Back to Journals » Nature and Science of Sleep » Volume 12
Comparing the Effects of Sleep and Rest on Memory Consolidation
Authors Tucker MA ![]() , Humiston GB
, Humiston GB ![]() , Summer T, Wamsley E
, Summer T, Wamsley E
Received 20 July 2019
Accepted for publication 28 November 2019
Published 3 February 2020 Volume 2020:12 Pages 79—91
DOI https://doi.org/10.2147/NSS.S223917
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Sutapa Mukherjee
Matthew A Tucker,1 Graelyn B Humiston,2 Theodore Summer,2 Erin Wamsley2
1University of South Carolina School of Medicine, Department of Biomedical Sciences, Greenville, SC, USA; 2Furman University, Department of Psychology and Program in Neuroscience, Greenville, SC, USA
Correspondence: Matthew A Tucker
University of South Carolina School of Medicine, Department of Biomedical Sciences, Health Sciences Administration Building (HSAB) 248, 701 Grove Road, Greenville, SC 29605, USA
Tel +1 864-455-8945
Email [email protected]
Introduction: There is ample evidence that overnight sleep and daytime naps benefit memory retention, compared to comparable amounts of active wakefulness. Yet recent evidence also suggests that a period of post-training rest (eg, quiet wakefulness with eyes closed) provides a similar memory benefit compared to wake. However, the relative benefits of sleep vs quiet waking rest on memory remain poorly understood. Here, we assessed the extent to which sleep provides a unique memory benefit, above and beyond that conferred by quiet waking rest.
Methods: In a sample of healthy undergraduate students (N=83), we tested the effect of 30 mins of post-learning sleep, rest, or active wake on concept learning (dot pattern classification) and declarative memory (word pair associates) across a 4-hr daytime training-retest interval.
Results and Conclusions: Contrary to our hypotheses, we found no differences in performance between the three conditions for either task. The findings are interpreted with reference to methodological considerations including the length of the experimental interval, the nature of the tasks used, and challenges inherent in creating experimental conditions that can be executed by participants.
Keywords: rest, sleep, memory consolidation, concept learning, declarative memory
Introduction
Numerous studies demonstrate that periods of sleep, whether obtained in long consolidated bouts (eg, overnight sleep), or even short daytime naps, result in better memory retention compared to typical waking activities, such as going about one’s daily routine.1,2 These findings have been interpreted as support for the hypothesis that sleep is uniquely equipped to support memory consolidation.2,3 More recently, however, there has been a growing interest in determining whether certain types of waking brain states might also represent an optimal condition for memory consolidation.4,5 For example, recent evidence suggests that even a brief period of waking rest with eyes closed following learning may confer a memory benefit similar to that seen following sleep for some memory tasks.4,6 However, the comparative benefits of sleep vs resting wakefulness remain poorly understood. Although it has frequently been hypothesized that sleep-specific mechanisms uniquely support memory consolidation,7–9 the extent to which post-training sleep confers a memory performance advantage compared to resting wakefulness remains unclear.
Even though memories are clearly retained over periods of wake, sleep is thought to represent a more opportune time for memory processing, as sensory processing is reduced to a minimum and the neurobiological milieu turns to favor consolidation, rather than encoding.5 However, certain forms of wakefulness may be equally beneficial for memory in this respect. During our waking hours, we go through periods in which we are actively focused on processing environmental stimuli (“online” or active wakefulness), but we also experience periods of disengagement from the sensory environment, in which our focus turns to internal thoughts and feelings (“offline” or quiet wakefulness).4,10 These offline waking states, in particular, have been proposed to facilitate the consolidation of memory,4 as well as other important cognitive processes such as insight formation (eg, “aha” moments).11
While there are clear neurobiological differences between sleep and resting wake, aspects of resting wake mirror some key features of sleep thought to account for its memory benefits. Relative to active wake (eg active focus on processing environmental stimuli), which is characterized by low amplitude, desynchronized electroencephalography (EEG) in the higher beta frequencies (>25 Hz), both resting wakefulness and sleep are marked by slower global brain rhythms. Relative to active wakefulness, both sleep and resting wake are also characterized by increased activation of default-mode network brain structures, including medial temporal and medial frontal regions associated with memory processing.12–14 In both rest and sleep, activation of these structures may play an important memory processing function.14–16 Finally, sleep and resting wake are also both characterized by reduced acetylcholine levels and the concurrent expression of hippocampal sharp-wave/ripples (SWRs), both of which may contribute to memory consolidation by facilitating hippocampal-cortical communication during offline states.17–19
While daytime naps have been shown to benefit diverse memory types (eg, declarative,20 motor,21 spatial,22 perceptual,23 and concept learning24), the benefit of waking brain states for memory has only recently been investigated. Thus far, however, it appears that resting wake may also benefit several different forms of learning and memory. A brief period of resting wake following learning results in better retention of verbal information, relative to an equivalent duration spent completing an unrelated distractor task.6,25 Similar effects have now been described for motor memory,26 spatial memory,27–29 and insight formation.11
In light of these recent observations, it is important to examine whether post-training sleep confers an additional benefit, above and beyond that which follows resting wakefulness. While it is possible that resting wake might facilitate exactly the same consolidation processes as those documented during sleep, it is also possible that the unique neurobiology of sleep might be necessary for consolidation of some forms of learning, or for retention of memories over longer periods of time. In a recent study from our own laboratory, for example, we found that, relative to a period of active wakefulness, post-training resting wake enhanced motor skill learning at immediate test, but the benefit was not maintained 4hrs later in the day.26 This suggested that resting wake might be sufficient for short-term stabilization of motor learning, but that sleep may be required for long-term maintenance of these gains. Another study found that, over a 4 hr retention interval containing sleep vs quiet wake (in this case, meditation), retention for some forms of declarative memory (free recall), but not others (eg, cued recall of number-name associates) was improved.30 These observations could suggest that while resting wake is sufficient to allow stabilization of some forms of memory, this effect and its duration may depend on the memory type examined.
A handful of recent studies have attempted to compare memory following a short bout of sleep to memory following an equivalent duration of resting wake. In a tightly controlled study, the ideal resting wakefulness comparison condition would be one where, as in sleep, participants are reclining with eyes closed in a quiet and dark environment, while not completing any task or being exposed to any experimental stimuli. However, this can be challenging due to the difficulty of having participants maintain wakefulness under these conditions for the typical duration of a nap study (60–90 mins). For this reason, investigators have mainly approached this problem either by having “rest” participants complete a non-demanding, passive task (eg, listening to music or stories31,32), or by intermittently alerting rest participants to remain awake using a tone or other reminder.33 These approaches have led to mixed results.
