Back to Journals » OncoTargets and Therapy » Volume 13
Clinicopathological Characteristics of TZAP Expression in Colorectal Cancers
Authors Jung SJ, Seo YR, Park WJ, Heo YR, Lee YH, Kim S ![]() , Lee JH
, Lee JH ![]()
Received 10 August 2020
Accepted for publication 26 November 2020
Published 17 December 2020 Volume 2020:13 Pages 12933—12942
DOI https://doi.org/10.2147/OTT.S274394
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Federico Perche
Soo-Jung Jung,1 Yu-Ri Seo,2 Won-Jin Park,1 Yu-Ran Heo,1 Yun-Han Lee,2 Shin Kim,3 Jae-Ho Lee1
1Department of Anatomy, Keimyung University School of Medicine, Daegu, South Korea; 2Department of Molecular Medicine, School of Medicine, Keimyung University, Daegu, South Korea; 3Department of Immunology, School of Medicine, Keimyung University, Daegu, South Korea
Correspondence: Jae-Ho Lee
Department of Anatomy, Keimyung University School of Medicine, 1035, Dalgubeol-Daero, Dalseo-Gu, Daegu 42601, Republic of Korea
Tel +82-53-258-7512
Email [email protected]
Shin Kim
Department of Immunology, Keimyung University School of Medicine, 1035, Dalgubeol-Daero, Dalseo-Gu, Daegu 42601, Republic of Korea
Email [email protected]
Purpose: The zinc finger protein, ZBTB48, is a telomere-associated protein. It was renamed as telomeric zinc finger-associated protein (TZAP) binding to elongated telomeres. However, its expression level was not measured in cancers.
Patients and Methods: We analyzed TZAP mRNA levels in 60 colorectal cancers (CRC) and its correlation with telomere length and TERT was studied.
Results: TZAP mRNA in CRC was higher statistically than that in paired non-cancerous tissues (p = 0.033). Higher TZAP was found in carcinoembryonic antigen (CEA)-positive CRCs (> 5 ng/mL) (p = 0.012). Shorter telomere was found in CRCs with high TZAP expression than that with low TZAP expression (p = 0.010). According to quantitative correlation analysis, TZAP has a correlation with age (r = − 0.349, p = 0.007), TERT (r = 0.279, p = 0.041) and telomere length (r = − 0.305, p = 0.021). TZAP expression did not harbor prognostic value in CRC. Inhibition of TZAP expression by siRNA suppresses cell growth in HT29 cells; however, it resulted in increased cell viability in HCT116 cells. TZAP inhibition induces a decrease in mRNA levels of TERT in both HT29 and HCT116 cells. TCGA data analysis showed higher expression of TZAP showed poorer overall survival in colon cancer (p = 0.001); however, it did not have a significance in rectal cancer (p = 0.951).
Conclusion: We suggested that TZAP may be a possible biomarker for CRC.
Keywords: TZAP, telomere, colorectal cancer, ZBTB48
Introduction
Telomeres are composed of 6-bp TTAGGG repeat sequences capping each end of eukaryotic chromosomes.1,2 In human somatic cells, an average length of telomere is 5–15 kilobases; however, approximately 30–200 base pairs were shortened at every cell division. It is counteracted by the telomerase reverse transcriptase or the alternative lengthening of telomeres (ALT) mechanism.3–5 Overly long telomeres are cut back to normal length by trimming mechanism.6–8 Although its precise regulation has not been recognized, a recent study found a novel protein, ZBTB48, regulating telomere length.9 As zinc finger protein, it was renamed telomeric zinc finger-associated protein (TZAP). In addition, they found that TZAP overexpression caused extremely short telomere. TZAP is located to chromosome 1p36, a region that is frequently rearranged or deleted in many cancers.10–12 Therefore, genetic change of TZAP could have a role in cancer pathogenesis; however, TZAP research has been studied rarely in cancer patients or cells.
Colorectal cancer (CRC) is one of the most common types of cancer in Korea and other developed countries.13 CRC carcinogenesis is induced by genetic and epigenetic changes and is influenced by tumor–host interactions.14,15 Many studies demonstrated molecular markers such as BRAF, KRAS, and MMR for CRC.16,17 Previous studies have demonstrated telomere length may be a possible prognostic marker for CRC.18,19 However, the regulation of telomere length by TZAP in CRC is uncertain. Here, we evaluate the association between TZAP expression and survival outcomes in CRC and investigate its molecular mechanism. Additionally, TERT mRNA expression and telomere length were also investigated and their results were compared with public available big data, The Cancer Genome Atlas (TCGA). This comprehensive data may suggest molecular mechanism of TZAP expression in colorectal carcinogenesis.
Patients and Methods
Patients and Tissues
A total of 60 male patients (63.6 ± 10.5 years old, 34–83) were diagnosed as CRC and included in this study. Tissue samples, colorectal adenocarcinomas and paired adjacent non-cancerous tissues were provided from Keimyung Human Bio-resource Bank, Korea. It was obtained from the patients undergoing surgery in Dongsan Medical Center (Daegu, Korea) between April 2008 and January 2010. All patients were explained the study purpose, and informed consent was obtained from each study participant. This study was approved by the Institutional Review Board (Keimyung University Dongsan Medical Center No. 2020-07-027).
