Back to Journals » ImmunoTargets and Therapy » Volume 15
Cryoablation Plus Immune Checkpoint Inhibitors Enhanced Dendritic Cell and T Cell Activation in TNBC Murine Model
Authors Sardela de Miranda F ![]() , Babcock RL, Mahecha MF, Gukhool PJ, Boligala GP, Garcia AK, Martinez-Marin D, Oliver AG, Hintelmann T, Furr KL, Bose C, Almodovar S, Singh SP, Pruitt K, Melkus MW, Layeequr Rahman R
, Babcock RL, Mahecha MF, Gukhool PJ, Boligala GP, Garcia AK, Martinez-Marin D, Oliver AG, Hintelmann T, Furr KL, Bose C, Almodovar S, Singh SP, Pruitt K, Melkus MW, Layeequr Rahman R
Received 16 November 2025
Accepted for publication 6 March 2026
Published 7 April 2026 Volume 2026:15 581990
DOI https://doi.org/10.2147/ITT.S581990
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Michael Shurin
Flavia Sardela de Miranda,1– 3 Rachel L Babcock,1– 4 Maria F Mahecha,1,2 Prrishti J Gukhool,1,2 Geetha P Boligala,3,4 Amanda K Garcia,5 Dalia Martinez-Marin,4 Ava G Oliver,5 Thomas Hintelmann,5 Kathryn L Furr,1,2 Chhanda Bose,6 Sharilyn Almodovar,5 Sharda P Singh,1,2,7 Kevin Pruitt,4 Michael W Melkus,1,2 Rakhshanda Layeequr Rahman1,2
1Department of Surgery, Texas Tech University Health Sciences Center, Lubbock, TX, 79430, USA; 2Breast Center of Excellence, Texas Tech University Health Sciences Center, Lubbock, TX, 79430, USA; 3Department of Cell Biology and Biochemistry, Texas Tech University Health Sciences Center, Lubbock, TX, 79430, USA; 4Department of Pharmacology, University of North Carolina at Chapel Hill, Chapel Hill, NC, 27599, USA; 5Department of Immunology and Molecular Microbiology, Texas Tech University Health Sciences Center, Lubbock, TX, 79430, USA; 6Department of Pharmacology and Neuroscience, Texas Tech University Health Sciences Center, Lubbock, TX, 79430, USA; 7Department of Internal Medicine, Tech University Health Sciences Center, Lubbock, TX, 79430, USA
Correspondence: Michael W Melkus, Department of Surgery, TTUHSC, 3601 4th Street, Lubbock, TX, 79430-8312, USA, Email [email protected] Rakhshanda Layeequr Rahman, Breast Health Center, Creighton University-CHI Health Lakeside, 16901 Lakeside Hills Ct, Omaha, NE, 68130, USA, Email [email protected]
Purpose: Cryoablation eradicates tumors through repeated freeze–thaw cycles and preserves tumor-associated antigens, triggering inflammatory signals capable of priming anti-tumor immunity, yet its therapeutic potential in triple-negative breast cancer (TNBC) remains largely unexplored. Immune checkpoint inhibitors (ICIs) have shown clinical benefit in TNBC but come with significant immune-related toxicities. Combining cryoablation with ICIs in TNBC may amplify the efficacy of cryoablation, which is significantly less toxic than ICIs, thereby providing opportunities for lowering the doses of ICIs in clinical practice. Here, we investigated the therapeutic impact of cryoablation with ICIs in an orthotopic bilateral murine TNBC model.
Methods: Two weeks after tumor induction, primary tumors were cryoablated while the abscopal tumors were not manipulated and represented distant tumors. Twenty-four hours pre- and post-cryoablation, mice received an intra-peritoneal injection of PBS or ICIs (anti-CTLA-4, PD-1, or PD-L1). Tumors, tumor-draining lymph nodes (TdLNs), spleen, and peripheral blood were assessed for immune profiling a week later.
Results: Preliminary analyses demonstrated that combining cryoablation with anti-CTLA-4 enhanced T cell activation systemically compared to cryoablation alone or in combination with PD-1/PD-L1 blockade. Notably, relative to cryoablation monotherapy, combination with anti-CTLA-4 increased the frequencies of activated CD4⁺ and CD8⁺ T cells in the abscopal tumors, while also inducing a higher frequency and activation of conventional dendritic cells in the abscopal TdLNs.
Conclusion: These results suggest combination of cryoablation with anti-CTLA-4 therapy enhances systemic antitumor immunity by boosting antigen presentation. Our results support further investigation into this combination strategy to prevent tumor recurrence and metastasis while minimizing toxicity of treatment.
Keywords: triple-negative breast cancer, cryoablation, immune checkpoint inhibitors, anti-tumor immune response, abscopal effect
Introduction
Surgery, radiotherapy, and chemotherapy remain central to cancer treatment; however, newer approaches such as targeted therapy and immunotherapy have recently revolutionized the oncology field.1 Initially, immunotherapy was mainly focused on the use of immunocytokines and cancer vaccines, which were highly toxic and mostly ineffective.2,3 However, with the development of immune checkpoint inhibitors (ICI), the field witnessed major achievements, and most immunotherapy efforts have ever since been focused on these molecules.
Immune checkpoints receptors are expressed on the surface of immune cells. Generally divided into co-inhibitory and co-stimulatory proteins, immune checkpoints maintain the balance between preserving self-tolerance and eliminating threats.4 T cell activation normally requires two signals: recognition of a specific antigen by the T cell receptor (TCR) in the context of major histocompatibility complex (MHC) molecules, and engagement of the co-stimulatory receptor CD28 with its co-stimulatory ligands, CD80/86. These signals promote clonal expansion, cytokine production, and the acquisition of effector functions, allowing T cells to mount an effective immune response.5 Co-inhibitory receptors, such as the Cytotoxic T-Lymphocyte Antigen 4 (CTLA-4) and the Programmed Cell Death Protein 1 (PD-1), function to suppress T cell activity and prevent overactivation of the immune system.
Tumors utilize the CTLA-4 and PD-1 pathways to evade host immune response. CTLA-4 is expressed on the surface of T cells during their early stages of activation, and when bound to co-stimulatory ligands CD80/86, it transmits inhibitory signals, increasing the threshold for T cell activation. Additionally, CTLA-4 has a higher affinity for and sequesters CD80/86, preventing their binding to CD28, which results in decreased availability of stimulatory signals for T cell activation.6,7 Specifically, on regulatory T cells (Tregs), CTLA-4 engagement also enhances their immunosuppressive activity.8 Meanwhile, PD-1 is predominantly expressed on the surface of activated T cells during its effector phase9 and it interacts with two ligands: Programmed Death Ligand 1 (PD-L1) or 2 (PD-L2).10 PD-L1 is expressed on the surface of various cells, including T cells, B cells, dendritic cells (DC), macrophages, and tumor cells, while PD-L2 is primarily found on macrophages, DC, and non-hematopoietic cells present, for example, in the lungs.1 Once bound to its ligands, PD-1 signals intracellularly to prevent T cell activation, cytokine production, and proliferation.11–13
As an important tumor evasion mechanism, the expression of immune checkpoint proteins and its ligands are frequently dysregulated by tumors to impair T cell anti-tumoral functions.14 Therefore, blockade of these inhibitory signals can enhance the immune system’s ability to fight cancer by removing the brakes on T cell activation and effector function. The first FDA approved ICI was ipilimumab, an anti-CTLA-4 blocking antibody; this was rapidly followed by the development of anti-PD-1 and anti-PD-L1 inhibitors, such as pembrolizumab and atezolizumab, which have become some of the most widely prescribed anticancer therapies, currently approved to treat various cancers,15,16 including triple-negative breast cancer (TNBC).17,18
TNBCs are characterized by the lack of expression of estrogen, progesterone, and HER2 receptors19 and, therefore, cannot be treated with established targeted therapies.20 Additionally, TNBCs present an inherently aggressive clinical behavior and remarkable heterogeneity, leading to poorer outcomes when compared to other breast cancer subtypes.20 Despite chemotherapy,21 patients remain at high risk for disease recurrence and metastasis,22–25 where ICI could provide hopes of better outcomes, since TNBC is the most immunogenic subtype of breast cancer. However, response rates to single agent ICI in metastatic TNBC have been modest and associated with immune-related adverse events, suggesting that combination of checkpoint blockade with other approaches might provide favorable outcomes and allow dose de-escalation, decreasing side effects.26
Cryoablation is a promising candidate for combination with immune checkpoint inhibition therapy. This minimally invasive procedure induces target tissue necrosis through controlled freeze–thaw cycles while preserving tumor-associated antigens (TAAs) and releasing inflammatory signals.27 These effects can stimulate antitumor immune responses and potentially enhance the abscopal effect - a systemic immune response that targets distant, untreated tumors.28–30 This contrasts with surgical resection, where the tumor and, consequently, TAAs are removed.30 Currently, cryoablation is clinically approved for the treatment of fibroadenomas and low-risk breast cancers31–35 and there is limited clinical data for its use in TNBC.
Pre-clinical evidence previously published by our group indicates that local treatment of TNBC by cryoablation increases systemic immune response compared to resection, where 40% of the mice in the resected group showed local recurrence and lung metastasis, while no recurrence or metastasis occurred post-cryoablation.36 This is in agreement with other papers that show superior immune activation with cryoablation compared to resection in breast cancer37 and colon cancer38 models. Furthermore, in a bilateral TNBC mouse model, cryoablation of the primary tumor led to significant increases in the frequency of NK cells at the secondary tumor, with similar trends for effector/effector memory CD4+ and CD8+ T cells, while a significant rise in the frequency of migratory dendritic cells was detected in the spleen. This superior immune activation is likely correlated to better tumor control, as secondary unmanipulated tumors from cryoablation were significantly smaller than secondary tumors from resection.39 This supports the use of cryoablation in TNBC, showing its potential to provide antigens and inflammatory signals to stimulate immune activation.
Despite the safety of cryoablation and reported benefits of antigen preservation, one challenge remains: immunosuppression. Although cryoablation recruits immune cells and generates tumor antigens for uptake by antigen-presenting cells (APCs), high expression of checkpoint molecules within the tumor immune microenvironment (TIME) or secondary lymphoid organs can suppress T cell activation and effector functions, thereby limiting therapeutic efficacy.40–42 Thus, combination of cryoablation with ICI offers a synergistic approach, combining the benefits of each approach, while tackling their weaknesses. In this study, we used a bilateral TNBC mouse model and flow cytometry to evaluate the combination of cryoablation to either anti-CTLA-4, anti-PD-1, or anti-PD-L1. We selected a 1-week immunological endpoint to capture the critical early phase of immune priming after cryoablation combined with immune checkpoint inhibition. Cryoablation induces rapid tumor necrosis, releasing tumor antigens and danger signals that drive acute inflammation, dendritic cell activation, and initiation of adaptive immunity within the first 3–7 days.37,39,43 Immune checkpoint inhibitors act primarily during early T-cell priming and expansion, amplifying emerging anti-tumor responses.44,45 Moreover, early immunological readouts have been shown to serve as predictive biomarkers of downstream adaptive immune responses and clinical outcomes.44 Furthermore, a deeper understanding of the fundamental mechanisms governing early T-cell activation may help explain the limited efficacy of ICI monotherapy in high-risk breast cancers.46 Therefore, assessment at 1 week reflects the convergence of antigen release and checkpoint-mediated immune activation, whereas later time points are more likely to represent secondary or compensatory processes rather than primary mechanisms of therapeutic synergy.
We found CTLA-4 inhibition best synergized with cryoablation, providing enhanced frequencies of activated CD4+ and CD8+ T cells in the spleen and blood. Additionally, cryoablation plus anti-CTLA-4 led to higher frequencies of activated CD4+ and CD8+ T cells in unmanipulated secondary (abscopal) tumors, mirrored by higher frequencies of central memory CD8+ T cells and conventional DCs (cDCs) in tumor-draining lymph nodes (TdLNs). These exciting results support the combination of cryoablation with anti-CTLA-4, which should next be tested on response longevity and specificity.
Materials and Methods
Cell Line
The luciferase-expressing triple-negative mammary carcinoma cell line, 4T1-12B, was generously provided by Dr. G. Gary Sahagian from TUFTS University.47 Cells were cultured at 37°C degrees and 5% CO2 concentration in Dulbecco’s Modified Eagle’s medium (DMEM) supplemented with 10% FBS and 1% penicillin/streptomycin.
Mice
Naïve female BALB/c mice (8–11 weeks old) were obtained from The Jackson Laboratory (RRID:IMSR_JAX:000651, Bar Harbor, ME, USA) and housed in a pathogen-free facility at the Laboratory Animal Resource Center (LARC) under a 12-hour light/12-hour dark cycle. Mice had free access to water and standard chow and were routinely monitored for health. All procedures were approved by the Institutional Animal Care and Use Committee (IACUC) of Texas Tech University Health Sciences Center (Protocol 24001) and conducted in accordance with the ARRIVE guidelines (Animal Research: Reporting of in vivo experiments).
