")
Back to Journals » Neuropsychiatric Disease and Treatment » Volume 17
CircRNA circPTK2 Might Suppress Cancer Cell Invasion and Migration of Glioblastoma by Inhibiting miR-23a Maturation
Authors Chen W, Wang N, Lian M
Received 12 December 2020
Accepted for publication 21 July 2021
Published 21 August 2021 Volume 2021:17 Pages 2767—2774
DOI https://doi.org/10.2147/NDT.S297108
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Yuping Ning
Wei Chen, Ning Wang, Minxue Lian
Department of Neurosurgery, First Affiliated Hospital of Xi’an Jiaotong University, Xi’an City, Shanxi Province, 710061, People’s Republic of China
Correspondence: Minxue Lian
Department of Neurosurgery, First Affiliated Hospital of Xi’an Jiaotong University, No. 277 West Yanta Road, Xi’an City, Shanxi Province, 710061, People’s Republic of China
Email [email protected]
Background: CircRNA circPTK2 plays opposite roles in different cancers, while its role in glioblastoma is unknown. The aim of this study was to explore the involvement of circPTK2 in glioblastoma.
Methods: Expression of circPTK2, mature miR-23a, and premature miR-23a in paired cancer and non-cancer tissues from glioblastoma patients (n = 60) was analyzed by RT-qPCR. Pearson’s correlation coefficient was used to analyze the correlations between gene expressions. The effects of circPTK2 overexpression on miR-23a maturation were analyzed by transfecting circPTK2 expression vector into glioblastoma cells, followed by determining the expression of mature miR-23a and premature miR-23a by RT-qPCR. Transwell assays were carried out to explore the role of circPTK2 and miR-23a in regulating glioblastoma cell invasion and migration.
Results: We found that circPTK2 was downregulated in GBM and was inversely correlated with mature miR-23a, but not premature miR-23a. GBM cells transfected with circPTK2 expression vector showed significantly downregulated mature miR-23a, but not premature miR-23a. Transwell assay analysis showed that circPTK2 overexpression decreased cell invasion and migration, while miR-23a increased cell invasion and migration. Moreover, miR-23a overexpression reversed the inhibitory effects of circPTK2 overexpression on cell behaviors.
Conclusion: CircPTK2 might suppress cancer cell invasion and migration by inhibiting the maturation of miR-23a.
Keywords: glioblastoma, circPTK2, miR-23a, maturation, invasion, migration
Background
Glioblastoma (GBM) multiforme is a rare type of malignant brain tumor. However, due to its highly aggressive nature,1 it has become a common cause of cancer deaths in clinical practice.1 GBM is characterized by its dismal outcome and poor prognosis, with a 5-year survival rate of < 5% and a median survival time of only 12–15 months.2,3 Despite the advances in treatment approaches, such as temozolomide-based chemotherapy, surgery, and radiotherapy, treatment of GBM is still technically challenging, mainly due to the development of therapy resistance and infiltrative tumor growth pattern, which could lead to tumor recurrence.1,4,5 Therefore, novel therapeutic approaches are needed.
Molecular alterations play critical roles in GBM growth and metastasis.6,7 With the increased understanding of molecular pathogenesis of GBM, certain molecular pathways, such as PI3K/Akt/mTOR signaling pathway and Notch pathway, have been proven to be potential targets for the development of novel molecularly targeted GBM therapy, which aims to treat GBM by regulating related gene expression.8–10 However, molecular targeted therapy is still under research. Particularly, safe and effective targets remain lacking. Circular RNAs (circRNAs, covalently closed single RNA strands) are emerging critical players in cancer biology.11 Rather than coding proteins, circRNAs participate in cancer biology by regulating gene expression at transcriptional or translational levels.12 Therefore, circRNAs might be potential targets for targeted therapy. CircRNA circPTK2 plays opposite roles in different cancers. Many studies have certified that circular RNA could function as a tumor promoter or suppressor in the development of various cancers. Recent reports have demonstrated that circPTK2 functions as a tumor promoter in bladder cancer but serves as a tumor suppressor in non-small cell lung cancer.13–15 Even though the biological function of circPTK2 had been identified in diverse cancers, the specific role of circPTK2 in GBM is unknown. We observed a close correlation between circPTK2 and miR-23a, a critical player in GBM,16 in our preliminary microarray dataset (data not shown). We, therefore, analyzed the interactions between circPTK2 and miR-23a in GBM.