DNA Isolation and Telomere Length Analysis
We isolated DNA from samples by QIAamp DNA mini kits (Qiagen, Inc., Valencia, CA, USA). Telomere length was analyzed using qPCR with specific primers for telomere (T) and β-globin (S), based on previous study.13 qPCR was performed by a LightCycler 480 II system (Roche Diagnostics, Basel, Switzerland) with the specific primer described in Table S1 and SYBR GREEN Premix (Toyobo, Japan). Relative telomere length was calculated T/S values using the following formula: T/S = 2−ΔCq, where ΔCq = mean CqT - mean CqS. β-Actin as a housekeeping gene was used for normalization. Each measurement was repeated in triplicate and five serially diluted control samples were included in each experiment.
RNA Isolation and mRNA Expression Analysis
We extracted RNA from tissues by the TRIzol reagent (Molecular Research Center Inc., Cincinnati, OH, USA). RNA quality was measured by Nanodrop 1000 (Thermo Scientific, Wilmington, Denmark). Each cDNA was synthesized from 2 μg of total RNA using M-MLV reverse transcriptase (Promega, Madison, WI, USA). Then, qPCR was performed with a specific primer in Table S1. Further examination was carried out as previously described.
Cell Culture and siRNA Transfection
Human colon cancer cell lines, HCT116 and HT29, were purchased from the Korean Cell Line Bank (KCLB; Seoul, Republic of Korea). They were maintained in RPMI-1640 medium (Corning Incorporated, Corning, NY, USA) with 10% of fetal bovine serum (FBS; HyClone Laboratories, Logan, UT, USA) and 1% of penicillin/streptomycin solution (HyClone Laboratories), in a 37°C with 5% CO2. For MTT and apoptosis assay, cells were plated at 30% density 24 h before transfection. Lipofectamine 2000 (Invitrogen, Carlsbad, CA, USA) was mixed with Opti-MEM (Invitrogen) to adjust the final concentration of 15 nM siRNA, then, the medium was replaced 6 h after transfection. TZAP siRNA duplexes (si-TZAP) were chemically synthesized by Ambion (Austin, TX, USA; siRNA ID# s6566, s6567, s6568). The negative control siRNA (si-NC) that does not target any endogenous transcript was used for control experiments. The sequences of si-NC (Bioneer, Daejeon, South Korea) are as follows: 5′-ACGUGACACGUUCGGAGAA-3′ (sense) and 5′-UUCUCCGAACGUGUCACGU-3′ (antisense).
Cell Viability Assay
Cell viability was analyzed by the MTT reagent as stated in the manufacturer’s protocol (Duchefa Biochemie, Haarlem, Netherlands). Log phase cells were trypsinized into single-cell suspension and HCT116 (2 × 103 cells) and HT29 (1 × 103 cells) were seeded into 96-well plates. After 24 h, cells were transfected as described above. After 96 h, 90 µL of plain media and 10 µL of MTT were added and cultured for 1–2 h in a 37°C humidified incubator. After 2h, 100 µL of dimethyl sulfoxide (DMSO, SIGMA-ALDRICH, St Louis, MO, USA) solution was added into each well. Then, OD value was measured at a wavelength of 540 nm by an Asys UVM 340 microplate reader (Biochrom, Cambridge, UK).
The Cancer Genome Atlas (TCGA) Data Analysis
We investigated the publically available TCGA datasets and their data were downloaded from the TCGA Data Portal at https://tcga-data.nci.nih.gov/tcga/. The microarray and RNA-Seq experiments and clinical data were downloaded directly from the TCGA website in May 2020.
Statistical Analysis
Chi-square, Fisher’s exact test and the Mann–Whitney U-test were used to analyze the association between variables. Univariate survival analysis constructed by the Log rank test with Kaplan–Meier curve. Overall survival was defined as the time between diagnosis and mortality. Disease-free survival was defined as the time between diagnosis, and disease recurrence or the development of distant metastasis. The correlations between telomere length and mRNA expressions and clinicopathologic parameters were assessed with the Pearson’s correlation coefficient analysis. A P-value of <0.05 was considered statistically significant.
Results
Expression Levels of TZAP and TERT mRNA and Telomere Length
TZAP and TERT mRNA expression levels were analyzed in CRC patients. The TZAP and TERT expression were normalized to β-actin. Then, the quantitative PCR (qPCR) data were measured by ∆CT values (Figure 1). TZAP mRNA in CRC was statistically higher than that in paired normal tissue (p = 0.033). However, TERT mRNA and telomere length were similar in CRC and non-cancerous tissues (p = 0.604 and 0.288, respectively). The average of TZAP and TERT expression levels in CRCs were 6.41 ± 1.43 and 4.13 ± 1.09, respectively, as calculated in CRC compared to that of normal tissues. Telomeres in CRC were an average 3.95-fold (SD = 1.39) longer than that in non-cancerous samples.
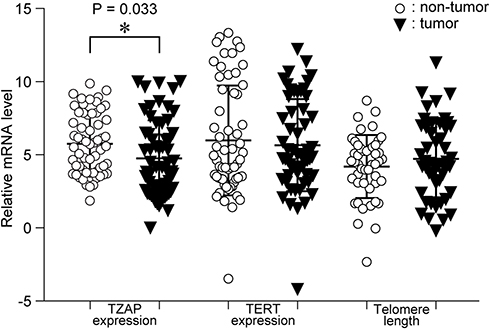 |
Figure 1 mRNA expression levels of TZAP and TERT, and telomere length in CRCs and paired non-cancerous tissues. |
Clinicopathological Characteristics of TZAP and TERT mRNA Expressions and Telomere Length in CRC
To identify the association between these changes and the clinicopathological characteristics, patients were divided into two groups by the average values of the Tumor/Non-tumor ratio. Clinicopathological characteristics of telomere length, TZAP, and TERT expression levels are summarized in Table 1. High TZAP and TERT mRNA levels and telomere shortening were shown in 52.5%, 31.6%, and 43.9% of CRCs, respectively. TZAP and TERT expression levels were different according to T stage; however, it did not achieve statistical significance (p = 0.067 and 0.072, respectively). Higher expression levels of TZAP were found in carcinoembryonic antigen (CEA)-positive CRCs (>5 ng/mL) (p = 0.012). Telomere length was shorter in CRCs with high TZAP expression than in CRCs with low TZAP expression (p = 0.010). Other clinicopathological parameters showed no association with telomere length, TZAP, and TERT mRNA levels.
