")
Back to Journals » Infection and Drug Resistance » Volume 17
Clinical Characteristics and Drug Resistance Mechanisms of Linezolid-Non-Susceptible Enterococcus in a Tertiary Hospital in Northwest China
Authors Wu M, Kang J , Tao J, Yang Y, Li G, Jia W
Received 25 October 2023
Accepted for publication 25 January 2024
Published 8 February 2024 Volume 2024:17 Pages 485—494
DOI https://doi.org/10.2147/IDR.S442105
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Sandip Patil
Mengying Wu,1 Jia Kang,1 Jia Tao,1 Yanwen Yang,2 Gang Li,1 Wei Jia1,3
1Medical Experimental Center, General Hospital of Ningxia Medical University, Yinchuan, People’s Republic of China; 2Department of Neurology, General Hospital of Ningxia Medical University, Yinchuan, People’s Republic of China; 3Ningxia Key Laboratory of Clinical and Pathogenic Microbiology, General Hospital of Ningxia Medical University, Yinchuan, People’s Republic of China
Correspondence: Wei Jia, Medical Experimental Center, General Hospital of Ningxia Medical University, Yinchuan, 750001, People’s Republic of China, Tel +86 951-6744013, Email [email protected]
Purpose: To understand the detection rate and distribution characteristics of Linezolid-nonsusceptible Enterococcus (LNSE) and analyze the molecular typing and main drug resistance mechanisms of LNSE, providing a theoretical basis for the precision prevention and control of LNSE hospital infections.
Methods: A total of 40 LNSE strains isolated from clinical specimens between January 1, 2012, and December 31, 2022, were collected. The LNSE isolates identified by instrument detection were confirmed using a microbroth dilution method. The WHONET 5.0 software was used for statistical analysis of LNSE detection rate, and the LNSE judgment was based on the 2022 CLSI criteria. PCR methods were used to detect 23S rRNA, cfr, optrA, and L3, L4 ribosomal RNA sites for linezolid resistance genes, and gene sequencing was used to verify the amplified PCR products. Multiple locus sequence typing (MLST) was performed to analyze the homology of LNSE strains.
Results: A total of 6924 Enterococcus isolates were separated and identified from January 1, 2012, to December 31, 2022, of which 40 were LNSE strains (26 Enterococcus faecalis, 14 Enterococcus faecium), with a detection rate of 0.58% (40/6924). Among them, 28 Linezolid-intermediated Enterococcus(LIE) were detected, accounting for 0.4% (28/6924), and 12 Linezolid-resistant Enterococcus(LRE) were detected, with a detection rate of 0.17% (12/6924). Among the LNSE strains, 23 were resistant to genes. The 40 LNSE strains could be divided into 20 different ST types, with ST16 being the main type, accounting for 12.5% (5/40).
Conclusion: The detection of LNSE strains was dominated by Enterococcus faecalis, and the main resistance mechanism of LRE strains was carrying the optrA gene, with 23S rRNA gene mutations also contributing to resistance. New resistance gene phenotypes (optrA +/23S rRNA+) emerged. Most LRE cases were sporadic, and clonal dissemination was observed in some strains.
Keywords: linezolid-nonsusceptible Enterococcus, PCR, MLST, ST16, optrA
Introduction
Enterococcus is a normal flora in the intestines of humans and animals and a common conditionally pathogenic bacterium causing hospital-acquired infections, which can lead to various diseases such as urinary tract infections, sepsis, and endocarditis.1 Due to the inherent and acquired resistance characteristics of Enterococcus, as well as the widespread use of various antibiotics in clinical practice, multidrug-resistant Enterococcus has increased, and even vancomycin-resistant Enterococcus has appeared. Linezolid is the first Oxazolidinone antibiotics used in clinical practice, which binds to the peptidyl transferase center of the bacterial ribosome 50s subunit, inhibiting the formation of the 70s initiation complex and interfering with protein synthesis to exert bacteriostatic effects.2 Since linezolid has a good treatment effect for most infections caused by Gram-positive bacteria, it is the last line of defense for clinical Staphylococcus aureus resistant to methicillin, vancomycin-resistant Enterococcus, and multidrug-resistant Streptococcus pneumoniae. However, with the extensive use of linezolid in clinical practice, linezolid-resistant Enterococcus continues to emerge and increase year by year.