An early and influential study of the effects of sleep on visual discrimination learning showed that while a post-training nap significantly benefitted subsequent performance, a similar performance benefit was not detected in a small control group (N=9) who rested while blindfolded and listened to an audio story.31 In a subsequent study using the same task, a post-training nap containing REM sleep similarly benefitted performance more than a rest period spent listening to music, although in this case sleep and rest differed only when stimuli had been subject to the effects of retroactive interference.32
Using different learning paradigms, other studies have concluded that sleep and quiet rest confer a similar memory benefit. For example, relative to active wakefulness, both sleep and quiet rest equally facilitated new learning in a hippocampus-dependent visual search task.34 Another study reported that both sleep and resting wakefulness (with alerting tones administered to maintain wakefulness) similarly benefitted performance on an auditory sequence-learning task, in comparison to active wake.33
Yet other recent studies have compared sleep, active wake, and quiet wake and found no difference between conditions (including, perhaps surprisingly, no difference between sleep and active wakefulness) for an alternating serial reaction time task35 and a paired associates task.36 While the above findings suggest that wakefulness and sleep might show equivalent memory benefits under some conditions, a direct comparison of the memory effects of sleep to those of completely task- and stimulus-free eyes closed rest has not yet been conducted.
Here, we report the first study directly comparing the memory benefit of sleep to that of a quiet eyes-closed resting condition in which participants were not exposed to any experimental stimuli or wake-promoting tones or other reminders. Importantly, this task-free and stimulus-free rest condition matches those in the above-reviewed studies demonstrating that brief (<20 mins) periods of waking rest benefit performance on verbal, procedural, spatial, and insight tasks, relative to equivalent periods spent completing a distractor task. We examined both a standard declarative memory task (paired associates learning), and a concept learning task that requires the abstraction of generalities from specific examples (dot-pattern classification).
Participants were trained on the dot pattern classification task and paired associates task immediately prior to a ≤30 mins interval filled with sleep, resting wake (reclining with eyes closed), or active wake (playing the video game Snood: http://snoodworld.com). This short time interval was selected as one that seemed to provide a reasonable compromise between being long enough for participants to obtain a substantive amount of sleep, yet short enough that participants could reasonably be expected to maintain wakefulness in the quiet rest condition. Participants were retested on these tasks ~3.5 hrs following the experimental intervention. We hypothesized that these three distinctly different brain states would differentially impact memory performance, with the sleeping brain providing the greatest performance benefit, but with resting wake also conferring a greater benefit than active wake.
Methods
Participants
Participants were 118 undergraduate students (N=69 female; Age=19.3±1.3 years (SD)) randomly assigned to one of three conditions (Sleep: N=41; Rest: N=44, Active Wake: N=33; see Figure 1 for experimental timeline). Fifteen participants were excluded due to EEG recording problems (N=5), failure to report for the delayed retest session at the appointed time (N=4), failure to follow instructions (N=1 participant attempted to write down the answers for the paired associates task, N=1 participant was discovered to have completed the paired associates task before in a prior study), or failure of the participant to correctly complete all training session procedures (N=4). Additionally, N=36 participants failed to successfully complete their assigned condition because they either fell asleep during the Rest condition or failed to sleep during the Sleep condition. However, N=16 of these participants did meet inclusion criteria for a different experimental group (see Table 1 and “Experimental Conditions”, below), and thus were reclassified for purposes of analysis: Sleep participants (N=7) who slept for less than 2 mins and never entered stage N2 sleep were reassigned to the Rest group, while Rest participants (N=9) who slept more than 10 mins were reassigned to the Sleep group. The remaining N=20 participants who failed to complete their assigned condition did not meet inclusion criteria for a different experimental group, and were excluded from all further analysis. Thus, the final sample consisted of 83 participants (Sleep: N=28; Rest: N=28, Active Wake: N=27). Statistical analyses for the original sample (randomly assigned to condition and not including the reassigned participants) are presented in the Supplemental Analyses and in Supplemental Table 1 and largely mirror the results reported below.
 |
Table 1 Participant Exclusions |
 |
Figure 1 Experimental Timeline. 00:00 represents study start time. Following EEG hookup, participants completed the PVT and training for the Concept Learning and Paired Associates tasks in randomized order, followed by one of three experimental conditions. Participants then left the laboratory and returned later to complete the second PVT and the Concept Learning and Paired Associates tests. Rest = Resting Wake, Wake = Active Wake. |
All participants signed informed consent and were compensated $10/hour or received course credit in an introductory psychology course. This research was approved by the Furman University Institutional Review Board, and was conducted in accordance with the Declaration of Helsinki.
Memory Tasks
Concept Learning – Dot Pattern Classification
The concept learning task was based on the dot pattern classification task originally developed by Posner (1967), and is identical to that described in our previous paper demonstrating that sleep benefits performance on this task.24 Participants learned to classify abstract patterns of dots into categories, based on their spatial pattern (Figure 2). Three categories were used (A, B, and C), based on the stimuli developed by Shin and Nosofsky.37 Each category consisted of 10 different patterns: six exemplars presented at training (“Old Exemplars”), three new exemplars presented at retest, which were not seen at training (“New Exemplars”), and the “Prototype” of each category, defined as the spatial average of the exemplars. Exemplars were created by distorting the category prototype through application of a statistical rule. New exemplars varied in their similarity to the prototype, having a low, medium, or high level of distortion.37 Old Exemplars in the current study were all medium-distortion patterns.