Treatments
Naïve female BALB/c mice underwent bilateral orthotopic transplant with 1×106 4T1-12B cells per side. At two weeks, the primary (left) tumors were cryoablated, while the abscopal (right) tumors were not manipulated to represent distant tumors, used to measure the abscopal effect (Supplementary Figure 1). Two days before cryoablation, primary and abscopal tumors were measured with calipers by the same individual to ensure measurement consistency, and mice were divided into different groups with similar tumor volume averages. The cryoablation procedure was performed in a blinded manner (the individual performing the cryoablation was blinded to group assignments) and under strict aseptic technique, as detailed below and consistent with our previously published methods.36,39 Mice were anesthetized with 2.5% inhaled isoflurane, and the skin was incised and retracted away from the primary tumor. The Visica 2 treatment system (Sanarus Technologies, Inc., Pleasanton, CA, USA) or ProSense treatment system (IceCure Medical, Caesarea, Israel) probe connected to liquid nitrogen cryoablation machines were placed directly on top of the tumor mass (Figure 1A); each primary tumor underwent two freeze/thaw cycles and was left in vivo to induce the abscopal effect. The freezing time was dependent on tumor size, while all tumors were equally given 3 minutes to completely thaw between the two freezing cycles. Next, the skin was closed using 4–0 Prolene sutures (Ethicon, Somerville, NJ). To manage pain and discomfort from the procedure, 0.05 mg/kg buprenorphine (Par Pharmaceuticals, Chestnut Ridge, NY) diluted in 200 µL of saline was administered intraperitoneally, and one cup of MediGel Hazelnut (Clear H2O®, Portland, ME) supplemented with 0.5 mg/mL Rimadyl (Zoetis, Parsippany, NJ) was provided per cage. Animals were monitored for 5 days for post-procedure complications.
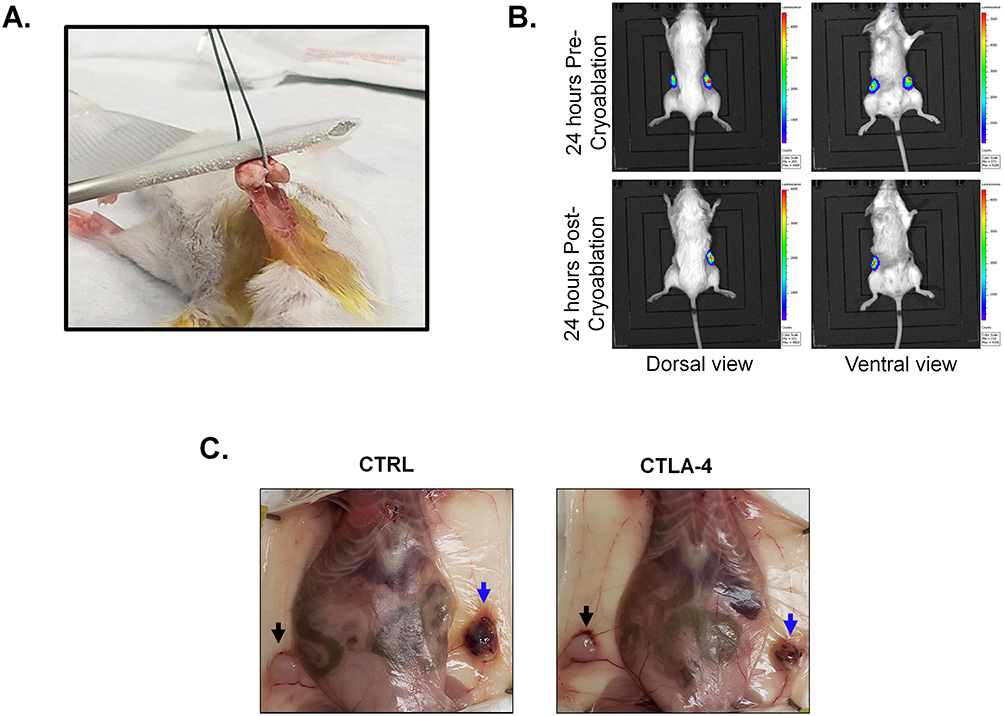 |
Figure 1 Mouse model. (A) Representative photo from the mouse mammary carcinoma cryoablation. (B) IVIS images 24 hours pre- and post-cryoablation (top and bottom, respectively). Images on the left show dorsal view, while images on the right show ventral view. (C) Representative photos of mouse necropsy (CTRL = control; CTLA-4 = anti-CTLA-4) showing the cryoablated (blue arrow) and abscopal (black arrow) tumors. |
Prior to cryoablation, mice received via IP injection an initial 100 µg dose of anti-CTLA4 (Clone UC10-4F10-11, Cat# BE0032, BioXcell, NH), anti-PD-1 (Clone J43, Cat# BP0033-2, BioXcell, NH) or anti-PD-L1 (Clone 10F.9G2, Cat# BP0101, BioXcell, NH) to prime the immune system, followed by a second post-procedure 100 µg ICI boosting dose (total ICI administration is 200 ug/mouse). The ICI dosage used for these experiments are well established in preclinical murine models of ICI blockade with 100–250 µg per mouse routinely achieving robust T-cell activation and anti-tumor efficacy without overt toxicity.34,48–50 This regimen provides a biologically relevant and validated exposure for evaluating early systemic immune responses and the synergistic effects of cryoablation with checkpoint inhibition. Control mice were administered PBS (Phosphate buffered saline). One-week post-cryoablation, all mice were euthanized, and their organs were processed for anti-tumor immune analyses. Two independent experiments were conducted in accordance with TTUHSC-IACUC policies and approved protocols. Animals found dead during the study were excluded from the analyses. The experimental approach is illustrated in Supplementary Figure 1.
Tumor Monitoring
To check the location of luciferase-expressing tumors before cryoablation and confirm efficiency of the procedure in eliminating the tumor cells, mice received 100 µL of D-luciferin, potassium salt (15 mg/mL) (GoldBio, St. Louis, MO) intraperitoneally (IP) and were imaged with the in vivo imaging system (IVIS) Lumina XR (Caliper Life Sciences, PerkinElmer™) twenty-four hours pre- and post- cryoablation, consistent with our previous published methods (Figure 1B).39
Tissue and Blood Collection
One week after cryoablation, euthanasia was performed using carbon dioxide (CO2) inhalation, delivered from a compressed gas cylinder at a flow rate of 20–30% of the chamber volume per minute, followed by cardiac puncture to ensure death (Figure 1C). All procedures were conducted in accordance with the American Veterinary Medical Association (AVMA) Guidelines for the Euthanasia of Animals (2020).51 Peripheral blood obtained by cardiac puncture was collected into tubes containing 0.5 M EDTA. Primary and abscopal tumors, tumor-draining lymph nodes (TdLNs), and spleens were collected into tubes containing sterile PBS. Tumors were divided for either histological analysis or flow cytometric processing.
Tumor-Infiltrating Lymphocyte (TIL) Quantification
Tumors designated for histology were formalin-fixed, paraffin-embedded, and sectioned. Slides were stained with hematoxylin and eosin (H&E). For each mouse, two distinct fields were selected from the invasive margin of the tumor. Lymphocytes within these areas were manually counted under light microscopy by a trained observer blinded to experimental groups. Each group consisted of five mice, resulting in ten individual scoring areas per group. Counts from the two fields per mouse were averaged to obtain a single TIL count per animal, which was then used for statistical analysis.
Sample Processing for Flow Cytometry
For all downstream steps and washes, cells were resuspended in FACS buffer (PBS supplemented with 5% FBS). Spleens and TdLNs were mechanically dissociated by pressing through a 70 µm nylon mesh filter. Tumors designated for flow cytometry were minced and enzymatically digested with 300 U/mL collagenase, 100 U/mL hyaluronidase, and 150 µg/mL DNase, then passed through a 70 µm nylon mesh filter to obtain single-cell suspensions. Red blood cells (RBCs) in blood, spleen, and tumor suspensions were removed using RBC lysis buffer (STEMCELL Technologies, Vancouver, Canada). Following lysis, tumor cell suspensions were resuspended in FACS buffer containing 25 µg/mL DNase to prevent clump formation.
Flow Cytometry Analysis for Immune Phenotyping
Isolated cells from blood, spleen, tumors, and TdLNs were counted to determine cell numbers and viability. 2×106 viable cells were aliquoted from each sample and divided equally between two wells – one stained with a lymphoid panel and the other with a myeloid panel. Flow antibodies used in this study are found in Supplementary Table 1. In cases where samples had less than 2×106 viable cells, the total sample, regardless of cell numbers, was divided equally between two wells and excluded from Median Fluorescence Intensity (MFI) analysis. Both panels included a Live/Dead Aqua viability dye (Thermo Fisher Scientific Inc., Waltham, MA). Prior to staining, Fc receptors were blocked with CD16/CD32 antibody mixture (Invitrogen, San Diego, CA). Flow cytometry data was acquired on a Cytek Northern Lights™-Clinical (NL-CLC) spectral analyzer (Cytek Biosciences, Freemont, CA) using SpectroFlo® software and analyzed in a blinded manner with FlowJo v10.8.1 software.
Debris, doublets, and dead cells were excluded from the analyses, and live immune cells were selected by gating on the CD45 expressing/live-and-dead negative population. Lymphoid and myeloid cells were defined according to the strategy used in our previous study.39 The summary of cell types according to marker expression is shown in Supplementary Table 2.
Power Analysis and Statistical Analysis
Sample sizes (cryoablation alone, cryo + anti-CTLA-4, cryo + anti-PD-1, cryo + anti-PD-L1; n = 10, 10, 5, 5, respectively) were selected to balance statistical rigor with the 3 Rs principle (Replacement, Reduction, Refinement) for ethical animal use in research. Group differences were analyzed by one-way ANOVA with post hoc correction, with a predefined primary comparison of cryo + anti-CTLA-4 versus cryo alone using an unpaired two-tailed Student’s t-test. Based on a 30% expected difference and 20–25% variation, n = 10 per group provided 82–92% power (Cohen’s d = 1.2–1.5, α = 0.05).
Statistical analyses were performed using GraphPad Prism software version 9.00 for Windows (La Jolla CA., https://www.graphpad.com). For preliminary comparisons between cryoablation monotherapy and combination treatments with anti-CTLA-4, anti-PD-1, or anti-PD-L1, one-way ANOVA with Tukey’s post-test was used for normally distributed data, while the Kruskal–Wallis test with Dunn’s post-test was applied for non-normally distributed data. Comparisons specifically between cryoablation alone and cryoablation plus anti-CTLA-4 were assessed using an unpaired t-test or Mann–Whitney U-test for normal and non-normal distributions, respectively. Standard deviation (SD) is reported for each bar. Differences were considered statistically significant when *p<0.05, ** p<0.01, ***p<0.001, and ****p<0.0001, with a 95% confidence interval (CI). Exact p values are reported when relevant to highlight trends.
Analyses of organ weights and lymphoid panel for flow cytometry of tumors, spleen and blood comparing all treatments were performed using five mice per group from one (PD-L1 and PD-1) or two (Control and CTLA-4) independent experiments (shown in Figure 2, and Supplementary Figure 2A; first experiment - pink dots, second experiment - blue dots). Analysis of lymphoid panel for tumors of cryoablation monotherapy and combination with anti-CTLA-4 were performed using five mice per group from two independent experiments (Figures 3 and 4; first experiment - pink dots, second experiment - blue dots). TIL counts were analyzed by evaluating two scoring areas per mouse, with five mice per group, in one experiment (Supplementary Figure 4). Analyses of lymph nodes and for the myeloid panel were conducted with five mice per group from one experiment (shown in Figures 5–7, and Supplementary Figures 5–8; second experiment - blue dots). Finally, analysis of the lymphoid panel for the spleen and blood of cryoablation monotherapy and combination with anti-CTLA-4 were performed using five mice per group from two independent experiments (Figure 8; first experiment - pink dots, second experiment - blue dots).
