Back to Journals » Infection and Drug Resistance » Volume 13
Cryptococcus neoformans/gattii Species Complexes from Pre-HIV Pandemic Era Contain Unusually High Rate of Non-Wild-Type Isolates for Amphotericin B
Authors Pharkjaksu S ![]() , Chongtrakool P
, Chongtrakool P ![]() , Chayakulkeeree M
, Chayakulkeeree M ![]() , Mitrpant C, Angkasekwinai P, Bennett JE, Kwon-Chung KJ, Ngamskulrungroj P
, Mitrpant C, Angkasekwinai P, Bennett JE, Kwon-Chung KJ, Ngamskulrungroj P ![]()
Received 30 October 2019
Accepted for publication 11 February 2020
Published 26 February 2020 Volume 2020:13 Pages 673—681
DOI https://doi.org/10.2147/IDR.S235473
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Sahil Khanna
Sujiraphong Pharkjaksu, 1 Piriyaporn Chongtrakool, 1 Methee Chayakulkeeree, 2 Chalermchai Mitrpant, 3 Pornpimon Angkasekwinai, 4 John E Bennett, 5 Kyung J Kwon-Chung, 5 Popchai Ngamskulrungroj 1
1Department of Microbiology, Faculty of Medicine Siriraj Hospital, Mahidol University, Bangkok, Thailand; 2Division of Infectious Diseases and Tropical Medicine, Department of Medicine, Faculty of Medicine Siriraj Hospital, Mahidol University, Bangkok, Thailand; 3Department of Biochemistry, Faculty of Medicine Siriraj Hospital, Mahidol University, Bangkok, Thailand; 4Department of Medical Technology, Faculty of Allied Health Sciences, Thammasat University, Pathumthani, Thailand; 5Laboratory of Clinical Immunology and Microbiology, National Institute of Allergy and Infectious Diseases, National Institutes of Health, Bethesda, MD, USA
Correspondence: Popchai Ngamskulrungroj
Department of Microbiology, Faculty of Medicine Siriraj Hospital, Mahidol University, 2 Wanglang Road, Siriraj, Bangkok Noi, Bangkok 10700, Thailand
Tel +66 2 419 7053
Fax +66 2 418 4148
Email [email protected]
Introduction: The Cryptococcus neoformans/gattii species complexes are a leading cause of fatality among HIV-infected patients. Despite the unavailability of clinical breakpoints (CBPs) for antifungal agents, epidemiological cutoff values (ECVs) were recently proposed, and non-wild-type isolates for polyenes and azoles are being increasingly reported. However, the distributions of the susceptibility patterns for pre-HIV-era isolates have not been studied.
Methods: We determined the in vitro antifungal susceptibility patterns of 233 Cryptococcus isolates, collected at the National Institutes of Health, USA, in pre-HIV pandemic era, to study minimum inhibitory concentrations (MICs) to the important drugs for cryptococcosis and to compare the results with strain genotypes. Amphotericin B susceptibility was compared to published ECV of C. neoformans.
Results: The 233 Cryptococcus strains consisted of 89.7% C. neoformans species complex and 10.3% C. gattii species complex. Most were from clinical sources (189, 81.1%), and the major molecular type was VNI (146, 62.7%). The highest geometric mean (GM) was observed for fluconazole (GM = 0.96 μg/mL) while the lowest was for itraconazole (GM = 0.10 μg/mL). MICs to fluconazole in C. gattii species complex were significantly higher than C. neoformans species complex (p < 0.001). Moreover, C. neoformans/VNI strains showed significantly higher MICs than others such as C. neoformans/VNII to fluconazole (p < 0.0001) and C. deneoformans/VNIV to amphotericin B (p = 0.022) and fluconazole (p = 0.008). In our collection of 167 clinical C. neoformans species complex strains, 85 (50.9%), 24 (14.4%), and 3 (1.8%) strains had an amphotericin B (AMB)-MIC of 1, 2, and 4 μg/mL, respectively. The high percentage (66.9%, 79/118 strains) of non-wild-type clinical C. neoformans VNI strains, using an AMB-ECV of 0.5 μg/mL, was found. Moreover, 25 of 28 (89.3%) C. neoformans VNI strains from environmental and veterinary sources also had AMB-MICs above 0.5 μg/mL. In general, there was no significant difference in GM AMB-MIC of the clinical strains isolated from patients with (35 patients) and without (78 patients) prior AMB treatment (0.85 vs 0.76; p = 0.624). GM MIC of the environmental strains was not significantly different from that of the prior AMB-treatment strains (0.98 vs 0.76, p = 0.159) and the post-AMB-treatment strains (0.98 vs 0.85, p = 0.488).
Conclusion: The high rate of non-wild-type among these otherwise naive isolates to amphotericin B is unexpected. Confirmation with more strains from a later era is needed.