Methods
Patients and Tissue Acquisitions
This study was approved by the Ethics Committee of First Affiliated Hospital of Xi’an Jiaotong University. At this hospital, we enrolled a total of 60 GBM patients (38 males and 22 females) between June 2017 and June 2020. Their age range was 58 to 72 years, with a mean of 65.4±± 6.7 years. Because other clinical disorders and the application of any therapeutic approaches can affect the expression of certain genes, patients were excluded from any initiated therapy for any clinical disorders within 3 months prior to the admission. Patients complicated with other clinical disorders were also excluded. Recurrent GBM cases and patients with a history of any malignancies were excluded. All the 60 GBM patients were diagnosed for the first time.
Prior to therapy, cancer and paired adjacent (within 3 cm around tumors) non-cancer tissue samples were collected from the 60 GBM patients using fine-needle aspiration. All samples were confirmed with histopathological analysis and stored in liquid nitrogen prior to the subsequent assays.
GBM Cell Lines and Cell Culture
Two human GBM cell lines, LN-229 and U-138, from ATCC, USA, were included and cultured in a medium composed of 90% EMEM (ATCC) and 10% FBS (ATCC) at 37°C in an incubator with 5% CO2 and 95% humidity. Both cell lines were cultured to 85% confluency prior to the subsequent assays.
Cell Transfections
To overexpress circPTK2 and miR-23a, LN-229 and U-138 cells (108) were transfected with circPTK2 expression vector or miR-23a mimic via transient transfections mediated by Lipofectamine 2000 (Invitrogen). The expression vector of circPTK2 was constructed with pcDNA3.1 (Invitrogen) as the backbone. Negative control (NC) miRNA and miR-23a mimic were purchased from Invitrogen. In each experiment, C (cells without transfection) and NC (empty vector- or NC miRNA-transfected cells) cells were included. Both cell lines were cultured in fresh media for 48h after transfection and prior to the subsequent experiments.
Preparations of RNA Samples
Total RNAs were isolated from paired tissue samples and LN-229 and U-138 cells using RNeasy Mini Kit (QIAGEN) and incubated with DNase I (Invitrogen) at 37 °C for 100 min to completely remove genomic DNA. RNA integrity was analyzed electrophoretically on 5% urea-PAGE gels, and RNA purity was examined using a NanoDrop™ 2000 spectrophotometer by determining OD260/280 ratios.
RT-qPCR Assays
cDNA samples were prepared through reverse transcriptions (RTs) with RNA samples with satisfactory integrity and an OD260/280 ratio close to 2.0 (pure RNA) as templates. All RTs were completed using a StaRT Reverse Transcription kit (AnyGenes). To determine circPTK2 expression, qPCRs were performed using SYBR Green Master Mix (Bio-Rad) with 18S rRNA as an endogenous control.
To determine the levels of mature miR-23a and premature miR-23a, RTs and qPCRs were performed using All-in-One™ miRNA qRT-PCR Reagent Kit (Genecopoeia). Prior to analyzing the mature miR-23a level, the addition of poly (A) was first performed, and ploy (T) was used as reverse primer. Sequence-specific primers were used to analyze premature miR-23a levels. Ct values of target genes were normalized to the internal control using the 2−ΔΔCt method.
Transwell Assay
To analyze cell invasion and migration, LN-229 and U-138 cells with transfections were subjected to Transwell assays using inserts (8 μm pore) from Corning. The insert membranes were coated with Corning® Matrigel® matrix for 12h at 37 °C prior to invasion assays, but not migration assays. To analyze cell migration and invasion, cells in serum-free medium were added into the upper chamber (3000 cells in 0.1 mL medium per well), and the lower chamber was filled with medium containing 20% FBS. Cells were cultured at 37 °C for 24h, and the membranes were collected. After removing cells on the upper surface of the membranes with cotton swabs, cells on the lower surface were stained using 0.5% crystal violet (Sigma-Aldrich) for 20 min and counted under a light microscope.
