Back to Journals » International Journal of Chronic Obstructive Pulmonary Disease » Volume 15
Circulating miR-1246 in the Progression of Chronic Obstructive Pulmonary Disease (COPD) in Patients from the BODE Cohort
Authors Cazorla-Rivero S, Mura-Escorche G, Gonzalvo-Hernández F, Mayato D, Córdoba-Lanús E, Casanova C
Received 12 July 2020
Accepted for publication 23 September 2020
Published 29 October 2020 Volume 2020:15 Pages 2727—2737
DOI https://doi.org/10.2147/COPD.S271864
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. Richard Russell
Sara Cazorla-Rivero,1,2 Glorian Mura-Escorche,1,2 Francisca Gonzalvo-Hernández,3 Delia Mayato,1 Elizabeth Córdoba-Lanús,1,2,4,* Ciro Casanova1– 3,*
1Research Unit, Hospital Universitario Nuestra Señora de Candelaria, Santa Cruz de Tenerife, Spain; 2University of La Laguna, San Cristóbal de La Laguna, Tenerife, Spain; 3Pulmonary Department, Hospital Universitario Nuestra Señora de Candelaria, Santa Cruz de Tenerife, Spain; 4Instituto de Enfermedades Tropicales y Salud Pública de Canarias (IUETSPC), San Cristóbal de La Laguna, Tenerife, Spain
*These authors contributed equally to this work
Correspondence: Elizabeth Córdoba-Lanús
University of La Laguna- IUETSPC, Av. Astrofísico Francisco Sanchez. Campus Anchieta, Apartado 456, San Cristobal de la Laguna CP 38200, Tenerife, Spain
Email [email protected]
Background: COPD is characterized by a persistent inflammatory response, especially against cigarette smoke. COPD patients may develop varying degrees of emphysematous destruction of the lungs. A pathophysiological role for miRNAs in COPD has been suggested in several studies. We examined changes in microRNAs expression profile during 10 years follow-up in relation to COPD progression.
Methods: Clinical and lung function parameters were registered from every subject included in the study. miRNAs expression was determined in 14 serum samples from 7 patients in two moments (4 smokers with COPD (BODE cohort) and 3 smokers without COPD) by next generation sequencing (NGS) at baseline and after 10 years follow-up. A validation study was performed by qPCR in 20 patients with COPD (13 emphysema-diagnosed by CTscan) and 10 smoker controls at baseline and after 10 years follow-up. hsa-miRNA-20a-5p and hsa-let-7d-5p were used as endogenous controls.
Results: A total of 198 miRNAs (≥ 10TPM) were identified by NGS. Between these, hsa-miR-1246 was found significantly downregulated in COPD patients after 10 years when compared to baseline (p< 0.0001, FDR=0.05). Seventy-five percent of these patients had an emphysema diagnose. In the validation analysis, when analyzed longitudinally, hsa-miR-1246 was significantly downregulated in COPD patients with emphysema after 10 years (p= 0.019). However, no association was found between the expression of miR-1246 and any other lung function parameters (FEV1, PaO2, DLCO, IC/TLC) within the follow-up period. GO and KEGG enrichment analysis revealed miR-1246 to be associated with target genes in several pathways involved in COPD/emphysema development.
Conclusion: Our findings suggest that hsa-miR-1246 may act as a biomarker of emphysema in COPD. Functional analysis is guaranteed to elucidate its role in COPD.
Keywords: COPD progression, miRNAs, emphysema
Introduction
Chronic obstructive pulmonary disease (COPD) is characterized by a progressive airflow obstruction and a persistent inflammatory response, especially against cigarette smoke.1,2 COPD is the third leading cause of chronic morbidity and mortality worldwide.3
Recent reports suggest that cigarette smoke induces oxidative stress-mediated DNA damage and triggers cellular senescence in the lungs, which results in several pathophysiological changes in the lung.2,3 The infiltration of small airways by neutrophils, macrophages and T lymphocytes is associated with further release of cytokines and proteolytic enzymes, which amplifies the inflammation and release proteases and reactive oxidative species that damage the parenchymal lung tissue, results in the destruction of alveolar structure and increased mucus secretion, contributing to the development of emphysema in COPD patients.4 This impaired tissue regeneration activates matrix remodeling and tissue repair mechanism such as cellular proliferation and senescence that are however insufficient to counteract its effect.
miRNAs are non-coding 18–24 nucleotide long RNA molecules. miRNAs play a critical role in several biological processes such as apoptosis, cell differentiation, proliferation, DNA damage repair, angiogenesis, stress response and stem cell division. They act primarily by suppressing mRNA translation and thereby reducing the levels of the proteins involved.5,6 Deregulation of miRNAs can be caused by various mechanisms, either genetic or epigenetic.7 It has been reported that miRNA expressions dysregulation is associated with the development and progression of several human diseases like cancer, neurological, cardiovascular diseases8 and pulmonary ones.9
A pathophysiological role for miRNAs in COPD has been suggested in several studies. Previous studies showed a differential expression of certain miRNAs in different types of samples such as sputum, bronchial and lung tissue10–12 or even serum or plasma13,14 in smokers and COPD cases. It has also been described how tobacco smoke affects the expression of miRNAs of the bronchial epithelium.15 It has been suggested several deregulated miRNAs to be involved in the development of tobacco-associated diseases such as COPD and its progression.16 A study by Xie et al demonstrated that the serum ratio of miR-21 to miR-181a could predict the risk of suffering COPD in asymptomatic smokers with a significant smoking burden.17 Moreover, Sato et al found that the expression of miR-146a was correlated with the severity of COPD.18 However, all research designs until now were cross-sectional. Longitudinal studies are needed to evaluate potentially useful miRNAs as biomarkers for early detection of disease-related molecular and genetic changes, as well as a risk marker of the disease evolution by early detection of pulmonary function loss, exacerbations and/or mortality.