Keywords: Cryptococcus neoformans/gattii species complexes, pre-HIV pandemic, genotype, antifungal susceptibility, epidemiologic cutoff values, non-wild type
Introduction
The advent of the AIDS pandemic in the 1980s caused a dramatic increase in cryptococcosis incidence. Nowadays, the global burden of cryptococcosis is estimated to be more than 900,000 cases per year with HIV-associated cryptococcal meningoencephalitis being highest in sub-Saharan Africa and Southeast Asia.1 An estimated 15% of AIDS patients worldwide die from cryptococcosis, and the major cause of fatality is due to delayed diagnosis, poor patient management, or emergence of antifungal drug resistance.1–3
Untreated cryptococcal meningitis is 100% fatal.4 The introduction of amphotericin B (AMB) in the 1950s decreased the mortality rate of cryptococcal meningoencephalitis to less than 50%.5 Currently, the standard therapy for cryptococcosis, was recommended by the World Health Organization and the Infectious Diseases Society of America, induction therapy. This consists of AMB in combination with 5-fluorocytosine (5FC), followed by triazoles such as fluconazole (FLC) as a consolidation therapy.2,6 Failures of cryptococcosis therapy by the standard antifungals are not uncommon.7,8 One study reported antibiotic persistence and microevolution as a possible resistance mechanism leading to therapy failure.9 According to one report, a reduction of treatment efficacy by FLC in the setting of meningitis in AIDS patients emerged after a prolonged treatment or prophylaxis with FLC.7 Although there is no clear relationship between fluconazole MIC and results of treatment with fluconazole alone, there are reports from some countries such as Cambodia,10 India,11 Taiwan,12 and Singapore13 which showed approximately 15% of treatment failure of FLC with MIC of the cryptococcal isolates up to ≥256 µg/mL. Such high MICs for FLC have occasionally been observed among both clinical and environmental C. neoformans species complex isolates. Small differences in MIC of antifungals were reported among different genotypes of C. gattii species complex, although there is no clear relationship between susceptibility and treatment outcome. One study found that the VGII (C. deuterogatti) had a higher geometric mean (GM) of MIC to FLC than VGI (C. gattii).14 Another study demonstrated higher GM MIC for azole drugs among the isolates of VGIV (C. tetragattii) as compared to VGI and VGIII (C. bacillisporus).15 There was no significant difference in MICs from C. neoformans, indicating that C. gattii can be effectively treated with the same antifungal regimens as C. neoformans.16,17
To predict the efficacy of antimicrobial treatment, the susceptibility of the infecting microorganism to an antimicrobial agent is one factor to be considered. Ideally, a clinical breakpoint (CBP) is used as a predictive MIC value to determine whether an infection is likely to respond to an antimicrobial agent. However, the CBP is usually not chosen for some less common pathogenic organisms. Therefore, an alternative value, an epidemiological cutoff value (ECVs), while not predictive of in vivo efficacy, has been offered by experts to determine whether a strain is wild type (in vitro susceptible) or non-wild type (in vitro resistant).18,19 This number is selected by a committee, such as the Clinical and Laboratory Standards Institute (CLSI), based on pharmacologic consideration and not clinical outcome. For example, the availability of reference methodologies has enabled the recognition of resistant isolates as well as proposed CBPs and ECVs for Candida spp. and Aspergillus spp. with regard to the most available antifungal agents. The ECVs of Cryptococcus species became available in the 2010s to interpret as a wild type or non-wild type based on the molecular types of the isolates.18,20
Since many antifungal drugs, such as fluconazole (a widely used azole drug currently used for therapy for cryptococcosis), were not available during the pre-HIV pandemic era, the MICs of these naïve isolates were never investigated. Therefore, we conducted this study to establish the antifungal susceptibility patterns of the C. neoformans and C. gattii species complexes recovered during the pre-HIV pandemic era and to investigate whether the antifungal susceptibility patterns varied between different genotypes or the source of isolation. Furthermore, changes in the antifungal susceptibility patterns in sequential clinical isolates from the same patients were also determined.
Materials and Methods
Study Isolates
Cryptococcal isolates and their associated demographic data which had been collected before 1980 (pre-HIV pandemic) at the Laboratory of Clinical Immunology and Microbiology of National Institute of Allergy and Infectious Diseases were obtained and maintained in a culture collection of Department of Microbiology, Siriraj Hospital, Thailand. Each isolate was cultured on Sabouraud dextrose agar (SDA; 4% dextrose, 1% peptone, 1.5% agar, with a final pH of 5.6 ± 0.2; Oxoid, UK) and incubated at 30°C for 48–72 h. All revived isolates were prepared for glycerol stock (25%) and maintained at −80°C. Information on each cryptococcal isolate was retrieved from previous reports21,22 (Supplementary Table 1). Each isolate including the sequential isolates from the same patient was considered as an individual strain as the different MIC data have been shown previously among longitudinal isolates from the same patients.9 This study was approved by the Siriraj Institutional Ethics Review Board (COA no. Si 091/2016). As there was no direct contact with the patients involved, the requirement for informed consents was waived by the institutional ethic committee.