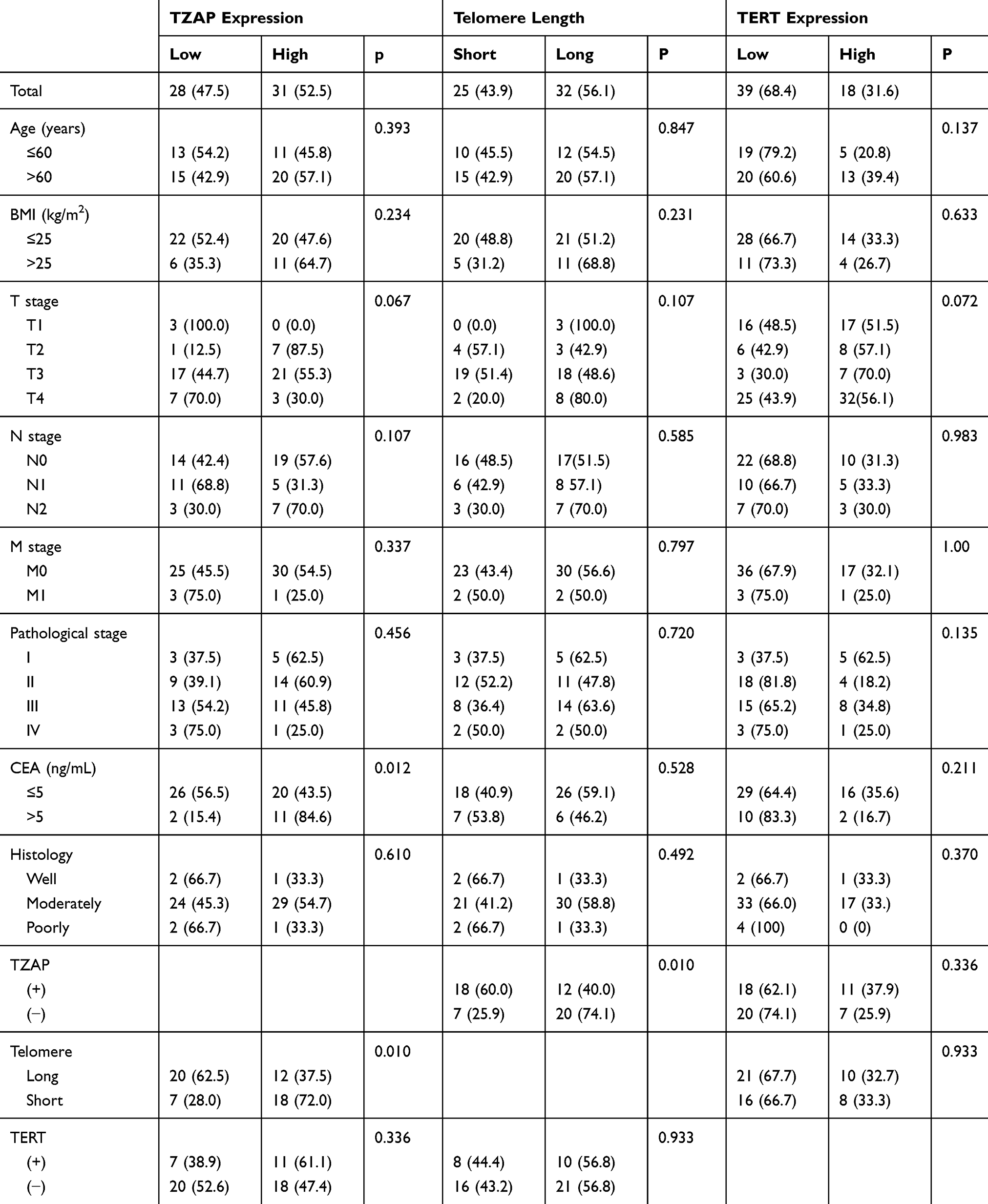 |
Table 1 Clinicopathological Characteristics of TZAP and TERT mRNA Expressions and Telomere Length in CRC |
Quantitative correlation analysis with clinical parameters was performed. The association between age, BMI, CEA, telomere length, TZAP, and TERT is presented in Table 2. Telomere length had no association with age (r = −0.118, p = 0.383). However, TZAP has a negative correlation with age (r = −0.349, p = 0.007, Figure 2A) and telomere length (r = −0.305, p = 0.021, Figure 2B). TZAP and TERT had a weak positive correlation each other (r = 0.279, p = 0.041, Figure 2C). Other parameters had no quantitative correlation with others.