The aim of the present study was to examine changes in a microRNA expression profile after 10 years-follow-up in relation to COPD progression serum samples in a very well characterized cohort of patients.
Methods
Study Individuals
Patients
Twenty-four individuals with a diagnosis of COPD recruited from the Hospital Universitario N/S de Candelaria, Tenerife, Spain, were included in this study. They are part of a cohort of 362 individuals with annual clinical follow-up since 2002, part of the BODE multicenter study.19 From these, four individuals were included in the screening step and other 20 were included in the subsequent validation step. Inclusion criteria: age >40 years, post-bronchodilator FEV1/FVC ratio <0.70 and clinically stable for at least 6 weeks at the time of evaluation. Spirometry lung volumes, pulmonary function test, and exercise capacity were measured according to ATS-ERS guidelines20–22 and severity was graded by the Global Initiative for Obstructive Lung Disease (GOLD).21 Dyspnoea was evaluated by mMRC scale23 and the BODE Index was calculated as previously described.19 Co-morbidities were quantified using the Charlson index.24 A pulmonologist visually scored the baseline for emphysema presence, using validated criteria established by the Fleischner Society.25 Exclusion criteria included any other respiratory diseases and uncontrolled comorbidities such as malignancy at baseline.
Controls
Based on the availability of tobacco-smoker controls without COPD (>44 years, smoking history of >15 pack-year and FEV1% pred >0.80; FEV1/FVC >0.70), with blood sample and with a follow-up of ten years, 13 individuals could be included in this study (3 smoker controls for screening step and 10 for the validation study in the second stage of this research).
The study was approved by the ethical committee board of Hospital Universitario N/S de Candelaria and written informed consent was obtained from all participants (PI 55/17). This study was conducted in accordance with the Declaration of Helsinki.
Sample Collection
A total of 74 serum samples were collected from participants (24 stable COPD patients and 13 control smokers without COPD at two moments: baseline and after 10 years of follow-up) between the screening and validation steps. Serum samples were isolated within 1 h after receiving whole blood and immediately stored at −80 °C until further use.
miRNA Screening in COPD Patients
RNA was isolated from 7 participants (4 smokers with COPD and 3 control smokers without COPD) at baseline and after 10 years follow-up (14 serum samples). Isolation of RNA from 200 µL serum of patients was performed using a miRNeasy Serum/Plasma Advanced Kit (Qiagen). miRNAs expression was determined by next generation sequencing (NGS) in an Illumina NextSeq500 platform at Exiqon A/S company (Denmark). After mapping the data and counting to relevant entries in miRBase v20 software (http://mirbase.org)26 the numbers of known microRNAs were calculated. miRNAs expression is expressed as Tags Per Million (“TPM”, the number of reads for a particular microRNA). microRNA stably expressed across all samples were identified using NormFinder software.27
Quantitative RT-PCR for miRNA Expression. Validation Assay
A validation study was performed in 20 patients with COPD (from them 13 had emphysema-diagnosed by CT scan) and 10 control smokers without COPD, at two moments: baseline and after 10 years follow-up (a total of 60 serum samples). Isolated RNA from serum samples was used in a retro transcription reaction to synthesize cDNA using miRCURY LNA RT kit (Qiagen Inc., Germany), according to the manufacturer instructions. We also used RNA spike-ins (UniSp2, UniSp4 and UniSp5, as an RNA isolation control. And cel-miR-39-3p in conjunction with the UniSp6 spike-in as cDNA synthesis and amplification controls) (miRCURY LNA RT Kit and RNA Spike-in Kit, Qiagen Inc., Germany). A qPCR analysis was used to quantify the expression of the resulting significant miRNAs from the NGS screening study at the first stage. In this case, hsa-miR-1246 expression was determined by using miRCURY LNA Sybr Green PCR Master Mix (Qiagen Inc., Germany). The reactions were performed in a final volume of 10 µL and contained 1X Sybr Green Master Mix, 200 nmol/L specific primer set (miRCURY LNA miRNA PCR assay, Qiagen Inc., Germany), using 3 ng cDNA per reaction. The conditions included an initial denaturation at 95 °C for 15 min., followed by 40 cycles of 94 °C for 15 s, 55 °C for 30 s, and 72 °C for 30 s. All the samples were performed in triplicates. Experiments were performed on a StepOnePlus real-time PCR system (Applied Biosystem, Foster city, CA, USA). hsa-miRNA-20a-5p and hsa-let-7d-5p, that resulted as the best candidates for normalization control in the first stage screening by the NormFinder software, were also used for normalization as the reference genes in this assay. A non-template control (NTC) was carried out in each experiment, crucial to identify any contamination that may increase the background.
The relative expression analysis of the target miRNAs was performed using the comparative threshold method 2−ΔΔCt.28
In silico Analysis
We also integrated the miRNA and the differentially expressed putative target mRNA using the miRBase v20 (http://www.mirbase.org/),26 TargetScan (http://www.targetscan.org/vert_72/)29 and the DIANA-microT_CDS softwares (http://diana.imis.athena-innovation.gr/DianaTools/index.php?r=MicroT_CDS/index).30 In the next step, the g:Profiler software was used to contrast the resulting genes proposed as targets of miR-1246.31
We performed functional and enrichment analyses through the Kyoto Encyclopedia of Genes and Genomes (KEGG) (https://www.kegg.jp/) and the Gene-ontology (GO) terms32,33 by using the miRTar.Human software (http://mirtar.mbc.nctu.edu.tw/human/index.php).
Statistical Analysis
For the cross-sectional analysis, a t-Student, ANOVA, Chi2, Fisher Exact, Kruskal–Wallis test were used to test differences in means and proportions of clinical and lung function characteristics between smoker patients with COPD and smokers without the disease.
miRNAs differential expression analysis resulting from NGS was performed using the EdgeR statistical software package (Bioconductor, http://bioconductor.org/).34 For normalization, the trimmed mean of M-values method based on log-fold and absolute gene-wise changes in expression levels between samples (TMM normalization) was used. The Benjamini-Hochberg false discovery rate (FDR) algorithm was used to corrected p-values for multiple testing.35
The association between baseline miRNA expression with clinical and/or pulmonary function variables was explored using Pearson’s correlation coefficients. χ2 Pearson test was used for comparisons of differential miRNAs expression between cases and the control group.
