Back to Journals » Journal of Inflammation Research » Volume 15
Comparison of the Concentration of Encephalitozoon cuniculi Genotypes I and III in Inflammatory Foci Under Experimental Conditions
Authors Sak B, Holubová N ![]() , Květoňová D, Hlásková L, Tinavská J, Kicia M, Zajączkowska Ż, Kváč M
, Květoňová D, Hlásková L, Tinavská J, Kicia M, Zajączkowska Ż, Kváč M
Received 22 February 2022
Accepted for publication 12 April 2022
Published 26 April 2022 Volume 2022:15 Pages 2721—2730
DOI https://doi.org/10.2147/JIR.S363509
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Ning Quan
Bohumil Sak,1 Nikola Holubová,1 Dana Květoňová,1 Lenka Hlásková,1 Jana Tinavská,1,2 Marta Kicia,3 Żaneta Zajączkowska,3 Martin Kváč1,4
1Institute of Parasitology, Biology Centre, Czech Academy of Science, České Budějovice, Czech Republic; 2Faculty of Science, University of South Bohemia in České Budějovice, České Budějovice, Czech Republic; 3Department of Biology and Medical Parasitology, Wroclaw Medical University, Wroclaw, Poland; 4Faculty of Agriculture, University of South Bohemia in České Budějovice, České Budějovice, Czech Republic
Correspondence: Bohumil Sak, Institute of Parasitology, Biology Centre, Czech Academy of Science, České Budějovice, Czech Republic, Tel +420387775421, Fax +420385310388, Email [email protected]
Background: Microsporidia of the genus Encephalitozoon are usually associated with severe infections in immunodeficient hosts while, in immunocompetent ones, microsporidiosis produces minimal clinically apparent disease. Despite their microscopic size, microsporidia are capable of causing systemic infection within a few days. However, the mechanisms by which microsporidia reach target tissues during acute infection remain unclear. Out of four genotypes of Encephalitozoon cuniculi, only three are available for experimental studies, with E. cuniculi genotype II being the best characterized.
Methods: In the present study, we tested the association between inflammation induction in immunocompetent and immunodeficient mice and the presence of spores of E. cuniculi genotypes I and III in selected organs using molecular methods and compared the results with previously published data on E. cuniculi genotype II.
Results: We reported the positive connection between inflammation induction and the significant increase of E. cuniculi genotypes I and III occurrence in inflammatory foci in both immunocompetent BALB/c and immunodeficient severe combined immunodeficient (SCID) mice in the acute phase of infection. The induction of inflammation resulted in increased concentration of E. cuniculi of both genotypes in the site of inflammation, as previously reported for E. cuniculi genotype II. Moreover, our study extended the spectrum of differences among E. cuniculi genotypes by the variations in dispersal rate within host bodies after experimentally induced inflammation.
Conclusion: The results imply possible involvement of immune cells serving as vehicles transporting E. cuniculi towards inflammation foci. The elucidation of possible connection with pro-inflammatory immune responses represents an important challenge with implications for human health and the development of therapeutic strategies.
Keywords: Encephalitozoon cuniculi genotype I, Encephalitozoon cuniculi genotype III, inflammation, targeted migration
Introduction
Microsporidia are unicellular, intracellular, spore-forming, ubiquitous parasites belonging to the phylum Microsporidia found to infect a wide range of hosts, including humans. The best described microsporidian is Encephalitozoon cuniculi, which infect a wide range of vertebrates and invertebrates.1 Currently, four genotypes of E. cuniculi have been determined based on molecular differences in the internal transcribed spacer (ITS) region of the rRNA genes.2,3 While E. cuniculi genotype I has rarely been recorded in asymptomatic birds, rodents, rabbits, dogs, horses, boars, buffalos, great apes, and seldom in humans4–18 and E. cuniculi genotype III has been identified only in lemmings, rabbits, birds, swine, dogs, blue foxes, snow leopard, tamarins and squirrel monkeys, and also in humans, causing a variety of pathologies ranging from asymptomatic to fatal infections,4,19–34 Encephalitozoon cuniculi genotype II has been found in many different species of hosts causing a wide spectrum of pathogenesis.35–40 The recently described genotype IV has been detected in cats, dogs, marmots, and in immunosuppressed humans,3,41,42 causing disseminated encephalitozoonosis in the latter species. However, contrary to the first three genotypes, genotype IV is not available for in vitro culture and thus for laboratory experiments.
Encephalitozoon species infect several cell types in mammalian hosts, including epithelial and endothelial cells, fibroblasts, macrophages, and astrocytes43 mainly in the gastrointestinal tract, kidney, brain, eyes, sinuses, and muscles.35,36,44–46 Recently, a possible involvement of immune cells connected with pro-inflammatory immune responses serving as vehicles transporting E. cuniculi genotype II purposefully across the whole host body towards inflammation was suggested, based on the occurrence of microsporidia in inflamed tissues,47 and experimentally verified.48
The present study was designed to compare the progress of concentration of Encephalitozoon cuniculi genotypes I and III in sites of inflammation with previously published data on E. cuniculi genotype II48 and to answer the question of whether individual genotypes of E. cuniculi differ in dispersal within the host body following experimentally induced inflammation.
Materials and Methods
Ethics Statement
All experimental procedures complied with the law of the Czech Republic (Act No 246/1992 Coll., on the protection of animals against cruelty). The study design was approved by ethical committees at the Biology Centre of CAS, the State Veterinary Administration, and the Central Commission for Animal Welfare under protocol no. 35/2020.
