreochromis niloticus thriving in Egyptian freshwater and brackish water lakes. Int J Ecotoxicol Ecobiol. 2017;2:172–177. doi:10.11648/j.ijee.20170204.16
27. Durand P, Sire C, Theron A. Isolation of microsatellite markers in the digenetic trematode Schistosoma mansoni from Guadeloupe island. Mol Ecol. 2000;9(7):997–998. doi:10.1046/j.1365-294x.2000.00939-4.x
28. Albus U. Guide for the Care and Use of Laboratory Animals.
29. Doenhoff MJ, Cioli D, Utzinger J. Praziquantel: mechanisms of action, resistance and new derivatives for schistosomiasis. Curr Opin Infect Dis. 2008;21(6):659–667. doi:10.1097/QCO.0b013e328318978f
30. Lotfy WM, Hishmat MG, El Nashar AS, Abu El Einin HM. Evaluation of a method for induction of praziquantel resistance in Schistosoma mansoni. Pharm Biol. 2015;53(8):1214–1219. doi:10.3109/13880209.2014.970289
31. Ji HF, Li XJ, Zhang HY. Natural products and drug discovery. EMBO Rep. 2009;10(3):194–200. doi:10.1038/embor.2009.12
32. Acorda JA, Mangubat IYEC, Divina BP. Evaluation of the in vivo efficacy of pumpkin (Cucurbita pepo) seeds against gastrointestinal helminths of chickens. Turk J Vet Anim Sci. 2019;43(2):206–211. doi:10.3906/vet-1807-39
33. Alhawiti AO, Toulah FH, Wakid MH. Anthelmintic potential of Cucurbita pepo Seeds on Hymenolepis nana. Acta Parasitologica. 2019;64(2):276–281. doi:10.2478/s11686-019-00033-z
34. de Moraes J, Nascimento C, Yamaguchi LF, Kato MJ, Nakano E. Schistosoma mansoni: in vitro schistosomicidal activity and tegumental alterations induced by piplartine on schistosomula. Exp Parasitol. 2012;132(2):222–227. doi:10.1016/j.exppara.2012.07.004
35. Seif HSA. Ameliorative effect of pumpkin oil (Cucurbita pepo L.) against alcohol-induced hepatotoxicity and oxidative stress in albino rats. BJBAS. 2014;3(3):178–185.
36. Rouag M, Berrouague S, Djaber N, et al. Pumpkin seed oil alleviates oxidative stress and liver damage induced by sodium nitrate in adult rats: biochemical and histological approach. Afr Health Sci. 2020;20(1):413–425.
37. Rezig L, Chouaibi M, Msaada K, Hamdi S. Chemical composition and profile characterisation of pumpkin (Cucurbita maxima) seed oil. Ind Crops Prod. 2012;37(1):82–87. doi:10.1016/j.indcrop.2011.12.004
38. Beshay E, Rady A, Afifi A, Mohamed A. Schistosomicidal, antifibrotic and antioxidant effects of Cucurbita pepo L. seed oil and praziquantel combined treatment for Schistosoma mansoni infection in a mouse model. J Helminthol. 2019;93(3):286–294. doi:10.1017/S0022149X18000317
39. De Oliveira R, Rehder V, Oliveira A, et al. Schistosoma mansoni: in vitro schistosomicidal activity of essential oil of Baccharis trimera (less) DC. Exp Parasitol. 2012;132:135–143. doi:10.1016/j.exppara.2012.06.005
40. Xiao S-H, Mei J-Y, Jiao P-Y. The in vitro effect of mefloquine and praziquantel against juvenile and adult Schistosoma japonicum. Parasitol Res. 2009;106(1):237–246. doi:10.1007/s00436-009-1656-x
41. Utzinger J, Chollet J, Tu Z, Shuhua X, Tanner M. Comparative study of the effects of artemether and artesunate on juvenile and adult Schistosoma mansoni in experimentally infected mice. Trans R Soc Trop Med Hyg. 2002;96(3):318–323. doi:10.1016/S0035-9203(02)90110-0