Statistical Analysis
Data of paired tissue samples were compared by paired t-test. Comparisons among multiple independent groups were performed using ANOVA Tukey’s test. Pearson’s correlation coefficient was applied to analyze correlations. P<0.05 was considered statistically significant.
Results
GBM Tissues Exhibited Altered Expression of circPTK2 and Mature miR-23a, but Not Premature miR-23a
Total RNAs were isolated from paired cancer and non-cancer tissues of 60 patients who suffered from glioblastoma and used to analyze the expression of circPTK2, mature miR-23a, and premature miR-23a using RT-qPCR. HemI 1.0 software was used to plot the heatmaps to represent differential gene expression. Compared to non-cancer tissues, cancer tissues exhibited downregulated circPTK2 (Figure 1A) and upregulated mature miR-23a (Figure 1B). However, no significant difference in premature miR-23a expression levels was observed between cancer and non-cancer tissues (Figure 1C).
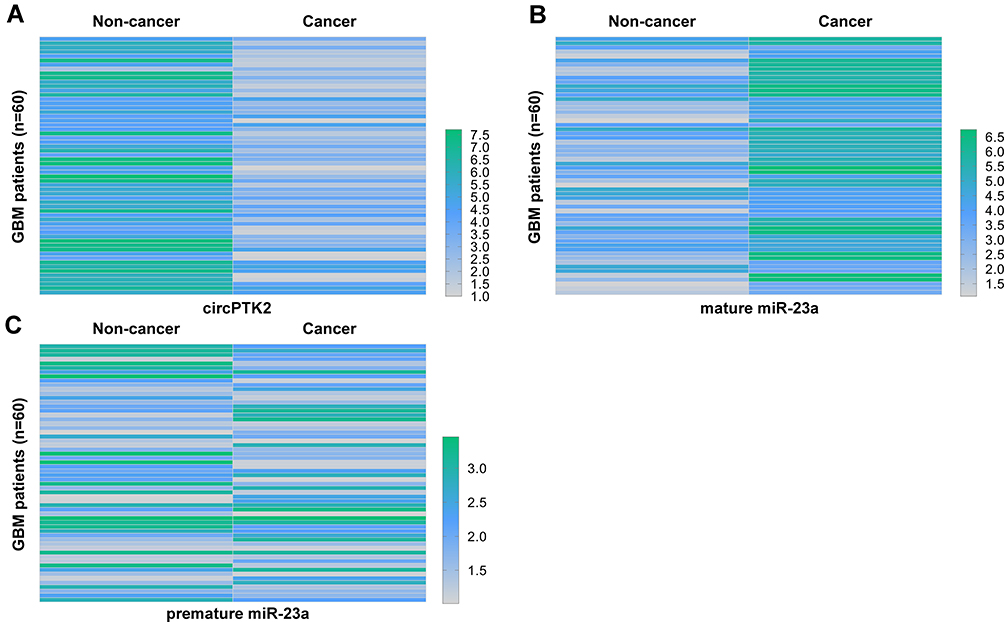 |
Figure 1 GBM tissues exhibited altered expression of circPTK2 and mature miR-23a but not premature miR-23a. Total RNAs were isolated from paired cancer and non-cancer tissues of 60 GBM patients, and the expression of circPTK2 (A), mature miR-23a (B), and premature miR-23a (C) were analyzed using RT-qPCR. HemI 1.0 software was used to plot heatmaps to represent differential gene expression. |
CircPTK2 and Mature miR-23a Were Inversely Correlated in Glioblastoma Tissues
Across cancer tissues, the correlations between circPTK2 and mature miR-23a or premature miR-23a were analyzed by Pearson’s correlation coefficient. A significant and inverse correlation was observed between circPTK2 and mature miR-23a (Figure 2A) but not between circPTK2 and premature miR-23a (Figure 2B). Therefore, the altered expression of circPTK2 in GBM might affect miR-23a maturation.