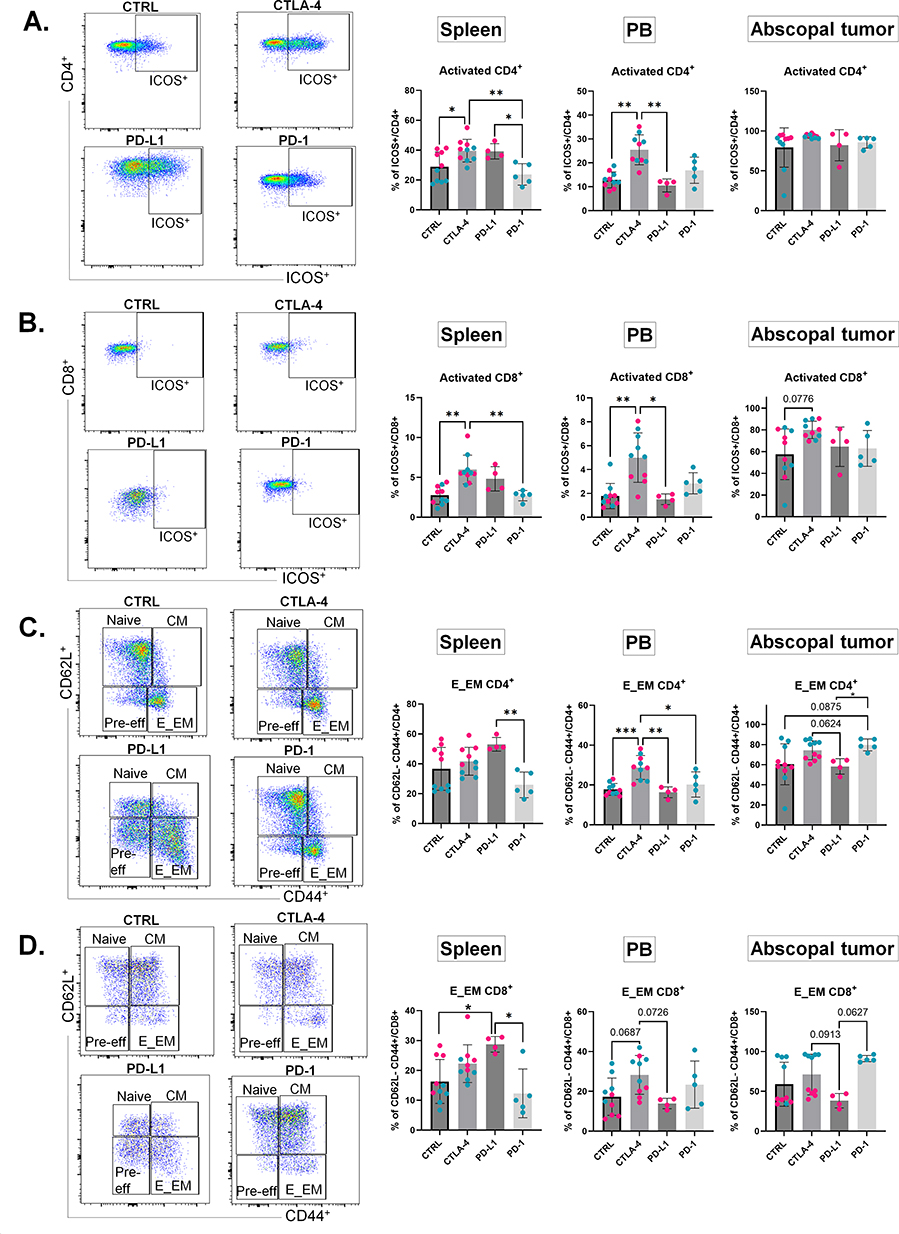 |
Figure 2 Preliminary comparisons of cryoablation monotherapy versus combination with ICIs show CTLA-4 blockade provides superior T cell activation. (A–D) Left: Representative spleen dot plots for CD4+ (A and C) or CD8+ (B and D) T cells in the four treatment groups (CTRL, CTLA-4, PD-L1, and PD-1). Right: Corresponding frequencies of ICOS+ (activated) T cells (A and B) or E_EM T cells (C and D), expressed as a percentage of the parent population, in spleen, peripheral blood, and abscopal tumor. Dot colors indicate individual experiments. One-way ANOVA for normally distributed data or Kruskal–Wallis test for non-normally distributed data was performed comparing cryoablation monotherapy to combination with ICI, with p<0.05 (*), p<0.01 (**), and p<0.001 (***) considered significant. E_EM = effector/effector memory; CM = central memory; CTRL = control, n = 10; CTLA-4 = anti-CTLA-4, n = 10; PD-L1 = anti-PD-L1, n = 5; PD-1 = anti-PD-1, n = 5. |
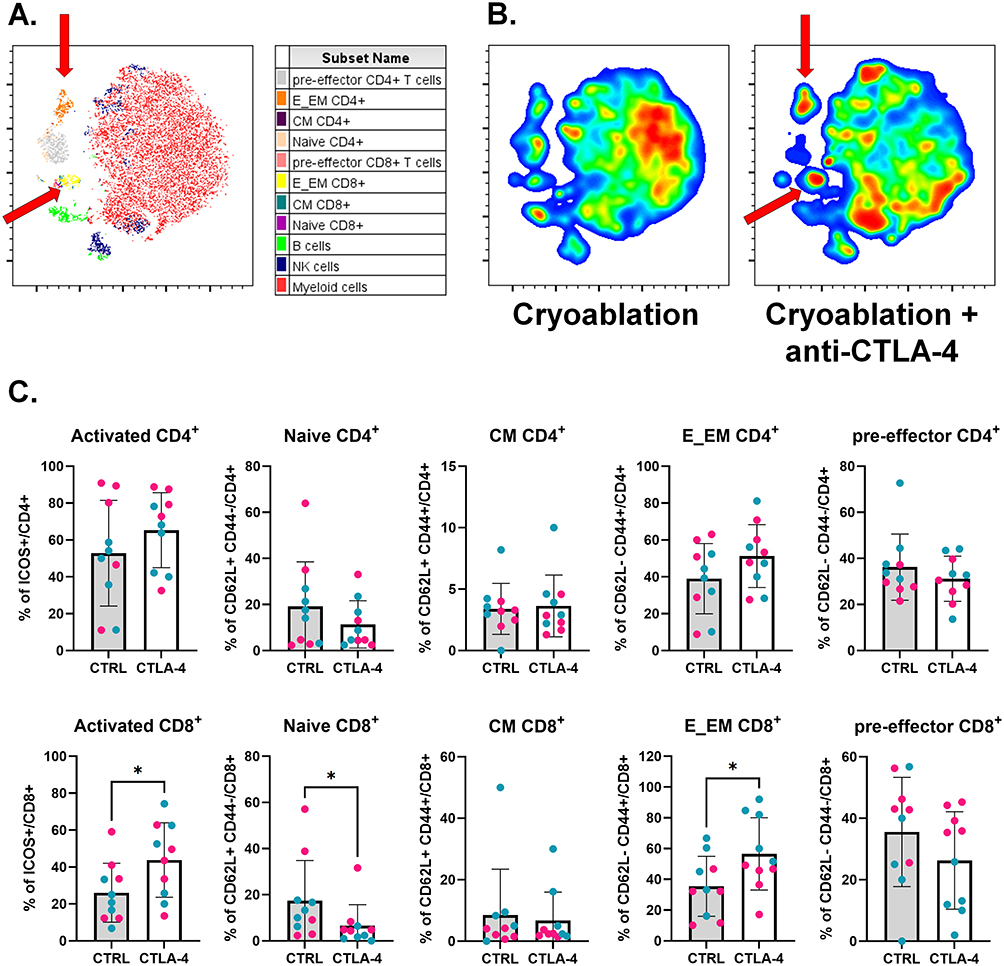 |
Figure 3 Combination of cryoablation with anti-CTLA-4 boosts infiltration of primary tumors by T cells. (A) Representative tSNE map from one experimental repeat, showing the clustering of different lymphoid populations at the primary tumors based on flow cytometry parameters’ similarity. Red arrows point to E_EM populations. (B) Representative tSNE heatmap from one experimental repeat, made from concatenated primary tumors from cryoablation monotherapy versus cryoablation plus anti-CTLA-4. Red arrows point to E_EM populations. (C) Subpopulations of T cells, analyzed as frequency of the parent (CD4+ or CD8+), at the primary tumor. The different dot colors represent different experiments. Unpaired Student’s t-test for normally distributed data or Mann–Whitney test for non-normally distributed data was performed comparing cryoablation monotherapy to combination with ICI, with p<0.05 (*) considered significant. CM = central memory; E_EM = effector/effector memory; CTRL = control; CTLA-4 = anti-CTLA-4. n = 10 per group. |
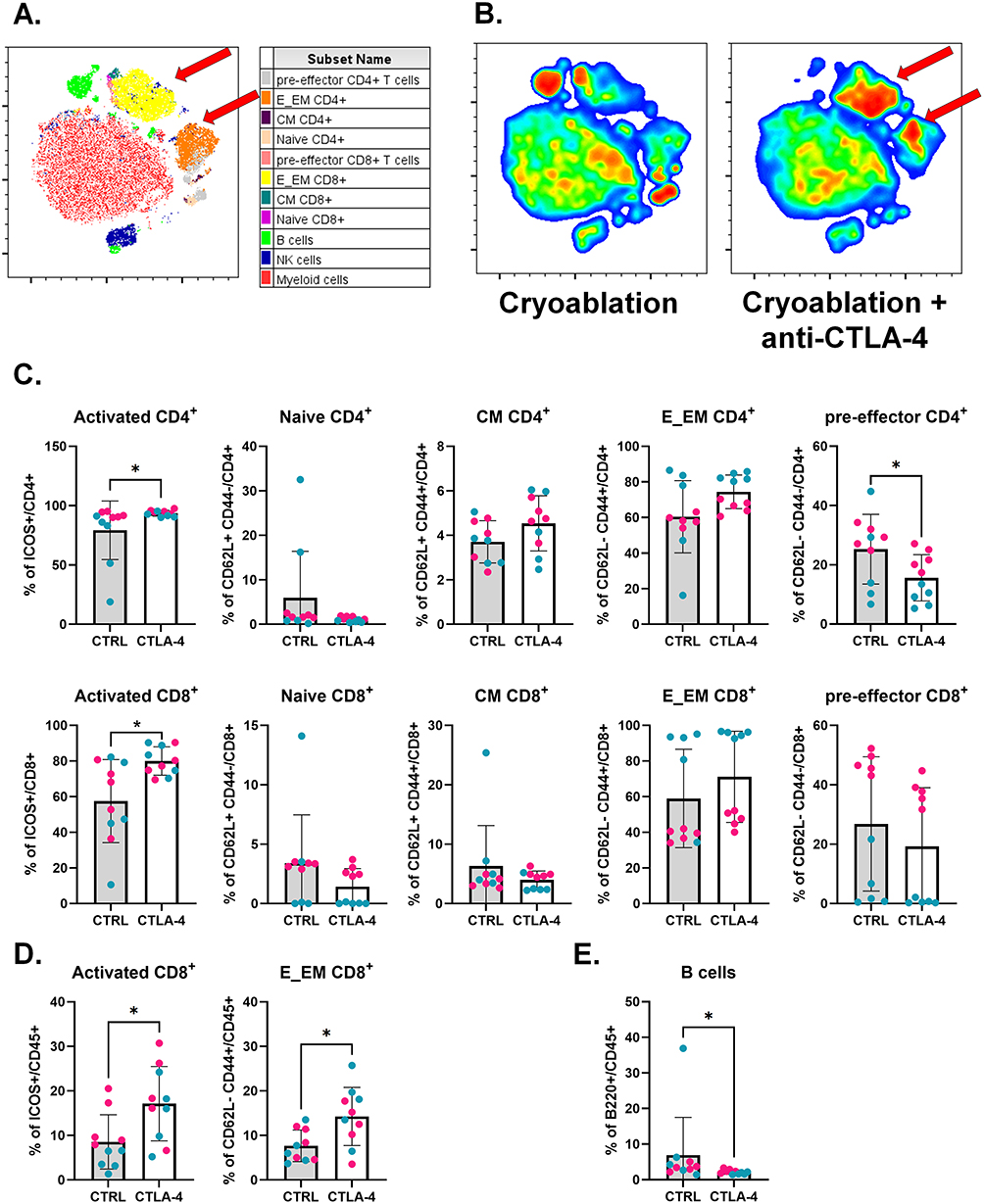 |
Figure 4 Cryoablation plus CTLA-4 blockade increases the frequencies of activated T cells at the abscopal tumors. (A) Representative tSNE map from one experimental repeat, showing the clustering of different lymphoid populations at the abscopal tumors based on flow cytometry parameters’ similarity. Red arrows point to E_EM populations. (B) Representative tSNE heatmap from one experimental repeat, made from concatenated abscopal tumors from cryoablation monotherapy versus cryoablation plus anti-CTLA-4. Red arrows point to E_EM populations. (C) Subpopulations of T cells, analyzed as frequency of the parent (CD4+ or CD8+), at the abscopal tumors. (D) Subpopulations of T cells (activated and effector/effector memory CD8+ T cells), analyzed as frequency of the live immune cells (CD45+), at the abscopal tumors. (E) Parent immune populations, analyzed as frequency of the live immune cells (CD45+), at the abscopal tumors. Dots are colored according to the experiment. Unpaired Student’s t-test for normally distributed data or Mann–Whitney test for non-normally distributed data was performed comparing cryoablation monotherapy to combination with ICI, with p<0.05 (*) considered significant. CM = central memory; E_EM = effector/effector memory; CTRL = control; CTLA-4 = anti-CTLA-4. n = 10 per group. |
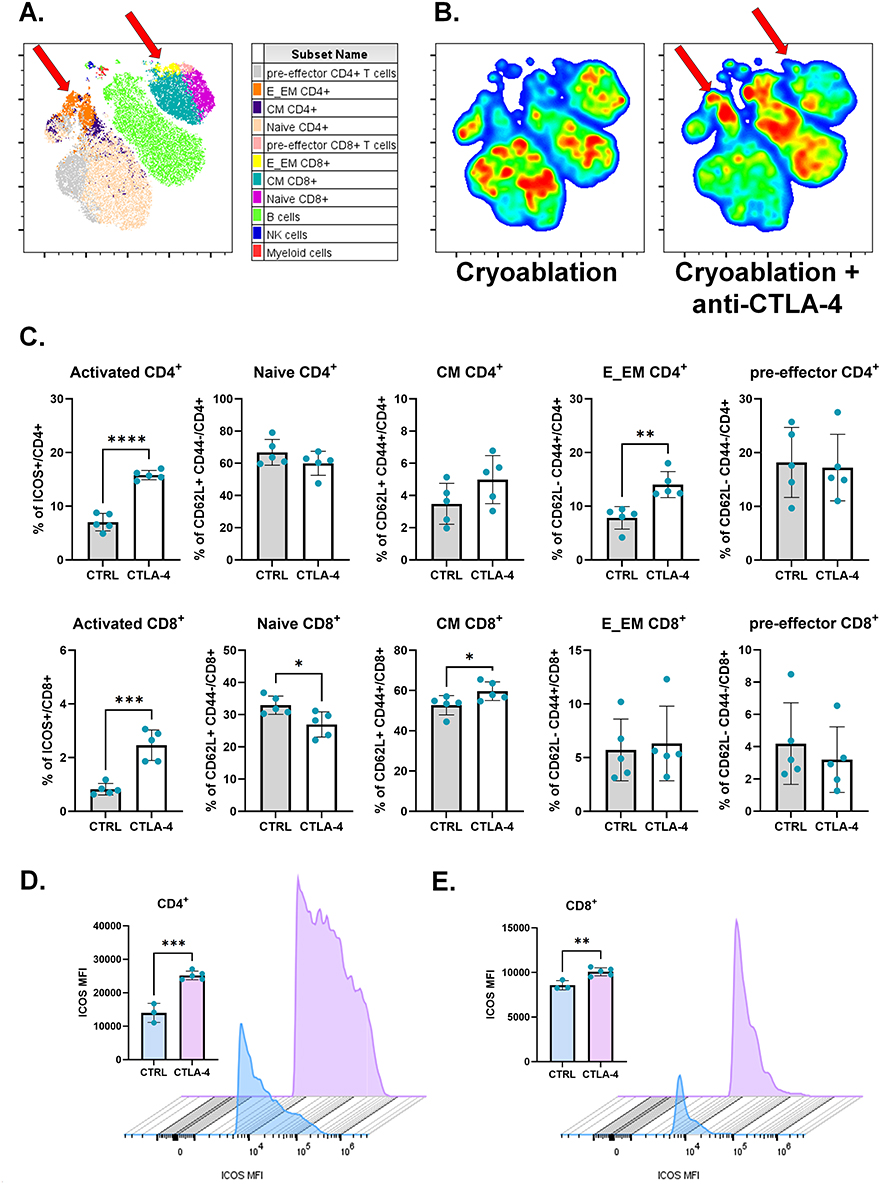 |
Figure 5 Anti-CTLA-4 addition to cryoablation treatment boosts T cell activation at the lymph nodes draining the primary tumor. (A) Representative tSNE map from one experimental repeat, showing the clustering of different lymphoid populations at the primary TdLNs based on flow cytometry parameters’ similarity. Red arrows point to E_EM CD4+ T cell and CM CD8+ T cell populations. (B) Representative tSNE heatmap from one experimental repeat, made from concatenated primary TdLNs from cryoablation monotherapy versus cryoablation plus anti-CTLA-4. Red arrows point to E_EM CD4+ T cell and CM CD8+ T cell populations. (C) Subpopulations of T cells, analyzed as frequency of the parent (CD4+ or CD8+), at the primary TdLNs. (D and E) ICOS median fluorescence intensity (MFI) per count in CD4+ (D) and CD8+ (E) T cells from the from the primary TdLNs. Histograms were generated with concatenated samples from one representative experiment. Unpaired Student’s t-test for normally distributed data or Mann–Whitney test for non-normally distributed data was performed comparing cryoablation monotherapy to combination with ICI, with p<0.05 (*), p<0.01 (**), p<0.001 (***), and p<0.0001 (****) considered significant. CM = central memory; E_EM = effector/effector memory; CTRL = control; CTLA-4 = anti-CTLA-4. n = 5 per group for frequency analysis; n = 3–5 per group for MFI analysis. |
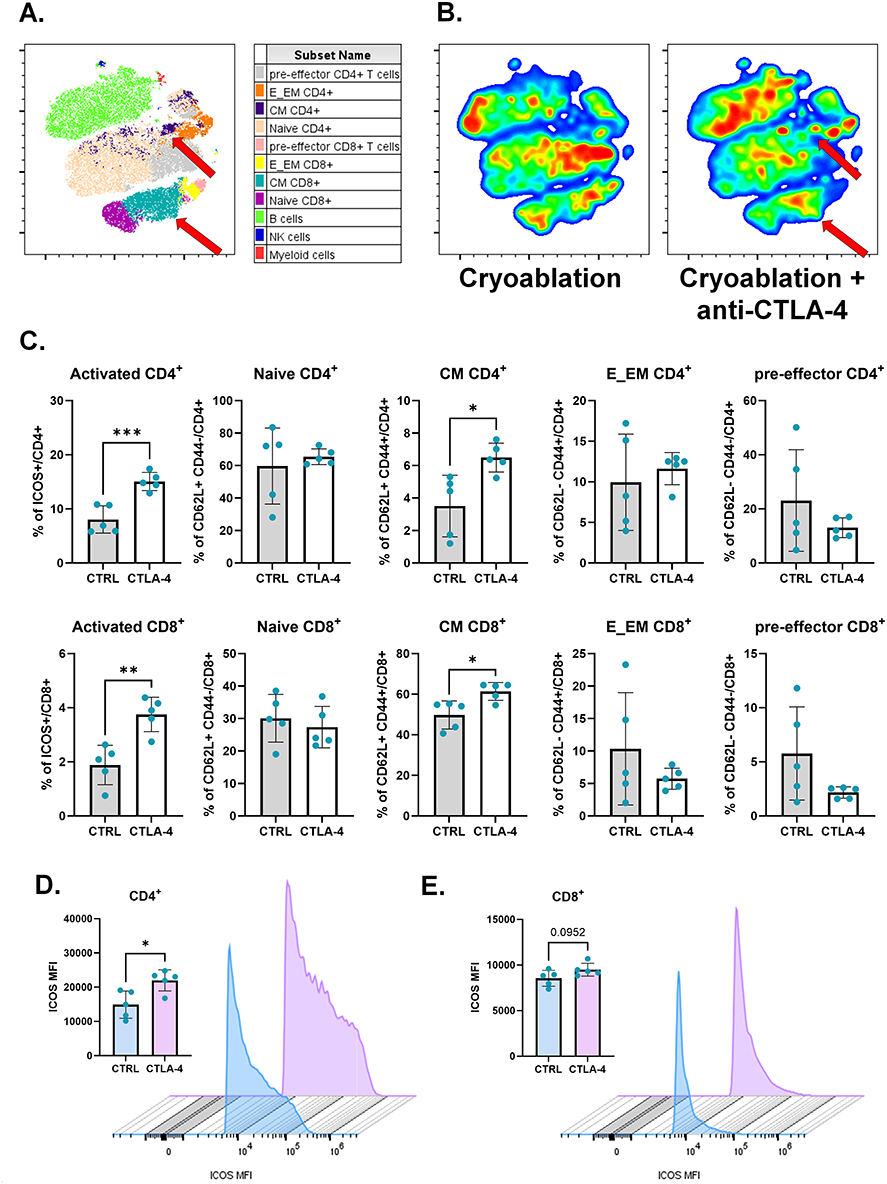 |