Mice
Eight-week-old Severe Combined ImmunoDeficient (SCID) and immunocompetent (BALB/c) mice originally obtained from Charles River (Sulzfeld, Germany) were bred in plastic cages with sterilized wood-chip bedding situated in flexible film isolators (BEM Znojmo, Czech Republic) with HEPA filters. Mice were supplied with a sterilized diet (TOP-VELAZ Praha, Czech Republic) and sterilized water ad libitum.
Parasites
The spores of E. cuniculi genotype I (lagomorph subtype, ATCC®-50503TM) originally isolated from rabbit, OH, USA in 1978 (ATCC® Product Sheet), and spores of E. cuniculi genotype III originally isolated from pet rodents, Steppe lemmings (Lagurus lagurus), CZE in 201432 were grown in vitro in Green monkey kidney cells (VERO, line E6) maintained in RPMI-1640 medium (Sigma-Aldrich, St. Louis, MO) supplemented with 2.5% heat inactivated fetal bovine serum. Spores were harvested and purified from cells by centrifugation over 50% Percoll (Sigma-Aldrich) at 1100×g for 30 min and washed three times in deionized water before storing in deionized water at 4°C supplemented with antibiotics (Sigma-Aldrich, 100 U/mL penicillin, 100 μg/mL streptomycin, and 2.5 μg/mL amphotericin B). The spores were washed with deionized water before use.
Experimental Protocol
Fifteen SCID and 27 BALB/c mice were per orally (P.O.) infected with the dose of 107 either E. cuniculi genotype I or III spores in 0.2 mL of deionized water using intragastric gavage. The inflammation in experimental SCID and BALB/c was induced by the usage of 50 μL of Freund’s Incomplete Adjuvant (Sigma-Aldrich) inoculated in the acute phase of infection49 to the hind limb muscle. Mice of both strains injected intramuscularly with 50 μL of sterile phosphate buffered solution (PBS) at the same intervals were used as negative controls. The animal’s health, mortality, and morbidity were recorded at 12-hour intervals.
Three mice from each group were euthanized by cervical dislocation every seventh day post-infection (DPI), and sterile samples for molecular detection and histology were prepared from limb muscles and liver by the usage of a different pair of sterile dissection tools.
DNA Isolation
Tissue samples were homogenized by bead disruption using a FastPrep®–24 Instrument (MP Biomedicals, CA) and 0.5 mm glass beads (Biospec Products, Inc., Bartlesville, OK) at a speed of 5.5 m/s for 1 min. Total DNA was extracted using a commercially available column based isolation kit, DNeasy Blood & Tissue Kit (QIAGEN, Hilden, Germany), according to the manufacturer’s instructions. Extracted DNA was stored at −20°C.
PCR Amplification
Nested PCR protocols were used for amplification of a partial sequence of 16S rRNA, using microsporidia-specific primers.50,51 DNA obtained from spores of E. cuniculi genotype II and ultrapure water was used as a positive and negative control, respectively. PCR products were evaluated by gel electrophoresis.
qRT-PCR
DNA samples from limb muscles and liver were processed using a quantitative RT-PCR protocol amplifying a 268-bp region of the 16S rRNA gene of E. cuniculi52 and a 137-bp region of murine β-actin as a housekeeping gene.53 Each run included negative controls consisting of unspiked specimens and diluent blanks. The total amount of spores in 1 g of individual tissue samples was calculated based on the number of β-actin copies in the tissue sample based on a method routinely established in the laboratory.54,55
Statistical Analysis
Differences in microsporidia presence in inflammatory foci compared to the non-induced site were analyzed by Student’s t-tests and the conformity of the variances of the tested groups was verified by an F-test at a confidence level of α≤0.05. All computations were made using Statistica 6.0 software (StatSoft CR, Praha, Czech Republic).
Results
The application of Freund’s Incomplete Adjuvant elicited a local inflammatory immune response in all animals characterized by the formation of a purulent inflammatory focus in the hind limb muscle. On the other hand, no macroscopic changes were observed in the PBS-inoculated mice.
Induction of Inflammation in SCID Mice
In immunodeficient SCID mice, E. cuniculi genotypes I and III were detected in all screened tissue already prior to induction of inflammation (Figure 1A and B) with a higher spore burden in E. cuniculi genotype III infected mice, reaching up to 5.2×102 spores per gram of tissue.
 |
Figure 1 Maximum value of E. cuniculi spores per 1 gram of tissue (number in white fields), frequency and spore burden of E. cuniculi spores per gram of tissue (colored squares; each square represents one mouse), and infection trend illustration (red fields) in immunodeficient SCID mice infected either with E. cuniculi genotype I (A) or E. cuniculi genotype III (B) and induced in the acute phase of infection. aPeroral infection 107 spores of E. cuniculi in 200 µL dH2O and intramuscular injection of PBS; bperoral infection 107 spores of E. cuniculi in 200 µL dH2O and intramuscular injection of Freund’s Incomplete Adjuvant; grey column, intramuscular injection of all animals in the group into the right thigh muscle; *significant difference between right and left leg; #significant difference between control and experimental group; colored squares, positive capture of microsporidia in the sample according to the quantity scale. |
In the control group of immunodeficient SCID mice infected with E. cuniculi genotype I (Figure 1A) and with no inflammation induction (injected with PBS only), no effect was observed and the number of spores increased continuously till the end of experiment 35 days post-infection (DPI), reaching up to 1.7×104 spores per gram of tissue (Figure 1A).