In Germany, the detection rate of linezolid-resistant Enterococcus faecium increased from less than 1% in 2008 to more than 9% in 2014.3 According to the China Antimicrobial Surveillance Network (CHINET) monitoring data, the resistance rate of Enterococcus faecalis to linezolid increased from 1.9% in 2018 to 3.5% in 2021. In 2017, a report from South Korea found that among 7720 Enterococcus faecalis and 3939 Enterococcus faecium from animal or animal carcass sources from 2003 to 2014, 12 (0.16%) and 27 (0.69%) were resistant to oxazolidinone, respectively. Among the resistant strains, 89.74% carried genes, 17.9% had mutations in the L4 protein of the ribosome, and no mutations were observed in the 23SrDNA gene.4 The LEADER antimicrobial resistance testing organization analyzed the susceptibility of 54,911 Gram-positive cocci collected from 60 medical research centers in the United States from 2004 to 2012. The results showed that the non-susceptibility rate (including resistance and intermediate) of Enterococcus to linezolid ranged from 0.34% to 1.83%, and the overall low-level resistance (MIC 4–8 μg/mL) was presented.5 The Zyvox (R) Annual Appraisal of Potency and Spectrum (ZAAPS) resistance monitoring study, which included 77 medical microbiological testing centers in 33 countries and collected 52,082 Gram-positive cocci worldwide, showed that the non-susceptibility rate of linezolid-resistant Enterococcus increased from 0.0% in 2004 to 0.8% in 2012. The three isolates of linezolid-resistant Enterococcus faecalis were all optrA-positive carriers.6 In 2016, 8325 Gram-positive isolates from 42 countries and 76 centers (excluding the United States) were found to have 8 linezolid-resistant Enterococcus faecalis (LREfs) and 2 linezolid-resistant Enterococcus faecium (LREfm). Enterococcus faecalis were optrA-positive carriers, and the 2 Enterococcus faecium isolates had G2576T mutation.7 Currently, most linezolid-resistant Enterococcus faecalis are isolated from Europe and Asia, while linezolid-resistant Enterococcus faecium are mainly isolated from Europe and North America.8
The increasing resistance of Enterococcus to linezolid requires urgent attention to solve this resistance problem. In addition to the main 23S rRNA gene mutations, acquired resistance genes associated with mobile genetic elements (MGE), such as cfr, cfr (B), optrA, and poxtA, also play an important role in the resistance of Enterococcus to linezolid.9 The oxazolidinone resistance genes have been found on a variety of MGEs, including plasmids, transposons, ICEs, prophages, and genomic islands, in various bacteria. These MGEs, but also IS-mediated, oxazolidinone resistance gene-carrying TUs, which can integrate into the aforementioned MGEs, play an important role in the spread of cfr, optrA, and poxtA genes across not only strain, species, and genus, but also family and order borders. In addition, these MGEs often carry additional resistance genes which support the coselection and persistence of the oxazolidinone resistance genes.10 Currently, several mechanisms of resistance, or reduced susceptibility, to oxazolidinones have been identified. They can be summarized as follows: (i) ribosomal mutations in 23S rRNA and/or in L3 and/or L4 ribosomal proteins; (ii) loss of the rlmN gene activity; (iii) active efflux; and (iv) transferable mechanisms including the Cfr and Cfr-like methyltransferases and the ABC-F proteins OptrA, PoxtA and PoxtA2.11 The presence and spread of resistance genes not only accelerate the development of bacterial resistance but also increase the difficulty of treating Enterococcus-related infectious diseases. Bacterial typing can explore the clonal relationships between different strains, determine the relationship between pathogens and infection sources, and use it to control the epidemic and cross-infection of pathogens. This study confirmed and analyzed LNSE strains isolated from General Hospital of Ningxia Medical University in China from January 1, 2012, to December 31, 2022, to understand the detection rate and resistance level of LNSE strains. PCR and sequencing methods were used to detect 23S rRNA, cfr, optrA, and L3, L4 ribosomal sites linezolid resistance genes to understand the main resistance genes of LNSE. To investigate the clonal relatedness of LNSE strains, multilocus sequence typing (MLST) was used to analyze the phylogenetic relationships between LNSE strains.
Materials and Methods
Sample Collection and Reference Strains
Clinical strains were collected from January 1, 2012, to December 31, 2022, at General Hospital of Ningxia Medical University. These include Linezolid-nonsusceptible Enterococcus (LNSE) and Linezolid-susceptible Enterococcus (LSE), all of which are non-repeat isolates. All participants have signed the informed consent form. Quality control strain: The standard strain of Enterococcus faecalis ATCC 29212 is preserved in our laboratory.
Main Reagents and Instruments
Vitek 2 Compact and matching GP identification cards, GP67 antimicrobial susceptibility cards (Biomerieux, France); 2×HieffTM PCR Master Mix (Nanjing Novozymes); PCR primers synthesized by Shanghai Shenggong; molecular weight DL2000 marker (Nanjing Novozymes); Genered nucleic acid dye (Biotium, USA); 50×TAE buffer (Boao Tuoda); MALDI-TOF bacterial mass spectrometry identification instrument (Biomerieux, France); PowerPac horizontal gel electrophoresis instrument (Bio-Rad, USA); Gel Doc XK gel imaging system (Bio-Rad, USA).
Drug Susceptibility Test
From January 2012 to December 2022, conventional bacterial cultivation methods were used for clinical microbial specimens, and the identification of Enterococcus species was performed using Vitek 2 Compact and MALDI-TOF. The routine drug susceptibility test of Enterococcus and its minimum inhibitory concentration (MIC) for linezolid were detected using the Gram-positive drug susceptibility card; the instrument-detected LNSE strains were confirmed using the microbroth dilution method recommended by CLSI M100 (M100, 30th).
Isolation of Bacterial DNA
Pick a pure culture colony and place it in a 0.5mL centrifuge tube, incubate at 100 °C for 10 minutes, and centrifuge at 13,000 × g for 30 seconds. The supernatant is the template solution for gene detection, stored at −20 °C for future use.
PCR Amplification and Sequencing
Adopting PCR method to amplify the gene of 23S rRNA domain V, cfr gene, optrA gene and L3, L4 ribosomal sites, synthesize primers according to literature,12,13 the primer pairs in this work are showed in Table 1. The reaction system is a total of 25 μL, including 2×HieffTM PCR Master Mix 12.5 μL, DNA template 2 μL, forward and reverse primers (10 μmol /L) each 1μL, and ddH2O 8.5 μL. PCR reaction conditions: 94 °C for 5 minutes, 94 °C for 30 seconds, 58°C for 45 seconds, 72 °C for 45 seconds, 30 cycles, 72 °C for 7 minutes. PCR products were subjected to 15 g/L agarose gel electrophoresis, positive amplification products were sent to Shanghai Shenggong Company for bidirectional sequencing, and sequencing results were queried on GenBank. And compare the sequence with the control strain ATCC29212, which is sensitive to linezolid.