A multiple logistic regression was performed to test the association of telomere length with COPD adjusting for age, sex and pack years. General linear model (GLIM) for repeated measures was used to assess disease progression from longitudinal clinical and lung function data. For genetic analysis, SPSS 21.0 IBM Co software was used for all statistical analyses and two-tailed p-values <0.05 were considered significant.
Results
Screening by NGS
A total of seven individuals (COPD cases and smoker controls) were included in the screening study in two moments. Both groups were similar in age, gender and smoking habit. Three COPD patients had emphysema diagnosed by CT scan. The main clinical characteristics of these individuals are shown in Supplemental Table 1.
One hundred and ninety-eight miRNAs (≥10TPM) were identified by Next Generation Sequencing (NGS) in the 7 patients (14 samples analyzed) (Figure 1). Between these, hsa-miR-184 was found differentially expressed between cases and controls at baseline (fold change=1.89, p=0.004) although it was discarded for further analysis as FDR>0.05 (Table 1). Interestingly, hsa-miR-1246 was found significantly downregulated in COPD patients after 10 years when compared to baseline (fold change=1.7; p<0.0001; FDR=0.05) (Figure 2). miR-1246 expression level in the control smoker group after 10-year follow-up was similar to the one observed at baseline (p=0.704). Comparison between smokers without COPD at baseline and after the follow-up resulted in hsa-miR-203a downregulated with a log fold change of 1.89 (p=0.0012), but FDR>0.05 so discarded for further analysis.
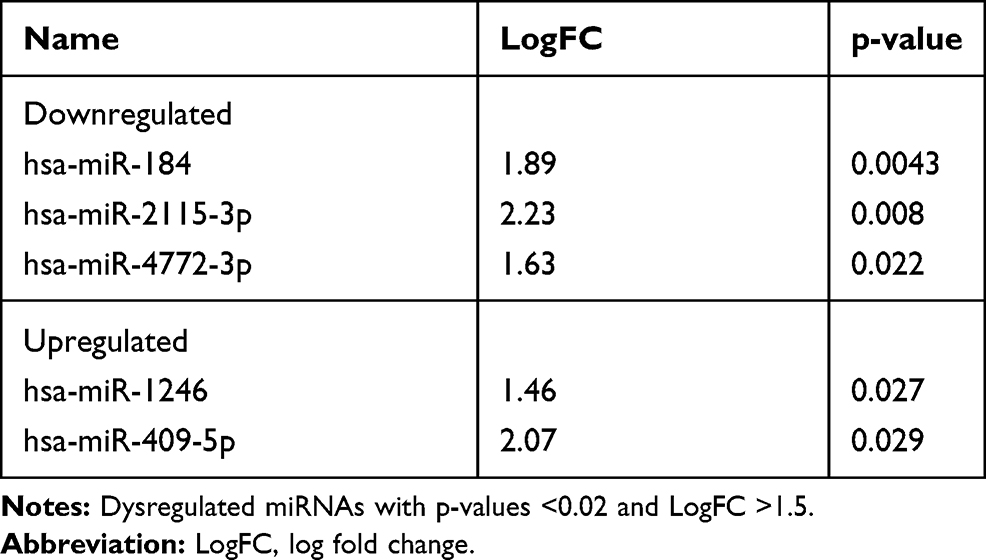 |
Table 1 Main Dysregulated miRNAs in Serum from Patients with COPD vs. Smokers Without the Disease at Baseline |
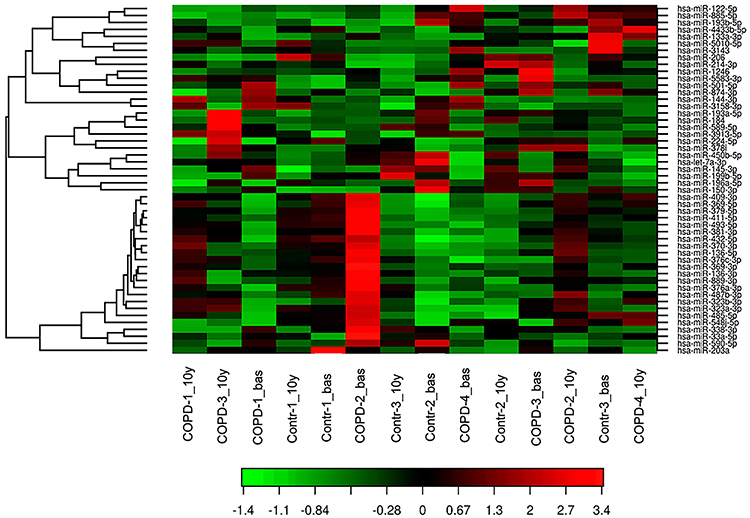 |
Figure 1 Heat Map showing the expression profiling of circulating miRNAs in patients with COPD and control smokers without COPD. miRNA assay by NGS was performed in 14 serum samples (7 corresponds to baseline (named COPD-1 to 4_bas and Controls-1 to 3_bas) and 7 to the sample individuals after 10 years of follow-up (named COPD-1 to 4_10y and Controls-1 to 3_10y)). Red represents a high level of gene expression while green represents a low level of expression. |
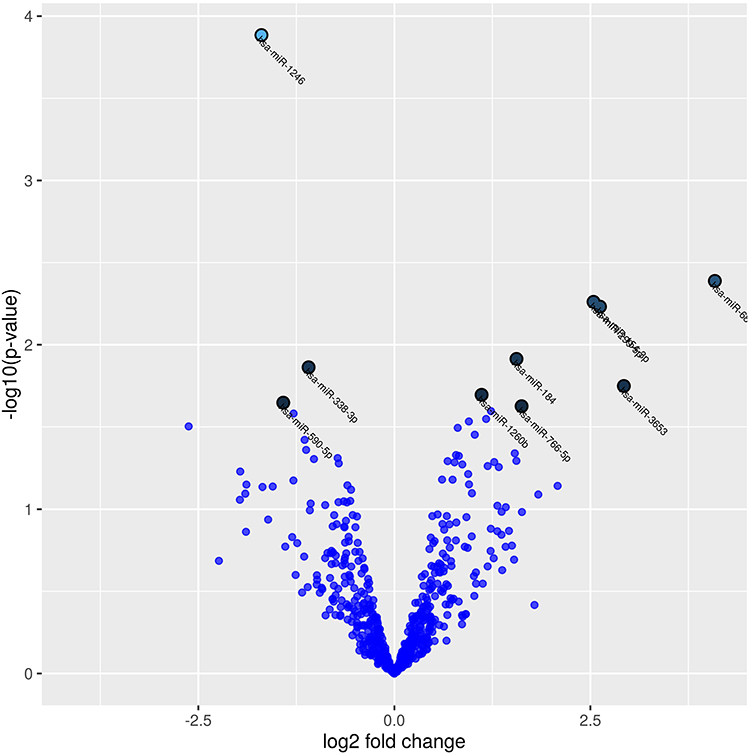 |
Figure 2 Volcano plot showing differentially expressed miRNAs in terms of the fold change in normalized expression in COPD patients at baseline compared to COPD patients after 10 years follow-up. The 10 microRNAs with the lowest p-values are marked with names on the plot (p-values <0.02 and log fold change (LogFC) >1). |
Validation Study