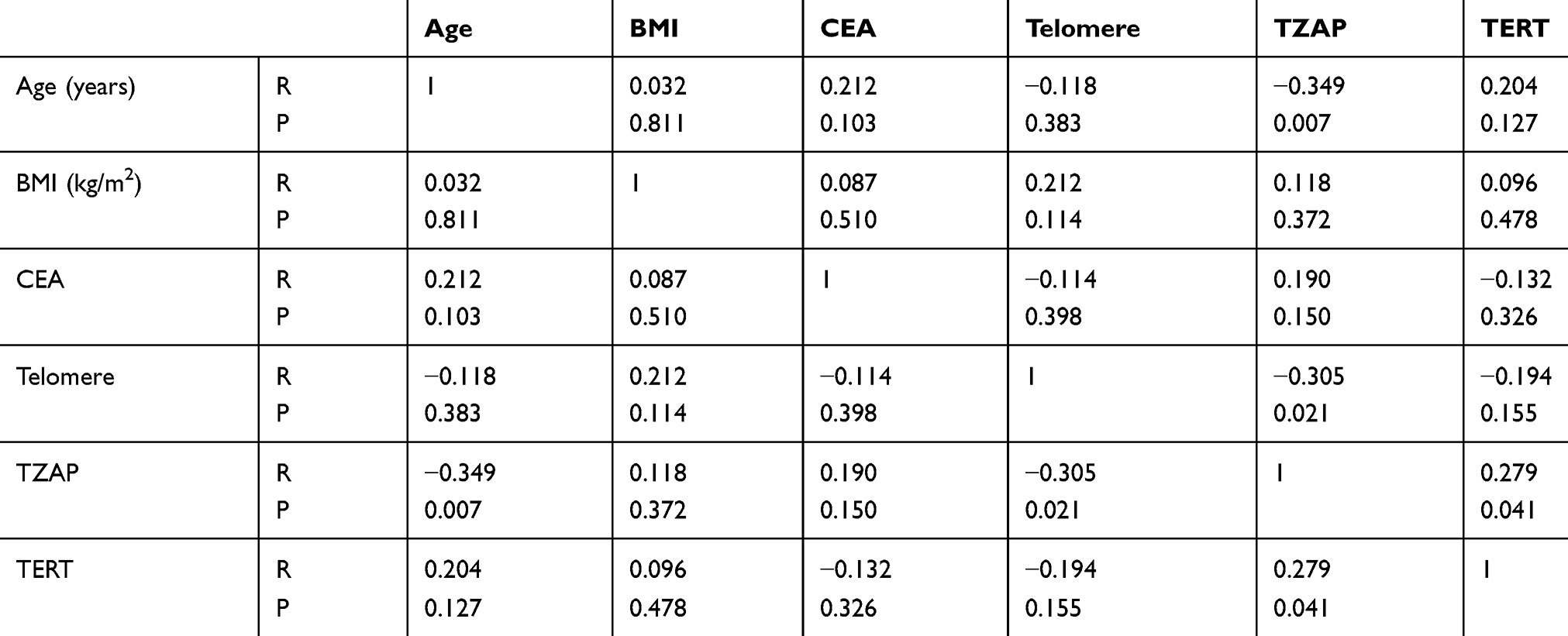 |
Table 2 Correlation Between TZAP Expression and the Clinical Parameters in CRC |
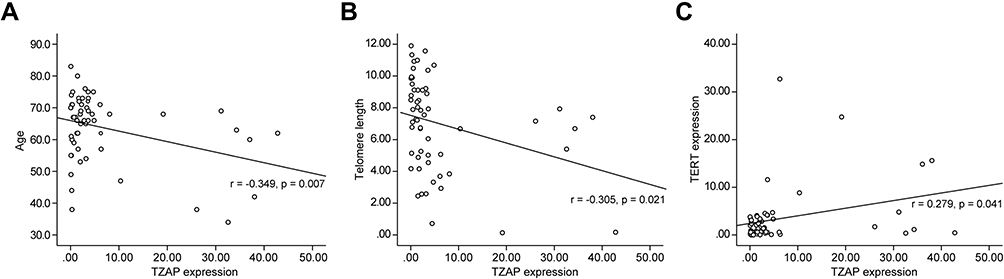 |
Figure 2 Correlation analysis. (A) Between TZAP expression and age. (B) Between TZAP and telomere length. (C) Between TZAP and TERT expressions. |
Prognostic Value of TZAP and TERT mRNA Expressions and Telomere Length in CRC
We evaluated survival analysis of CRC to show prognostic value of TZAP and TERT mRNA expression levels and telomere length. The median follow-up period was 60.2 months (2.5–88.9). Univariate survival analysis showed no significance of TZAP expression for overall survival in CRC (87.3 vs 76.8 months, p = 0.372) (Figure 3A). Disease-free survival was not different according to the TZAP expression (82.8 vs 74.8 months, p = 0.601). TERT expression (overall survival: 78.7 vs 80.0 months, p = 0.967; disease-free survival: 75.7 vs 76.6 months, p = 0.952) and telomere length (overall survival: 83.4 vs 68.8 months, p = 0.305; disease-free survival: 80.7 vs 64.5 months, p = 0.313) did not have any prognostic value in CRCs. When stratified patients by TZAP expression and telomere length, patients with lower TZAP and short telomere length showed poorer prognoses (46.6 vs 83.4 months, p = 0.003, Figure 3B). Other stratification of these markers did not show any significance.