 |
Table 1 PCR Primers Sequences and Product Sizes for Target Genes |
LNSE Homology Analysis
We conducted a homology analysis of LNSE strains using MLST. We referred to the seven endogenous housekeeping gene sequences from the MLST website (https://pubmlst.org/). PCR reaction matrix is the same as mentioned above; Enterococcus faecalis(efa)-MLST PCR reaction conditions: 94°C for 5 minutes, 94°C for 30 seconds, 52°C for 30 seconds, 72°C for 1 minute, 30 cycles, and 72°C for 7 minutes; Enterococcus faecium(efm)-MLST PCR reaction conditions: 95°C for 15 minutes, 94°C for 30 seconds, 50°C for 30 seconds, 72°C for 30 minutes, 35 cycles, and 72°C for 5 minutes. The PCR amplification products were sent to Shanghai Shenggong Company for sequencing. The BioNumerics software was used to construct a minimum spanning tree. This software is a platform developed by the Belgian Applied Maths company for managing, storing, and statistical analysis of all types of biological data. By importing the table file of MLST housekeeping gene results, we analyzed the phylogenetic relationships between different ST types.
Results
The Ratio of Enterococcus Species in the Population
From January 1, 2012 to December 31, 2022, our hospital detected a total of 6924 Enterococcus strains, among which 4162 were Enterococcus faecium, accounting for 60.1%; 1826 were Enterococcus faecalis, accounting for 26.37%; 275 were Enterococcus gallinarum, accounting for 3.97%; 258 were Enterococcus plumboflavus, accounting for 3.73%; 152 were Enterococcus avium, accounting for 2.2%; 104 were Enterococcus raffinosus, accounting for 1.5%; 89 were Enterococcus hirae, accounting for 1.29%; and 58 were other Enterococcus species, accounting for 0.84%.
Drug Susceptibility Analysis of LNSE Strain