Analysis of Genotype
DNA was extracted using the phenol–chloroform–isoamyl alcohol (25:24:1, v: v: v) method.19 The URA5 gene was amplified with the following primers, URA5 (5ʹATGTCCTCCCAAGCCCTCGACTCCG3ʹ) and SJ01(5ʹTTAAGACC TCTGAACACCGTACTC3ʹ). Genotypes were determined with a restriction fragment length polymorphism analysis (RFLP) of the URA5 gene digested with restriction enzymes HhaI and Sau96I (Thermo Fisher Scientific, MA, USA).23 A set of standard laboratory reference strains representing each of the eight major molecular types were used for the molecular typing: WM148 (C. neoformans/VNI), WM626 (C. neoformans/VNII), WM 628 (C. neoformans × deneoformans hybrid/VNIII), WM 629 (C. deneoformans/VNIV), WM 179 (C. gattii/VGI), WM 178 (C. deuterogattii/VGII), WM 175 (C. bacillisporus/VGIII), and WM 779 (C. tetragattii/VGIV).23
Antifungal Susceptibility Testing (AFST)
The drug susceptibilities of each isolate were determined by the broth microdilution method.18,20 Initially, plates containing serial two-fold dilutions of the antifungal agents were prepared as follows: fluconazole (FLC): 0.06–64 µg/mL; itraconazole (ITC): 0.03–16 µg/mL; amphotericin B (AMB): 0.03–16 µg/mL; and 5-fluorocytosine (5FC): 0.06–64 µg/mL. The antifungal agents were diluted in an antifungal susceptibility testing (AFST) medium (RPMI-1640 with 0.165 M morpholine propanesulfonic acid [MOPS]) to 2X concentration.24 One hundred microliters of each agent in 2X concentration were added onto a 96-well plate and stored at −70°C pending their use.
On the day of the test, 100 µL of yeast suspension (McFarland standard No. 0.5; approximately 107 cell/mL) was suspended into 9.9 mL of AFST medium, to yield a final concentration of approximately 5×102 to 2.5×103 cells/mL of working yeast suspension. The AFST panels were resuspended with 100 μL of working yeast suspension in each well with a multichannel pipetting device. The AFST plate was covered with adhesive seals and incubated at 35 ºC for 48 h in a non-CO2 incubator. If the growth control showed negative, the plates were incubated for another 24 hrs before being re-examined.
The amount of growth in each well was compared with that of the growth control. For amphotericin B, the MIC was determined as the lowest drug concentration that prevented any visible growth. As to the 5FC and azole antifungals, the MIC was determined as the lowest drug concentration yielded a 50% inhibition of growth relative to the growth control. Quality control of the AFST plate was performed with Issatchenkia orientalis (Candida krusei) ATCC 6258 and Candida parapsilosis ATCC 22019 before the testing of each lot.20 Furthermore, standard strains of Cryptococcus neoformans/gattii species complexes, H99 and R265, were included as control strains. Repeats of both the MIC and E-test methods were also done for any controversial results. The criteria used to interpret the ECVs were based on the guidelines of the CLSI guidelines but they only were available for some molecular types: AMB in C. neoformans/VNI = 0.5, C. gattii/VGI = 0.5, C. deuterogattii/VGII = 1; 5FC in C. neoformans/VNI = 8, C. gattii/VGI = 4, C. deuterogattii/VGII = 32; ITC in C. neoformans/VNI = 0.25, C. gattii/VGI = 0.5, C. deuterogattii/VGII = 1; FLC in C. neoformans/VNI = 8, C. gattii/VGI = 16, C. deuterogattii/VGII = 32.18 ECVs for the other molecular types are currently not available. To compare the MIC to the AMB among the sequential isolates, the first and the last strains that were isolated from each patient were used as representative strains for the pre- and post-treatment groups, respectively. The MIC90 and MIC50 values were defined as the lowest concentration of the antifungals at which 90% and 50% of the strains were inhibited, respectively.
Statistical Analysis
Data were calculated as MIC range, geometric mean MIC (GM MIC), MIC50, and MIC90 in Microsoft Excel version 2019. For geometric means, isolates with MIC’s designated as “equal to or greater than” were given the value as equal to the number. A two-tailed Fisher’s exact test and a two-tailed unpaired t test implemented in GraphPad QuickCals (https://www.graphpad.com/quickcalcs/) were applied to determine the correlation analysis, including the environmental or clinical sample sources. The Mann–Whitney U-test or Kruskal–Wallis test with Dunn’s multiple comparisons test was implemented in GraphPad Prism version 8.0.2 (GraphPad Software, California, USA) to examine the in vitro antifungal susceptibility testing results. A value of p < 0.05 was considered significant.
Results
VNI Was the Most Predominant Genotype Among the Isolates of C. neoformans/gattii Species Complexes Recovered in Pre-HIV Era
As ECVs are only available for some molecular types of the C. neoformans/gattii species complexes, molecular typing via a URA5-RFLP was conducted. Among the 233 isolates of globally collected C. neoformans/gattii species complexes (Supplementary Table 1), C. neoformans/VNI was the most prevalent genotype (146 strains, 62.7%) followed by C. neoformans/VNII (34 strains, 14.6%), C. deneoformans/VNIV (24 strains, 10.3%), C. bacillisporus/VGIII (17 strains, 7.3%), C. gattii/VGI (6 strains, 2.6%), C. neoformans × deneoformans hybrid/VNIII (5 strains, 2.1%), and C. deuterogattii/VGII (1 strain, 0.4%). Most isolates (n = 189 strains, 81.1%) were clinical strains isolated from 154 patients. 37 (15.9%) and 7 (3.0%) were environmental and veterinary strains, respectively. Based on the geographic distributions, 199 strains (85.4%) had been recovered from the USA followed by Thailand (14 strains, 6.0%), Denmark (10 strains, 4.3%), Italy (9 strains, 3.9%), and Canada (1 strain, 0.4%). Fifty-three clinical isolates were sequential strains isolated from 18 patients (Supplementary Table 2).