After induction of inflammation of 7 DPI in the experimental group, in the non-induced leg, the spore burden was similar to the control group; however, a significant (p<0.05) increase in the number of spores in the induced right hind leg was observed, starting at 14 DPI and reaching up to 2.4×105 spores per gram of tissue at 35 DPI, compared to the left hind leg, containing maximally 3.1×104 spores per gram of tissue at 35 DPI. In addition, there was a significant difference (p<0.05) in spore burden in induced animals compared to the control mice, and the infection trend was more intense in induced animals (Figure 1A).
A similar situation was observed in SCID mice infected with E. cuniculi genotype III. There was no difference within control groups (Figure 1B), except that the spore burden in individual tissues was higher compared to E. cuniculi genotype I, reaching up to 4.7×106 spores per gram of tissue at 35 DPI.
In the experimental group infected with E. cuniculi genotype III with induced inflammation, there was a one order shift toward the induced site starting at 14 DPI reaching 2.7×102, 2.1×104, 2.1×106, and 1.0×107 spores per gram of tissue, compared to 1.5×101, 2.2×103, 1.5×105, and 1.3×106 spores per gram of tissue in the control group (Figure 1B). Surprisingly, a significant decrease in spore burden was observed in the non-induced site at 14 DPI (eg, 1.5×106 spores per gram of tissue). Moreover, there was a significant difference (p<0.05) in spore burden in induced animals compared to the control mice starting from 21 DPI (Figure 1B). The infection trend was significantly more intense and reached a higher spore burden in the induced site compared to SCID mice infected with E. cuniculi genotype I (Figure 1A).
Induction of Inflammation in BALB/c Mice
Also in immunocompetent BALB/c mice were E. cuniculi genotypes I and III, detected prior to induction of inflammation in all screened tissue samples at least in one animal per group (Figure 2A and B).
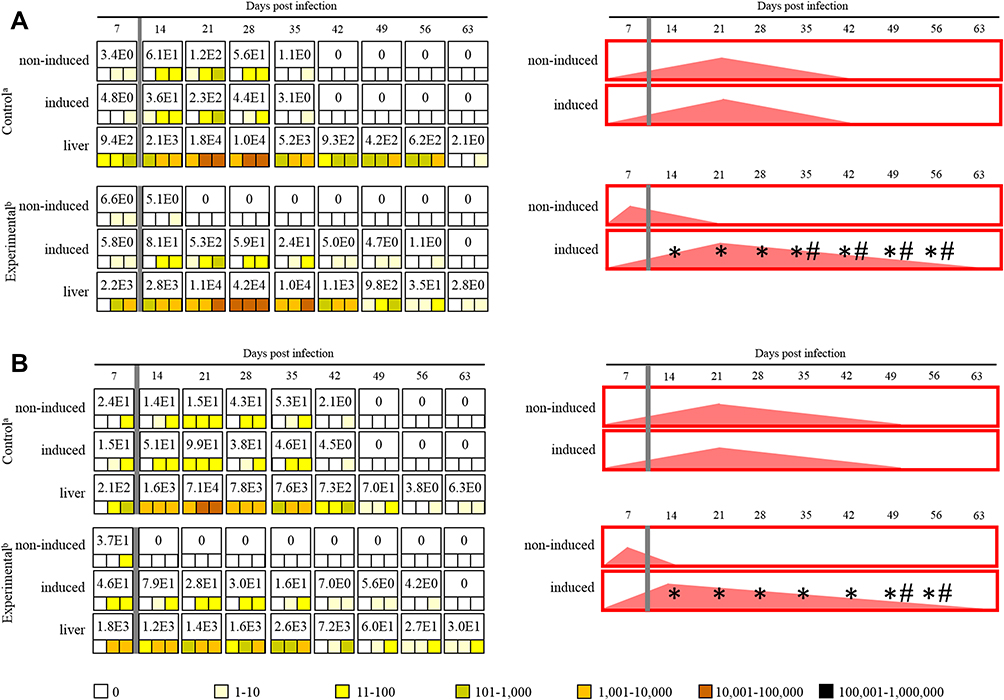 |
Figure 2 Maximum value of E. cuniculi spores per 1 gram of tissue (number in white fields), frequency and spore burden of E. cuniculi spores per gram of tissue (colored squares; each square represents one mouse), and infection trend illustration (red fields) in immunocompetent BALB/c mice infected either with E. cuniculi genotype I (A) or E. cuniculi genotype III (B) and induced in the acute phase of infection. aPeroral infection 107 spores of E. cuniculi in 200 µL dH2O and intramuscular injection of PBS; bperoral infection 107 spores of E. cuniculi in 200 µL dH2O and intramuscular injection of Freund’s Incomplete Adjuvant; grey column, intramuscular injection of all animals in the group into the right thigh muscle; *significant difference between right and left leg; #significant difference between control and experimental group; colored squares, positive capture of microsporidia in the sample according to the quantity scale. |
In the control group of immunocompetent BALB/c mice infected with E. cuniculi genotypes I and III, no differences in microsporidia distribution were observed and the number of spores was comparable in both legs, increased at the beginning of the infection, reaching up to 2.3×102 and 9.9×101 spores per 1 g of tissue, respectively, and followed by continuous disappearance and clearance of microsporidia in latter weeks of infection from muscles, namely 42 DPI (E. cuniculi genotype I) and 49 DPI (E. cuniculi genotype III) (Figure 2A and B).