Figure 6 Cryoablation plus anti-CTLA-4 results in higher memory formation at lymph nodes draining the abscopal tumors. (A) Representative tSNE map from one experimental repeat, showing the clustering of different lymphoid populations at the abscopal TdLNs based on flow cytometry parameters’ similarity. Red arrows point to CM populations. (B) Representative tSNE heatmap from one experimental repeat, made from concatenated abscopal TdLNs from cryoablation monotherapy versus cryoablation plus anti-CTLA-4. Red arrows point to CM populations. (C) Subpopulations of T cells, analyzed as frequency of the parent (CD4+ or CD8+), at the abscopal TdLN. (D and E) ICOS median fluorescence intensity (MFI) per count in CD4+ (D) and CD8+ (E) T cells from the abscopal TdLNs. Histograms were generated with concatenated samples from one representative experiment. Unpaired Student’s t-test for normally distributed data or Mann–Whitney test for non-normally distributed data was performed comparing cryoablation monotherapy to combination with ICI, with p<0.05 (*), p<0.01 (**), and p<0.001 (***) considered significant. CM = central memory; E_EM = effector/effector memory; CTRL = control; CTLA-4 = anti-CTLA-4. n = 5 per group for frequency analysis; n = 3–5 per group for MFI analysis. |
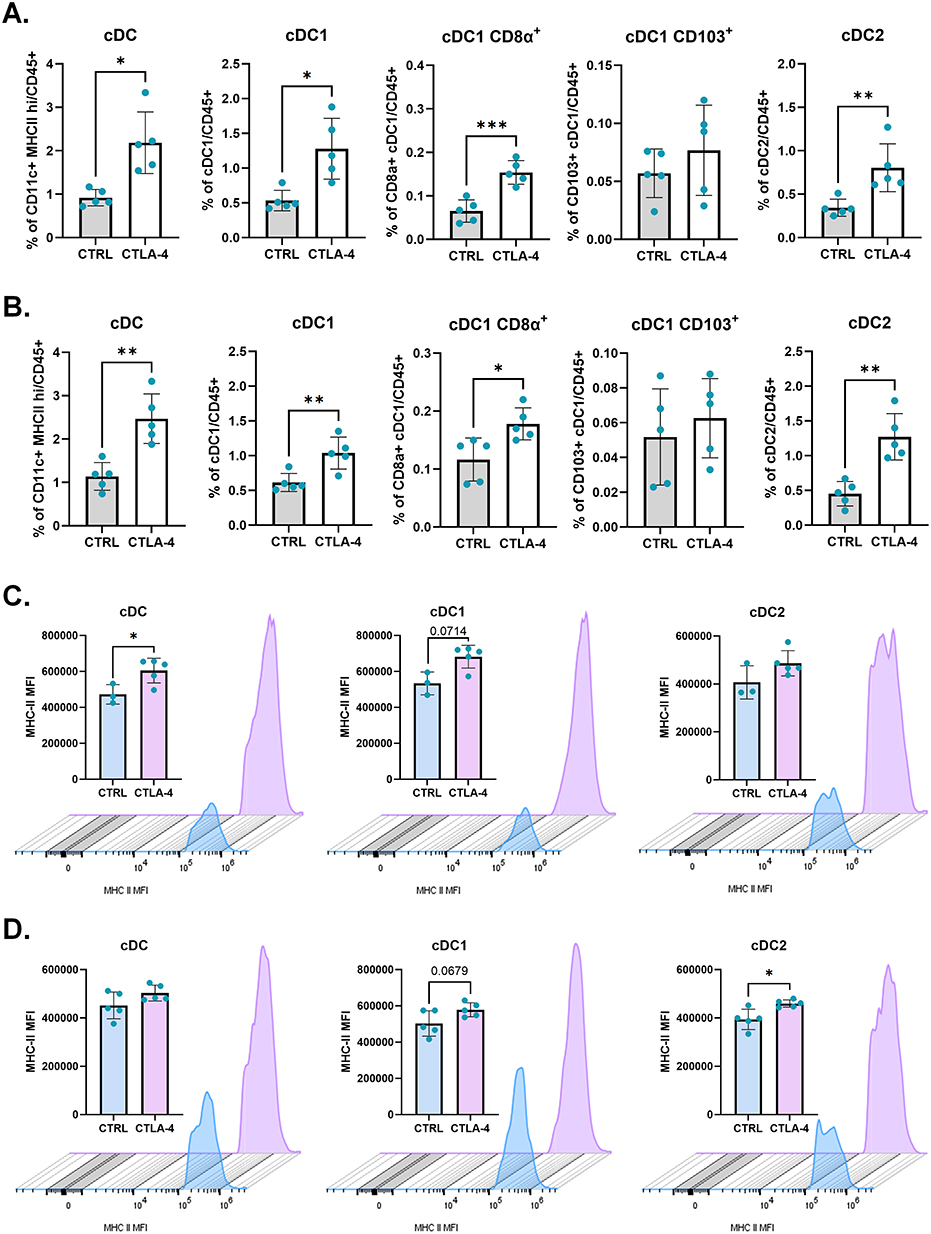 |
Figure 7 Blockade of CTLA-4 leads to enhancement of dendritic cell frequencies at the TdLNs when combined to cryoablation. Frequency of conventional dendritic cells and their subpopulations, analyzed as frequency of the live (CD45+), at the primary (A) and abscopal (B) TdLNs. (C and D) From left to right, MHC class II mean fluorescence intensity (MFI) per count in cDC, cDC1, and cDC2, from the primary (C) abscopal (D) TdLNs. Histograms were generated with concatenated samples from one representative experiment. Unpaired Student’s t-test for normally distributed data or Mann–Whitney test for non-normally distributed data was performed comparing cryoablation monotherapy to combination with ICI, with p<0.05 (*), p<0.01 (**) and p<0.001 (***) considered significant. cDC = conventional dendritic cell; CTRL = control; CTLA-4 = anti-CTLA-4. n = 5 per group for frequency analysis; n = 3–5 per group for MFI analysis. |
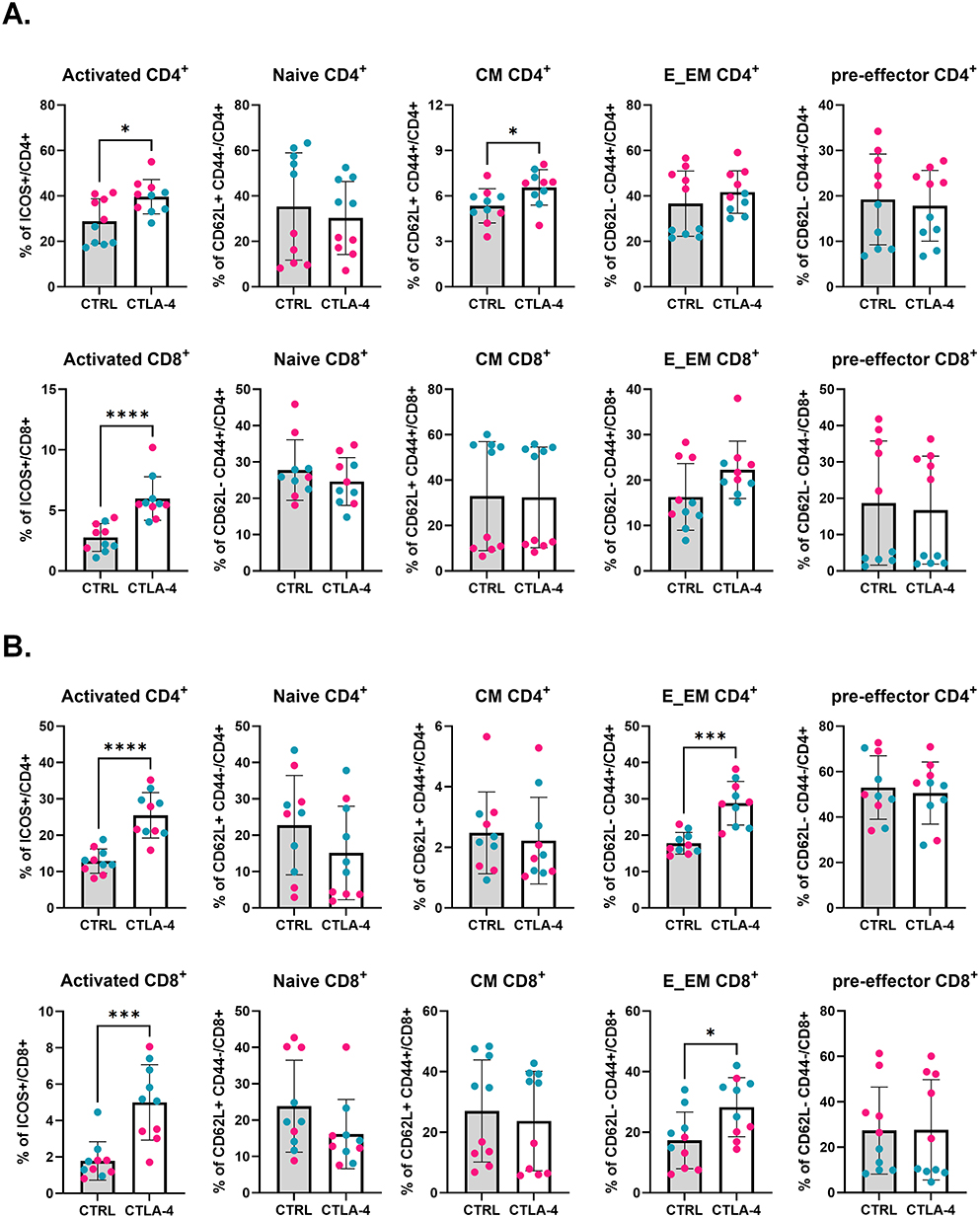 |
Figure 8 The effects of cryoablation with anti-CTLA-4 over T cell activation are not restricted to tumor proximity. (A) Subpopulations of T cells, analyzed as frequency of the parent (CD4+ or CD8+), at the spleen (A) and peripheral blood (B). The different dot colors represent different experiments. Unpaired Student’s t-test for normally distributed data or Mann–Whitney test for non-normally distributed data was performed comparing cryoablation monotherapy to combination with ICI, with p<0.05 (*), p<0.001 (***), and p<0.0001 (****) considered significant. CM = central memory; E_EM = effector/effector memory; CTRL = control; CTLA-4 = anti-CTLA-4. n = 10. |
Results
Inhibition of Anti-CTLA-4 is a Better Combination to Cryoablation than Blockade of PD-1/PD-L1 Axis
Twenty-four hours before and after cryoablation, mice were treated with PBS (control), anti-CTLA-4, anti-PD-1, or anti-PD-L1 antibodies. One mouse from the anti-PD-L1 group died after treatment and was excluded from the analysis. Mice were sacrificed a week later to identify early events involved with the establishment of the anti-tumor immune response. The cryoablation procedure between each experiment and treatment were consistent (Supplementary Figure 2B) demonstrating uniformity and rigor of the experimental design and reproducibility across independent cohorts. Preliminary analyses comparing all four groups showed no differences in the weight of primary tumors, abscopal tumors, or spleens a week after treatment (Supplementary Figure 2A and B). However, analysis of immune cell frequency by flow cytometry revealed that, systemically, cryoablation plus anti-CTLA-4 resulted in significant increases in the frequency of activated CD4+ and CD8+ T cells in the spleen and peripheral blood (Figure 2A and B). Additionally, there was also a significant increase in the frequency of effector/effector memory CD4+ and CD8+ T cells in the peripheral blood (Figure 2C and D). The combination of cryoablation to anti-CTLA-4 was also associated with trends for higher frequencies of activated CD8+ T cells and effector/effector memory CD4+ T cells in the abscopal tumors (Figure 2B and C). Consistently, the majority of ICOS+ cells were effector/effector memory T cells (Supplementary Figure 3A and B). This suggests that combination of cryoablation with anti-CTLA-4 is optimal compared to PD-1/PD-L1 blockade, resulting in enhanced T cell activation. Therefore, further analyses were focused on cryoablation monotherapy versus combination with CTLA-4 (referred to as “combination” on the results below) to dissect the treatment outcomes.
Anti-CTLA-4 Therapy Enhances CD8+ T Cell Infiltration on Primary Tumors a Week After Cryoablation