C. gattii Species Complex Had Significantly Higher Geometric Mean Than C. neoformans Species Complex to FLC
MIC ranges, GM MIC, MIC50, and MIC90 of the 4 antifungal agents are presented in Supplementary Table 3. The MIC ranges for each of the antifungal drugs were ≤0.06–4 µg/mL for AMB, 0.12 - ≥128 µg/mL for 5FC, ≤0.06–8 µg/mL for ITC, and 0.12–8 µg/mL for FLC. Overall, the highest GM MIC was observed with FLC (GM = 0.96 µg/mL) while the lowest GM MIC was observed with ITC (GM = 0.10 µg/mL). The geometric mean of the C. gattii species complex to FLC was significantly higher than that of the C. neoformans species complex (1.68 vs 0.90 µg/mL; p < 0.001). No statistically significant difference was observed for the other drugs (Figure 1).
 |
Figure 1 Geometric mean (GM) MIC of four antifungal agents between C. neoformans species complex (black bar) and C. gattii species complex (white bar). Only the p-value of the statistically significant difference is shown. Abbreviations: AMB, amphotericin B; 5FC, 5-fluorocytosine; ITC, itraconazole; FLC, fluconazole. |
Among C. neoformans Species Complex, VNI Isolates Generally Had Significantly Higher Geometric Means to AMB and FLC Than Other Genotypes
Comparisons of the MICs between molecular types revealed that the molecular type C. neoformans/VNI had a higher GM MIC than C. neoformans/VNII to FLC (1.06 vs 0.49 µg/mL; p < 0.0001). However, 6 of the 8 strains with high resistance to 5FC (≥128 µg/mL) following 5FC monotherapy were VNI and only one was VNII. Strains of VNI also had a higher GM MIC than C. deneoformans/VNIV to AMB and FLC (0.88 vs 0.49 µg/mL; p = 0.022 and 1.06 vs 0.77 µg/mL; p = 0.008, respectively). However, the molecular type C. neoformans/VNI had slightly lower GM MIC than C. deneoformans/VNIV to 5FC (0.13 vs 0.17 µg/mL; p = 0.017; Figure 2).
 |
Figure 2 Geometric mean (GM) MIC of four antifungal agents among different genotypes in C. neoformans species complex. Only the p-value of the statistically significant difference is shown. Abbreviations: AMB, amphotericin B; 5FC, 5-fluorocytosine; ITC, itraconazole; FLC, fluconazole. |
Unusually High Rate of Non-Wild Type Strains to AMB Was Observed Among Pre-HIV Era Cryptococcal Isolates
According to the ECVs, a high rate of non-wild type strain to AMB was observed for both the C. neoformans/VNI strains (104 strains, 71.2%) and C. gattii/VGI and VGII strains (3 strains, 42.9%). As to the other agents, most if not all, of the isolates were wild type (Table 1). There was no significant difference in the number of wild type isolates to any drug between the C. neoformans/VNI strains and the C. gattii/VGI and VGII strains (AMB: p = 0.198, 5FC: p > 0.999, ITC: p > 0.999, and FLC: p > 0.999).
 |
Table 1 Number of Wild Type Strains Among C. neoformans/gattii Species Complexes Based on Epidemiological Cutoff Values (ECVs) |
The Environmental Strains Showed a Higher Rate of Non-Wild Type Strains Than the Clinical Strains
Due to the unusually high proportion of non-wild type strains to AMB, we investigated if this occurred only in the clinical isolates, the population in which acquired resistance is more likely to occur. Surprisingly, we found that the percentage of non-wild type was significantly higher for the environmental isolates (96.0%) than the clinical isolates (66.9%) of C. neoformans/VNI (p = 0.003, Table 2). The comparison, however, could not be performed with the C. gattii/VGI isolates as only one veterinary and no environmental isolates were available. Considering together all the genotypes of C. neoformans, 34 (81%) of 42 isolates from environmental or veterinary sources had an amphotericin B above 0.5 μg/mL, compared to 112 (67%) of 167 clinical isolates.
 |
Table 2 Association Analysis of Amphotericin B Susceptibility Patterns Based on ECVs Among C. neoformans/VNI Between Clinical and Environmental Isolates |
No Significant Difference Between Clinical Strains with and Without Prior AMB Treatment
In general, there was no significant difference in GM AMB-MIC of the clinical strains isolated from patients with (35 patients) and without (78 patients) prior AMB treatment (0.85 vs 0.76; p = 0.624) (Supplementary Table 1). Similarly, no significant difference in non-wild type frequency of the VNI clinical strains isolated from patients with and without prior AMB treatment (60.0% vs 68.2%; p = 0.498) (Table 3). In fact, strains from only 1 out of 7 patients (patient M) exhibited 2 or more dilutions difference in AMB-MIC between the pre- and post-AMB therapy and strains with very high 5FC-MIC (≥128 µg/mL) was found in 3 patients (patient A, N, and O) with failed 5FC therapy (Supplementary Table 2). Moreover, GM MIC of the environmental strains were not significantly different from that of the prior AMB-treatment strains (0.98 vs 0.76, p = 0.159) and the post AMB-treatment strains (0.98 vs 0.85, p = 0.488) (Table 4).