From January 1, 2012 to December 31, 2022, a total of 6924 strains of Enterococcus were isolated and identified, of which 6884 were susceptible to linezolid. Among them, 40 strains of LNSE were detected by both instrumental and microbroth dilution methods (26 Enterococcus faecalis and 14 Enterococcus faecium), with a detection rate of 0.58% (40/6924). Among them, 28 strains of LIE were detected, with a detection rate of 0.4% (28/6924), and 12 strains of LRE were detected, with a detection rate of 0.17% (12/6924). The overall resistance rate of Enterococcus faecalis to linezolid is 0.4% (8/1826), and the overall resistance rate of Enterococcus faecium to linezolid is 0.1% (4/4162). Twelve LRE strains were isolated, with MIC values of 8–32μg/mL. LNSE strains identified in the study often exhibit multiple drug-resistant phenotypes (insensitive to at least one drug among three antimicrobial agents), with resistance rates of >40% to ciprofloxacin, levofloxacin, high-level gentamicin, erythromycin, and tetracycline, all strains are susceptible to teicoplanin, and a resistance rate of <4% to vancomycin (Tables 2–4).
 |
Table 2 Linezolid Susceptibility Results (MIC Values) Against LRE Strains by Instrumental Method and Broth Dilution Method |
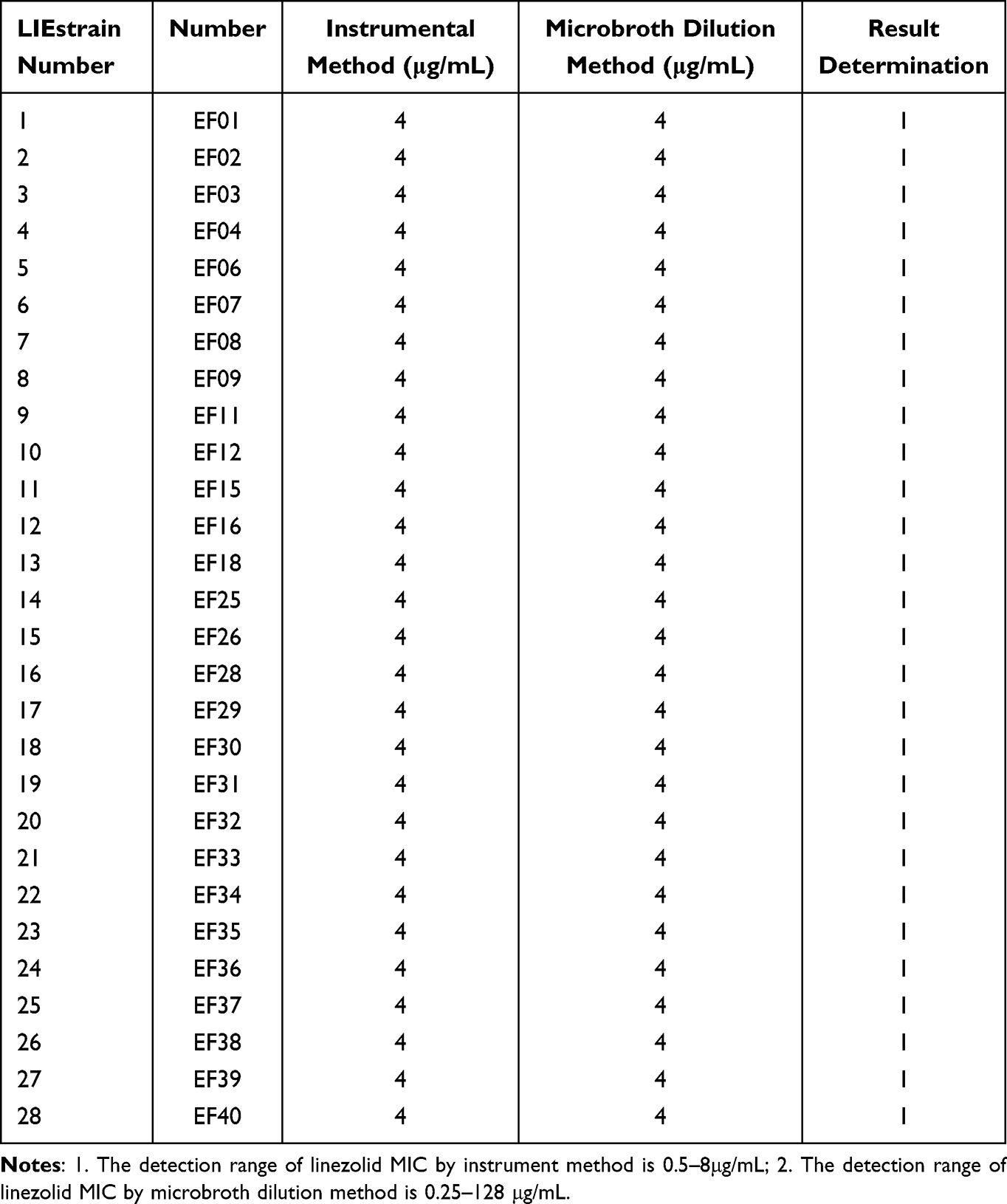 |
Table 3 Linezolid Susceptibility Results (MIC Values) Against LIE Strains by Instrument Method and Broth Dilution Method |
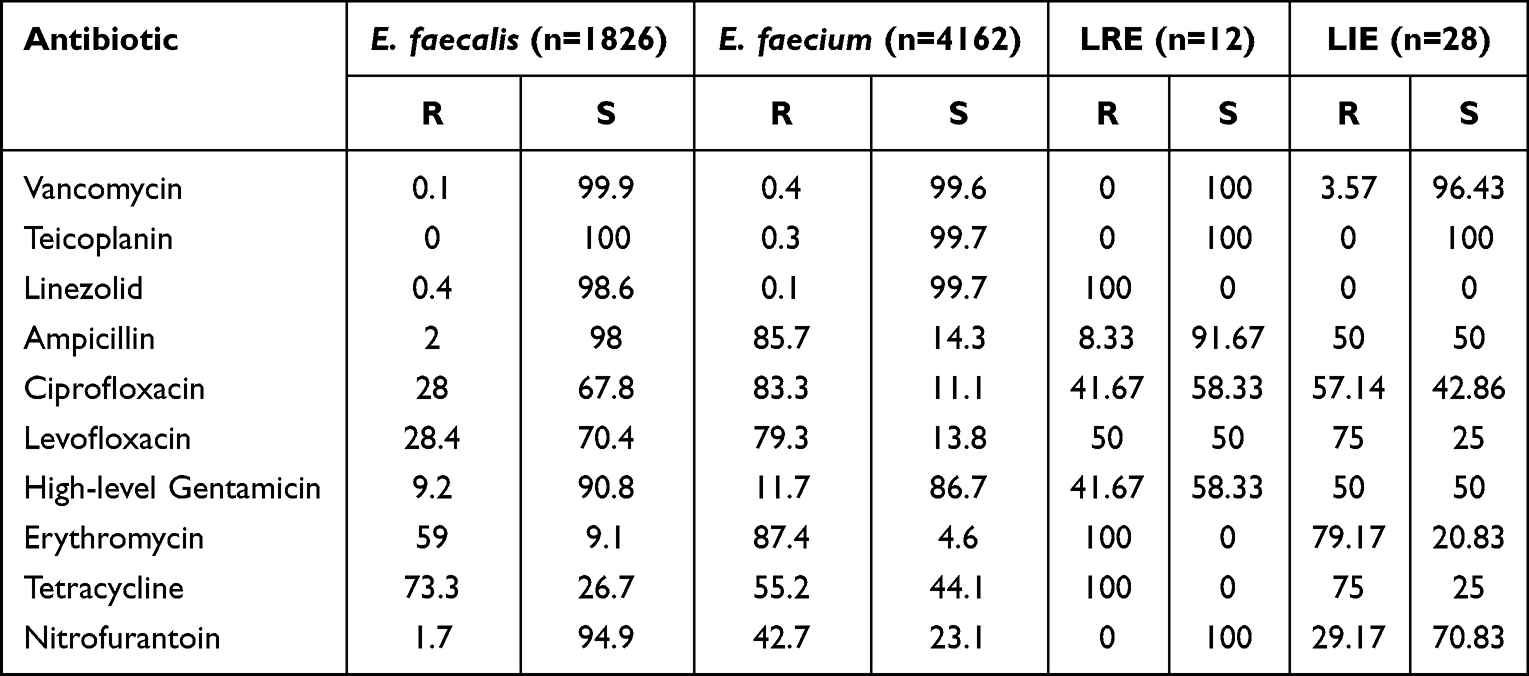 |
Table 4 Resistance and Sensitivity Rate (%) to Antibiotics Among Enterococcus Strains |
The Department and Specimen Type Distribution of LNSE Strain
The department distribution of LNSE strain is as follows: hepatobiliary surgery accounts for 25% (10/40), emergency department accounts for 17.5% (7/40), intensive care unit accounts for 10% (4/40), and the rest are urology, gastrointestinal surgery, pediatric surgery, cardiology and gynecology, etc. In terms of specimen types, urine accounts for 35% (14/40), bile accounts for 20% (8/40), secretions account for 17.5% (7/40), blood accounts for 7.5% (3/40), drainage fluid accounts for 7.5% (3/40), and the rest account for 12.5% (5/40).