42. Utzinger J, Keiser J, Shuhua X, Tanner M, Singer BH. Combination chemotherapy of schistosomiasis in laboratory studies and clinical trials. Antimicrob Agents Chemother. 2003;47(5):1487–1495. doi:10.1128/AAC.47.5.1487-1495.2003
43. Pica-Mattoccia L, Cioli D. Sex-and stage-related sensitivity of Schistosoma mansoni to in vivo and in vitro praziquantel treatment. Int J Parasitol. 2004;34(4):527–533. doi:10.1016/j.ijpara.2003.12.003
44. Miranda Filho CAL, de Oliveira Barbosa M, Oliveira AR, et al. In vitro and in vivo activities of multi-target phtalimido-thiazoles on Schistosomiasis mansoni. Eur J Pharma Sci. 2020;146:105236. doi:10.1016/j.ejps.2020.105236
45. Mendonca AMB, Feitosa APS, Veras DL, et al. The susceptibility of recent isolates of Schistosoma mansoni to praziquantel. Rev Inst Med Trop Sao Paulo. 2016;58. doi:10.1590/S1678-9946201658007
46. Manneck T, Haggenmüller Y, Keiser J. Morphological effects and tegumental alterations induced by mefloquine on schistosomula and adult flukes of Schistosoma mansoni. Parasitology. 2010;137(1):85–98. doi:10.1017/S0031182009990965
47. Shuhua X, Binggui S, Chollet J, Utzinger J, Tanner M. Tegumental changes in adult Schistosoma mansoni harbored in mice treated with artemether. J Parasitol. 2000;86(5):1125–1132. doi:10.1645/0022-3395(2000)086[1125:TCIASM]2.0.CO;2
48. Srivastava D, Ahmad MM, Shamim M, et al. Modulation of Gene Expression by Microsatellites in Microbes. In: Singh HB, Gupta VK, Jogaiah S, eds. New and Future Developments in Microbial Biotechnology and Bioengineering. Elsevier; 2019:209–218.
49. Nojadeh JN, Sharif SB, Sakhinia E. Microsatellite instability in colorectal cancer. EXCLI J. 2018;17:159.
50. Mourah S, Cussenot O, Vimont V, et al. Assessment of microsatellite instability in urine in the detection of transitional‐cell carcinoma of the bladder. Int J Cancer. 1998;79(6):629–633. doi:10.1002/(SICI)1097-0215(19981218)79:6<629::AID-IJC13>3.0.CO;2-1
51. Lidzbarsky G, Gutman D, Shekhidem HA, Sharvit L, Atzmon G. Genomic instabilities, cellular senescence, and aging: in vitro, in vivo and aging-like human syndromes. Front Med. 2018;5:104. doi:10.3389/fmed.2018.00104
52. Degtyareva NP, Greenwell P, Hofmann ER, et al. Caenorhabditis elegans DNA mismatch repair gene msh-2 is required for microsatellite stability and maintenance of genome integrity. Proc Natl Acad Sci. 2002;99(4):2158–2163. doi:10.1073/pnas.032671599
53. Strand M, Prolla TA, Liskay RM, Petes TD. Destabilization of tracts of simple repetitive DNA in yeast by mutations affecting DNA mismatch repair. Nature. 1993;365(6443):274–276. doi:10.1038/365274a0
54. Buermeyer AB, Wilson-van Patten C, Baker SM, Liskay RM. The human MLH1 cDNA complements DNA mismatch repair defects in Mlh1-deficient mouse embryonic fibroblasts. Cancer Res. 1999;59(3):538–541.
55. Grzybek M, Kukula-Koch W, Strachecka A, et al. Evaluation of anthelmintic activity and composition of pumpkin (Cucurbita pepo L.) seed extracts—in vitro and in vivo studies. Int J Mol Sci. 2016;17(9):1456. doi:10.3390/ijms17091456
56. Dkhil MA, Moneim AEA, Al-Quraishy S. Berberine protects against Schistosoma mansoni-induced oxidative damage in renal and testicular tissues of mice. Pak J Zool. 2014;46(3).
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.