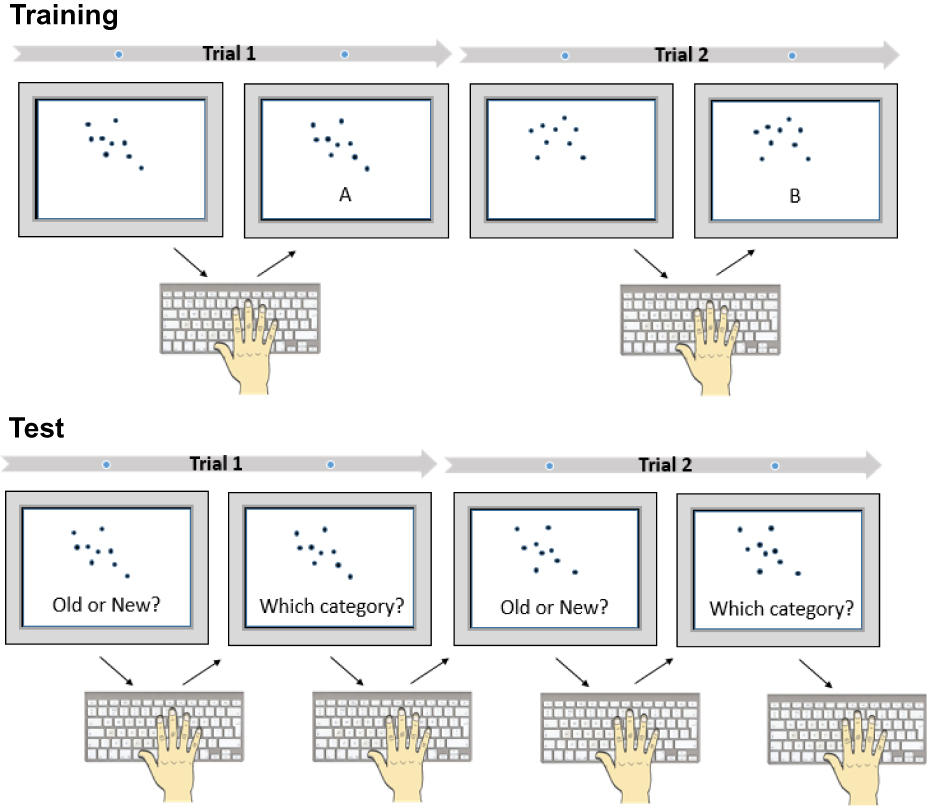 |
Figure 2 Dot Pattern Classification Task (Concept Learning). At training, participants are presented a series of dot patterns (exemplars) to classify these into three categories (A, B, and C). Only categories A and B are shown in this example. At test, participants must classify the dot patterns seen during training (Old Exemplars), novel dot patterns not previously seen but belonging to the same three categories (New Exemplars), and the prototypes from which each category was derived, also not previously seen (Prototypes). |
The task was presented using OpenSesame.38 The training session consisted of the presentation of 270 medium-distortion dot patterns. When each dot pattern was presented, the participant classified the pattern as “A”, “B”, or “C”, followed by feedback about whether the classification was correct. During the retest session, participants were presented with both the Old Exemplars seen at training, New Exemplars that they had not previously seen, and the Prototype pattern for each category (also not seen before). Participants again classified dot patterns into the A, B, and C categories, and also indicated whether the presented pattern had been seen before (“Old”) or not (“New”). Participants classified 30 patterns (6 Old Exemplars, 3 New Exemplars, and 1 Prototype for each category) in 3 blocks of 30 trials, with two untimed breaks in between. At retest, classification performance was quantified as the number of stimuli correctly classified. Recognition performance for exemplars was quantified as positive predictive value (hits/hits+false alarms), and for prototypes was quantified as (hits/hits+misses).
Paired Associates
The Paired Associates task consisted of 40 semantically unrelated word pairs (eg, alligator-cigar) used in Payne et al.39 During training, each pair was presented on the screen for 3 s. Following presentation of all 40 word pairs, participants were tested on their memory for the pairs using a cued recall paradigm. The first word from each pair was presented, and participants had to type the word that was paired with it (eg, alligator-________). Feedback was given after each answer. If a participant answered at least 60% (24 out of 40) of the pairs correctly, the test phase ended. If they did not reach 60% correct, they were shown all 40 of the word pairs and tested again, repeating the process until they reached the 60% criterion. During the retest session, participants completed one final cued recall test, with no feedback. Performance was measured as number correct at training, raw change from training to retest (# correct at retest - # correct at training), and percent change from training to retest (((# correct at retest - # correct at training)/# correct at training) *100).
Experimental Conditions
We approached the problem of designing length-equivalent rest and sleep conditions by yoking the duration of a stimulus-free eyes-closed resting wake condition to the time that it took sleep participants to obtain a minimum of 10 mins of sleep. In this between-subjects design, condition (Sleep, Rest, or Active Wake) was assigned by triads, with the first participant in each triad assigned to the Sleep condition, the second to the Rest condition, and the third to the Active Wake condition. For each triad, experimental condition length was yoked to that of the Sleep participant, such that if the Sleep participant’s nap opportunity was 22 mins, the duration of the Rest and Active Wake condition would also be 22 mins. During the experimental phase room conditions were kept as similar as possible across the three conditions, except where differences are described below. Light levels were the same (lights on), the door was closed after the researcher started the session, and participants were video monitored during the experimental phase.
Sleep
Participants were instructed to lie on the bed in the testing room and try to fall asleep, and to minimize movement. The light was left on and the door was closed. The interval ended when the participant had a) been in the room for 30 mins, b) had slept for 20 mins, or c) had fallen asleep and then woken up, but had been asleep for at least 10 mins and had been in the room for at least 20 mins. To be included in analysis for this condition, it was decided a priori that participants must obtain at least 10 total minutes of sleep. None of the participants in this sleep condition obtained REM sleep (see Table 2) and all sleep participants obtained N2 sleep.