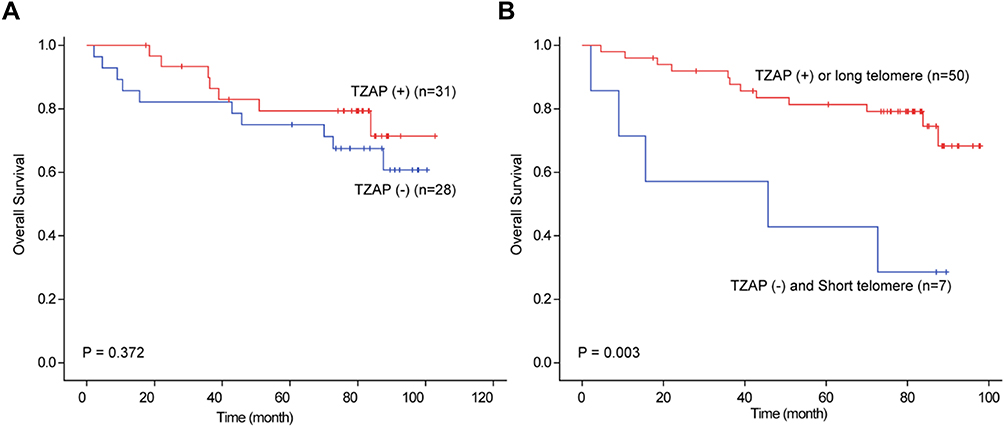 |
Figure 3 Survival analysis in CRCs. (A) Overall survival according to TZAP expression. (B) Overall survival according to TZAP expression and telomere length. |
Effect of TZAP Silencing in Cell Viability and TERT mRNA Expression
To show the effect of TZAP silencing on cell survival, HCT116 and HT29 cells were transfected with a negative control (NC) siRNA or TZAP siRNA. Firstly, we determined the efficacy of TZAP siRNA by qPCR. As shown in Figure 4A, TZAP mRNA was significantly lower in HCT116 and HT29 transfected with TZAP siRNA than cells transfected with NC siRNA. Cell viability of the HCT116 and HT29 cells was assessed by using an MTT assay. The viability of the TZAP-siRNA-treated cells was increased by about 170% in HCT116 cells, when compared to that of the control group. However, in HT29 cells, it was reduced by about 60% (Figure 4B). To investigate the association between TZAP and TERT, mRNA expression of TERT was analyzed in TZAP silencing HCT116 and HT29 cells. TZAP silencing induces a decrease in mRNA levels of TERT in both cells (Figure 4C). These findings suggest that the function of TZAP is related to the TERT pathway in CRC progression. However, telomere length was not significantly changed in TZAP silencing HCT116 (p = 0.48) and HT29 cells (p = 0.73).
 |