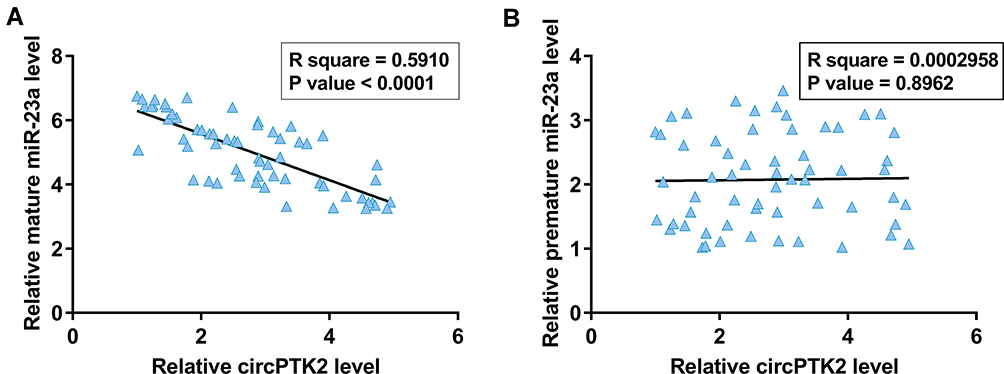 |
Figure 2 A significant and inverse correlation was observed between circPTK2 and mature miR-23a (A) but not between circPTK2 and premature miR-23a (B). |
MiR-23a Maturation Was Suppressed by circPTK2 Overexpression in Glioblastoma Cell Lines
The glioblastoma cell lines, LN-229 and U-138, were transfected with circPTK2 expression vector or miR-23a mimic, and the overexpression was checked every 24h until 96h. It was observed that circPTK2 and mature miR-23a were significantly overexpressed between 24h and 96h after transfecting circPTK2 expression vector or miR-23a mimic, respectively (Figure 3A, p<0.05). LN-229 and U-138 cells transfected with circPTK2 expression vector showed significantly downregulated mature miR-23a between 24h and 96h after transfection (Figure 3B, p<0.05), while premature miR-23a expression was not affected at each time point (Figure 3C, p<0.05). In addition, overexpression of miR-23a mimic failed to significantly alter circPTK2 expression at each time point (Figure 3D). These data suggested that miR-23a maturation in glioblastoma cells might be suppressed by circPTK2 overexpression.
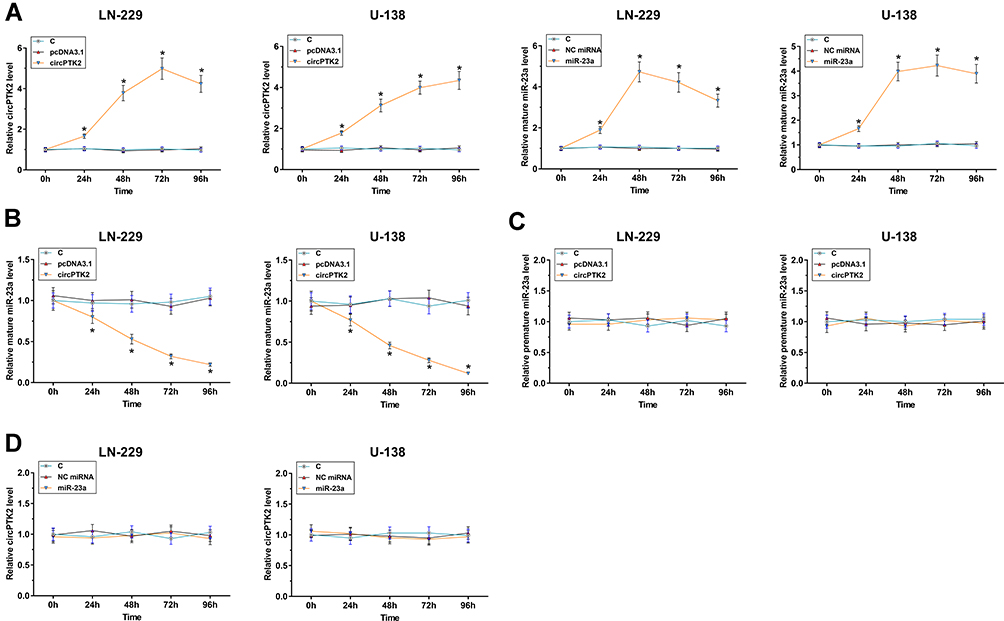 |
Figure 3 The correlations between circPTK2 and mature miR-23a or premature miR-23a across GBM tissues were analyzed by Pearson’s correlation coefficient. LN-229 and U-138 cells were transfected with circPTK2 expression vector or miR-23a mimic, and the overexpression was checked every 24h until 96h (A). The effects of circPTK2 expression vector transfection on mature miR-23a (B) and premature miR-23a (C) and the effects of miR-23a mimic transfection on the expression of circPTK2 (D) at each time point were also analyzed by RT-qPCR. *p<0.05. |
CircPTK2 Overexpression Suppressed GBM Cell Invasion and Migration by Inhibiting miR-23a Maturation