Table 2 shows the main clinical characteristics of the 30 individuals included in the validation study. Patients with COPD and smoker controls were similar in age, gender and body mass index (BMI). Although COPD cases had a higher number of pack-years smoked, this was not significant and neither existed differences between individuals who continued smoking in both groups. Patients with COPD presented worse lung function and walked less. More than a half of the cases presented emphysema diagnosed by CT scan.
 |
Table 2 Baseline Clinical and Lung Function Characteristics of the COPD Patients and Control Smokers Included in the Validation Study |
Cross Sectional
In the validation analysis, we did not find any significant difference between the expression of miR-1246 and the groups of patients with COPD and control smokers at baseline (Supplemental Figure 1). When we investigate the relation between the clinical and pulmonary function parameters or the presence of emphysema with differential expression of this miRNA in COPD patients, no significant differences were found between groups. When we compared the expression of miR-1246 in COPD with emphysema vs. COPD without emphysema at baseline, we did not find any difference (p=0.817).
Longitudinal Analysis
In addition, in the longitudinal analysis, miR-1246 was found to be less expressed although no significant, in patients with COPD after 10 years of follow-up when compared to baseline (fold change=1.73). The smokers without COPD did not show any differential expression for this miRNA during the monitoring period. Clinical and lung function differences achieved in COPD patients during the monitoring time are shown in Table 3, as well as for controls in Supplemental Table 1. Interestingly, when we analyzed only patients with emphysema, this miRNA was significantly less expressed (p= 0.019) in this group after the 10-year follow-up period in contrast to baseline (Figure 3). However, no association was found between lung function parameters (FEV1, PaO2, IC/TLC) including the diffusion capacity (DLCO), a subrogate marker of emphysema, and miR-1246 when evaluating emphysema progression during monitoring follow-up.
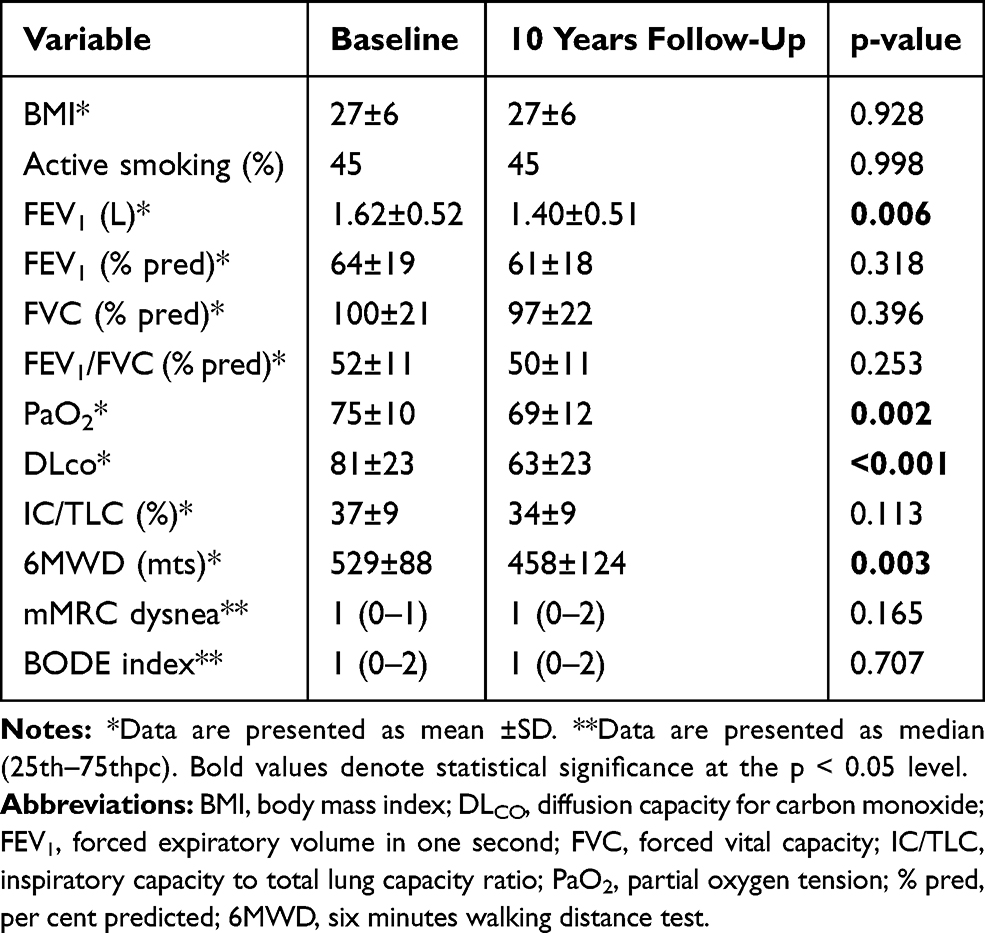 |
Table 3 Baseline and 10-Years Follow-Up Clinical and Lung Function Characteristics of the COPD Patients Included in the Validation Study |
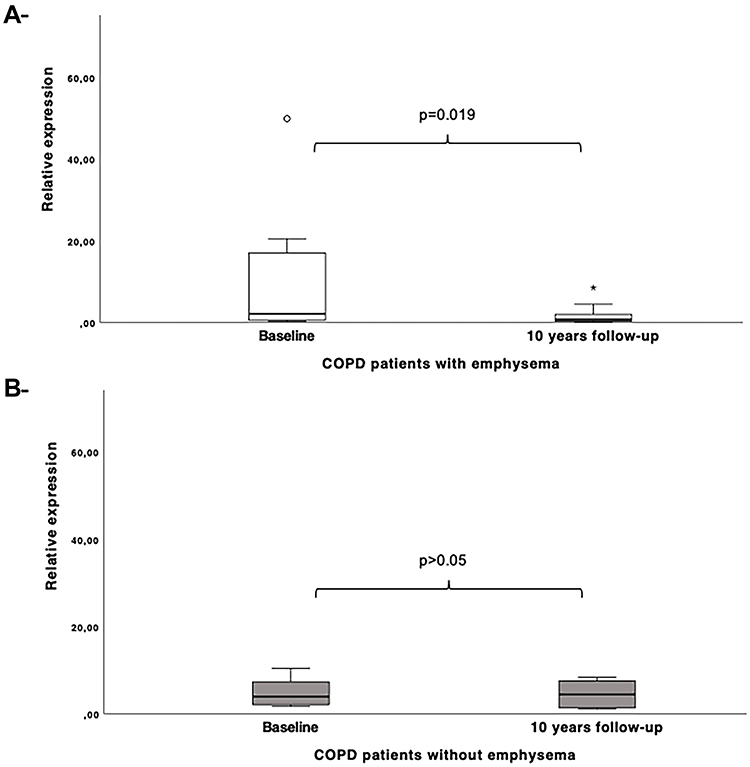 |
Figure 3 Dysregulated expression of miR-1246 in COPD patients with emphysema (A) and patients without emphysema (B) at 10 years-follow-up in contrast to baseline. |
In silico Analysis
Our functional analysis, through GO enrichment of the DEGs on the biological process (BP), molecular function (MF) and cellular component (CC) categories and the KEGG enrichment pathways revealed miR-1246 to be significantly associated with target genes enriched in several functions and signaling pathways. Between them, are of particular interest, the vascular smooth muscle signaling pathway, the MAPK signaling cascade, the chemokine signaling pathway and the Wnt cascade (Table 4), all reported to be involved in COPD/emphysema development.