Figure 4 siRNA knockdown of TZAP and growth of colorectal cancer cells. (A) Detection of cell viability after transfection in HCT116 and HT29 cells. (B) Detection of TZAP mRNA expression in HCT116 and HT29 cells at 48 h after transfection. (C) Detection of TERT mRNA expression in HCT116 and HT29 cells at 48 h after transfection. Data represent three independent experiments (*P<0.05 and **P<0.01 by Student’s t-test). Abbreviations: si-NC, negative control siRNA; si-TZAP, TZAP-specific siRNA. |
The Cancer Genome Atlas (TCGA) Data
We used the publically available colon and rectal cancer dataset in the TCGA database. In colon cancer, TZAP expression was related with lymphatic invasion (p = 0.039) and TERT expression (p = 0.004) (Table 3). In rectal cancer, TZAP did not have any clinical characteristics; however, TERT expression had an association with T stage (p = 0.017) and lymphatic invasion (p = 0.17) (Table 4). TZAP and TERT expression tended to be correlated; however, it did not get a significance (p = 0.08).
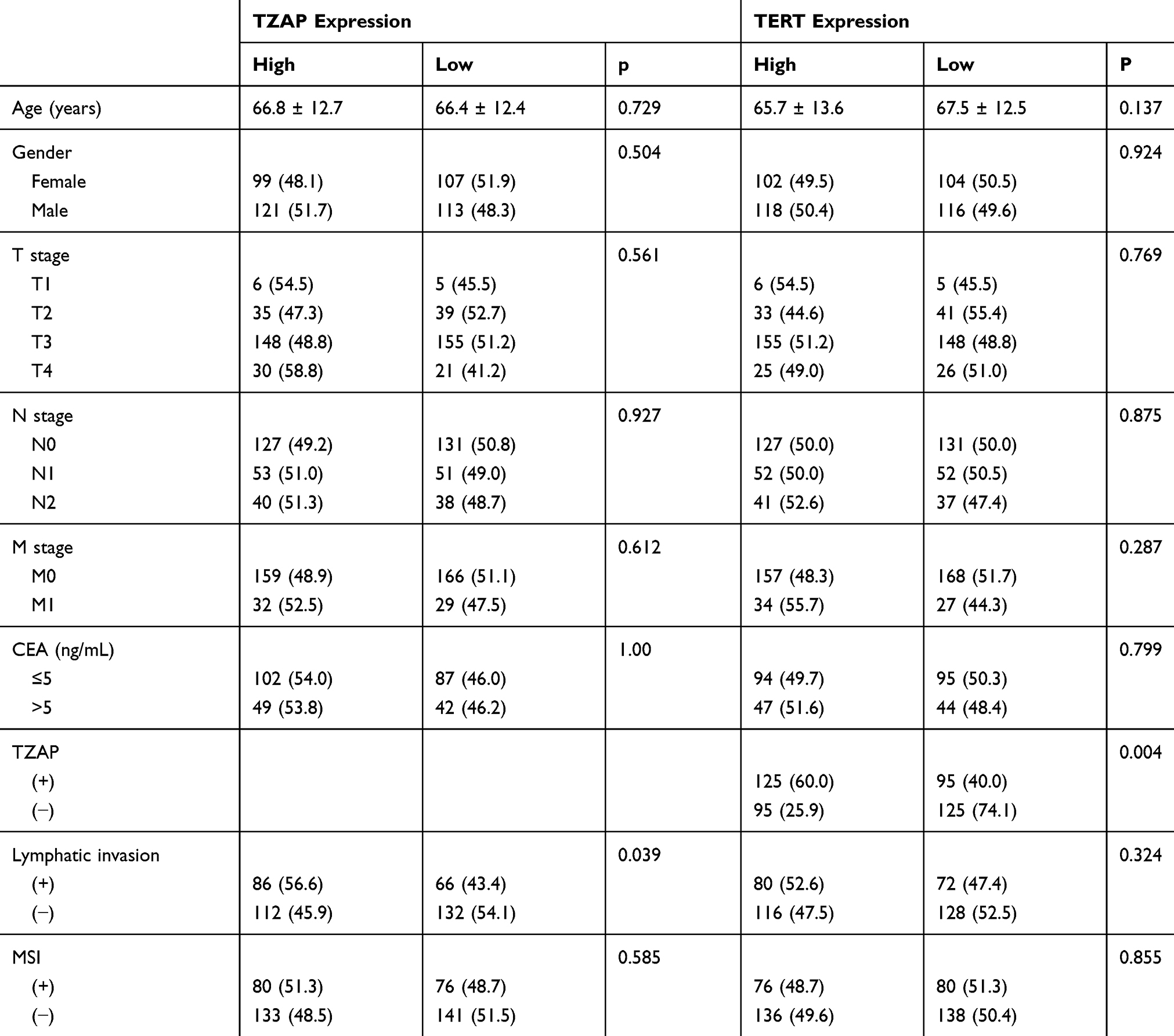 |
Table 3 TCGA Data of Clinicopathological Characteristics of TZAP and TERT mRNA Expressions in Colon Cancer |
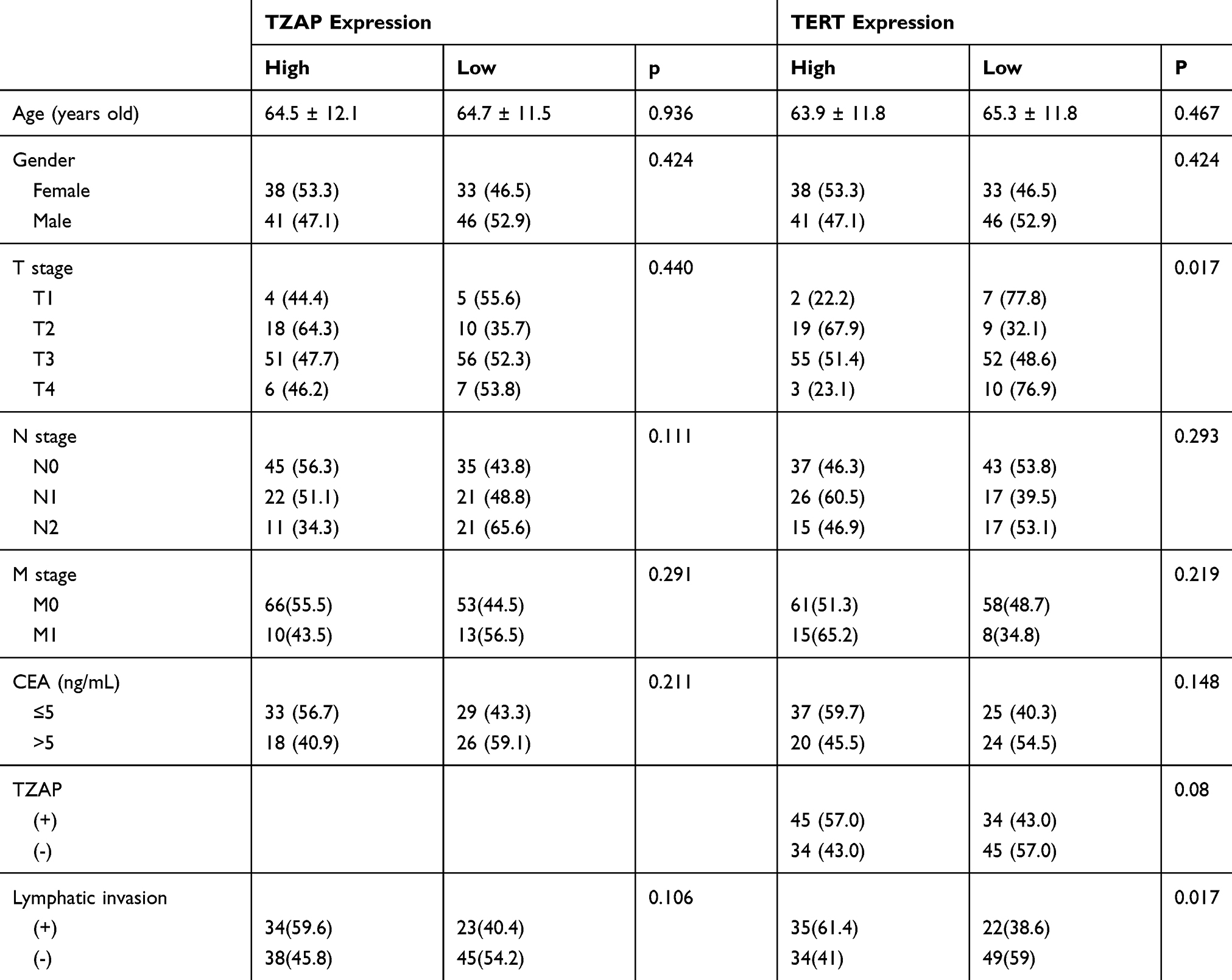 |
Table 4 TCGA Data of Clinicopathological Characteristics of TZAP and TERT mRNA Expressions in Rectal Cancer |
In our previous report, quantitative correlation analysis also showed that TZAP and TERT mRNA levels were positively correlated in colon cancer (r = 0.187, p < 0.001) and rectal cancer (r = 0.186 p = 0.020).20 Survival analysis showed that higher TZAP mRNA levels predicted poorer prognoses in colon cancer (p < 0.001, Figure 5A). However, it did not have any prognostic value in rectal cancer (p = 0.119, Figure 5B).