The induction of inflammation in immunocompetent BALB/c mice infected with E. cuniculi genotype I during the acute phase of infection (7 DPI) resulted in the disappearance of microsporidia from the non-induced left hind leg and an increase of their concentration toward the induced right hind leg in all experimental mice (p<0.05) starting from 14 DPI (Figure 2A), reaching up to 5.3×102 spores per 1 g of tissue reported 2 weeks post-inflammation induction. Moreover, the inflammation induction resulted in a significant increase (p<0.05) in concentration of spores in the induced site in the experimental group 35 DPI and prolonged persistence of microsporidia till 56 DPI in the induced leg compared to the control group (Figure 2A).
One order shift towards the induced site was also reported in immunocompetent BALB/c mice infected with E. cuniculi genotype III with induced inflammation, starting at 14 DPI and reaching the maximum of 7.9×101 spores per gram of tissue at 14 DPI. The persistence of microsporidia was prolonged for 2 weeks in the induced right hind leg compared to the control group (Figure 2B).
Discussion
Of the several species of microsporidia that infect humans, E. cuniculi belongs to the most common.39 Horizontally transmitted environmentally resistant life cycle stage spores access host tissues by germinating in the gut lumen, where infections are typically initiated in gut epithelial cells, and sometimes muscle cells. However, the target tissues – those in which microsporidia develop to the infectious stage, represent a species-specific interaction with its host. As E. cuniculi lacks the organ/cell specificity, it can infect a wide spectrum of host cells including epithelial cells, vascular endothelial cells, kidney tubule cells, and possibly other cell types, and can be found in most tissues, with a predilection, in mammals, for the brain and kidney.49,55–58 However, microsporidian spores are non-motile and short distance host dissemination is limited to a unique mechanism of host cell invasion using 10–50 μm long polar tubes that deliver this organism into the host cell. As Encephalitozoon species are able to survive and replicate in a variety of immune cells, including resident and migratory macrophages and other phagocytic cells such as neutrophils, and eosinophils, monocytes, and dendritic cells, which may contribute to the dissemination of E. cuniculi throughout the host organism,59,60 suggesting that induction of chemokines for inducing innate immune inflammation may also promote recruitment of host cells for continued infection and dissemination.61–63 Recently, a role for the involvement of such immune cells connected with pro-inflammatory immune responses (eg, trafficking macrophages and other phagocytic cells such as neutrophils, monocytes, dendritic cells, and eosinophils) in the expansion of E. cuniculi infection in host tissues has been suggested based on the occurrence of microsporidia in inflamed tissues47 and the targeted migration seen following experimental induction of inflammation towards inflammation foci.48
Although four genotypes of E. cuniculi have been identified to date, human infections are mostly associated with E. cuniculi genotypes I and II,64 which are found primarily in HIV-infected patients,65–67 and in transplant recipients.3,31,68,69
In animals, E. cuniculi genotype III seems to be more aggressive and infections caused by this strain can lead to the death of immunocompetent hosts,28,32,33 whereas E. cuniculi genotypes I and II mostly represent a parasite that occurs in host species in a latent asymptomatic form.4,6–8,10–12,17,70
As the particular genotypes of E. cuniculi differ not only in infection onset speed and extent of infection, but also in chemotherapy sensitivity,49,55,57,58,71,72 thus host survival and progression of the infection depend not only on the immune status of the host but also on the E. cuniculi genotype causing the infection. Under experimental conditions, the infection extent and albendazole sensitivity of E. cuniculi genotype I was comparable to E. cuniculi genotype II and the infection onset speed and mortality rate were similar to E. cuniculi genotype III.72
As the results of the present study show, the migration potential of E. cuniculi genotypes and their concentration in induced inflammatory foci varied in both immunocompetent and immunodeficient mice.
In SCID mice the concentration of microsporidia was the most intense in E. cuniculi genotype III, reaching 1.0×107 spores per gram of tissue at 35 DPI, compared to 2.4×105 and 5.0×105 reported in genotypes I and II, respectively (present study),48 (Figure 3A). This correspond to the data published by Sak et al55,58,72 and Kotková et al,49,71 ie, the infection intensity of E. cuniculi genotypes I and II was comparable, whereas E. cuniculi genotype III reached up to two orders higher spore burden.