To examine the role of circPTK2 in regulating glioblastoma cell invasion and migration, we performed Transwell assay using LN-229 and U-138 cell lines. Our data showed that circPTK2 overexpression decreased cell invasion and migration, while miR-23a overexpression increased cell invasion (Figure 4A, p<0.05) and migration (Figure 4B, p<0.05). Moreover, miR-23a overexpression reversed the inhibitory effects of circPTK2 overexpression on cell behaviors.
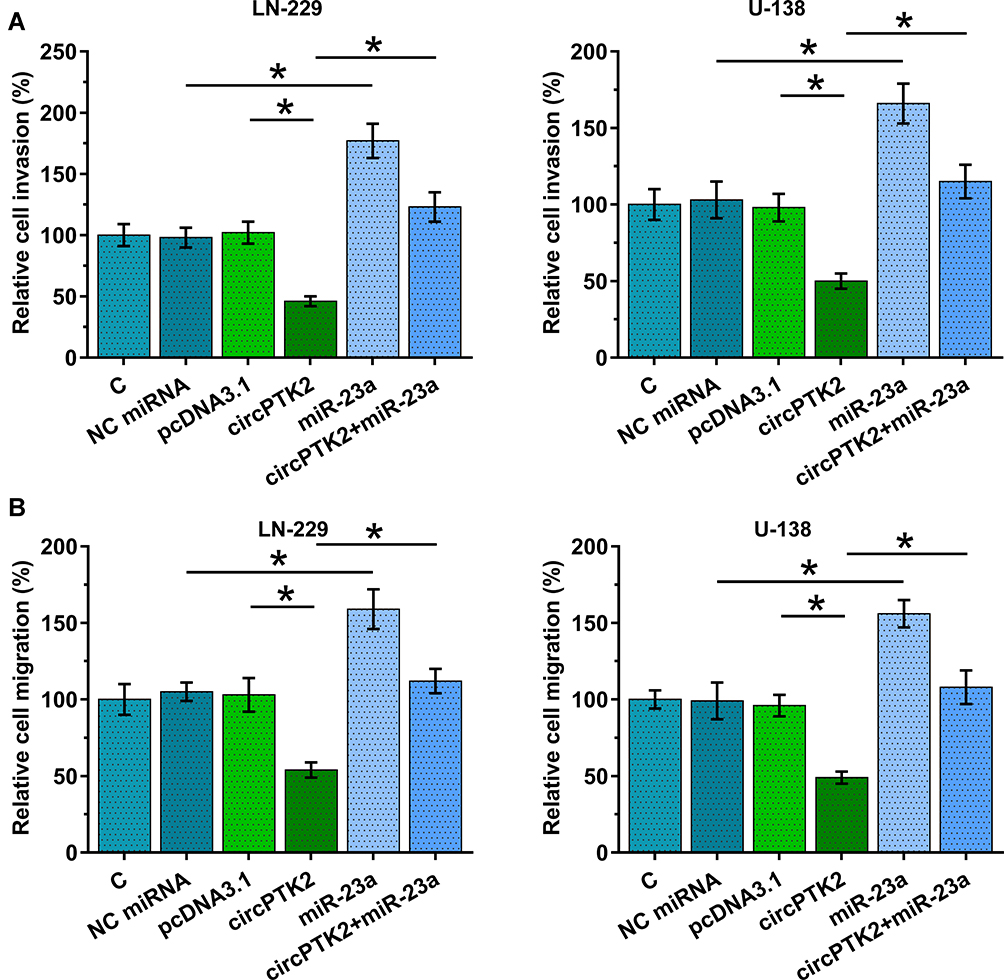 |
Figure 4 CircPTK2 overexpression suppressed GBM cell invasion and migration via miR-23a. Transwell assay was performed to analyze the role of circPTK2 (A) and miR-23a (B) in regulating the invasion and migration of LN-229 and U-138 cells. *p<0.05. |
Discussion
This study analyzed the potential involvement of circPTK2 in GBM and explored its interactions with miR-23a. Our data showed that circPTK2 was under-expressed in GBM and could decrease miR-23a maturation to suppress cancer cell invasion and migration.
Interestingly, previous studies have reported the opposite roles of circPTK2 in different types of cancers.13–15 In non-small cell lung cancer, circPTK2 is under-expressed, and its overexpression suppresses tumor metastasis and TGF-β-induced epithelial-mesenchymal transition (EMT) by controlling TIF1γ.13 In contrast, circPTK2 is overexpressed in bladder cancer and promotes cancer cell migration and proliferation.14 Similarly, circPTK2 overexpression was also observed in colorectal cancer, in which circPTK2 binds vimentin to suppress EMT.15 Therefore, previously reported functions of circPTK2 cannot be used to speculate its function in other cancers. In this study, we observed the downregulation of circPTK2 in GBM, as well as its inhibitory effects on cancer cell invasion and migration, suggesting the tumor-suppressive roles of circPTK2 in GBM. Therefore, overexpression of circPTK2 might serve as a potential target to suppress GBM metastasis. However, in vivo experiments are needed to verify our conclusions.