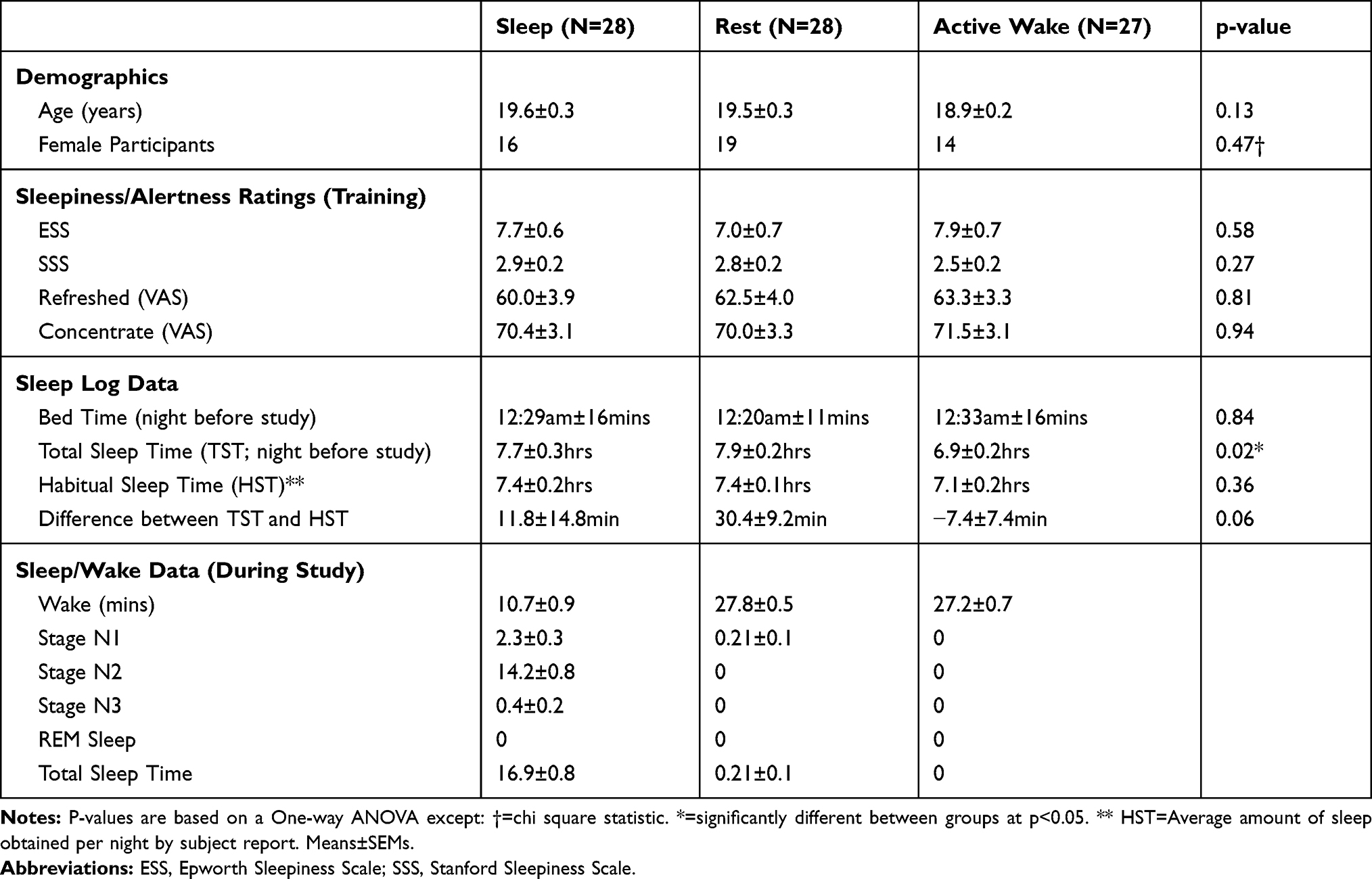 |
Table 2 Participant Information |
Rest
Participants were instructed to sit upright against a pillow on the bed in the testing room, and to minimize movement, keep their eyes closed, and stay awake. The length of the interval was determined by the length of the yoked Sleep participant. To be included in analysis for this condition, it was decided a priori that participants must not obtain more than 2 mins of sleep during the interval, and that they not enter N2 sleep (ie, any sleep obtained was N1 sleep). Of the final Rest group included in analysis, N=5 slept, obtaining less than 2 mins of N1 sleep and no N2 sleep, and the remainder obtained no sleep.
Active Wake
Participants in this condition played the computer game Snood (http://snoodworld.com) at “medium” difficulty. They were instructed to play the game continuously, starting a new game each time they won or lost. The duration of game play was determined by the length of the yoked Sleep participant.
Procedure
Training began between 9:00am and 1:30pm (Mean: 11:32am±1 hr 22 mins (SD); Figure 1), and participants were instructed not to consume caffeine after 10am on the day of their session. Training lasted approximately 90 mins, and the retest session began ~4 hrs after the end of training. After arriving at the laboratory, participants completed a demographics questionnaire, a general information form, a 3-day retrospective sleep log, and the Epworth Sleepiness Scale (ESS), a measure of trait sleepiness.40
While participants completed the initial forms, scalp electrodes were applied to record sleep and wake EEG activity. Electrodes were applied to F3/F4, C3/C4, O1/O2, each referenced to the contralateral mastoid. Electrodes applied near the eyes (right and left outer canthus) and chin recorded eye movements and muscle activity, respectively. EEG signals were recorded at 400hz with a Grass-Telefactor Aura amplifier. Impedances for all EEG recordings were kept below 10kΩ. All EEG recordings were scored in 30-s epochs according to the standardized criterial established by the American Academy of Sleep Medicine.47
After electrode placement, participants completed the Stanford Sleepiness Scale (SSS), a measure of state sleepiness,41 and two visual analog scales rating how “refreshed” the participant felt and their ability to “concentrate”. These were followed by the Psychomotor Vigilance Task (PVT).42 In this computerized vigilance task, participants kept their eyes fixed on the center of the computer screen, and pressed the spacebar each time a red dot appeared. Participants were then trained on the Paired Associates task and the Concept Learning task. The order of the tasks was counterbalanced so that an equal number of participants did each order (main sample: Concept Learning first (N=42), Paired Associates first (N=41)). Within condition, Concept Learning was completed first for 12 of 28 in the Sleep condition, 16 of 28 in the Rest condition, and 14 of 27 in the Wake condition.
After Concept Learning and Paired Associates training, participants began their assigned experimental condition (Sleep, Rest, or Active Wake) in the testing room with the lights on, with continuous EEG recording.
Participants’ subjective experience during each experimental condition is of interest to our lab in light of our prior work suggesting that the consolidation of memory during offline states of rest and sleep may be reflected in the contents of conscious experience.43 Thus, following each experimental condition, participants completed a subjective experience questionnaire while the experimenter removed the electrodes. This questionnaire had participants estimate the percentage of time they spent in 15 pre-defined mental categories: “Thinking about learning tasks from earlier”, “Thinking about something that happened earlier today”, “Thinking about something that happened yesterday to a week ago”, “Thinking about something that happened last year or several years ago”, “Thinking about the remainder of the day”, “Thinking about something that will happen tomorrow to next week”, “Thinking about something that will happen in the next year or several years”, “Thinking about relaxation”, “Thinking about something else”, “Mind was blank”, “Counting the time”, “Focused meditation”, “Sleeping”, “Snood (if applicable)”, and “Other”. Participants were then instructed to return for the retest, which was scheduled to start 5 hrs following the start of the training session. Participants were allowed to leave the laboratory during this time. They were instructed to refrain from napping and from consuming drugs or alcohol. When they returned for the test, participants were asked whether they had napped, or consumed drugs or alcohol. No participants responded in the affirmative to these questions.
Upon returning to the lab for the retest session (4.43±0.17 hrs after the start of task training), participants again completed the SSS and visual analog scales, followed by the PVT. Participants were then retested on the Paired Associates and Concept Learning tasks in the same order as during Session 1. After finishing the tests, participants were debriefed and compensated.
Statistical Analyses
To statistically compare performance across experimental conditions, one-way ANOVAs were calculated with condition entered as the independent variable. Pearson’s correlations were conducted to test the associations between subjective experience questionnaire data and task performance. All statistical analyses were conducted using IBM SPSS Statistics for Windows, Version 22.0, Armonk, NY. For parametric tests, all dependent variables conformed to the normal distribution. Results for each analysis are presented as the mean±SEM unless otherwise specified. Statistical significance was set to p<0.05.