 |
Table 4 The Enriched KEGG Pathway of Predicted Genes |
Discussion
The main finding of this study is the significant downregulation of miR-1246 observed in COPD patients with emphysema after a follow-up of 10 years. GO ontology and KEGG enrichment analysis revealed miR-1246 to be associated with target genes in several pathways involved in COPD/emphysema development.
Altered miRNAs expression was reported in several studies performed on different samples as sputum, serum and lung tissue of patients with COPD when compared to smokers without the disease.10,12,17,36,37 Dysregulated miRNAs were observed in relation to COPD, and in relation clinical severity18 and to increase the risk of acute exacerbations in stable patients with COPD.37
Other authors investigated potential miRNAs to be implicated in the destruction of the emphysematous lung in COPD. Christenson et al found several miRNAs to be dysregulated in patients with severe emphysema and between them, demonstrated a role for miR-638 in promoting the maturity of emphysematous lung tissue and lung fibroblasts.38 Savarimuthu et al found miR-34b and miR-34c to be less significantly expressed in lung tissue of patients with moderate emphysema when compared to patients with mild forms of the disease, suggesting this miRNA to modulate SERPINE gene expression.39 Another study on lung tissue from patients with COPD showed increased expression of miR-15b in areas of emphysema as well as fibrosis.12 A recent study that analyzed dysregulated miRNAs in PBMCs found miR-335 to be downregulated in patients with severe emphysema with PiZZ AATD when compared to mild ones suggesting a role for this miRNA in the activation of pathways related to inflammation and angiogenesis.40 miR-1246 expression has also been reported to be altered in relation to several cancer: breast cancer41 hepatocellular carcinoma,42 pancreatic cancer,43 and lung cancer development44,45 and therapeutics.46
In our study a diminished expression of miRNA-1246 was observed particularly in COPD patients with emphysema after 10 years. Although this novel relationship, our longitudinal analysis over the follow-up period did not find an association between the progression of emphysema monitored by the diffusion capacity (DLCO). Further studies in a larger cohort that may include a longitudinal evaluation of emphysema by CT scan could contribute to understand the possible role of this miRNA in this type of patients.
To our knowledge, a prognostic value of miRNAs that may be involved in a worse progression of COPD has not been investigated by previous studies. Furthermore, there are no previous longitudinal studies exploring deregulated miRNAs in relation to the progression of COPD. Our research expanded over the past ones in studying miRNA expression dysregulation in patients suffering COPD and smoker controls during 10 years of follow-up.