The effects of the anti-CTLA-4 dose administered to the mice via intraperitoneal injection are apparent in the primary tumor (treated locally with cryoablation), as observed by a significant increase in the TIL counts a week after treatment (Supplementary Figure 4; the TIL counts in the abscopal tumor will be discussed later on section 3.3). To reveal which subsets of TILs are being recruited to these tumors, flow cytometry phenotyping was performed. Unsupervised non-linear dimensionality reduction by t-distributed stochastic neighbor embedding (t-SNE) analysis indicated primary tumors from mice treated with cryoablation plus anti-CTLA-4 presented increased frequencies of effector/effector memory CD4+ and CD8+ when compared to cryoablation monotherapy (Figure 3A and B). Quantitative analysis confirmed there was a significantly higher percentage of activated and effector/effector memory CD8+ T cells in primary tumors when anti-CTLA-4 was added, while a significant decrease in the frequency of naïve CD8+ T cells was also observed (Figure 3C).
Even though the t-SNE analysis indicated potential differences in myeloid infiltration in the primary tumors (Supplementary Figure 5A and B), quantitative comparisons revealed no significant differences in the frequency of macrophages, neutrophils, or dendritic cells in the TIME (Supplementary Figure 5C). Thus, results suggest cryoablation plus anti-CTLA-4 increased recruitment of T cells into the primary tumor, while not affecting infiltration of myeloid subtypes at this early timepoint.
Combination of Cryoablation to Anti-CTLA-4 Also Affects Distant Tumors
Previous work from our group showed cryoablation by itself can increase the infiltration of abscopal tumors by anti-tumor immune cells, especially NK cells.39 Here, while TIL count results found no increase in the overall infiltration by T cells with the combination of cryoablation with anti-CTLA-4 (Supplementary Figure 4), we importantly revealed changes in the frequencies of T cell subpopulations. Analysis by t-SNE indicated the combination increased the frequencies of infiltrating effector/effector memory CD4+ and CD8+ T cells in abscopal tumors, compared to cryoablation monotherapy (Figure 4A and 4B). Quantitative analysis showed a significant increase in the frequencies of activated CD4+ and CD8+ T cells (Figure 4C). Specific assessment of the T cell subtypes within the total live immune population found combination with anti-CTLA-4 also increased the frequency of effector/effector memory CD8+ T cells, in agreement with the t-SNE findings (Figure 4D). Additionally, the combination resulted in a significant decrease in the frequency of B cells (Figure 4E). Similarly to primary tumors, there were no differences in infiltration by myeloid cells in the abscopal tumors (Supplementary Figure 6A–C). These data suggest that, although distant from the primary cryoablation site, abscopal tumors exhibited a significantly higher frequency of activated CD4⁺ and CD8⁺ T cells following systemic treatment with anti-CTLA-4.
Cryoablation Plus Anti-CTLA-4 Treatment Induces Higher Frequencies of Activated T Cell Populations in the Tumor-Draining Lymph Nodes
Tumor-draining lymph nodes are the primary sites at which anti-tumor lymphocytes are activated by TAAs and were therefore examined. Both t-SNE and quantitative analyses demonstrated that treatment of mice with cryoablation plus anti-CTLA-4 increased the frequency of activated CD4+ and CD8+ T cells, effector/effector memory CD4+ T cells and central memory CD8+ T cells in the primary TdLNs (Figure 5A–C), and induced significant increases in ICOS median fluorescence intensity (MFI) on T cells (Figure 5D and E). Overall, the treatment seemed to decrease the frequency of T cells relative to all immune cells in those lymph nodes, however, further analysis indicated the outcome corresponds to diminished proportion of naïve CD4+ and CD8+ T cells (Supplementary Figure 7). Furthermore, the combination also resulted in increased frequencies of B and NK cells (Supplementary Figure 7).
Meanwhile, in the abscopal TdLNs, t-SNE and quantitative analysis revealed augmented frequencies of activated and central memory CD4+ and CD8+ T cells when anti-CTLA-4 is combined with cryoablation (Figure 6A–C). The combination also increased the level of ICOS expression on those cells, as demonstrated by significant increases in ICOS median fluorescence intensity (MFI) (Figure 6D and E). Therefore, cryoablation plus anti-CTLA-4 leads to higher frequency of activated T cells and central memory T cells proximally and distantly from the primary site of treatment.
Frequency and Activation of Conventional Dendritic Cells in the Tumor-Draining Lymph Nodes are Enhanced by Combination of Cryoablation with Anti-CTLA-4
Cryoablation results in the release of TAAs that can be used by APCs to activate T cells locally or distantly.27 Relative to cryoablation monotherapy, the addition of anti-CTLA-4 resulted in significantly higher frequencies of cDCs at both the primary and abscopal TdLNs, where both type 1 cDCs (cDC1) and type 2 cDCs (cDC2) populations were increased (Figure 7A and B). To understand whether the cDC1s were migrating from the primary site of treatment or were resident cDCs capturing TAAs transported through the lymph, cDC1s were further categorized as resident or migratory based on CD8a+ and CD103+ expression, respectively. Analyses found higher frequencies of resident rather than migratory dendritic cells in both the primary and abscopal TdLNs (Figure 7A and B).