Results
Sample Characteristics and Measures of Alertness
Age and sex data, subjective alertness/sleepiness reports, sleep log data, and sleep/wake data for each condition are described in Table 2. Mean reaction times on the PVT (omitting lapses (reaction times > 500ms)) were comparable between groups at the time of training (Sleep: 377.8±10.9 ms, Rest: 363.3±7.8, Active Wake: 357.6±7.2, one-way ANOVA, F(2,79)=1.38, p=0.26). From training to retest, PVT reaction times decreased (became faster) similarly for the three conditions (Sleep: −14.4±8.1 ms, Rest: −12.8±6.0, Active Wake: −11.6±6.1, one-way ANOVA, F(2,71)=0.04, p=0.96). One-way ANOVAs examining change in subjective ratings from training to retest for the remaining measures of alertness revealed no significant effect of group on change over time in ability to concentrate (positive values = greater able to concentrate; Sleep: 7.9±2.9, Rest: 6.4±2.7, Active Wake: 2.2±2.1, F(2,80)=1.25, p=0.29), how refreshed participants felt (positive values = more refreshed; Sleep: 11.8±4.3, Rest: 9.6±3.1 Active Wake: 5.6±3.1, F(2,80)=0.79, p=0.46), or SSS (negative values = more alert; Sleep: −0.8±0.2, Rest: −0.7±0.2 Active Wake: −0.3±0.1, F(2,80)=2.61, p=0.08).
Concept Learning: Dot Pattern Classification
At retest, participants in the three conditions did not differ in their ability to classify Old Exemplars (Sleep: 35.7±1.9, Rest: 37.9±1.9, Wake: 36.9±1.6, F(2,80)=0.36, p=0.70, ηp2=0.009), New Exemplars (Sleep: 14.2±0.9, Rest: 14.0±1.0, Wake: 14.6±0.9, F(2,80)=0.10, p=0.90, ηp2=0.003), or Prototypes (Sleep: 5.2±0.4, Rest: 5.7±0.4, Wake: 5.7±0.4, F(2,79)=0.62, p=0.54, ηp2=0.016; Figure 3). Recognition performance also did not differ between the conditions for Exemplars (Sleep: 0.63±0.02, Rest: 0.66±0.02, Wake: 0.61±0.02, F(2,79)=1.62, p=0.20, ηp2=0.039) or Prototypes (Sleep: 0.44±0.03, Rest: 0.45±0.04, Wake: 0.38±0.04, F(2,79)=1.22, p=0.30, ηp2=0.030; Figure 4). We did not observe any significant correlations between sleep parameters (total sleep time, time spent in stages N1, N2, and N3) in the sleep group and concept learning (correct identifications or recognition performance) (Pearson’s correlations, all p values > 0.16). Task order (concept learning first vs paired associated first) did not interact with condition for Exemplar categorization (Old, p=0.64; New, p=0.52, Prototype, p=0.12), Exemplar recognition (p=0.97), or Prototype recognition (p=0.41), nor were there any significant main effects of task order on performance (all p values > 0.09).
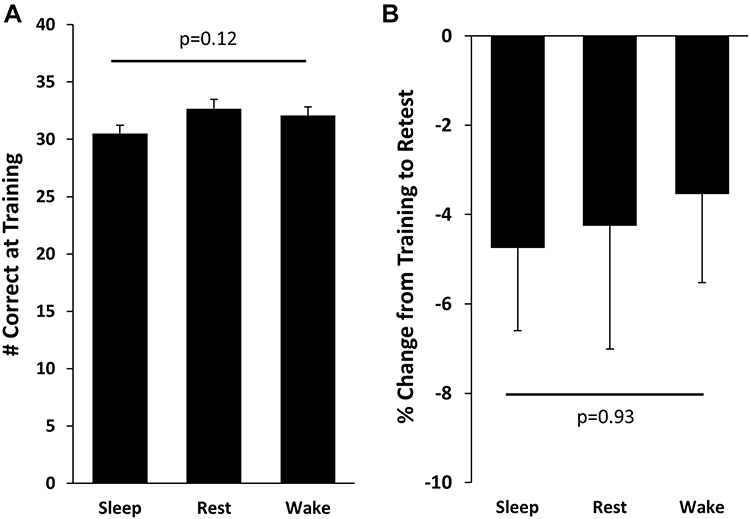 |
Figure 3 Paired Associates Performance. (A) Number correct at the end of the training session;(B) Percent change from training to retest. Bars represent Means±SEMs. |
 |
Figure 4 Pattern Classification Performance – Correct Classifications. At retest, participants indicated whether dot patterns belonged to category (A, B, or C). Means±SEMs. |
Because total sleep time the night before the study, as reported on the 3-day sleep log, was shorter in the Active Wake group compared to Sleep and Rest (Table 2), we conducted the same analyses with total sleep time entered as a covariate. This did not impact the above results (greatest p-value change was from p=0.45 to p=0.20 for prototype recognition performance). For the Paired Associates and Concept Learning tasks, the time that the training session occurred was not associated with task performance at training or retest (all p values for Pearson’s correlations >0.09). Time of training also did not differ between experimental groups (F(2,82)=0.39, p=0.68). Prior research with both the paired associated task39,44,45 and concept learning task24 has failed to detect time-of-day effects on either encoding or retrieval.
Paired Associates
The number of trials required to reach 60% criterion at training was similar across conditions (Sleep: 2.3±0.1 trials, Rest: 2.0±0.1, Wake: 2.3±0.1, F(2,80)=1.61, p=0.21) as was the number of items correct at immediate test at the end of the training session (Sleep: 30.5±0.7 correct, Rest: 32.7±0.8, Wake: 32.1±0.8, F(2,80)=2.20, p=0.12; Figure 5A). At retest, participants in all conditions demonstrated similar declines in performance, both in terms of raw change (Sleep: −1.5±0.5 word pairs, Rest: −1.3±0.7, Wake: −1.2±0.6, F(2,80)=0.06, p=0.94, ηp2=0.002; Figure 5B) and percentage change from training to retest (Sleep: −4.8±1.9%, Rest: −4.3±2.7%, Wake: −3.5±2.0%, F(2,80)=0.08, p=0.93, ηp2=0.002). We did not observe any significant relationships between sleep parameters (total sleep time, time spent in stages N1, N2, and N3) in the sleep group and raw change/percent change in recall from training to retest (Pearson’s correlations, all p values > 0.08). There was no main effect of task order on change in paired associates (p=0.34), nor was there an interaction effect (p=0.50). Entering total sleep time from the 3-day sleep log as a covariate had no influence on the findings for paired associates (correct items at training, F(2,69)=1.61, p=0.21; raw change from training to retest, F(2,69)=0.09, p=0.91; % change from training to retest, F(2,69)=0.05, p=0.95). For the Paired Associates and concept learning tasks, the time of day that the training session occurred was not associated with task performance at training or retest (all p values for Pearson’s correlations >0.09). Time of training also did not differ between experimental groups (F(2,82)=0.39, p=0.68). An analysis of sex differences in concept learning and paired associates performance yielded one significant difference for recognition of prototypes (p=0.04), which did not remain significant after correction for multiple comparisons using the least significant difference method (LSD).