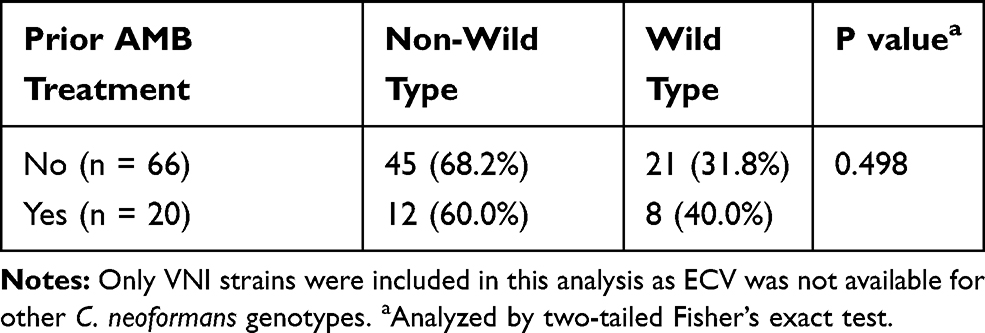 |
Table 3 Comparative Amphotericin B Susceptibility Patterns of Clinical VNI Strains with and Without Prior Amphotericin Therapy |
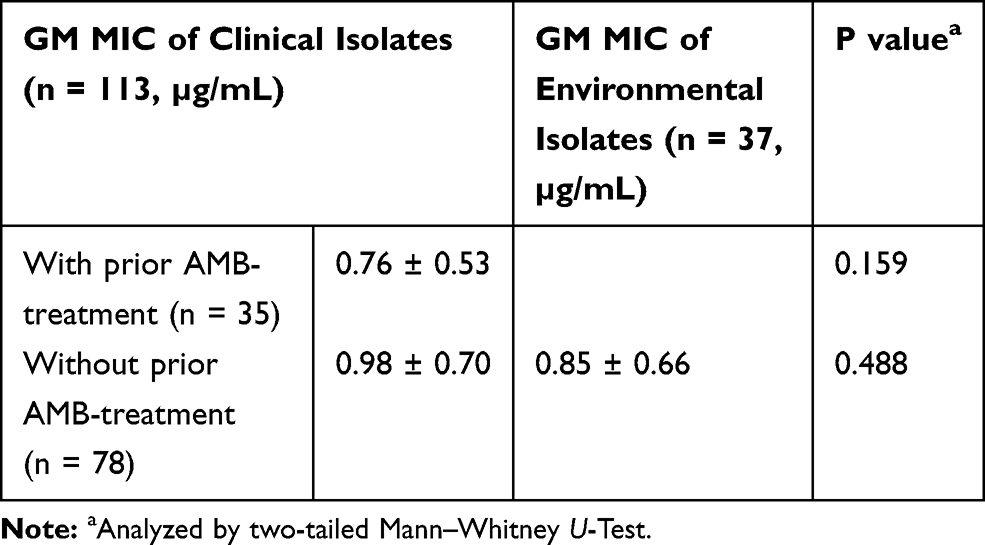 |
Table 4 Comparative Amphotericin B Susceptibility Patterns of Clinical Strains Between with or Without Prior AMB-Treatment with Environmental Strains Among C. neoformans Species Complex |
Discussion
Generally, determination of the MIC in the laboratory has been the method of choice for monitoring antifungal resistance.20 However, the antifungal resistance of Cryptococcus is difficult to define in the laboratory due to the absence of interpretive breakpoints.25 Based on the recent standardized protocol and ECVs, cryptococcal isolates from the pre-HIV pandemic were studied for antifungal susceptibility patterns and their association with epidemiological characteristics.