PCR Amplification results
Genes related to resistance to linezolid, such as 23S rRNA, cfr, L3, L4, and optrA, were selected for detection. It was found that 23 LNSE strains were positive for oxazolidinone resistance genes, which could be divided into two categories: single drug resistance genes and double drug resistance genes. Single drug resistance genes: optrA gene positive for 17 strains, 23S rRNA gene mutation for 14 strains. Double drug resistance genes: simultaneous carriage of optrA gene and 23S rRNA gene for 8 strains. The genes encoding L3 ribosomal protein rplC and L4 ribosomal protein rplD, as well as the cfr gene, did not mutate, and 12 LNSE strains without any resistance genes were found. Electrophoresis analysis charts are shown in Figure 1A and B, as well as Figure 2A and B.
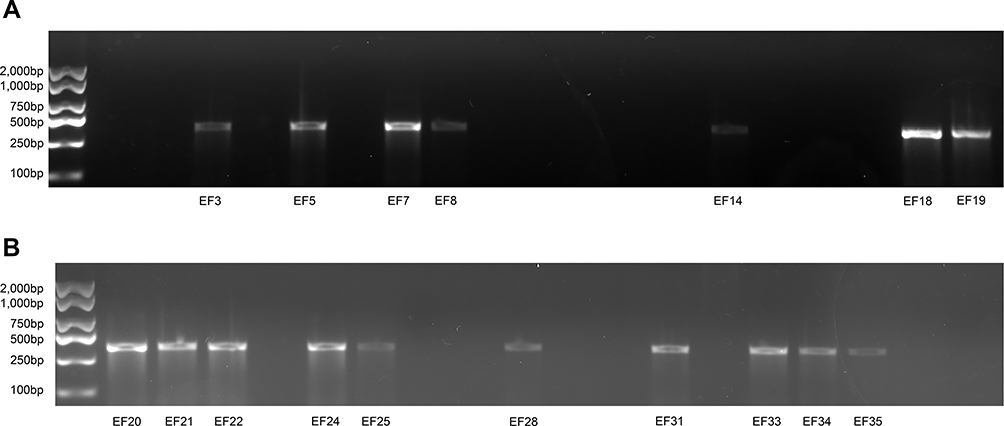 |
Figure 1 (A) electrophoresis-analysis-of-optra-resistance-gene-in-40-LNSE-strains. (B) electrophoresis-analysis-of-optra-resistance-gene-in-40-LNSE-strains. |
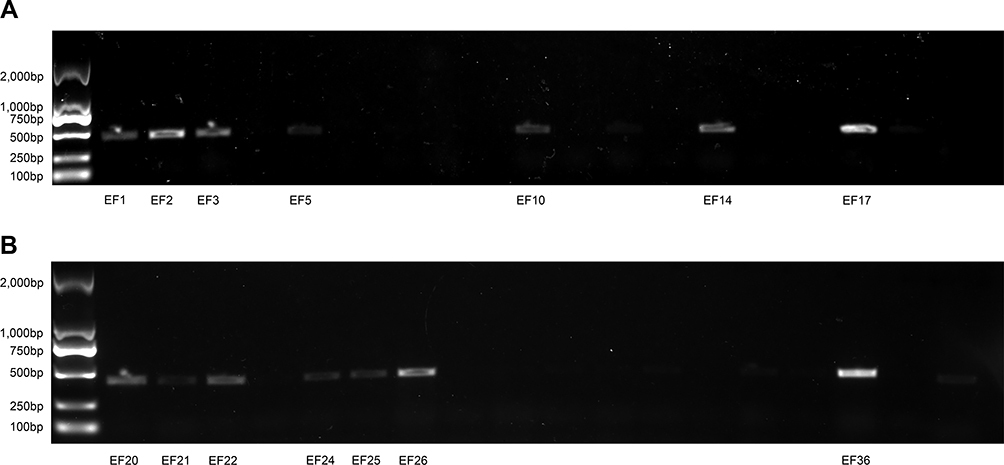 |
Figure 2 (A) electrophoresis-analysis-of-23srna-resistance-gene-in-40-LNSE-strains. (B) electrophoresis-analysis-of-23srna-resistance-gene-in-40-LNSE-strains. |
MLST Analysis of LNSE Strain
The seven housekeeping genes (gdh, gyd, pastS, gki, aroE, xpt, yqiL) of 26 Enterococcus faecalis strains among the 40 LNSE strains and the seven housekeeping genes (adk, atpA, ddl, gdh, gyd, purK, pstS) of 14 Enterococcus faecium strains among the 40 LNSE strains were uploaded to the MLST official website for comparison. In total, 20 ST types were found, among the Enterococcus faecalis, 5 strains of ST16 type were found, 3 strains of ST25 type, and 2 strains of ST506 type, the others are ST969, ST954, ST300, ST330, ST480, ST179, ST714, ST191, ST217, ST241, ST436, ST65, ST256, ST475, and ST1184. Only 2 strains of ST78 and 1 strain of ST17 were found in the Enterococcus faecium. The A6 strain of Enterococcus faecalis and the B1, B2, B3, B7, B10, B12, B13, and B14 strains of Enterococcus faecium were not found in the website, which may indicate new ST types and further verification and confirmation are needed. New alleles that must be submitted to the pubMLST platform to obtain a new sequence type. The analysis results are shown in Figures 3 and 4.