 |
Figure 5 Pattern Classification Performance – Recognition Accuracy. At retest, participants indicated whether the dot patterns had been presented during the training session (Old Exemplars) or not (New Exemplars and Prototypes). (A) Accuracy for Exemplars: Hits/(Hits+FalseAlarms) = Positive Predictive Value. (B) Accuracy for prototypes: Hits/(Hits+Misses) = % Correct. Means±SEMs. |
Subjective Experience Questionnaire Data
A subjective experience questionnaire was administered to all participants to document the percentage of time spent engaged in each of 15 thought processes during the 30-min experimental sleep/wake/rest interval (Figure 6). After correcting for multiple comparisons using the Benjamini–Hochberg method to control for false discovery rate,46 we found no significant correlations between these thought processes and task performance.
 |
Figure 6 Subjective Experience Questionnaire Data. Participants estimated the percentage of time spent engaged in 15 thought processes during the 30-min experimental phase, 11 of which were collapsed into three categories: Thinking about the past = “Thinking about something that happened earlier today”+ “Thinking about something that happened yesterday to a week ago”+ “Thinking about something that happened last year or several years ago”; Thinking about the future = “Thinking about the remainder of the day” + “Thinking about something that will happen tomorrow to next week” + “Thinking about something that will happen in the next year or several years”; Thinking not related to the past or future = “Thinking about relaxation” + “Thinking about something else” + “Mind was blank” + “Counting the time” + “Focused meditation”. |
Discussion
There is a considerable body of research describing the benefits of sleep (compared to active wake) for various types of memory, including declarative, motor, and perceptual memory.1,47 Recent studies also show that a post-training period of resting wake (versus active wake) can confer similar memory benefits.25,26,48 Thus far, it remains unclear whether sleep provides a unique additional memory benefit beyond that described to occur following eyes-closed rest. The current study is one of a small number attempting to directly compare the memory effects of sleep to those of waking rest, and is to our knowledge the first attempt to do this using a completely task- and stimulus-free resting condition. Contrary to expectations, we found no evidence that our sleep, active wake, and resting wake conditions differentially affected retention for either the declarative word pairs task or the concept learning task. Although sleep has most often been described as a brain state that functions to stabilize and strengthen memory, it may also support qualitative memory transformations, such as the extraction of commonalities from specific, correlated examples (gist extraction: Payne, Stickgold et al 2008;49 probabilistic learning: Barsky, et al 2015;50 Durrant, et al 201151). A related form of learning that exemplifies this particular cognitive skill is concept learning, in which patterns extracted from learned information facilitate the identification of similar (though novel) patterns.52,53 A previous study from our lab, using a 12hr training-retest interval that spanned a full night of sleep or a day of wake, showed that overnight sleep facilitated the accurate classification of novel dot pattern exemplars, compared to active wake.24 However, in the current study, which employed a ≤30 mins retention interval and a 3.5–4 hr training-retest interval, we found that sleep, resting wake, and active wake did not differentially impact categorization or recognition performance. Little-studied in the current literature, it is possible that concept formation as instantiated in this dot pattern classification task follows a different consolidation timeline than other forms of memory, one that requires closer to 12 hrs for effects of post-learning state to emerge, as opposed to the much shorter interval used in the present study.
But in contrast, performance on the word-pair associate task has been demonstrated to benefit from even a brief nap in numerous prior studies.20,54–57 As such, we had strongly expected to replicate the benefit of sleep relative to active wakefulness for this task, hypothesizing a graded effect on performance, such that sleep would impart the greatest benefits, with resting wake also being superior to active wake, and possibly equivalent to sleep.
Although failure to detect this basic effect of sleep vs active wakefulness could be a result of type II error arising from sampling error, we suspect that features of our study design may have prevented detection of this oft-reported memory effect. In order to achieve a task-free and stimulus-free waking rest comparison condition, we opted for sleep participants to engage in a very short nap opportunity, with participants obtaining less than 20 mins of sleep on average. However, in almost all prior studies examining the benefits of short bouts of sleep on memory, participants are allowed a much longer opportunity to sleep, usually 60–90 mins.20,23 This duration allows participants to obtain a significant amount of slow wave sleep and/or REM sleep, both of which are thought to play a critical role in memory consolidation.1 Although exceptions exist (see58), it may be that the typically reported memory benefits of sleep are expressed most robustly when participants are afforded at least an hour-long sleep opportunity in which slow wave sleep and/or REM sleep is observed.
Indeed, a few prior studies have suggested that very short naps in which participants do not enter SWS may not provide the same memory benefit as longer naps. For example, Alger et al59 reported that a 10-min nap did not impact memory as strongly as a 60-min nap. Thus, although the small number of participants obtaining SWS in the current study prevents us from comparing those who did vs did not enter this sleep stage, prior research suggests the possibility that some lower threshold amount of N2 and/or N3 sleep is required to observe robust memory effects. It is also possible that our failure to observe a memory effect here is due to the problems we experienced with differential attrition across groups. A substantial subset of participants was unable to fulfil the conditions of the study (remaining awake in the rest condition and sleeping in the sleep condition), and were therefore re-assigned to a new condition based on a-priori classification criteria. However, analyses based on original group assignment yielded essentially the same results (see Supplemental Analyses). Additionally, with or without the reassignment of these individuals, there was little indication that subjective (sleepiness scales) or objective measures (reaction times) of sleepiness/alertness differed across the three conditions.
This study highlights the difficulty of designing a protocol that simultaneously allows sensitive detection of the classically reported memory benefit of sleep, and the more recently observed benefit of completely task-free and stimulus-free rest. Here, the length of the experimental phase (Sleep, Rest, or Wake) was carefully considered to maximize the chance of observing group differences in memory, while ensuring that participants would be able to comply with instructions. However, this required creating an experimental situation in which participants (college students) spent up to 30 mins attempting to stay awake in a relaxing environment with little sensory stimulation (Rest), or were given only 30 mins to obtain a minimum of 10 mins of sleep (Sleep). These challenges, which may limit the methodological scope of this type of study, should be appreciated and carefully addressed in future research examining the effects of short intervals of post-training brain states on memory consolidation.