Treatment with cryoablation plus anti-CTLA-4 also led to significant enhancements in the activation of cDCs, as evidenced by higher MHC class II MFI values (Figure 7C and D). This suggests that the higher availability of TAAs and decreased blocks for T cell activation, provided by the combination of cryoablation with anti-CTLA-4, leads to expansion of cDC in the TdLNs. The heightened expression of MHC-II may result in increased antigen presentation activity as a probable mechanism supporting the generation of higher frequencies of effector T cells in the TdLN, which can then migrate into the tumors.
Cryoablation Plus Anti-CTLA-4 Augments Immune Activation Systemically
Besides TdLNs, the spleen is another important secondary lymphoid organ where antigen presentation and T cell activation occurs. The combination of cryoablation with anti-CTLA-4 increased the frequencies of activated CD4+ and CD8+ T cells in the spleen, compared to cryoablation monotherapy (Figure 8A). Similarly to the abscopal TdLNs, a higher percentage of central memory CD4+ T cells was also observed in mice from the combination group (Figure 8A).
Interestingly, the higher percentage of activated T cells was not mirrored by the frequency of cDCs - analyses did not detect any differences in cDC frequencies between the two groups evaluated here (Supplementary Figure 8A). The combination however did lead to a significant increase in the frequencies of myeloid cells, specifically, Ly6G intermediate neutrophils, and similar trends for M1-like TAMs and Ly6G high neutrophils. Hence, the findings suggest that the increased T cell activation is not correlated to increased frequencies of cDC in the spleen, and that intense immune activation might be triggering a balance response operated by myeloid cells.
The increased frequencies of activated CD4+ and CD8+ T cells, observed throughout different organs in response to cryoablation plus anti-CTLA-4, was also observed in the peripheral blood (Figure 8B). Specifically, a higher percentage of effector/effector memory CD4+ and CD8+ T cells were found in animals treated with combination therapy (Figure 8B). There were no noticeable changes in the myeloid compartment besides a significant decrease in the frequency of cDC1s, and a trend of higher percentages of myeloid cells and Ly6G intermediate neutrophils (Supplementary Figure 8B). Peripheral blood is the easiest sample to collect from patients; the results therefore suggest that the effects from the combination therapy might be traceable in the peripheral blood, which could be potentially used in a clinical setting to measure patient responses.
Discussion
Cryoablation is a promising alternative to surgical resection, offering the dual advantage of inducing local tumor necrosis whilst simultaneously stimulating the anti-tumor immune response via release of TAAs and DAMPs. Meanwhile, ICIs have revolutionized cancer treatment due to their ability to release immune cells from inhibitory signals that hinder their activation or effector function. However, both therapies have limitations, especially in TNBC. For example, although cryoablation shows promise in treating early-stage, low-risk breast tumors, there is only limited clinical data on cryoablation use for TNBC. This is because most trials involve older women with small, low-grade lesions and exclude more aggressive subtypes due to larger tumor size at presentation, poorly defined invasive margins, and higher risk of local recurrence, all of which limit the feasibility of complete tumor ablation without surgical resection.46,52 Additionally, ICIs are frequently associated with the development of immune-related adverse events and, as a monotherapy, have demonstrated only modest clinical benefit for TNBC patients in the metastatic setting.17,53 Therefore, combination of cryoablation with ICI is promising, as it would potentiate immune activation, decreasing the likelihood of local recurrence and allowing ICI dosing de-escalation, which could result in lower rates of immune-related adverse events.54–57
Currently, the most common ICIs used in clinical practice are anti-CTLA-4, anti-PD-1, and anti-PD-L1. These antibodies target different phases of the T cell life and, therefore, combination of ICI therapy needs to take into consideration the strategy that will provide the highest synergy. Specifically for TNBC, only anti-PD-1 is FDA-approved in combination with chemotherapy for early-stage high-risk TNBC58 and metastatic PD-L1+ TNBC.59 However, our results suggest that, for the purpose of combination with cryoablation, anti-CTLA-4 is a better candidate than PD-1 or PD-L1 inhibition, leading to higher frequency of activated T cells at the spleen and peripheral blood, with similar trends in abscopal (secondary) tumors.
Cryoablation completely destroys the tumor and the tumor immune microenvironment; this initiates the Cancer-Immunity Cycle, where dendritic cells will uptake TAAs to activate T cells locally, or migrate into the TdLNs or other secondary lymphoid organs to further activate T cells.60 Additionally, antigens can be transported to secondary lymphoid organs through the lymph.61 Thus, it is possible that PD-1/PD-L1 inhibition is not as effective as CTLA-4 due to fewer effector T cells and tumor cells for the antibodies to act upon after cryoablation, while the TAAs and inflammatory signals will boost T cell recruitment and activation, providing an excellent window for CTLA-4 inhibition to take place. This rationale underlies our focus on early immune activation. Therefore, all additional analyses in this study were focused on CTLA-4, to further understand the effects of its combination with cryoablation.
We previously reported that, even though cryoablation completely kills the tumor and associated TIME, infiltration by newly recruited immune cells - especially, naïve CD4+ T cells and myeloid cells - can be detected a week later in the treated tumor.39 Here, we show that the increased immune infiltration in the primary tumor is further stimulated by the addition of anti-CTLA-4, as observed by significant enhancements in the frequency of activated CD8+ T cells, specifically, effector/effector memory CD8+ T cells. This boost in immune infiltration was restricted to the lymphoid compartment, as no significant differences in myeloid frequencies were observed in the primary tumor. This suggests that CTLA-4 blockade was efficient, allowing T cells to be activated by the APCs presenting TAAs preserved by cryoablation.
The increased T cell activation upon anti-CTLA-4 combination with cryoablation was not limited to the primary tumor. Even though TIL scoring did not indicate any differences in the frequencies of lymphocytes between the tested groups, flow cytometry results showed abscopal tumors from cryoablation plus anti-CTLA-4 also harbored increased frequencies of activated CD4+ and CD8+ T cells. Similarly to the primary tumors, no differences were observed in the myeloid compartment of abscopal tumors. While cryoablation alone resulted in trends of enhanced effector/effector memory CD8+ T cell infiltration in abscopal tumors,39 the combination with anti-CTLA-4 led to distinguished increased frequencies of activated and effector/effector memory CD8+ T cells, suggesting that blockade of CTLA-4 further overcomes immunosuppressive signals and improves cryoablation efficacy. The discrepancy between TIL counts and flow cytometry results is not unexpected, as TIL counts - while clinically relevant - measure only the total lymphocyte population without distinguishing subtypes, and are based on a single tumor section rather than the entire tissue.
Lymph nodes are vital components of the immune system, capturing, processing, and presenting tumor antigens.62 Indeed, a systemic immune response was essential for ICI efficacy in a spontaneous breast cancer mouse model study, where lymphocyte egress blockade prevented treatment response.63 The significant increase in the frequency and activation status of cDCs at both primary and abscopal TdLNs indicates that, to some extent, T cells found within the tumor were likely activated in the sentinel lymph nodes and then recruited to the tumor. This agrees with previous studies showing that, upon ICI treatment, TdLNs become immunogenic and act as a source of tumor-reactive T cells,62,64–66 highlighting the importance of this tissue to the anti-tumor immune response.
Besides giving rise to effector/effector memory lymphocytes that will migrate into the tumor, TdLNs also harbor tumor-reactive memory-like CD8+ T cells, which are critical for durable protection post-immunotherapy.67,68 These cells are chemoattracted to lymph nodes and can rapidly differentiate into effector T cells upon inflammatory antigenic re-challenge.69 Interestingly, the combination of CTLA-4 blockade with cryoablation resulted in a significant increase in the frequency of central memory CD8+ T cells at both the primary and abscopal TdLNs. This suggests that combination treatment leads to the establishment of a long-term protective response, where patrolling for recurrence and metastasis is mediated by central memory T cells. However, additional studies evaluating TCR specificity and longevity of the anti-tumor immune response are needed to confirm such hypotheses.
Importantly, lymph node involvement has been linked to distant metastasis, as tumor-draining lymph nodes can be reprogrammed by cancer cells into an immunosuppressive microenvironment that disrupts effective immune response.70–72 Therefore, surgical removal or irradiation of regional lymph nodes remains a standard of care for many cancers, including breast cancer.62 However, the results found here, along with accumulating preclinical and clinical evidence, indicate that the anti-tumor immune response and their enhancement by ICI treatment occur primarily within the TdLN,64,67,73–75 challenging the current standard practice of lymph node removal. In our model, the combination of cryoablation with anti-CTLA-4 not only increased the frequencies of activated T cells, but also augmented their activation status, as shown by elevated ICOS expression (higher MFI).
Despite growing insights into the roles of TdLNs in antitumor immunity, murine models that preserve lymph nodes while incorporating primary tumor treatment remain limited. Most models either fail to develop LN metastases, are constrained by ethical endpoints that preclude late-stage disease, or primarily model metastasis to distant organs, which limits the interpretation of the current data.62 To illustrate, the beneficial role of preserving the TdLN might be dependent on the immunogenicity of the cancer type. For example, a study by Dammeijer et al demonstrated that the nonimmunogenic pancreatic cancer cell KPC3 resulted in TdLN harboring low concentrations of overall activated T cells compared to nondraining lymph nodes. However, when OVA expression was induced in those tumors, which increases tumor immunogenicity, a robust infiltration of tumor specific T cells was observed particularly within the TdLN.75 Thus, the discussion of whether sentinel lymph nodes should be removed must consider the tumor entity, stage, and location,62 and more pre-clinical studies are needed to further guide this decision.
Nevertheless, the combination of cryoablation with anti-CTLA-4 resulted in increased frequency and activation of cDC in the TdLN, potentially being the mechanism leading to increased frequencies of activated T cells locally and systemically. In the spleen, there was a significant increase in the frequencies of activated CD4+ and CD8+ T cells, as well as central memory CD4+ T cells, indicating that this secondary lymphoid organ might also be important for sustained anti-tumor protection. However, further studies are warranted to confirm the functional relevance of the expanded memory compartment. Similarly to the tumors, the DC frequencies in the spleen were also not affected by the addition of anti-CTLA-4 to the treatment.
Interestingly, a significant increase in the frequency of myeloid cells was observed in spleens of mice from the cryoablation plus anti-CTLA-4 group. Among the myeloid populations, the frequency of Ly6G intermediate neutrophils was significantly higher, while similar trends were observed for Ly6G high neutrophils and M1-like macrophages. Ly6G high expression is typical of classical neutrophils, while intermediate Ly6G expression has recently been linked to an immature or transitioning neutrophil population presenting immunosuppressive functions in lung cancer and acute spleen infection studies.76–78 This suggests that these cells are potentially performing regulatory roles, balancing the extensive T cell activation that was observed, however, additional studies are needed to test the suppressive function of these cells.
Superior T cell activation was also observed in the peripheral blood of mice from the combination group, with significant increases in the frequencies of effector/effector memory CD4+ and CD8+ T cells as well. It is possible that this event reflects the migration of T cells from the secondary lymphoid organs, where they are activated, to the tumor, though tracking assays need to be performed to confirm such a fact. Despite additional functional read-outs, these findings are highly relevant as they indicate that response to cryoablation plus anti-CTLA-4 therapy is measurable in the peripheral blood. Blood is the most accessible and minimally invasive source of patient-derived immune cells, so changes in the frequencies of circulating cell populations can serve as early and clinically meaningful indicators of disease-associated immunological shifts.79 It is important to note that peripheral blood immune responses are not always representative of those occurring at the site of the tumor.80 In this study though, the changes observed in the peripheral blood closely mirrored changes observed at the TIME, increasing confidence in the read-out in this scenario. This means that response to the combination therapy could potentially be evaluated in clinical settings by peripheral blood draws, should the use of cryoablation plus anti-CTLA-4 get translated into clinical application.
Our in vivo experiments were performed in two independent replicates with 5 animals per group in each, with highly consistent results across experiments. The effect sizes observed were robust, and similar results were observed when data from both replicates were analyzed independently and when pooled. These reproducible outcomes, coupled with the concordance of immunological readouts obtained by flow cytometry, support the reliability of our findings. Future studies with additional replicates and complementary methodologies will be valuable to further validate and expand upon these results.