As both FLC and ITC became available in the 1980s, we expected that acquired resistance to both drugs was unlikely.26 Although the highest GM MIC value (0.96 µg/mL) was observed with FLC, the values were approximately a half to a quarter of the values for the isolates recovered from the present HIV pandemic era.27–30 In fact, all isolates were wild type to both FLC and ITC as expected. In the case of 5FC, almost all isolates from the pre-HIV pandemic were also wild type. Although infrequent, antifungal therapy failure for cryptococcosis has been reported during HIV pandemic era.31–33 In addition, our C. gattii isolates had higher GM MIC to almost all antifungal agents compared to C. neoformans which agrees with previous studies.34,35
One would expect low rate of acquired resistance to AMB among isolates of pre-HIV era comparing to the rate of the present day. Surprisingly, the percentage of non-wild type clinical C. neoformans VNI strains to AMB (MIC > 0.5 µg/mL) was 66.9% (79/118 strains) as very high and the rate was even higher 89.3% (25/28 strains) among the environmental and veterinary isolates. Our results differ from the reports that the majority of HIV pandemic isolates were susceptible to AMB.27,36,37 For example, based on the same AFST method and ECV, a recent study in Germany showed no non-wild type isolate to AMB among 102 C. neoformans/VNI strains isolated in the present days.38 Moreover, the most comprehensive cryptococcal AFST study with strains collected from six laboratories during 2010–2012,39 VNI, VGI, and VGII strains showed only 2.8% (28/1002 strains), 0.8% (2/259 strains), and 0.2% (1/470 strains) non-wild type rate. The difference in antifungal susceptibility pattern between pre-HIV and HIV pandemic might be explained by the fact that ECV was developed mainly from strains collected from HIV pandemic. However, this remains to be further tested with more strains from the later era.
Unlike CBPs, ECVs cannot be used to distinguish between AMB-susceptible and AMB-resistant isolates of the C. neoformans/gattii species complexes due to its lack of clinical correlation coupled with its limitation to only some molecular types. The high non-wild type rate to AMB of the pre-HIV isolates could also be due to the fact that ECV was developed mainly based on the strains collected from the HIV patients.18,40 Moreover, based on genotyping analysis, C. neoformans/VNI isolates had a significantly higher GM MIC to FLC than C. neoformans/VNII and C. deneoformans/VNIV, and a significantly higher GM MIC to AMB than C. deneoformans/VNIV. However, the difference of GM MIC to 5FC and ITC among each molecular type was either very minimal or not observed. This is similar to one study reported that no significant differences were detected among the genotypes AFLP1/VNI, AFLP1A/VNII and AFLP1B/VNII and their respective antifungal susceptibility profiles.41 These results highlight that the interpretation of cryptococcal antifungal susceptibility among different molecular type/species should be done with caution.39,42-44
Conclusion
The impact of time on antifungal susceptibility was presented. However, differences in antifungal susceptibility between centers are well known, despite using the same standard methods. Further amphotericin B susceptibility testing by multi-center with more strains of C. gattii and strains from a later era is needed.
Acknowledgments
The study was supported by the Thailand Research Fund through the Royal Golden Jubilee Ph.D. Program (grant number PHD/0056/2561, to PN and SP). PN was also supported by Siriraj Research Development Fund, Faculty of Medicine Siriraj Hospital, Mahidol University (grant number (IO) R016133012). SP was supported by the Siriraj Graduate Scholarship. The reference strains were obtained through the courtesy of Dr. Wieland Meyer, The University of Sydney, Westmead Hospital, Westmead, NSW, Australia.
Author Contributions
All authors contributed to the data analysis and the writing of this paper, gave final approval of the version to be published, and agreed to be accountable for all aspects of the work.
Disclosure
The authors declare that they have no conflicts of interest.
References
1. Rajasingham R, Rolfes MA, Birkenkamp KE, Meya DB, Boulware DR. Cryptococcal meningitis treatment strategies in resource-limited settings: a cost-effectiveness analysis. PLoS Med. 2012;9:e1001316. doi:10.1371/journal.pmed.1001316
2. WHO. Guidelines for the Diagnosis, Prevention and Management of Cryptococcal Disease in HIV-Infected Adults, Adolescents and Children: Supplement to the 2016 Consolidated Guidelines on the Use of Antiretroviral Drugs for Treating and Preventing HIV Infection. Geneva, Switzerland: World Health Organization; 2018.
3. Brown GD, Denning DW, Gow NA, Levitz SM, Netea MG, White TC. Hidden killers: human fungal infections. Sci Transl Med. 2012;4:165rv113. doi:10.1126/scitranslmed.3004404
4. Mwaba P, Mwansa J, Chintu C, et al. Clinical presentation, natural history, and cumulative death rates of 230 adults with primary cryptococcal meningitis in Zambian AIDS patients treated under local conditions. Postgrad Med J. 2001;77:769–773. doi:10.1136/pmj.77.914.769
5. Perfect JR, Cox GM. Drug resistance in Cryptococcus neoformans. Drug Resist Updat. 1999;2:259–269. doi:10.1054/drup.1999.0090
6. Perfect JR, Dismukes WE, Dromer F, et al. Clinical practice guidelines for the management of cryptococcal disease: 2010 update by the infectious diseases society of america. Clin Infect Dis. 2010;50:291–322. doi:10.1086/649858
7. Bongomin F, Oladele RO, Gago S, Moore CB, Richardson MD. A systematic review of fluconazole resistance in clinical isolates of Cryptococcus species. Mycoses. 2018;61:290–297. doi:10.1111/myc.12747
8. Pappas PG. Antifungal trials: progress, approaches, new targets, and perspectives in cryptococcosis. In: Heitman J, Kozel T, Kwon-Chung K, Perfect J, editors. Cryptococcus from Human Pathogen to Model Yeast. Washington (DC): American Society of Microbiology; 2011:527–536.