In conclusion, this study, while failing to detect significant differences in memory across three distinctly different experimental conditions, may highlight some of the factors necessary to detect robust memory effects of sleep and/or quiet rest, including the selected training-retest interval, and the length of experimental phase.
Acknowledgment
This work was supported by grant 1R15MH107891 from the National Institute of Mental Health.
Disclosure
Dr Erin Wamsley reports grants from NIMH during the conduct of the study and grants from BIAL Foundation, Portugal, outside the submitted work. The authors report no other conflicts of interest in this work.
References
1. Diekelmann S, Born J. The memory function of sleep. Nat Rev Neurosci. 2010;11(2):114–126. doi:10.1038/nrn2762
2. Rasch B, Born J. About sleep’s role in memory. Physiol Rev. 2013;93(2):681–766.
3. Abel T, Havekes R, Saletin JM, Walker MP. Sleep, plasticity and memory from molecules to whole-brain networks. Curr Biol. 2013;23(17):R774–R788. doi:10.1016/j.cub.2013.07.025
4. Wamsley EJ. Memory consolidation during waking rest. Trends Cogn Sci. 2019;23(3):171–173. doi:10.1016/j.tics.2018.12.007
5. Mednick SC, Cai DJ, Shuman T, Anagnostaras S, Wixted JT. An opportunistic theory of cellular and systems consolidation. Trends Neurosci. 2011;34(10):504–514. doi:10.1016/j.tins.2011.06.003
6. Dewar M, Alber J, Butler C, Cowan N, Della Sala S. Brief wakeful resting boosts new memories over the long term. Psychol Sci. 2012;23(9):955–960. doi:10.1177/0956797612441220
7. Penagos H, Varela C, Wilson MA. Oscillations, neural computations and learning during wake and sleep. Curr Opin Neurobiol. 2017;44:193–201. doi:10.1016/j.conb.2017.05.009
8. Girardeau G, Zugaro M. Hippocampal ripples and memory consolidation. Curr Opin Neurobiol. 2011;21(3):452–459. doi:10.1016/j.conb.2011.02.005
9. Timofeev I. Neuronal plasticity and thalamocortical sleep and waking oscillations. Prog Brain Res. 2011;193:121–144.
10. Smallwood J, Schooler JW. The science of mind wandering: empirically navigating the stream of consciousness. Annu Rev Psychol. 2015;66:487–518. doi:10.1146/annurev-psych-010814-015331
11. Craig M, Ottaway G, Dewar M. Rest on it: awake quiescence facilitates insight. Cortex. 2018;109:205–214. doi:10.1016/j.cortex.2018.09.009
12. Mason MF, Norton MI, Van Horn JD, Wegner DM, Grafton ST, Macrae CN. Wandering minds: the default network and stimulus-independent thought. Science. 2007;315(5810):393–395. doi:10.1126/science.1131295
13. Buckner RL, Vincent JL. Unrest at rest: default activity and spontaneous network correlations. Neuroimage. 2007;37(4):
14. Wang K, Yu C, Xu L, Qin W, Li K, Jiang T. Offline memory reprocessing: involvement of the brain’s default network in spontaneous thought processes. PLoS One. 2009;4(3):e4867. doi:10.1371/journal.pone.0004867
15. Wamsley EJ. Dreaming, waking conscious experience, and the resting brain: report of subjective experience as a tool in the cognitive neurosciences. Front Psychol. 2013;4:637. doi:10.3389/fpsyg.2013.00637
16. Huo L, Li R, Wang P, Zheng Z, Li J. The default mode network supports episodic memory in cognitively unimpaired elderly individuals: different contributions to immediate recall and delayed recall. Front Aging Neurosci. 2018;10:6. doi:10.3389/fnagi.2018.00006
17. Hasselmo ME, McGaughy J. High acetylcholine levels set circuit dynamics for attention and encoding and low acetylcholine levels set dynamics for consolidation. Prog Brain Res. 2004;145:207–231.
18. Buzsaki G. The hippocampo-neocortical dialogue. Cereb Cortex. 1996;6(2):81–92. doi:10.1093/cercor/6.2.81
19. Khodagholy D, Gelinas JN, Buzsaki G. Learning-enhanced coupling between ripple oscillations in association cortices and hippocampus. Science. 2017;358(6361):369–372. doi:10.1126/science.aan6203
20. Tucker MA, Hirota Y, Wamsley EJ, Lau H, Chaklader A, Fishbein W. A daytime nap containing solely non-REM sleep enhances declarative but not procedural memory. Neurobiol Learn Mem. 2006;86(2):241–247. doi:10.1016/j.nlm.2006.03.005
21. Nishida M, Walker MP. Daytime naps, motor memory consolidation and regionally specific sleep spindles. PLoS One. 2007;2(4):e341. doi:10.1371/journal.pone.0000341
22. Nguyen ND, Tucker MA, Stickgold R, Wamsley EJ. Overnight sleep enhances hippocampus-dependent aspects of spatial memory. Sleep. 2013;36(7):1051–1057. doi:10.5665/sleep.2808
23. Mednick S, Nakayama K, Stickgold R. Sleep-dependent learning: a nap is as good as a night. Nat Neurosci. 2003;6(7):697–698. doi:10.1038/nn1078
24. Graveline YM, Wamsley EJ. The impact of sleep on novel concept learning. Neurobiol Learn Mem. 2017;141:19–26. doi:10.1016/j.nlm.2017.03.008
25. Brokaw K, Tishler W, Manceor S, et al. Resting state EEG correlates of memory consolidation. Neurobiol Learn Mem. 2016;130:17–25. doi:10.1016/j.nlm.2016.01.008
26. Humiston GB, Wamsley EJ. A brief period of eyes-closed rest enhances motor skill consolidation. Neurobiol Learn Mem. 2018;155:1–6. doi:10.1016/j.nlm.2018.06.002
27. Craig M, Wolbers T, Strickland S, Achtzehn J, Dewar M. Rapid improvement of cognitive maps in the awake state. Hippocampus. 2019;29:862–868. doi:10.1002/hipo.v29.9
28. Craig M, Dewar M, Harris MA, Della Sala S, Wolbers T. Wakeful rest promotes the integration of spatial memories into accurate cognitive maps. Hippocampus. 2016;26(2):185–193. doi:10.1002/hipo.22502