 |
Figure 3 Maximum value of E. cuniculi spores per 1 gram of tissue (number in white fields), frequency and spore burden of E. cuniculi spores per gram of tissue (colored squares; each square represents one mouse), and infection trend illustration (red fields) in immunodeficient SCID mice (A) and immunocompetent BALB/c mice (B) infected with E. cuniculi genotype II (modified from Brdíčková et al48). aPeroral infection 107 spores of E. cuniculi in 200 µL dH2O and intramuscular injection of PBS; bperoral infection 107 spores of E. cuniculi in 200 µL dH2O and intramuscular injection of Freund’s Incomplete Adjuvant; grey column, intramuscular injection of all animals in the group into the right thigh muscle; *significant difference between right and left leg; #significant difference between control and experimental group; colored squares, positive capture of microsporidia in the sample according to the quantity scale. |
Induction of inflammation in the experimental group of SCID mice infected with E. cuniculi genotype I or III resulted in a significant increase of spore burden at the site of inflammation, while, in the control group, the amount of spores per gram of tissue was approximately identical in both legs. Moreover, the significant concentration of microsporidia in the inflammation-induced site was comparable in the case of E. cuniculi genotypes I and III and was reported from 14 DPI, whereas the only significant difference between induced and non-induced site was described in SCID mice infected E. cuniculi genotype II at 21 DPI48 (Figure 3A). The elevated concentration of spores in muscle tissues at the end of the experiment regardless of inflammation induction could be attributed to the systemic spread of E. cuniculi in the terminal phase infection, as described already by Kotková et al.49
Also, in control immunocompetent BALB/c mice, the course of infection caused by E. cuniculi I was similar to E. cuniculi genotype II with a random occurrence of microsporidia in the limbs and infection clearance starting from 42 DPI, compared to E. cuniculi genotype III with the infection clearance at 49 DPI (present study),48 (Figure 3B), suggesting that, during the experiment, microsporidia may have been eliminated by the host immune system or microsporidia remaining in the body in undetectable amounts. The peak of infection in muscles was reported earlier in E. cuniculi genotypes II and III (21 DPI) than in infection with E. cuniculi genotype II (28 DPI) (present study),48 (Figure 3B).
The induction of inflammation in experimental BALB/c mice led to the disappearance of all genotypes of E. cuniculi from the non-induced leg starting either from 14 DPI (genotypes II and III) or 21 DPI (genotype I) (present study),48 (Figure 3B). Moreover, a significant increase of microsporidia towards the sites of inflammation in the induced leg was observed from 14 DPI in all genotypes, reaching a peak of E. cuniculi concentration either at 14 DPI (genotype III) or 21 DPI (genotypes I and III). On the other hand, no difference was observed between the occurrence frequencies and spore burden of microsporidia in induced and non-induced legs in the particular control groups (present study),48 (Figure 3B).
Conclusions
Based on our results we can conclude that the induction of inflammation had caused increased concentrations of E. cuniculi genotypes I and III in inflammatory foci, as previously reported for E. cuniculi genotype II. The frequency of microsporidia occurrence was not random, but shifted in favor of the site of induced inflammation compared to control groups, where spores were accidental or their specific DNA was not detected at all.
Moreover, our study confirmed differences among E. cuniculi genotypes reported previously based on the infection onset, infection extent, and host mortality rate. Based on the present study, we can conclude that particular genotypes of E. cuniculi also differ in dispersal rate within host bodies after experimentally induced inflammation and E. cuniculi genotype I possesses a combination of features reported in genotypes II and III.
As the mechanisms of microsporidia dispersal as well as the potential vehicle cells remain speculative, detailed specification is needed. The understanding of the connection of microsporidia dispersal with pro-inflammatory immune responses, addressing the tissue specificity and the mechanisms of transfer to target tissues, represents an important challenge with potential consequences for human health and brings novel insight into microsporidia problems and developing reliable preventive and therapeutic strategies.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising, or critically reviewing the article; gave final approval of the version to be published; and have agreed on the journal to which the article has been submitted.
Funding