The functional activities of both dendritic cells and T cells were directly evaluated in this study. Immune functionality was demonstrated by upregulation of MHC molecules, indicating enhanced antigen-presenting capacity; migration of DCs to draining lymphoid tissues, supporting effective T cell priming; increased ICOS expression as evidence of T cell activation; and differentiation into effector and memory T cell subsets, consistent with the development of sustained antitumor immunity. The study was designed to interrogate early immune event mechanisms following combination treatment with cryoablation and distinct immune checkpoint inhibitors; accordingly, mice were sacrificed one week after treatment for organ collection and analyses. This early time point likely explains why, despite marked immunologic differences among treatment groups, no significant decreases in abscopal tumor weight were detected. The significant enhancements on T cell activation upon combination with anti-CTLA-4, specifically, effector/effector memory CD8+ T cells in the tumors and central memory CD8+ T cells in the lymph nodes, suggest that this treatment setting provides superior results when compared to cryoablation monotherapy. However, additional studies are required to evaluate the antigen specificity of this enlarged T cell population, which will directly reflect on their potential to eliminate tumor cells in the long-term. The high abundance of tumor-associated antigens (TAAs) released and preserved by cryoablation likely suggests that the T cells activated in this context are tumor-specific. If this proves true in future assays, we may be one step closer to combining cryoablation with anti-CTLA-4 therapy for patients with triple-negative breast cancer (TNBC), potentially providing a safer, less invasive treatment that could also reduce immune-related adverse events by enabling lower ICI dosing.
This study has several limitations. First, the analysis was confined to a predefined early 1-week immunological endpoint. Although this time point is biologically appropriate for capturing early immune priming following cryoablation and its potential synergy with checkpoint blockade, it does not permit evaluation of longer-term immune dynamics or the durability of anti-tumor responses. Furthermore, while final tumor weights and representative images enable cross-sectional comparisons between groups (Figure 1C and Supplementary Figure 2B), they do not fully capture treatment kinetics or sustained tumor control over time. Second, immune modulation was assessed primarily at the phenotypic level, focusing on markers associated with activation and anti-tumor potential. Comprehensive functional validation such as quantification of effector cytokine secretion (eg., IFN-γ), direct cytotoxicity assays, and in vivo tumor re-challenge experiments was beyond the scope of the present study. Future investigations will incorporate longitudinal immune profiling, functional T-cell assays, and re-challenge models to evaluate memory responses and durable anti-tumor protection. These approaches will help clarify the mechanistic relationship between early immune activation and sustained therapeutic efficacy, thereby strengthening the translational relevance of this combination strategy.
Conclusions
The combination of cryoablation with ICIs has the potential to overcome the challenges imposed by monotherapy with these approaches, providing higher availability of tumor antigens and inflammation signals, while releasing immune cells from inhibitory signals mediated by checkpoint molecules. In the TNBC mouse model used in this study, combination of cryoablation with anti-CTLA-4 provided superior T cell activation locally and systematically, which was most likely supported by higher frequencies and improved activation status of cDCs in the TdLNs. Future efforts should evaluate the antigen specificity of the expanded T cell population, and the long-term protection provided by the combination to prevent tumor recurrence and metastasis.
Abbreviations
APCs, Antigen-presenting cells; cDCs, Conventional Dendritic Cells; CM, Central memory; Cryo, Cryoablation; CTLA-4, Cytotoxic T-Lymphocyte Antigen 4; CTRL, Control; DAMPS, Damage-associated molecular patterns; E_EM, Effector/Effector Memory; Hematoxylin and Eosin; HER2, Human Epidermal Growth Factor Receptor 2; Hi, high; H&E, IACUC, Institutional Animal Care and Use Committee; ICI, Immune checkpoint inhibitors; ICOS, Inducible T-cell costimulatory; Int, intermediate; IP, intraperitoneal; IVIS, in vivo Imaging System; Ly6G, Lymphocyte antigen 6 complex locus G6D; MFI, Median Fluorescence Intensity; MHC, Major histocompatibility complex; NK, Natural Killer cells; OVA, Ovalbumin; PB, Peripheral Blood; PBS, Phosphate buffered Saline; PD-1, Programmed Cell Death Protein 1; PD-L1, Programmed Death Ligand 1; Pre_eff, Pre effector; RBCs, Red blood cells; TAAs, Tumor-associated antigens; TAMs, Tumor-associated macrophages; TCR, T cell receptor; TdLNs, Tumor-draining lymph nodes; TILs, Tumor-infiltrating lymphocytes; TIME, Tumor immune microenvironment; TNBC, Triple-negative breast cancer; Tregs, Regulatory T cells; t-SNE, t-distributed stochastic neighbor embedding.
Data Sharing Statement
The data that support the findings of this study are available from the corresponding author, MWM, upon reasonable request.
Ethics Statement
All animal procedures were reviewed and approved by the Institutional Animal Care and Use Committee (IACUC) of Texas Tech University Health Sciences Center. The study was conducted in strict accordance with the institutional guidelines, local legislation, and the requirements for the care and use of laboratory animals.
Acknowledgments
We thank, Ice Cure (Leigh Anne Smith and Shad Good) for providing the Prosense Ice Cure Machine through their academic outreach research program, Elizabeth Garcia, LVN and Analicia Ramirez, RN for their expertise in operating the ICE3 machine. TTUHSC Immunology Core Lab for flow cytometry analysis, TTUHSC LARC and staff for assisting with animal care. Figure 1 was created with BioRender.com.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was supported by a research grant from The CH Foundation and from ASCO Equipment endowment to TTUHSC Excellence in Women’s Health.
Disclosure
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
1. Arafat Hossain M. A comprehensive review of immune checkpoint inhibitors for cancer treatment. Int Immunopharmacol. 2024;143(Pt 2):113365. doi:10.1016/j.intimp.2024.113365
2. Rosenberg SA, Yang JC, Restifo NP. Cancer immunotherapy: moving beyond current vaccines. Nat Med. 2004;10(9):909–22. doi:10.1038/nm1100
3. Robert C. A decade of immune-checkpoint inhibitors in cancer therapy. Nat Commun. 2020;11(1):3801. doi:10.1038/s41467-020-17670-y
4. Esfahani K, Roudaia L, Buhlaiga N, Del Rincon SV, Papneja N, Miller WH Jr. A review of cancer immunotherapy: from the past, to the present, to the future. Curr Oncol. 2020;27(Suppl 2):S87–S97. doi:10.3747/co.27.5223
5. Chen L, Flies DB. Molecular mechanisms of T cell co-stimulation and co-inhibition. Nat Rev Immunol. 2013;13(4):227–242. doi:10.1038/nri3405
6. Redmond WL, Ruby CE, Weinberg AD. The role of OX40-mediated co-stimulation in T-cell activation and survival. Crit Rev Immunol. 2009;29(3):187–201. doi:10.1615/critrevimmunol.v29.i3.10
7. Rudd CE, Taylor A, Schneider H. CD28 and CTLA-4 coreceptor expression and signal transduction. Immunol Rev. 2009;229(1):12–26. doi:10.1111/j.1600-065X.2009.00770.x
8. Wing K, Onishi Y, Prieto-Martin P, et al. CTLA-4 control over Foxp3+ regulatory T cell function. Science. 2008;322(5899):271–275. doi:10.1126/science.1160062
9. Agata Y, Kawasaki A, Nishimura H, et al. Expression of the PD-1 antigen on the surface of stimulated mouse T and B lymphocytes. Int Immunol. 1996;8(5):765–772. doi:10.1093/intimm/8.5.765
10. Zhang X, Schwartz JC, Guo X, et al. Structural and functional analysis of the costimulatory receptor programmed death-1. Immunity. 2004;20(3):337–347. doi:10.1016/s1074-7613(04)00051-2
11. Yokosuka T, Takamatsu M, Kobayashi-Imanishi W, Hashimoto-Tane A, Azuma M, Saito T. Programmed cell death 1 forms negative costimulatory microclusters that directly inhibit T cell receptor signaling by recruiting phosphatase SHP2. J Exp Med. 2012;209(6):1201–1217. doi:10.1084/jem.20112741
12. Chemnitz JM, Parry RV, Nichols KE, June CH, Riley JL. SHP-1 and SHP-2 associate with immunoreceptor tyrosine-based switch motif of programmed death 1 upon primary human T cell stimulation, but only receptor ligation prevents T cell activation. J Immunol. 2004;173(2):945–954. doi:10.4049/jimmunol.173.2.945
13. Karwacz K, Bricogne C, MacDonald D, et al. PD-L1 co-stimulation contributes to ligand-induced T cell receptor down-modulation on CD8+ T cells. EMBO Mol Med. 2011;3(10):581–592. doi:10.1002/emmm.201100165
14. Pardoll DM. The blockade of immune checkpoints in cancer immunotherapy. Nat Rev Cancer. 2012;12(4):252–264. doi:10.1038/nrc3239
15. Brahmer JR, Hammers H, Lipson EJ. Nivolumab: targeting PD-1 to bolster antitumor immunity. Future Oncol. 2015;11(9):1307–1326. doi:10.2217/fon.15.52
16. Wojtukiewicz MZ, Rek MM, Karpowicz K, et al. Inhibitors of immune checkpoints-PD-1, PD-L1, CTLA-4-new opportunities for cancer patients and a new challenge for internists and general practitioners. Cancer Metastasis Rev. 2021;40(3):949–982. doi:10.1007/s10555-021-09976-0
17. Schmid P, Rugo HS, Adams S, et al. Atezolizumab plus nab-paclitaxel as first-line treatment for unresectable, locally advanced or metastatic triple-negative breast cancer (IMpassion130): updated efficacy results from a randomised, double-blind, placebo-controlled, phase 3 trial. Lancet Oncol. 2020;21(1):44–59. doi:10.1016/S1470-2045(19)30689-8
18. Shah M, Osgood CL, Amatya AK, et al. FDA approval summary: pembrolizumab for neoadjuvant and adjuvant treatment of patients with high-risk early-stage triple-negative breast cancer. Clin Cancer Res. 2022;28(24):5249–5253. doi:10.1158/1078-0432.CCR-22-1110
19. Bianchini G, Balko JM, Mayer IA, Sanders ME, Gianni L. Triple-negative breast cancer: challenges and opportunities of a heterogeneous disease. Nat Rev Clin Oncol. 2016;13(11):674–690. doi:10.1038/nrclinonc.2016.66
20. Malorni L, Shetty PB, De Angelis C, et al. Clinical and biologic features of triple-negative breast cancers in a large cohort of patients with long-term follow-up. Breast Cancer Res Treat. 2012;136(3):795–804. doi:10.1007/s10549-012-2315-y
21. Lehmann BD, Bauer JA, Chen X, et al. Identification of human triple-negative breast cancer subtypes and preclinical models for selection of targeted therapies. J Clin Invest. 2011;121(7):2750–2767. doi:10.1172/JCI45014
22. Dent R, Trudeau M, Pritchard KI, et al. Triple-negative breast cancer: clinical features and patterns of recurrence. Clin Cancer Res. 2007;13(15 Pt 1):4429–4434. doi:10.1158/1078-0432.CCR-06-3045
23. Wang L, Zhai Q, Lu Q, et al. Clinical genomic profiling to identify actionable alterations for very early relapsed triple-negative breast cancer patients in the Chinese population. Ann Med. 2021;53(1):1358–1369. doi:10.1080/07853890.2021.1966086
24. Bianchini G, De Angelis C, Licata L, Gianni L. Treatment landscape of triple-negative breast cancer - expanded options, evolving needs. Nat Rev Clin Oncol. 2022;19(2):91–113. doi:10.1038/s41571-021-00565-2
25. Chaudhary LN, Wilkinson KH, Kong A. Triple-negative breast cancer: who should receive neoadjuvant chemotherapy? Surg Oncol Clin N Am. 2018;27(1):141–153. doi:10.1016/j.soc.2017.08.004