29. Craig M, Dewar M, Della Sala S, Wolbers T. Rest boosts the long-term retention of spatial associative and temporal order information. Hippocampus. 2015;25(9):1017–1027. doi:10.1002/hipo.v25.9
30. Schonauer M, Pawlizki A, Kock C, Gais S. Exploring the effect of sleep and reduced interference on different forms of declarative memory. Sleep. 2014;37(12):1995–2007. doi:10.5665/sleep.4258
31. Mednick SC, Nakayama K, Cantero JL, et al. The restorative effect of naps on perceptual deterioration. Nat Neurosci. 2002;5(7):677–681. doi:10.1038/nn864
32. McDevitt EA, Duggan KA, Mednick SC. REM sleep rescues learning from interference. Neurobiol Learn Mem. 2015;122:51–62. doi:10.1016/j.nlm.2014.11.015
33. Gottselig JM, Hofer-Tinguely G, Borbely AA, et al. Sleep and rest facilitate auditory learning. Neuroscience. 2004;127(3):557–561. doi:10.1016/j.neuroscience.2004.05.053
34. Mednick SC, Makovski T, Cai DJ, Jiang YV. Sleep and rest facilitate implicit memory in a visual search task. Vision Res. 2009;49(21):2557–2565. doi:10.1016/j.visres.2009.04.011
35. Simor P, Zavecz Z, Horvath K, et al. Deconstructing procedural memory: different learning trajectories and consolidation of sequence and statistical learning. Front Psychol. 2018;9:2708. doi:10.3389/fpsyg.2018.02708
36. Piosczyk H, Holz J, Feige B, et al. The effect of sleep-specific brain activity versus reduced stimulus interference on declarative memory consolidation. J Sleep Res. 2013;22(4):406–413. doi:10.1111/jsr.12033
37. Shin HJ, Nosofsky RM. Similarity-scaling studies of dot-pattern classification and recognition. J Exp Psychol Gen. 1992;121(3):278–304. doi:10.1037/0096-3445.121.3.278
38. Mathot S, Schreij D, Theeuwes J. OpenSesame: an open-source, graphical experiment builder for the social sciences. Behav Res Methods. 2012;44(2):314–324. doi:10.3758/s13428-011-0168-7
39. Payne JD, Tucker MA, Ellenbogen JM, et al. Memory for semantically related and unrelated declarative information: the benefit of sleep, the cost of wake. PLoS One. 2012;7(3):e33079. doi:10.1371/journal.pone.0033079
40. Johns MW. A new method for measuring daytime sleepiness: the Epworth sleepiness scale. Sleep. 1991;14(6):540–545. doi:10.1093/sleep/14.6.540
41. Hoddes E, Zarcone V, Smythe H, Phillips R, Dement WC. Quantification of sleepiness: a new approach. Psychophysiology. 1973;10(4):431–436. doi:10.1111/psyp.1973.10.issue-4
42. Lim J, Dinges DF. Sleep deprivation and vigilant attention. Ann N Y Acad Sci. 2008;1129:305–322. doi:10.1196/nyas.2008.1129.issue-1
43. Wamsley EJ, Tucker M, Payne JD, Benavides JA, Stickgold R. Dreaming of a learning task is associated with enhanced sleep-dependent memory consolidation. Curr Biol. 2010;20(9):850–855. doi:10.1016/j.cub.2010.03.027
44. Tucker MA, Tang SX, Uzoh A, Morgan A, Stickgold R. To sleep, to strive, or both: how best to optimize memory. PLoS One. 2011;6(7):e21737. doi:10.1371/journal.pone.0021737
45. Ellenbogen JM, Hulbert JC, Jiang Y, Stickgold R. The sleeping brain’s influence on verbal memory: boosting resistance to interference. PLoS One. 2009;4(1):e4117. doi:10.1371/journal.pone.0004117
46. Benjamini Y, Hochberg Y. Controlling the false discovery rate: a practical and powerful approach to multiple testing. J R Stat Soc Series B Stat Methodol. 1995;57(1):289–300.
47. Dudai Y, Karni A, Born J. The consolidation and Transformation of memory. Neuron. 2015;88(1):20–32. doi:10.1016/j.neuron.2015.09.004
48. Craig M, Dewar M. Rest-related consolidation protects the fine detail of new memories. Sci Rep. 2018;8(1):6857. doi:10.1038/s41598-018-25313-y
49. Payne JD, Stickgold R, Swanberg K, Kensinger EA. Sleep preferentially enhances memory for emotional components of scenes. Psychol Sci. 2008;19(8):781–788. doi:10.1111/j.1467-9280.2008.02157.x
50. Barsky MM, Tucker MA, Stickgold R. REM sleep enhancement of probabilistic classification learning is sensitive to subsequent interference. Neurobiol Learn Mem. 2015;122:63–68. doi:10.1016/j.nlm.2015.02.015
51. Durrant SJ, Taylor C, Cairney S, Lewis PA. Sleep-dependent consolidation of statistical learning. Neuropsychologia. 2011;49(5):1322–1331. doi:10.1016/j.neuropsychologia.2011.02.015
52. Posner MI, Goldsmith R, Welton KE
53. Posner MI, Keele SW. On the genesis of abstract ideas. J Exp Psychol. 1968;77(3):353–363. doi:10.1037/h0025953
54. Alger SE, Lau H, Fishbein W. Delayed onset of a daytime nap facilitates retention of declarative memory. PLoS One. 2010;5(8):e12131. doi:10.1371/journal.pone.0012131
55. Lau H, Tucker MA, Fishbein W. Daytime napping: effects on human direct associative and relational memory. Neurobiol Learn Mem. 2010;93(4):554–560. doi:10.1016/j.nlm.2010.02.003
56. Lau EYY, McAteer S, Leung CNW, Tucker MA, Li C. Beneficial effects of a daytime nap on verbal memory in adolescents. J Adolesc. 2018;67:77–84. doi:10.1016/j.adolescence.2018.06.004
57. Sopp MR, Michael T, Mecklinger A. Effects of early morning nap sleep on associative memory for neutral and emotional stimuli. Brain Res. 2018;1698:29–42. doi:10.1016/j.brainres.2018.06.020
58. Lahl O, Wispel C, Willigens B, Pietrowsky R. An ultra short episode of sleep is sufficient to promote declarative memory performance. J Sleep Res. 2008;17(1):3–10. doi:10.1111/jsr.2008.17.issue-1
59. Alger SE, Lau H, Fishbein W. Slow wave sleep during a daytime nap is necessary for protection from subsequent interference and long-term retention. Neurobiol Learn Mem. 2012;98(2):188–196. doi:10.1016/j.nlm.2012.06.003
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.