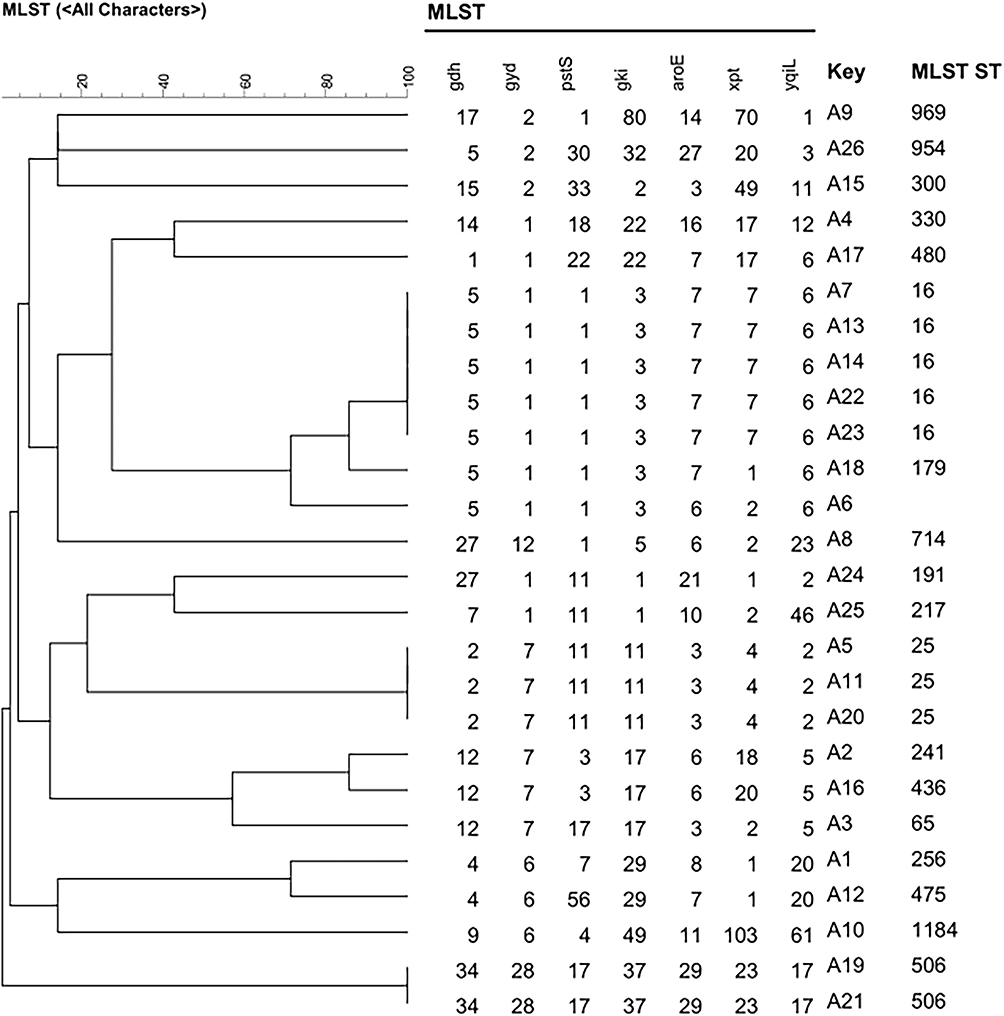 |
Figure 3 MLST phylogenetic tree of 26 Enterococcus faecalis strains among 40 LNSE strains. |
 |
Figure 4 MLST phylogenetic tree of 14 Enterococcus faecium strains among 40 LNSE strains. |
Discussion
Enterococcus is an important Gram-positive bacterium responsible for hospital infections after Staphylococcus aureus. Their cell walls are firm and thick, and they often appear as multi-drug resistant bacteria due to their biological characteristics. Linezolid is the main oxazolidinone antibiotic used in clinical practice for the treatment of severe infections caused by multidrug-resistant Gram-positive bacteria.14 Currently, the vast majority of Enterococcus strains are highly sensitive to linezolid, but there have been reports of linezolid-resistant Enterococcus isolates worldwide, including in China. The official website of the CHINET shows that the resistance rate of Enterococcus faecalis to linezolid in 2018 was 1.9%.15 The total resistance rate of Enterococcus faecalis in our hospital from 2012 to 2021 was 0.4%, which is lower than the data of the 2018 CHINET. Among the 40 LNSE strains isolated clinically, 12 were LRE strains and 28 were LIE strains. The MIC values of the 12 LRE strains ranged from 8–32μg/mL. Some literature suggests that an MIC range of 8–16mg/L should be defined as low-level resistance, and MIC > 64mg/L should be defined as high-level resistance.16 Low-level resistance may pave the way for the development of higher-level resistance, which may lead to the occurrence of “superbugs” in hospitals.17 Therefore, we conducted molecular typing and main resistance mechanism research on LRE strains in our hospital.
The 40 LNSE strains were mainly distributed in departments such as hepatobiliary surgery, emergency department, and intensive care unit, and the specimen sources were mainly urine, bile, secretions, blood, and drainage fluid. MLST analysis showed that the 40 LNSE strains could be divided into 20 different ST types. ST 16 type efa was the most common strain in this study (5/40), mainly distributed in hepatobiliary surgery, suggesting the existence of ST16 type-dominated multiclonal dissemination in hepatobiliary surgery. Other ST type strains were relatively scattered and distributed in various departments, showing sporadic epidemics and no clonal dissemination within the hospital.