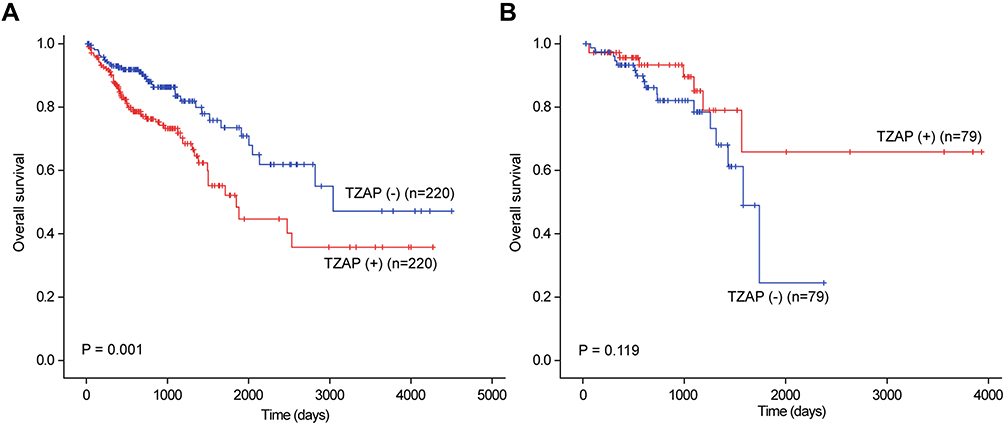 |
Figure 5 TCGA data of TZAP expression (ZBTB48) in colon cancer (A) and rectal cancer (B). |
Discussion
In this study, we showed clinicopathological significance of TZAP mRNA expression in CRC patients, cell lines, and public data for the first time. Previous studies suggested that TZAP expression might prevent telomere elongation occurring during cancer progression.4,9 Therefore, its insufficient expression predicts poorer prognoses in many cancers by TCGA data.21 However, higher TZAP expression was associated with poorer survival, resulting in some cancers, especially CRC. In the present study, we analyze TZAP mRNA expression and telomere length in 60 CRC tissues and paired non-cancerous tissues. TZAP expression was significantly higher in CRCs and it was associated with higher CEA and shorter telomere length. To clarify the role of TZAP in telomere regulation, TERT expression inducing telomere elongation was examined. Quantitative correlation analysis showed an age-dependent in TZAP mRNA levels and telomere shortening. Interestingly, TZAP expression was positively correlated with TERT expression. Our previous study using TCGA data showed this correlation in bladder, colorectal, glioblastoma, head and neck squamous cell carcinoma, hepatocellular carcinoma, lung, melanoma, and uterine endometrial carcinomas.20 To confirm their association, TERT mRNA was analyzed in TZAP-silenced HCT116 and HT29 cells. The results showed that TZAP silencing decreased TERT mRNA expression in both cells. For a first time, this association was confirmed equally in CRC tissues, cell lines, and big data. Although TZAP has been known to compete with TRF1 and TRF2, our results suggested that TZAP and TERT were expressed competitively and proportionally. Unexpectedly, TZAP silencing did not induce the change of telomere length. The paradoxical effect of TZAP and TERT on the regulation of telomere length should be confirmed further.
The public data demonstrated that TZAP mRNA expression in colon cancer was associated with poorer prognoses. It was in agreement with previous hypothesis suggesting telomere shortening via TZAP expression may induce cell death.9 In rectal cancer, TZAP mRNA expression tended to be associated with better survival result; however, it did not have significance. Moreover, TZAP silencing induced cancer cell growth in HCT116 cells; however, it decreased the viability of HT29 cells. Both data and cell line results showed a bipolar effect of TZAP expression; therefore, its pivot factor should be investigated. Considering the genetic difference of HCT116 and HT29 cells, P53 mRNA expression was analyzed. However, there was no significant difference in both cell lines (data not shown). In CRC patients, TZAP did not have prognostic significance; however, the combination of TZAP and telomere length showed powerful prognostic values. We hypothesized that short telomere uncontrolled without TZAP may be important for cancer progression. These controversial results of TZAP expression indicated that many factors might co-operate in cancer progression via telomere regulation. An unknown physiological mechanism may be present in this process. Moreover, TZAP mRNA level and activity of TZAP protein may be different. The molecular mechanism of TZAP should be confirmed further and its clinical value needed to be discussed.