9. Mlinaric-Missoni E, Hagen F, Chew WH, Vazic-Babic V, Boekhout T, Begovac J. In vitro antifungal susceptibilities and molecular typing of sequentially isolated clinical Cryptococcus neoformans strains from Croatia. J Med Microbiol. 2011;60:1487–1495. doi:10.1099/jmm.0.031344-0
10. Chandenier J, Adou-Bryn KD, Douchet C, et al. In vitro activity of amphotericin B, fluconazole and voriconazole against 162 Cryptococcus neoformans isolates from Africa and Cambodia. Eur J Clin Microbiol Infect Dis. 2004;23:506–508. doi:10.1007/s10096-004-1136-2
11. Datta K, Jain N, Sethi S, Rattan A, Casadevall A, Banerjee U. Fluconazole and itraconazole susceptibility of clinical isolates of Cryptococcus neoformans at a tertiary care centre in India: a need for care. J Antimicrob Chemother. 2003;52:683–686. doi:10.1093/jac/dkg399
12. Chen YC, Chang TY, Liu JW, et al. Increasing trend of fluconazole-non-susceptible Cryptococcus neoformans in patients with invasive cryptococcosis: a 12-year longitudinal study. BMC Infect Dis. 2015;15:277. doi:10.1186/s12879-015-1023-8
13. Soogarun S, Wiwanitkit V, Palasuwan A, et al. Detection of Cryptococcus neoformans in bird excreta. Southeast Asian J Trop Med Public Health. 2006;37:768–770.
14. Hagen F, Khayhan K, Theelen B, et al. Recognition of seven species in the Cryptococcus gattii/Cryptococcus neoformans species complex. Fungal Genet Biol. 2015;78:16–48. doi:10.1016/j.fgb.2015.02.009
15. Lockhart SR, Iqbal N, Bolden CB, et al. Epidemiologic cutoff values for triazole drugs in Cryptococcus gattii: correlation of molecular type and in vitro susceptibility. Diagn Microbiol Infect Dis. 2012;73:144–148. doi:10.1016/j.diagmicrobio.2012.02.018
16. Tay ST, Tanty Haryanty T, Ng KP, Rohani MY, Hamimah H. In vitro susceptibilities of Malaysian clinical isolates of Cryptococcus neoformans var. grubii and Cryptococcus gattii to five antifungal drugs. Mycoses. 2006;49:324–330. doi:10.1111/j.1439-0507.2006.01242.x
17. Andrade-Silva L, Ferreira-Paim K, Mora DJ, et al. Susceptibility profile of clinical and environmental isolates of Cryptococcus neoformans and Cryptococcus gattii in Uberaba, Minas Gerais, Brazil. Med Mycol. 2013;51:635–640. doi:10.3109/13693786.2012.761737
18. CLSI. Epidemiological Cutoff Values for Antifungal Susceptibility Testing, CLSI Suppl M59 Document.
19. Danesi P, Firacative C, Cogliati M, Otranto D, Capelli G, Meyer W. Multilocus sequence typing (MLST) and M13 PCR fingerprinting revealed heterogeneity amongst Cryptococcus species obtained from Italian veterinary isolates. FEMS Yeast Res. 2014;14:897–909. doi:10.1111/1567-1364.12178
20. CLSI. Reference Method for Broth Dilution Antifungal Susceptibility Testing of Yeasts, 4th Suppl., M27-S4. Wayne (PA): Clinical Laboratory Standards Institute; 2012.
21. Bennett JE. Susceptibility of Cryptococcus neoformans to amphotericin B. Antimicrob Agents Chemother (Bethesda). 1966;6:405–410.