The V region point mutations in 23S rRNA are the most common resistance mechanism for Linezolid-resistant Enterococcus faecalis strains, with the main mutation types being G2576T, followed by G2247T and G2505A. In addition, mutations have also been found at G2576U, T2500A, G2192T, G2447T, C2461T, A2503G, T2504C, G2505A, G2766T, G2512U, G2513U, and C2610G sites.18 In 2015, Wang et al19 first reported the optrA resistance gene in Enterococcus isolated from human and animal sources in China, which was the second transferable LZD resistance gene reported after the cfr gene. The optrA gene encodes a ABC transporter protein ATP-binding cassette F (ABC-F) family protein, belonging to the ABC family. This ABC transporter protein confers resistance to the oxazolidinones (linezolid and tedizolid) and amidol (chloramphenicol and fluphenicol) through ribosome protection, and this resistance gene can be easily transferred between Enterococcus strains.19–21 The results of this study showed that 17 strains (42.5%) were positive for the optrA gene, among which 7 were LRE and 10 were LIE. Fourteen strains had gene mutations in 23S rRNA, eight of which were LRE and six were LIE. Eight strains (EF03, EF05, EF14, EF20, EF21, EF22, EF24, EF25) carried both the optrA gene and the 23S rRNA gene, among which seven were LRE and one was LIE. This phenomenon indicates that co-carrying the optrA gene and the 23S rRNA gene is the main resistance mechanism for LRE strains in our hospital. Further conventional and continuous screening for optrA gene risk factors is needed, as these factors may be related to the rapid development and spread of linezolid resistance.
The cfr gene encodes a ribosomal RNA (rRNA) methyltransferase that can catalyze the methylation of C8 at position 2503 in the 23S rRNA V region, indirectly affecting the binding of specific antibiotics and resulting in resistance. It is a multidrug resistance gene, leading to resistance to oxazolidinone, chloramphenicol, lincomycin and streptogramin and decreased susceptibility to macrolide classes.22 Since the first report in 2012 of plasmid-mediated cfr gene in human Enterococcus faecalis (ST16) leading to resistance against linezolid,23 there have been several similar case reports.19,24–26 Mutations in ribosomal proteins L3, L4, or L22 may be another mechanism causing non-transferable resistance to linezolid. Studies have shown that alterations in the conserved structural domains of ribosomal proteins L3 and L4 encoded by the rplC and rplD genes are associated with non-susceptibility to linezolid in Enterococcus (MICs 4–8μg/mL).27 No mutations in the rplC gene encoding the L3 ribosomal protein, the rplD gene encoding the L4 ribosomal protein, or the cfr gene were detected in this study. Twelve strains were found to have no resistant genes. The formation of bacterial biofilms is considered to enhance bacterial resistance to antimicrobial agents in terms of bacterial adaptive resistance. Therefore, it cannot be ruled out that biofilm formation may contribute to the reduced susceptibility to linezolid. Whether biofilms participated in the formation of linezolid resistance in this study’s LNSE strain remains to be further investigated. The sample size of this study’s LRE is small, and further experiments will continue to collect more samples and further explore the mechanism of drug resistance and improve it.
Conclusion
The detection rate of LNSE strain in our hospital is 0.58%, mainly Enterococcus faecalis, mainly distributed in hepatobiliary surgery, emergency department and intensive care unit, with higher detection rate. At the same time, carrying optrA gene and 23S rRNA gene is the main resistance mechanism for LRE strain in our hospital. Epidemiological investigation showed that most LRE cases are sporadic, and some strains have clonal dissemination. The spread of LRE strain in hepatobiliary surgery is mainly caused by SST16 type strain clonal dissemination.
Ethics Approval
The research protocol has been approved by the Medical Research Ethics Review Committee of the General Hospital of Ningxia Medical University (No. 2020-690).
Acknowledgments
This work was supported by the Key Research and Development Project of Ningxia Hui Autonomous Region (No. 2021BEG03090).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Byappanahalli MN, Nevers MB, Korajkic A, et al. Enterococci in the environment. Microbiol Mol Biol Rev. 2012;76(4):685–706. doi:10.1128/MMBR.00023-12
2. Long KS, Wester B. Resistance to linezolid caused by modifications at its binding site on the ribosome. Antimicrob Agents Chemother. 2012;56(2):603–612. doi:10.1128/AAC.05702-11
3. Klare I, Fleige C, Geringer U, et al. Increased frequency of linezolid resistance among clinical Enterococcus faecium isolates from German hospital patients. J Glob Antimicrob Resist. 2015;3(2):128–131. doi:10.1016/j.jgar.2015.02.007
4. Tamang MD, Moon DC, Kim SR, et al. Detection of novel oxazolidinone and phenicol resistance gene optrA in enterococcal isolates from food animals and animal carcasses. Vet Microbiol. 2017;201:252–256. doi:10.1016/j.vetmic.2017.01.035
5. Mendes RE, Flamm RK, Hogan PA, Ross JE, Jones RN. Summary of linezolid activity and resistance mechanisms detected during the 2012 LEADER surveillance program for the United States. Antimicrob Agents Chemother. 2014;58(2):1243–1247. doi:10.1128/AAC.02112-13