26. Thomas R, Al-Khadairi G, Decock J. Immune checkpoint inhibitors in triple negative breast cancer treatment: promising future prospects. Front Oncol. 2020;10:600573. doi:10.3389/fonc.2020.600573
27. Abdo J, Cornell DL, Mittal SK, Agrawal DK. Immunotherapy plus cryotherapy: potential augmented abscopal effect for advanced cancers. Front Oncol. 2018;8:85. doi:10.3389/fonc.2018.00085
28. Baust JG, Gage AA. The molecular basis of cryosurgery. BJU Int. 2005;95(9):1187–1191. doi:10.1111/j.1464-410X.2005.05502.x
29. Pusceddu C, Paliogiannis P, Nigri G, Fancellu A. Cryoablation in the management of breast cancer: evidence to date. Breast Cancer. 2019;11:283–292. doi:10.2147/BCTT.S197406
30. Regen-Tuero HC, Ward RC, Sikov WM, Littrup PJ. Cryoablation and immunotherapy for breast cancer: overview and rationale for combined therapy. Radiol Imaging Cancer. 2021;3(2):e200134. doi:10.1148/rycan.2021200134
31. Habrawi Z, Melkus MW, Khan S, et al. Cryoablation: a promising non-operative therapy for low-risk breast cancer. Am J Surg. 2021;221(1):127–133. doi:10.1016/j.amjsurg.2020.07.028
32. Niu L, Wu B, Xu K. Cryosurgery for breast fibroadenomas. Gland Surg. 2012;1(2):128–131. doi:10.3978/j.issn.2227-684X.2012.08.02
33. Kaufman CS, Littrup PJ, Freeman-Gibb LA, et al. Office-based cryoablation of breast fibroadenomas with long-term follow-up. Breast J. 2005;11(5):344–350. doi:10.1111/j.1075-122X.2005.21700.x
34. Littrup PJ, Jallad B, Chandiwala-Mody P, D’Agostini M, Adam BA, Bouwman D. Cryotherapy for breast cancer: a feasibility study without excision. J Vasc Interv Radiol. 2009;20(10):1329–1341. doi:10.1016/j.jvir.2009.06.029
35. Paepke S, Metz S, Brea Salvago A, Ohlinger R. Benign breast tumours - diagnosis and management. Breast Care. 2018;13(6):403–412. doi:10.1159/000495919
36. Khan SY, Melkus MW, Rasha F, et al. Tumor-infiltrating lymphocytes (TILs) as a biomarker of abscopal effect of cryoablation in breast cancer: a pilot study. Ann Surg Oncol. 2022;29(5):2914–2925. doi:10.1245/s10434-021-11157-w
37. Sabel MS, Nehs MA, Su G, Lowler KP, Ferrara JL, Chang AE. Immunologic response to cryoablation of breast cancer. Breast Cancer Res Treat. 2005;90(1):97–104. doi:10.1007/s10549-004-3289-1
38. Wu Y, Cao F, Zhou D, et al. Cryoablation reshapes the immune microenvironment in the distal tumor and enhances the anti-tumor immunity. Front Immunol. 2022;13:930461. doi:10.3389/fimmu.2022.930461
39. de Miranda F S, Martinez-Marin D, Babcock RL, et al. Cryoablation of primary breast cancer tumors induces a systemic abscopal effect altering TIME (Tumor Immune Microenvironment) in distant tumors. Front Immunol. 2024;15:1498942. doi:10.3389/fimmu.2024.1498942
40. Pauken KE, Wherry EJ. Overcoming T cell exhaustion in infection and cancer. Trends Immunol. 2015;36(4):265–276. doi:10.1016/j.it.2015.02.008
41. Wei SC, Duffy CR, Allison JP. Fundamental mechanisms of immune checkpoint blockade therapy. Cancer Discov. 2018;8(9):1069–1086. doi:10.1158/2159-8290.CD-18-0367
42. Farhood B, Najafi M, Mortezaee K. CD8(+) cytotoxic T lymphocytes in cancer immunotherapy: a review. J Cell Physiol. 2019;234(6):8509–8521. doi:10.1002/jcp.27782
43. den Brok MH, Sutmuller RP, Nierkens S, et al. Efficient loading of dendritic cells following cryo and radiofrequency ablation in combination with immune modulation induces anti-tumour immunity. Br J Cancer. 2006;95(7):896–905. doi:10.1038/sj.bjc.6603341
44. Pulendran B, Ahmed R. Immunological mechanisms of vaccination. Nat Immunol. 2011;12(6):509–517. doi:10.1038/ni.2039
45. Sharma P, Allison JP. The future of immune checkpoint therapy. Science. 2015;348(6230):56–61. doi:10.1126/science.aaa8172
46. de Miranda F S, Castro M, Remmert N, Singh SP, Layeequr Rahman R, Melkus MW. Leveraging cryoablation and checkpoint inhibitors for high-risk triple negative breast cancer. Front Immunol. 2023;14:1258873. doi:10.3389/fimmu.2023.1258873
47. Tao K, Fang M, Alroy J, Sahagian GG. Imagable 4T1 model for the study of late stage breast cancer. BMC Cancer. 2008;8:228. doi:10.1186/1471-2407-8-228
48. Redmond WL, Linch SN, Kasiewicz MJ. Combined targeting of costimulatory (OX40) and coinhibitory (CTLA-4) pathways elicits potent effector T cells capable of driving robust antitumor immunity. Cancer Immunol Res. 2014;2(2):142–153. doi:10.1158/2326-6066.CIR-13-0031-T
49. Swancutt KL, Elghonaimy EA, Nicholson JH, et al. Immune stromal components impede biological effectiveness of carbon ion therapy in a preclinical model of pancreatic ductal adenocarcinoma. Mol Cancer Ther. 2026;25(2):333–345. doi:10.1158/1535-7163.MCT-25-0501
50. Kemeny HR, Elsamadicy AA, Farber SH, et al. Targeting PD-L1 initiates effective antitumor immunity in a murine model of Cushing disease. Clin Cancer Res. 2020;26(5):1141–1151. doi:10.1158/1078-0432.CCR-18-3486
51. AVMA. AVMA Guidelines for the Euthanasia of Animals: 2020 Edition. American Veterinary Medical Association; 2020.
52. Fermanian J, Ward RC, Holmes DR, et al. Cryoablation and intratumoral immunotherapy for breast cancer: a future path to cost-effective de-escalation for larger tumors, lymph nodes and metastatic disease. Cancers. 2025;17(12):1915. doi:10.3390/cancers17121915
53. Emens LA, Cruz C, Eder JP, et al. Long-term clinical outcomes and biomarker analyses of atezolizumab therapy for patients with metastatic triple-negative breast cancer: a phase 1 study. JAMA Oncol. 2019;5(1):74–82. doi:10.1001/jamaoncol.2018.4224
54. Low JL, Huang Y, Sooi K, et al. Low-dose pembrolizumab in the treatment of advanced non-small cell lung cancer. Int J Cancer. 2021;149(1):169–176. doi:10.1002/ijc.33534
55. Lebbe C, Meyer N, Mortier L, et al. Evaluation of two dosing regimens for nivolumab in combination with ipilimumab in patients with advanced melanoma: results From the Phase IIIb/IV CheckMate 511 trial. J Clin Oncol. 2019;37(11):867–875. doi:10.1200/JCO.18.01998
56. Rozeman EA, Menzies AM, van Akkooi ACJ, et al. Identification of the optimal combination dosing schedule of neoadjuvant ipilimumab plus nivolumab in macroscopic stage III melanoma (OpACIN-neo): a multicentre, phase 2, randomised, controlled trial. Lancet Oncol. 2019;20(7):948–960. doi:10.1016/S1470-2045(19)30151-2
57. Long GV, Robert C, Butler MO, et al. Standard-dose pembrolizumab plus alternate-dose ipilimumab in advanced melanoma: KEYNOTE-029 Cohort 1C, a Phase 2 randomized study of two dosing schedules. Clin Cancer Res. 2021;27(19):5280–5288. doi:10.1158/1078-0432.CCR-21-0793
58. Schmid P, Cortes J, Dent R, et al. Event-free survival with pembrolizumab in early triple-negative breast cancer. N Engl J Med. 2022;386(6):556–567. doi:10.1056/NEJMoa2112651
59. Cortes J, Cescon DW, Rugo HS, et al. Pembrolizumab plus chemotherapy versus placebo plus chemotherapy for previously untreated locally recurrent inoperable or metastatic triple-negative breast cancer (KEYNOTE-355): a randomised, placebo-controlled, double-blind, phase 3 clinical trial. Lancet. 2020;396(10265):1817–1828. doi:10.1016/S0140-6736(20)32531-9
60. Chen DS, Mellman I. Oncology meets immunology: the cancer-immunity cycle. Immunity. 2013;39(1):1–10. doi:10.1016/j.immuni.2013.07.012
61. Itano AA, McSorley SJ, Reinhardt RL, et al. Distinct dendritic cell populations sequentially present antigen to CD4 T cells and stimulate different aspects of cell-mediated immunity. Immunity. 2003;19(1):47–57. doi:10.1016/s1074-7613(03)00175-4
62. von Renesse J, Lin MC, Ho PC. Tumor-draining lymph nodes - friend or foe during immune checkpoint therapy? Trends Cancer. 2025;11(7):676–690. doi:10.1016/j.trecan.2025.04.008
63. Spitzer MH, Carmi Y, Reticker-Flynn NE, et al. Systemic immunity is required for effective cancer immunotherapy. Cell. 2017;168(3):487–502e15. doi:10.1016/j.cell.2016.12.022
64. Fransen MF, Schoonderwoerd M, Knopf P, et al. Tumor-draining lymph nodes are pivotal in PD-1/PD-L1 checkpoint therapy. JCI Insight. 2018;3(23). doi:10.1172/jci.insight.124507
65. Rahim MK, Okholm TLH, Jones KB, et al. Dynamic CD8(+) T cell responses to cancer immunotherapy in human regional lymph nodes are disrupted in metastatic lymph nodes. Cell. 2023;186(6):1127–1143e18. doi:10.1016/j.cell.2023.02.021
66. Prokhnevska N, Cardenas MA, Valanparambil RM, et al. CD8(+) T cell activation in cancer comprises an initial activation phase in lymph nodes followed by effector differentiation within the tumor. Immunity. 2023;56(1):107–124e5. doi:10.1016/j.immuni.2022.12.002
67. Huang Q, Wu X, Wang Z, et al. The primordial differentiation of tumor-specific memory CD8(+) T cells as bona fide responders to PD-1/PD-L1 blockade in draining lymph nodes. Cell. 2022;185(22):4049–4066e25. doi:10.1016/j.cell.2022.09.020
68. Schenkel JM, Herbst RH, Canner D, et al. Conventional type I dendritic cells maintain a reservoir of proliferative tumor-antigen specific TCF-1(+) CD8(+) T cells in tumor-draining lymph nodes. Immunity. 2021;54(10):2338–2353e6. doi:10.1016/j.immuni.2021.08.026
69. Vokali E, Yu SS, Hirosue S, et al. Lymphatic endothelial cells prime naive CD8(+) T cells into memory cells under steady-state conditions. Nat Commun. 2020;11(1):538. doi:10.1038/s41467-019-14127-9
70. Hoshida T, Isaka N, Hagendoorn J, et al. Imaging steps of lymphatic metastasis reveals that vascular endothelial growth factor-C increases metastasis by increasing delivery of cancer cells to lymph nodes: therapeutic implications. Cancer Res. 2006;66(16):8065–8075. doi:10.1158/0008-5472.CAN-06-1392
71. Burton JB, Priceman SJ, Sung JL, et al. Suppression of prostate cancer nodal and systemic metastasis by blockade of the lymphangiogenic axis. Cancer Res. 2008;68(19):7828–7837. doi:10.1158/0008-5472.CAN-08-1488
72. He Y, Kozaki K, Karpanen T, et al. Suppression of tumor lymphangiogenesis and lymph node metastasis by blocking vascular endothelial growth factor receptor 3 signaling. J Natl Cancer Inst. 2002;94(11):819–825. doi:10.1093/jnci/94.11.819
73. Saddawi-Konefka R, O’Farrell A, Faraji F, et al. Lymphatic-preserving treatment sequencing with immune checkpoint inhibition unleashes cDC1-dependent antitumor immunity in HNSCC. Nat Commun. 2022;13(1):4298. doi:10.1038/s41467-022-31941-w
74. Darragh LB, Gadwa J, Pham TT, et al. Elective nodal irradiation mitigates local and systemic immunity generated by combination radiation and immunotherapy in head and neck tumors. Nat Commun. 2022;13(1):7015. doi:10.1038/s41467-022-34676-w
75. Dammeijer F, van Gulijk M, Mulder EE, et al. The PD-1/PD-L1-checkpoint restrains T cell immunity in tumor-draining lymph nodes. Cancer Cell. 2020;38(5):685–700e8. doi:10.1016/j.ccell.2020.09.001
76. Pfirschke C, Engblom C, Gungabeesoon J, et al. Tumor-promoting Ly-6G(+) SiglecF(high) cells are mature and long-lived neutrophils. Cell Rep. 2020;32(12):108164. doi:10.1016/j.celrep.2020.108164
77. Liao C, Luo S, Liu X, et al. Siglec-F(+) neutrophils in the spleen induce immunosuppression following acute infection. Theranostics. 2024;14(6):2589–2604. doi:10.7150/thno.93812
78. Deniset JF, Surewaard BG, Lee WY, Kubes P. Splenic Ly6G(high) mature and Ly6G(int) immature neutrophils contribute to eradication of S. pneumoniae. J Exp Med. 2017;214(5):1333–1350. doi:10.1084/jem.20161621
79. Martins I, Ribeiro IP, Jorge J, et al. Liquid biopsies: applications for cancer diagnosis and monitoring. Genes. 2021;12(3):349. doi:10.3390/genes12030349
80. Malyguine AM, Strobl SL, Shurin MR. Immunological monitoring of the tumor immunoenvironment for clinical trials. Cancer Immunol Immunother. 2012;61(2):239–247. doi:10.1007/s00262-011-1148-6
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.