The study was supported by grants from the Grant Agency of the Czech Republic (20-10706S) and National Science Centre, Poland (2020/39/O/NZ6/02313).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Canning EU, Lom J, Dyková I. The Microsporidia of Vertebrates. London: Academic Press; 1986:289.
2. Didier ES, Vossbrinck CR, Baker MD, et al. Identification and characterization of three Encephalitozoon cuniculi strains. Parasitology. 1995;111:411–421. doi:10.1017/S0031182000065914
3. Talabani H, Sarfati C, Pillebout E, et al. Disseminated infection with a new genovar of Encephalitozoon cuniculi in a renal transplant recipient. J Clin Microbiol. 2010;48:2651–2653. doi:10.1128/JCM.02539-09
4. Kašičková D, Sak B, Kváč M, et al. Sources of potentially infectious human microsporidia: molecular characterisation of microsporidia isolates from exotic birds in the Czech Republic, prevalence study and importance of birds in epidemiology of the human microsporidial infections. Vet Parasitol. 2009;165:125–130. doi:10.1016/j.vetpar.2009.06.033
5. Engelhardt S, Buder A, Pfeil K, et al. Nephritis in a Staffordshire Terrier puppy caused by Encephalitozoon cuniculi genotype I. Tierarztl Prax Ausg K Kleintiere Heimtiere. 2017;45(6). doi:10.15654/TPK-161123
6. Sak B, Kváč M, Petrželková K, et al. Diversity of microsporidia (Fungi: microsporidia) among captive great apes in European zoos and African sanctuaries: evidence for zoonotic transmission? Folia Parasitol. 2011;58:81–86. doi:10.14411/fp.2011.008
7. Sak B, Petrželková KJ, Květoňová D, et al. Long-term monitoring of microsporidia, Cryptosporidium and Giardia infections in western Lowland Gorillas (Gorilla gorilla gorilla) at different stages of habituation in Dzanga Sangha Protected Areas, Central African Republic. PLoS One. 2013;8:e71840. doi:10.1371/journal.pone.0071840
8. Sak B, Petrželková KJ, Květoňová D, et al. Diversity of microsporidia, Cryptosporidium and Giardia in mountain gorillas (Gorilla beringei beringei) in Volcanoes National Park, Rwanda. PLoS One. 2014;9:e109751. doi:10.1371/journal.pone.0109751
9. Sak B, Kváč M, Květoňová D, et al. The first report on natural Enterocytozoon bieneusi and Encephalitozoon spp. infections in wild East-European House Mice (Mus musculus musculus) and West-European House Mice (M. m. domesticus) in a hybrid zone across the Czech Republic–Germany border. Vet Parasitol. 2011;178:246–250. doi:10.1016/j.vetpar.2010.12.044
10. Tsukada R, Tsuchiyama A, Sasaki M, et al. Encephalitozoon infections in Rodentia and Soricomorpha in Japan. Vet Parasitol. 2013;198:193–196. doi:10.1016/j.vetpar.2013.08.018
11. Wagnerová P, Sak B, Květoňová D, et al. Enterocytozoon bieneusi and Encephalitozoon cuniculi in horses kept under different management systems in the Czech Republic. Vet Parasitol. 2012;190:573–577. doi:10.1016/j.vetpar.2012.07.013
12. Laatamna A, Wagnerová P, Sak B, et al. Microsporidia and Cryptosporidium in horses and donkeys in Algeria: detection of a novel Cryptosporidium hominis subtype family (Ik) in a horse. Vet Parasitol. 2015;208:135–142. doi:10.1016/j.vetpar.2015.01.007
13. Kimura M, Aoki M, Ichikawa-Seki M, et al. Detection and genotype of Encephalitozoon cuniculi DNA from urine and feces of pet rabbits in Japan. J Vet Med Sci. 2013;75:1017–1020. doi:10.1292/jvms.12-0567
14. Deng L, Chai Y, Xiang L, et al. First identification and genotyping of Enterocytozoon bieneusi and Encephalitozoon spp. in pet rabbits in China. BMC Vet Res. 2020;16:212. doi:10.1186/s12917-020-02434-z
15. Němejc K, Sak B, Květoňová D, et al. Prevalence and diversity of Encephalitozoon spp. and Enterocytozoon bieneusi in wild boars (Sus scrofa) in Central Europe. Parasitol Res. 2014;113:761–767. doi:10.1007/s00436-013-3707-6
16. Ditrich O, Chrdle A, Sak B, et al. Encephalitozoon cuniculi genotype I as a causative agent of brain abscess in an immunocompetent patient. J Clin Microbiol. 2011;49:2769–2771. doi:10.1128/JCM.00620-11
17. Halánová M, Valenčáková A, Malčeková B, et al. Occurrence of microsporidia as emerging pathogens in Slovak Roma children and their impact on public health. Ann Agric Environ Med. 2013;20:695–698.
18. Tavalla M, Mardani-Kateki M, Abdizadeh R, et al. Molecular identification of Enterocytozoon bieneusi and Encephalitozoon spp. in immunodeficient patients in Ahvaz, Southwest of Iran. Acta Trop. 2017;172:107–112. doi:10.1016/j.actatropica.2017.04.015
19. Botha WS, Dormehl IC, Goosen DJ. Evaluation of kidney function in dogs suffering from canine encephalitozoonosis by standard clinical pathological and radiopharmaceutical techniques. J S Afr Vet Assoc. 1986;57:79–86.
20. Cutlip RC, Beall CW. Encephalitozoonosis in Arctic lemmings. Lab Anim Sci. 1989;39:331–333.
21. van Dellen AF, Stewart CG, Botha WS. Studies of encephalitozoonosis in vervet monkeys (Cercopithecus pygerythrus) orally inoculated with spores of Encephalitozoon cuniculi isolated from dogs (Canis familiaris). Onderstepoort J Vet Res. 1989;56:1–22.
22. Didier ES, Varner PW, Didier PJ, et al. Experimental microsporidiosis in immunocompetent and immunodeficient mice and monkeys. Folia Parasitol. 1994;41:1–11.