22. Meyer W, Aanensen DM, Boekhout T, et al. Consensus multi-locus sequence typing scheme for Cryptococcus neoformans and Cryptococcus gattii. Med Mycol. 2009;47:561–570. doi:10.1080/13693780902953886
23. Meyer W, Castaneda A, Jackson S, Huynh M, Castaneda E, IberoAmerican Cryptococcal Study G. Molecular typing of IberoAmerican Cryptococcus neoformans isolates. Emerg Infect Dis. 2003;9:189–195. doi:10.3201/eid0902.020246
24. Rex JH, Pfaller MA, Walsh TJ, et al. Antifungal susceptibility testing: practical aspects and current challenges. Clin Microbiol Rev. 2001;14:643–658. doi:10.1128/cmr.14.4.643-658.2001
25. Aller AI, Martin-Mazuelos E, Lozano F, et al. Correlation of fluconazole MICs with clinical outcome in cryptococcal infection. Antimicrob Agents Chemother. 2000;44:1544–1548. doi:10.1128/aac.44.6.1544-1548.2000
26. Pierard GE, Arrese JE, Pierard-Franchimont C. Itraconazole. Expert Opin Pharmacother. 2000;1:287–304. doi:10.1517/14656566.1.2.287
27. Pan W, Khayhan K, Hagen F, et al. Resistance of Asian Cryptococcus neoformans serotype A is confined to few microsatellite genotypes. PLoS One. 2012;7:e32868. doi:10.1371/journal.pone.0032868
28. Hagen F, Hare Jensen R, Meis JF, Arendrup MC. Molecular epidemiology and in vitro antifungal susceptibility testing of 108 clinical Cryptococcus neoformans sensu lato and Cryptococcus gattii sensu lato isolates from Denmark. Mycoses. 2016;59:576–584. doi:10.1111/myc.12507
29. Worasilchai N, Tangwattanachuleeporn M, Meesilpavikkai K, et al. Diversity and antifungal drug susceptibility of cryptococcus isolates in Thailand. Med Mycol. 2017;55:680–685. doi:10.1093/mmy/myw130
30. Chowdhary A, Randhawa HS, Sundar G, et al. In vitro antifungal susceptibility profiles and genotypes of 308 clinical and environmental isolates of Cryptococcus neoformans var. grubii and Cryptococcus gattii serotype B from north-western India. J Med Microbiol. 2011;60:961–967. doi:10.1099/jmm.0.029025-0
31. Pfaller M, Zhang J, Messer S, et al. Molecular epidemiology and antifungal susceptibility of Cryptococcus neoformans isolates from Ugandan AIDS patients. Diagn Microbiol Infect Dis. 1998;32:191–199. doi:10.1016/S0732-8893(98)00095-9
32. Franzot SP, Hamdan JS. In vitro susceptibilities of clinical and environmental isolates of Cryptococcus neoformans to five antifungal drugs. Antimicrob Agents Chemother. 1996;40:822–824. doi:10.1128/AAC.40.3.822
33. Archibald LK, Tuohy MJ, Wilson DA, et al. Antifungal susceptibilities of Cryptococcus neoformans. Emerg Infect Dis. 2004;10:143–145. doi:10.3201/eid1001.020779
34. Trilles L, Fernandez-Torres B, Lazera Mdos S, Wanke B, Guarro J. In vitro antifungal susceptibility of Cryptococcus gattii. J Clin Microbiol. 2004;42:4815–4817. doi:10.1128/jcm.42.10.4815-4817.2004
35. Chen YC, Chang SC, Shih CC, et al. Clinical features and in vitro susceptibilities of two varieties of Cryptococcus neoformans in Taiwan. Diagn Microbiol Infect Dis. 2000;36:175–183. doi:10.1016/S0732-8893(99)00137-6
36. Mitchell TG, Perfect JR. Cryptococcosis in the era of AIDS-100 years after the discovery of Cryptococcus neoformans. Clin Microbiol Rev. 1995;8:515–548. doi:10.1128/CMR.8.4.515
37. Pfaller MA, Messer SA, Jones RN, Castanheira M. Antifungal susceptibilities of Candida, Cryptococcus neoformans and Aspergillus fumigatus from the Asia and Western Pacific region: data from the SENTRY antifungal surveillance program (2010-2012). J Antibiot (Tokyo). 2015;68:556–561. doi:10.1038/ja.2015.29
38. Selb R, Fuchs V, Graf B, et al. Molecular typing and in vitro resistance of Cryptococcus neoformans clinical isolates obtained in Germany between 2011 and 2017. Int J Med Microbiol. 2019;309:151336. doi:10.1016/j.ijmm.2019.151336
39. Espinel-Ingroff A, Chowdhary A, Cuenca-Estrella M, et al. Cryptococcus neoformans-Cryptococcus gattii species complex: an international study of wild-type susceptibility endpoint distributions and epidemiological cutoff values for amphotericin B and flucytosine. Antimicrob Agents Chemother. 2012;56:3107–3113. doi:10.1128/AAC.06252-11
40. O’Shaughnessy EM, Lyman CA, Walsh TJ. Amphotericin B: polyene resistance mechanisms. In: Mayers DL, editor. Antimicrobial Drug Resistance: Mechanisms of Drug Resistance. Totowa, NJ: Humana Press; 2009:295–305.
41. Herkert PF, Meis JF, Lucca de Oliveira Salvador G, et al. Molecular characterization and antifungal susceptibility testing of Cryptococcus neoformans sensu stricto from southern Brazil. J Med Microbiol. 2018;67:560–569. doi:10.1099/jmm.0.000698
42. Espinel-Ingroff A, Aller AI, Canton E, et al. Cryptococcus neoformans-Cryptococcus gattii species complex: an international study of wild-type susceptibility endpoint distributions and epidemiological cutoff values for fluconazole, itraconazole, posaconazole, and voriconazole. Antimicrob Agents Chemother. 2012;56:5898–5906. doi:10.1128/aac.01115-12
43. Chong HS, Dagg R, Malik R, Chen S, Carter D. In vitro susceptibility of the yeast pathogen cryptococcus to fluconazole and other azoles varies with molecular genotype. J Clin Microbiol. 2010;48:4115–4120. doi:10.1128/jcm.01271-10
44. Iqbal N, DeBess EE, Wohrle R, et al. Correlation of genotype and in vitro susceptibilities of Cryptococcus gattii strains from the Pacific Northwest of the United States. J Clin Microbiol. 2010;48:539–544. doi:10.1128/jcm.01505-09
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.