Our functional in silico analysis suggests miR-1246 to be linked to genes mediators in various signaling pathways that may be related to emphysema. Our results showed mir-1246 to target genes like AKT1 in the insulin growth factor (IGF) pathway. A recent study by Cottage et al (2019) reveals that levels of pro-growth mediators IGF1 and Akt are diminished in COPD lung compared with normal subjects, suggesting that a defective IGF1 pathway may mediate in part, the compromised tissue regeneration seen in COPD lungs.47 The age-related loss of the replicative and regenerative signaling is known to be associated with the diminution of the insulin/insulin like growth factor (IGF1) pathway. The IGF1 pathway is involved in various mechanisms of the cell cycle, including proliferation, survival, and differentiation through downstream Akt activity.48 In this sense miR-1246 may have interesting role in this pathway that is to be unravel in relation to emphysema and COPD.
It was proposed that imbalance between protease and antiprotease process that results on emphysema could also have direct implication on chemokines degradation. The CXCL12-CXCR4 axis in chemokine-mediated signaling pathway has been the focus target of many strategies based on its neutralization49 and targeting the CXCL12-CXCR4 axis may be promising for COPD treatment. We found miR-1246 to be associated with CXCL12 in this pathway. Barbiwska et al (2018) demonstrated that in a mouse model of cigarette smoke exposure, the intermittent administration of plerixafor (FDA-approved CXCR4 antagonist) decreases emphysema damages, without affecting CXCL12 level and inflammation in BALF.50
Our predictive analysis also suggested a link between miR-1246 and genes in the Wnt signaling pathway. This cascade has been deeply studied and its known to serve as a critical regulator in lung development and also in physiological and pathophysiological processes of lung in adulthood.51 A recent research by Yang et al (2019) demonstrated in an in vitro study that miR-1246 regulates Wnt/β-catenin pathway through targeting GSK-3β/β-catenin.52
This study has several limitations: firstly, we used a small number of participants in the screening phase to identify key miRNAs to be later validated in the total sample. Although, this is a common limitation inherent to this type of analysis due to its complexity and economic cost. Second, we did not study the expression of miR-1246a in lung tissue. However, circulating miRNAs has demonstrated to be feasible biomarkers in the diagnosis of several diseases. For example, miRNA profiles in some biological fluids have been shown to adequately reflect tumor characteristics, without the need for tissue biopsies.53 Third, the lack of longitudinal evaluation of emphysema by CT scan. However, we used instead of the annuals records of DLCO, a recognized subrogate marker of emphysema.
In conclusion, our longitudinal observational study on COPD showed a novel link with hsa-miR-1246 consistent with a differential downregulated expression pattern in COPD patients with emphysema after 10 years of follow-up. Functional analyses are guaranteed to elucidate its role in emphysema progression in COPD.
Abbreviations
COPD, chronic obstructive pulmonary disease; miRNA, microRNA; hsa-miRNA, homo sapiens microRNA; FEV1, forced expiratory volume in one second; FVC, forced vital capacity; BMI, body mass index; PaO2, partial oxygen tension; DLCO, diffusion capacity for carbon monoxide; IC/TLC, inspiratory capacity to total lung capacity ratio; 6MWD, six-minute walking distance test; CT scan, computed tomography scanner; NGS, next generation sequencing; TPM, tags per million; FDR, false discovery rate; qPCR, quantitative real time polymerase chain reaction; GLIM, general linear modelling for repeated measures test.
Data Sharing Statement
Data are available upon reasonable request. All data relevant to the study are included in the article or uploaded as Supplemental Table 1 and Supplemental Figure 1.
Ethics Approval
Ethical approval was obtained from Hospital Universitario La Candelaria, Tenerife, Spain (PI 55/17).
Patient Consent
Written informed consent was obtained from all participants.
Acknowledgments
We will like to thank Hilaria González Acosta for her excellent technical assistance and the Instituto Universitario de Enfermedades Tropicales y Salud Pública de Canarias (IUETSPC) for the use of laboratory equipment. Elizabeth Córdoba-Lanús is supported by Cabildo de Tenerife (Agustín de Betancourt programme).
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; and agree to be accountable for all aspects of the work.
Funding
This study was supported by AstraZeneca (2017).
Disclosure
Sara Cazorla-Rivero, Glorian Mura-Escorche, Francisca Gonzalvo-Hernández, Delia Mayato, and Elizabeth Córdoba-Lanús, declare not to have any financial or personal conflict of interests. Ciro Casanova Macario reports grants from Astra-Zeneca, GSK, and Menarini, advisory board for Astra-Zeneca and GSK, financial support for congress from Boehringer, Novartis and Menarini, personal fees from Bial, and lectures for Novartis, outside the submitted work; and declares to have received lectures and/or scientific advice from Laboratorios Bial, Boehringer-Ingelheim, AstraZeneca, GSK, Esteve, Menarini, Novartis and Rovi in the last three years. The authors report no other potential conflicts of interest for this work.
References
1. Global Initiative for Chronic Obstructive Lung Disease (GOLD). Global strategy for the diagnosis, management, and prevention of chronic obstructive pulmonary disease (2018 report). Available from: https://goldcopd.org/wp-content/uploads/2017/11/GOLD-2018-v6.0-FINAL-revised-20-Nov_WMS.pdf.