23. Guscetti F, Mathis A, Hatt JM, et al. Overt fatal and chronic subclinical Encephalitozoon cuniculi microsporidiosis in a colony of captive emperor tamarins (Saguinus imperator). J Med Primatol. 2003;32:111–119. doi:10.1034/j.1600-0684.2003.00016.x
24. Reetz J, Wiedemann M, Aue A, et al. Disseminated lethal Encephalitozoon cuniculi (genotype III) infections in cotton-top tamarins (Oedipomidas oedipus)-a case report. Parasitol Int. 2004;53:29–34. doi:10.1016/j.parint.2003.10.003
25. Reetz J, Nöckler K, Reckinger S, et al. Identification of Encephalitozoon cuniculi genotype III and two novel genotypes of Enterocytozoon bieneusi in swine. Parasitol Int. 2009;58:285–292. doi:10.1016/j.parint.2009.03.002
26. Mathis A, Weber R, Deplazes P. Zoonotic potential of the microsporidia. Clin Microbiol Rev. 2005;18:423–445. doi:10.1128/CMR.18.3.423-445.2005
27. Asakura T, Nakamura S, Ohta M, et al. Genetically unique microsporidian Encephalitozoon cuniculi strain type III isolated from squirrel monkeys. Parasitol Int. 2006;55:159–162. doi:10.1016/j.parint.2006.02.002
28. Juan-Sallés C, Garner MM, Didier ES, et al. Disseminated encephalitozoonosis in captive, juvenile, cotton-top (Saguinus oedipus) and neonatal emperor (Saguinus imperator) tamarins in North America. Vet Pathol. 2006;43:438–446. doi:10.1354/vp.43-4-438
29. Snowden KF, Lewis BC, Hoffman J, et al. Encephalitozoon cuniculi infections in dogs: a case series. J Am Anim Hosp Assoc. 2009;45:225–231. doi:10.5326/0450225
30. Valencakova A, Balent P, Ravaszova P, et al. Molecular identification and genotyping of Microsporidia in selected hosts. Parasitol Res. 2011;110:689–693. doi:10.1007/s00436-011-2543-9
31. Hocevar SN, Paddock CD, Spak CW, et al. Microsporidiosis acquired through solid organ transplantation: a public health investigation. Ann Intern Med. 2014;160:213–220. doi:10.7326/M13-2226
32. Hofmannová L, Sak B, Jekl V, et al. Lethal Encephalitozoon cuniculi genotype III infection in Steppe lemmings (Lagurus lagurus). Vet Parasitol. 2014;205:357–360. doi:10.1016/j.vetpar.2014.07.008
33. Meng X, Zheng J, He X, et al. First characterization in China of Encephalitozoon cuniculi in the blue fox (Alopex lagopus). J Eukaryot Microbiol. 2014;61:580–585. doi:10.1111/jeu.12135
34. Scurrell EJ, Holding E, Hopper J, et al. Bilateral lenticular Encephalitozoon cuniculi infection in a snow leopard (Panthera uncia). Vet Ophthalmol. 2015;18:143–147. doi:10.1111/vop.12218
35. Weber R, Bryan RT. Microsporidial infections in immunodeficient and immunocompetent patients. Clin Infect Dis. 1994;19:517–521. doi:10.1093/clinids/19.3.517
36. Weber R, Bryan RT, Schwartz DA, et al. Human microsporidial infections. Clin Microbiol Rev. 1994;7:426–461. doi:10.1128/CMR.7.4.426
37. Mertens RB, Didier ES, Fishbein MC, et al. Encephalitozoon cuniculi microsporidiosis: infection of the brain, heart, kidneys, trachea, adrenal glands, and urinary bladder in a patient with AIDS. Mod Pathol. 1997;10:68–77.
38. Didier ES, Snowden KF, Shadduck JA. Biology of microsporidian species infecting mammals. Adv Parasitol. 1998;40:283–320. doi:10.1016/S0065-308X(08)60125-6
39. Didier ES. Microsporidiosis: an emerging and opportunistic infection in humans and animals. Acta Trop. 2005;94:61–76. doi:10.1016/j.actatropica.2005.01.010
40. Perec-Matysiak A, Leśniańska K, Buńkowska-Gawlik K, et al. The opportunistic pathogen Encephalitozoon cuniculi in wild living Murinae and Arvicolinae in Central Europe. Eur J Protistol. 2019;69:14–19. doi:10.1016/j.ejop.2019.02.004
41. Nell B, Fuchs-Baumgartinger A, Fritsche J, et al. Distribution of Encephalitozoon cuniculi infections in the cat resulting in ophthalmological symptoms in Europe. Vet Ophthalmol. 2014;17:E24.
42. Milnes E, Delnatte P, Cai HY, et al. Systemic Encephalitozoonosis due to Encephalitozoon cuniculi strain IV in a Vancouver Island Marmot (Marmota Vancouverensis). J Zoo Wildl Med. 2018;49:484–488. doi:10.1638/2017-0175.1
43. Didier ES, Stovall ME, Green LC, et al. Epidemiology of microsporidiosis: sources and modes of transmission. Vet Parasitol. 2004;126:145–166. doi:10.1016/j.vetpar.2004.09.006
44. Weiss LM. and now microsporidiosis. Ann Intern Med. 1995;123:954–956. doi:10.7326/0003-4819-123-12-199512150-00012
45. Weiss LM, Keohane EM. Microsporidia at the turn of the millenium: Raleigh 1999. J Eukaryot Microbiol. 1999;46:3S–5S. doi:10.1111/j.1550-7408.1999.tb06055.x