6. Mendes RE, Hogan PA, Streit JM, Jones RN, Flamm RK. Zyvox® Annual Appraisal of Potency and Spectrum (ZAAPS) program: report of linezolid activity over 9 years (2004–12). J Antimicrob Chemother. 2014;69(6):1582–1588. doi:10.1093/jac/dkt541
7. Mendes RE, Deshpande L, Streit JM, et al. ZAAPS programme results for 2016: an activity and spectrum analysis of linezolid using clinical isolates from medical centres in 42 countries. J Antimicrob Chemother. 2018;73(7):1880–1887. doi:10.1093/jac/dky099
8. Bi R, Qin T, Fan W, Ma P, Gu B. The emerging problem of linezolid-resistant enterococci. J Glob Antimicrob Resist. 2018;13:11–19. doi:10.1016/j.jgar.2017.10.018
9. Kuroda M, Sekizuka T, Matsui H, et al. Complete genome sequence and characterization of linezolid-resistant Enterococcus faecalis clinical isolate KUB3006 Carrying a cfr(B)-transposon on its chromosome and optrA-Plasmid. Front Microbiol. 2018;9:2576. doi:10.3389/fmicb.2018.02576
10. Schwarz S, Zhang W, Du XD, et al. Mobile oxazolidinone resistance genes in gram-positive and gram-negative bacteria. Clin Microbiol Rev. 2021;34(3):e0018820. PMID: 34076490; PMCID: PMC8262807. doi:10.1128/CMR.00188-20
11. Brenciani A, Morroni G, Schwarz S, Giovanetti E. Oxazolidinones: mechanisms of resistance and mobile genetic elements involved. J Antimicrob Chemother. 2022;77(10):2596–2621. PMID: 35989417. doi:10.1093/jac/dkac263
12. Tsiodras S, Gold HS, Sakoulas G, et al. Linezolid resistance in a clinical isolate of Staphylococcus aureus. Lancet. 2001;358(9277):207–208. doi:10.1016/S0140-6736(01)05410-1
13. Mendes RE, Deshpande LM, Farrell DJ, et al. Assessment of linezolid resistance mechanisms among Staphylococcus epidermidis causing bacteraemia in Rome, Italy. J Antimicrob Chemother. 2010;65(11):2329–2335. doi:10.1093/jac/dkq331
14. Shinabarger DL, Marotti KR, Murray RW, et al. Mechanism of action of oxazolidinones: effects of linezolid and eperezolid on translation reactions. Antimicrob Agents Chemother. 1997;41(10):2132–2136. doi:10.1128/AAC.41.10.2132
15. Hu F, Guo Y, Yang Y, et al.; China Antimicrobial Surveillance Network (CHINET) Study Group. Resistance reported from China antimicrobial surveillance network (CHINET) in 2018. Eur J Clin Microbiol Infect Dis. 2019;38(12):2275–2281. doi:10.1007/s10096-019-03673-1
16. Cho SY, Kim HM, Chung DR, et al. Resistance mechanisms and clinical characteristics of linezolid-resistant Enterococcus faecium isolates: a single-centre study in South Korea. J Glob Antimicrob Resist. 2018;12:44–47. doi:10.1016/j.jgar.2017.09.009
17. Baquero F. Low-level antibacterial resistance: a gateway to clinical resistance. Drug Resist Updat. 2001;4(2):93–105. doi:10.1054/drup.2001.0196
18. Doern CD, Park JY, Gallegos M, Alspaugh D, Burnham CA, Richter SS. Investigation of linezolid resistance in Staphylococci and Enterococci. J Clin Microbiol. 2016;54(5):1289–1294. doi:10.1128/JCM.01929-15
19. Wang Y, Lv Y, Cai J, et al. A novel gene, optrA, that confers transferable resistance to oxazolidinones and phenicols and its presence in Enterococcus faecalis and Enterococcus faecium of human and animal origin. J Antimicrob Chemother. 2015;70(8):2182–2190. doi:10.1093/jac/dkv116
20. Cai J, Wang Y, Schwarz S, et al. High detection rate of the oxazolidinone resistance gene optrA in Enterococcus faecalis isolated from a Chinese anorectal surgery ward. Int J Antimicrob Agents. 2016;48(6):757–759. doi:10.1016/j.ijantimicag.2016.08.008
21. Sharkey LK, Edwards TA, O’Neill AJ, Wright GD. ABC-F proteins mediate antibiotic resistance through ribosomal protection. mBio. 2016;7(2):e01975. doi:10.1128/mBio.01975-15
22. Long KS, Poehlsgaard J, Kehrenberg C, Schwarz S, Vester B. The Cfr rRNA methyltransferase confers resistance to Phenicols, Lincosamides, Oxazolidinones, Pleuromutilins, and Streptogramin A antibiotics. Antimicrob Agents Chemother. 2006;50(7):2500–2505. doi:10.1128/AAC.00131-06
23. Diaz L, Kiratisin P, Mendes RE, Panesso D, Singh KV, Arias CA. Transferable plasmid-mediated resistance to linezolid due to cfr in a human clinical isolate of Enterococcus faecalis. Antimicrob Agents Chemother. 2012;56(7):3917–3922. doi:10.1128/AAC.00419-12
24. Wang L, He Y, Xia Y, Wang H, Liang S. Investigation of mechanism and molecular epidemiology of linezolid-resistant Enterococcus faecalis in China. Infect Genet Evol. 2014;26:14–19. doi:10.1016/j.meegid.2014.05.001
25. Liu Y, Wang Y, Schwarz S, et al. Investigation of a multiresistance gene cfr that fails to mediate resistance to phenicols and oxazolidinones in Enterococcus faecalis. J Antimicrob Chemother. 2014;69(4):892–898.
26. Li B, Ma CL, Yu X, et al. Investigation of mechanisms and molecular epidemiology of linezolid nonsusceptible Enterococcus faecalis isolated from a teaching hospital in China. J Microbiol Immunol Infect. 2016;49(4):595–599. doi:10.1016/j.jmii.2015.05.010
27. Chen H, Wu W, Ni M, et al. Linezolid-resistant clinical isolates of enterococci and Staphylococcus cohnii from a multicentre study in China: molecular epidemiology and resistance mechanisms. Int J Antimicrob Agents. 2013;42(4):317–321. doi:10.1016/j.ijantimicag.2013.06.008
 © 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.