2. John-Schuster G, Günter S, Hager K, et al. Inflammaging increases susceptibility to cigarette smoke-induced COPD. Oncotarget. 2016;7(21):30068–30083. doi:10.18632/oncotarget.4027
3. Lee J, Sandford A, Man P, Sin DD. Is the aging process accelerated in chronic obstructive pulmonary disease? Curr Opin Pulm Med. 2011;17(2):90–97. doi:10.1097/MCP.0b013e328341cead
4. Sethi S, Mahler DA, Marcus P, et al. Inflammation in COPD: implications for management. Am J Med. 2012;125(12):1162–1170. doi:10.1016/j.amjmed.2012.06.024
5. Wu L, Fan J, Belasco JG. MicroRNAs direct rapid deadenylation of mRNA. Proc Natl Acad Sci U S A. 2006;103(11):4034–4039. doi:10.1073/pnas.0510928103
6. Meister G. miRNAs get an early start on translational silencing. Cell. 2007;131(1):25–28. doi:10.1016/j.cell.2007.09.021
7. Croce CM. Causes and consequences of microRNA dysregulation in cancer. Nat Rev Genet. 2009;10(10):704–714. doi:10.1038/nrg2634
8. Dai R, Ahmed SA. MicroRNA, a new paradigm for understanding immunoregulation, inflammation, and autoimmune diseases. Transl Res. 2011;157(4):163–179. doi:10.1016/j.trsl.2011.01.007
9. Rupani H, Sanchez-Elsner T, Howarth P. MicroRNAs and respiratory diseases. Eur Respir J. 2013;41(3):695–705. doi:10.1183/09031936.00212011
10. Van Pottelberge GR, Mestdagh P, Bracke KR, et al. MicroRNA expression in induced sputum of smokers and patients with chronic obstructive pulmonary disease. Am J Respir Crit Care Med. 2011;183(7):898–906. doi:10.1164/rccm.201002-0304OC
11. Schembri F, Sridhar S, Perdomo C, et al. MicroRNAs as modulators of smoking-induced gene expression changes in human airway epithelium. Proc Natl Acad Sci USA. 2009;106(7):2319–2324. doi:10.1073/pnas.0806383106
12. Ezzie ME, Crawford M, Cho JH, et al. Gene expression networks in COPD: microRNA and mRNA regulation. Thorax. 2012;67:122–131. doi:10.1136/thoraxjnl-2011-200089
13. Akbas F, Coskunpinar E, Aynaci E, et al. Analysis of serum micro-RNAs as potential biomarker in chronic obstructive pulmonary disease. Exp Lung Res. 2012;38(6):286–294. doi:10.3109/01902148.2012.689088
14. Soeda S, Ohyashiki JH, Ohtsuki K, et al. Clinical relevance of plasma miR-106b levels in patients with chronic obstructive pulmonary disease. Int J Mol Med. 2013;31(3):533–539. doi:10.3892/ijmm.2013.1251
15. Graff J, Powers L, Dickson A, et al. Cigarette smoking decreases global microRNA expression in human alveolar macrophages. PLoS One. 2012;7(8):e44066. doi:10.1371/journal.pone.0044066
16. Banerjee A, Luettich K. MicroRNAs as potential biomarkers of smoking-related diseases. Biomark Med. 2012;6(5):671–684. doi:10.2217/bmm.12.50
17. Xie L, Wu M, Lin H, et al. An increased ratio of serum miR-21 to miR-181a levels is associated with the early pathogenic process of chronic obstructive pulmonary disease in asymptomatic heavy smokers. Mol Biosyst. 2014;10(5):1072–1081. doi:10.1039/C3MB70564A
18. Sato T, Liu X, Nelson A, et al. Reduced miR-146a increases prostaglandin E 2 in chronic obstructive pulmonary disease fibroblasts. Am J Respir Crit Care Med. 2010;182(8):1020–1029. doi:10.1164/rccm.201001-0055OC
19. Celli BR, Cote C, Marin JM, et al. The body mass index, airflow obstruction, dyspnea, exercise performance (BODE) index in chronic obstructive pulmonary disease. N Engl J Med. 2004;350(10):1005–1012. doi:10.1056/NEJMoa021322
20. ATS Committee on Proficiency Standards for Clinical Pulmonary Function Laboratories. ATS statement: guidelines for the six-minute walk test [published correction appears in Am J Respir Crit Care Med. 2016 May 15;193(10):1185]. Am J Respir Crit Care Med. 2002;166(1):111–117.
21. American Thoracic Society Statement. Lung function testing: selection of reference values and interpretative strategies. Am Rev Respir Dis. 1991;144(5):1202–1218. doi:10.1164/ajrccm/144.5.1202
22. Macintyre N, Crapo RO, Viegi G, et al. Standardisation of the single-breath determination of carbon monoxide uptake in the lung. Eur Respir J. 2005;26(4):720–735. doi:10.1183/09031936.05.00034905
23. Mahler DA, Wells CK. Evaluation of clinical methods for rating dyspnea. Chest. 1988;93(3):580–586. doi:10.1378/chest.93.3.580
24. Charlson M, Szatrowski TP, Peterson J, Gold J. Validation of a combined comorbidity index. J Clin Epidemiol. 1994;47(11):1245–1251. doi:10.1016/0895-4356(94)90129-5
25. Lynch DA, Austin JH, Hogg JC, et al. CT-definable subtypes of chronic obstructive pulmonary disease: a statement of the fleischner society. Radiology. 2015;277(1):192–205. doi:10.1148/radiol.2015141579
26. Kozomara A, Birgaoanu M, Griffiths-Jones S. miRBase: from microRNA sequences to function. Nucleic Acids Res. 2019;47(D1):D155–D162. doi:10.1093/nar/gky1141
27. Andersen CL, Jensen JL, Ørntoft TF. Normalization of real-time quantitative reverse transcription-PCR data: a model-based variance estimation approach to identify genes suited for normalization, applied to bladder and colon cancer data sets. Cancer Res. 2004;64(15):5245–5250. doi:10.1158/0008-5472.CAN-04-0496
28. Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods. 2001;25(4):