46. Wittner M, Weiss L. The Microsporidia and Microsporidiosis. Washington DC: ASM Press; 1995.
47. Kicia M, Weselowska M, Kopacz Z, et al. Disseminated infection of Encephalitozoon cuniculi associated with osteolysis of hip periprosthetic tissue. Clin Infect Dis. 2018;67:1228–1234. doi:10.1093/cid/ciy256
48. Brdíčková K, Sak B, Holubová N, et al. Encephalitozoon cuniculi genotype II concentrates in inflammation foci. J Inflamm Res. 2020;13:583–593. doi:10.2147/JIR.S271628
49. Kotková M, Sak B, Květoňová D, et al. Latent microsporidiosis caused by Encephalitozoon cuniculi in immunocompetent hosts: a murine model demonstrating the ineffectiveness of the immune system and treatment with albendazole. PLoS One. 2013;8:e60941. doi:10.1371/journal.pone.0060941
50. De Bosscuere H, Wang Z, Orlandi PA. First diagnosis of Encephalitozoon intestinalis and E. hellem in a European brown hare (Lepus europaeus) with kidney lesions. Zoonoses Public Health. 2007;54:131–134. doi:10.1111/j.1863-2378.2007.01034.x
51. Katzwinkel-Wladarsch S, Lieb M, Helse W, et al. Direct amplification and species determination of microsporidian DNA from stool specimens. Trop Med Int Health. 1996;1:373–378. doi:10.1046/j.1365-3156.1996.d01-51.x
52. Wolk DM, Schneider SK, Wengenack NL, et al. Real-time PCR method for detection of Encephalitozoon intestinalis from stool specimens. J Clin Microbiol. 2002;40:3922–3928. doi:10.1128/JCM.40.11.3922-3928.2002
53. Dai J, Wang P, Adusumilli S, et al. Antibodies against a tick protein, Salp15, protect mice from the Lyme disease agent. Cell Host Microbe. 2009;6:482–492. doi:10.1016/j.chom.2009.10.006
54. Sak B, Jandová A, Doležal K, et al. Effects of selected Indonesian plant extracts on E. cuniculi infection in vivo. Exp Parasitol. 2017;181:94–101. doi:10.1016/j.exppara.2017.07.014
55. Sak B, Kotková M, Hlásková L, et al. Limited effect of adaptive immune response to control encephalitozoonosis. Parasite Immunol. 2017;39:e12496. doi:10.1111/pim.12496
56. Gannon J. A survey of Encephalitozoon cuniculi in laboratory animal colonies in the United Kingdom. Lab Anim. 1980;14:91–94. doi:10.1258/002367780780942917
57. Kotková M, Sak B, Kváč M. Differences in the intensity of infection caused by Encephalitozoon cuniculi genotype II and III – comparison using quantitative real-time PCR. Exp Parasitol. 2018;192:93–97. doi:10.1016/j.exppara.2018.07.019
58. Sak B, Brdíčková K, Holubová N, et al. Encephalitozoon cuniculi genotype III evinces a resistance to albendazole treatment in both immunodeficient and immunocompetent Mice. Antimicrob Agents Chemother. 2020;64:e00058–20. doi:10.1128/AAC.00058-20
59. Couzinet S, Cejas E, Schittny J, et al. Phagocytic uptake of Encephalitozoon cuniculi by nonprofessional phagocytes. Infect Immun. 2000;68:6939–6945. doi:10.1128/IAI.68.12.6939-6945.2000
60. Nassonova E, Tokarev YS, Trammer T, et al. Phagocytosis of Nosema grylli (Microsporidia, Nosematidae) spores in vivo and in vitro. J Eukar Microbiol. 2001;48:83–84. doi:10.1111/j.1550-7408.2001.tb00462.x
61. Fischer J, Tran D, Juneau R, et al. Kinetics of Encephalitozoon spp. infection of human macrophages. J Parasitol. 2008;94:169–175. doi:10.1645/GE-1303.1
62. Niederkorn JY, Shadduck JA. Role of antibody and complement in the control of Encephalitozoon cuniculi infections by rabbit macrophages. Infect Immun. 1980;27:995–1002. doi:10.1128/iai.27.3.995-1002.1980
63. Weidner E. Interactions between Encephalitozoon cuniculi and macrophages. Parasitophorous vacuole growth and the absence of lysosomal fusion. Z Parasitenkd. 1975;47:1–9. doi:10.1007/BF00418060
64. Didier ES, Weiss LM. Microsporidiosis: not just in AIDS patients. Curr Opin Infect Dis. 2011;24:490–495. doi:10.1097/QCO.0b013e32834aa152
65. Zender HO, Arrigoni E, Eckert J, et al. A case of Encephalitozoon cuniculi peritonitis in a patient with AIDS. Am J Clin Pathol. 1989;92:352–356. doi:10.1093/ajcp/92.3.352
66. Tosoni A, Nebuloni M, Ferri A, et al. Disseminated microsporidiosis caused by Encephalitozoon cuniculi III (dog type) in an Italian AIDS patient: a retrospective study. Mod Pathol. 2002;15:577–583. doi:10.1038/modpathol.3880566
67. Torres G, Izquierdo F, Capo V, et al. Genital microsporidiosis in women with AIDS: a post-mortem study. Rev Iberoam Micol. 2013;30:47–50. doi:10.1016/j.riam.2012.04.001
68. Kicia M, Wesolowska M, Jakuszko K, et al. Concurrent infection of the urinary tract with Encephalitozoon cuniculi and Enterocytozoon bieneusi in a renal transplant recipient. J Clin Microbiol. 2014;52:1780–1782. doi:10.1128/JCM.03328-13
69. Ladapo TA, Nourse P, Pillay K, et al. Microsporidiosis in pediatric renal transplant patients in Cape Town, South Africa: two case reports. Pediatr Transplant. 2014;8:E220–226. doi:10.1111/petr.12327
70. Neumayerová H, Juránková J, Jeklová E, et al. Seroprevalence of Toxoplasma gondii and Encephalitozoon cuniculi in rabbits from different farming systems. Vet Parasitol. 2014;204:184–190. doi:10.1016/j.vetpar.2014.04.020
71. Kotková M, Sak B, Hlásková L, et al. The course of infection caused by Encephalitozoon cuniculi genotype III in immunocompetent and immunodeficient mice. Exp Parasitol. 2017;182:16–21. doi:10.1016/j.exppara.2017.09.022
72. Sak B, Brdíčková K, Holubová N, et al. The course of infection of Encephalitozoon cuniculi genotype I in mice possess combination of features reported in genotypes II and III. Exp Parasitol. 2021;224:108101. doi:10.1016/j.exppara.2021.108101
 © 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.