Taken together, we demonstrated, for the first time, clinical and prognostic values of TZAP in CRC. The correlation between TZAP and TERT was consistent in patients, cell lines, and public data, however, some results were different in these data. Therefore, the issue of telomere regulation in cancer therapy remains to be determined. To date, various molecular targets were exploited to trigger cancer cell death via different mechanisms and pathways. TZAP may be associated with telomere regulation and cancer cell survival in CRCs. Therefore, targeting TZAP may be a new avenue for cancer therapy.
Conclusion
In conclusion, we demonstrated the clinical implication of TZAP expression in CRC tissues and cells. Big data results also support the significant role of TZAP expression in cancers. Further studies about its functional mechanism are warranted to validate these results.
Acknowledgments
The biospecimens and data used for this study were provided by the Biobank of Keimyung University Dongsan Hospital Biobank, a member of the Korea Biobank Network.
Funding
This research was supported by grants from the Basic Science Research Program through the National Research Foundation of Korea (NRF), funded by the Korean Government (MSIP), grant number 2014R1A5A2010008.
Disclosure
The authors declare no conflicts of interest.
References
1. Harley CB, Futcher AB, Greider CW. Telomeres shorten during ageing of human fibroblasts. Nature. 1990;345(6274):458–460. doi:10.1038/345458a0
2. Blackburn EH, Greider CW, Szostak JW. Telomeres and telomerase: the path from maize, tetrahymena and yeast to human cancer and aging. Nat Med. 2006;12(10):1133–1138. doi:10.1038/nm1006-1133
3. Xin H, Liu D, Songyang Z. The telosome/shelterin complex and its functions. Genome Biol. 2008;9(9):232. doi:10.1186/gb-2008-9-9-232
4. Shay W, Wright WE. Role of telomeres and telomerase in cancer. Semin Cancer Biol. 2011;21(6):349–353. doi:10.1016/j.semcancer.2011.10.001
5. Jafri MA, Ansari SA, Alqahtani MH, et al. Roles of telomeres and telomerase in cancer, and advances in telomerase-targeted therapies. Genome Med. 2016;8(1):69. doi:10.1186/s13073-016-0324-x
6. Pickett HA, Cesare AJ, Johnston RL, et al. Control of telomere length by a trimming mechanism that involves generation of t-circles. EMBO J. 2009;28(7):799–809. doi:10.1038/emboj.2009.42
7. Pickett HA, Reddel RR. The role of telomere trimming in normal telomere length dynamics. Cell Cycle. 2012;11(7):1309–1315. doi:10.4161/cc.19632
8. Sfeir A, de Lange T. Removal of shelterin reveals the telomere end-protection problem. Science. 2012;336(6081):593–597. doi:10.1126/science.1218498
9. Li JS, Miralles FJ, Simavorian T, et al. TZAP: a telomere-associated protein involved in telomere length control. Science. 2017;355(6325):638–641. doi:10.1126/science.aah6752
10. Maris JM, Jensen SJ, Sulman EP, et al. Cloning, chromosomal localization, physical mapping, and genomic characterization of HKR3. Genomics. 1996;35(2):289–298. doi:10.1006/geno.1996.0359
11. White PS, Maris JM, Sulman EP, et al. Molecular analysis of the region of distal 1p commonly deleted in neuroblastoma. Eur J Cancer. 1997;33(12):1957–1961. doi:10.1016/S0959-8049(97)00311-0
12. Maris JM, Jensen J, Sulman EP, et al. Human Krüppel-related 3 (HKR3): a candidate for the 1p36 neuroblastoma tumour suppressor gene? Eur J Cancer. 1997;33(12):1991–1996. doi:10.1016/S0959-8049(97)00279-7
13. Shin A, Kim KZ, Jung KW, et al. Increasing trend of colorectal cancer incidence in Korea, 1999–2009. Cancer Res Treat. 2012;44(4):219–226. doi:10.4143/crt.2012.44.4.219
14. Arnold M, Sierra MS, Laversanne M, et al. Global patterns and trends in colorectal cancer incidence and mortality. Gut. 2017;66(4):683–691. doi:10.1136/gutjnl-2015-310912
15. Colussi D, Brandi G, Bazzoli F, et al. Molecular pathways involved in colorectal cancer: implications for disease behavior and prevention. Int J Mol Sci. 2013;14:16365–16385.
16. Di Caro G, Marchesi F, Laghi L, et al. Immune cells: plastic players along colorectal cancer progression. J Cell Mol Med. 2013;17(9):1088–1095. doi:10.1111/jcmm.12117
17. Ogino S, Galon J, Fuchs CS, et al. Cancer immunology–analysis of host and tumor factors for personalized medicine. Nat Rev Clin Oncol. 2011;8(12):711–719. doi:10.1038/nrclinonc.2011.122
18. Kroupa M, Rachakonda SK, Liska V. Relationship of telomere length in colorectal cancer patients with cancer phenotype and patient prognosis. Br J Cancer. 2019;121(4):344–350. doi:10.1038/s41416-019-0525-3
19. Nersisyan L, Hopp L, Loeffler-Wirth H, et al. Telomere length maintenance and its transcriptional regulation in lynch syndrome and sporadic colorectal carcinoma. Front Oncol. 2019;9:1172. doi:10.3389/fonc.2019.01172
20. Heo YR, Park WJ, Lee JH. Positive correlation between TZAP and TERT in most cancers: a new player in cancer diseases. Ann Transl Med. 2018;6(10):197. doi:10.21037/atm.2018.03.14
21. Park WJ, Heo YR, Lee JH, et al. Prognostic value of TZAP expression in various cancers: TCGA data analysis. Keimyung Med J. 2018;37:17–21.
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.