29. Agarwal V, Bell GW, Nam JW, Bartel DP. Predicting effective microRNA target sites in mammalian mRNAs. Elife. 2015;4:e05005. doi:10.7554/eLife.05005
30. Paraskevopoulou MD, Georgakilas G, Kostoulas N, et al. DIANA-microT web server v5.0: service integration into miRNA functional analysis workflows. Nucleic Acids Res. 2013;41(Web Server issue):W169–W173. doi:10.1093/nar/gkt393
31. Raudvere U, Kolberg L, Kuzmin I, et al. g:Profiler: a web server for functional enrichment analysis and conversions of gene lists (2019 update). Nucleic Acids Res. 2019;47(W1):W191–W198. doi:10.1093/nar/gkz369
32. Ashburner M, Ball CA, Blake JA, et al. Gene ontology: tool for the unification of biology. The gene ontology consortium. Nat Genet. 2000;25(1):
33. Mi H, Muruganujan A, Ebert D, et al. PANTHER version 14: more genomes, a new PANTHER GO-slim and improvements in enrichment analysis tools. Nucleic Acids Res. 2019;47(D1):D419–D426. doi:10.1093/nar/gky1038
34. Huber W, Carey V, Gentleman R, et al. Orchestrating high-throughput genomic analysis with bioconductor. Nat Methods. 2015;12(2):115–121. doi:10.1038/nmeth.3252
35. Storey JD, Tibshirani R. Statistical significance for genomewide studies. Proc Natl Acad Sci U S A. 2003;100(16):9440–9445. doi:10.1073/pnas.1530509100
36. Dang X, Qu X, Wang W, et al. Bioinformatic analysis of microRNA and mRNA regulation in peripheral blood mononuclear cells of patients with chronic obstructive pulmonary disease. Respir Res. 2017;18(1):4. doi:10.1186/s12931-016-0486-5
37. Chen B-B, Li Z-H, Gao S. Circulating miR-146a/b correlates with inflammatory cytokines in COPD and could predict the risk of acute exacerbation COPD. Medicine (Baltimore). 2018;97(7):e9820. doi:10.1097/MD.0000000000009820
38. Christenson SA, Brandsma CA, Campbell JD, et al. miR-638 regulates gene expression networks associated with emphysematous lung destruction. Genome Med. 2013;5:114. doi:10.1186/gm519
39. Savarimuthu Francis SM, Davidson MR, Tan ME, et al. MicroRNA-34c is associated with emphysema severity and modulates SERPINE1 expression. BMC Genomics. 2014;15(1):88. doi:10.1186/1471-2164-15-88
40. Esquinas C, Janciauskiene S, Gonzalo R, et al. Gene and miRNA expression profiles in PBMCs from patients with severe and mild emphysema and PiZZ alpha1-antitrypsin deficiency. Int J Chron Obstruct Pulmon Dis. 2017;12:3381–3390. doi:10.2147/COPD.S145445
41. Dvinge H, Git A, Gräf S, et al. The shaping and functional consequences of the microRNA landscape in breast cancer. Nature. 2013;497(7449):378–382. doi:10.1038/nature12108
42. Moshiri F, Salvi A, Gramantieri L, et al. Circulating miR-106b-3p, miR-101-3p and miR-1246 as diagnostic biomarkers of hepatocellular carcinoma. Oncotarget. 2018;9(20):15350–15364. doi:10.18632/oncotarget.24601
43. Xu Y-F, Hannafon BN, Zhao YD, et al. Plasma exosome miR-196a and miR-1246 are potential indicators of localized pancreatic cancer. Oncotarget. 2017;8(44):77028–77040. doi:10.18632/oncotarget.20332
44. Zhang WC, Chin TM, Yang H, et al. Tumour-initiating cell-specific miR-1246 and miR-1290 expression converge to promote non-small cell lung cancer progression. Nat Commun. 2016;7(1):11702. doi:10.1038/ncomms11702
45. Kim G, An H-J, Lee M-J, et al. Hsa-miR-1246 and hsa-miR-1290 are associated with stemness and invasiveness of non-small cell lung cancer. Lung Cancer. 2016;91:15–22. doi:10.1016/j.lungcan.2015.11.013
46. Yuan D, Xu J, Wang J, et al. Extracellular miR-1246 promotes lung cancer cell proliferation and enhances radioresistance by directly targeting DR5. Oncotarget. 2016;7(22):32707–32722. doi:10.18632/oncotarget.9017
47. Bott A, Erdem N, Lerrer S, et al. miRNA-1246 induces pro-inflammatory responses in mesenchymal stem/stromal cells by regulating PKA and PP2A. Oncotarget. 2017;8(27):43897–43914. doi:10.18632/oncotarget.14915
48. Tran D, Bergholz J, Zhang H, et al. Insulin-like growth factor-1 regulates the SIRT 1-p53 pathway in cellular senescence. Aging Cell. 2014;13(4):669–678. doi:10.1111/acel.12219
49. Henrot P, Prevel R, Berger P, et al. Chemokines in COPD: from implication to therapeutic use. Int J Mol Sci. 2019;20(11):2785. doi:10.3390/ijms20112785
50. Barwinska D, Oueini H, Poirier C, et al. AMD3100 ameliorates cigarette smoke-induced emphysema-like manifestations in mice. Am J Physiol Lung Cell Mol Physiol. 2018;315(3):L382–L386. doi:10.1152/ajplung.00185.2018
51. Skronska-Wasek W, Gosens R, Konigshoff M, et al. WNT receptor signalling in lung physiology and pathology. Pharmacol Ther. 2018;187:150–166. doi:10.1016/j.pharmthera.2018.02.009
52. Yang F, Xiong H, Duan L, et al. MiR-1246 promotes metastasis and invasion of A549 cells by targeting GSK-3β‒mediated Wnt/β-catenin pathway. Cancer Res Treat. 2019;51(4):1420–1429. doi:10.4143/crt.2018.638
53. Passiglia F, Bronte G, Castiglia M, et al. Prognostic and predictive biomarkers for targeted therapy in NSCLC: for whom the bell tolls? Expert Opin Biol Ther. 2015;15(11):1553–1566. doi:10.1517/14712598.2015.1071348
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.