MiR-23a also plays different roles in different cancers.16,17 For instance, miR-23a is under-expressed in osteosarcoma and targets RUNX2 and CXCL12 to suppress cancer cell invasion and migration.17 In contrast, miR-23a is upregulated in GBM and increases cancer cell invasion through HOXD10-regulated glial-mesenchymal transition.16 Consistently, we also observed the upregulation of mature miR-23a in GBM and its enhancing effects on cancer cell invasion and migration. However, premature miR-23a was not significantly altered in GBM. Therefore, the increased miR-23a maturation might promote GBM.
Previous studies have shown that circRNAs could interact with miRNAs by serving as their molecular sponges.11,12 However, no promising binding site of miR-23a was found in circPTK2. Instead, circPTK2 overexpression in GBM cells suppressed miR-23a maturation. Therefore, we speculate that circPTK2 downregulation in GBM might increase miR-23a maturation, thereby increasing cancer cell invasion and migration. Because the formation of mature miRNAs requires transportation of premature miRNAs from the nucleus to the cytoplasm, circPTK2 might suppress the movement of premature miR-23a. The other possible reason is that the maturation of miRNAs relies heavily on efficient work of the core microprocessor complex, namely, the intranuclear RNase III enzyme Drosha and the double-stranded RNA-binding protein DGCR8. A previous study has demonstrated that circLONP2 indirectly recruit DGCR8/Drosha microprocessor through DDX1 to facilitate the maturation processing of pri-miR-17 in regulating colorectal carcinoma invasion and metastasis.18 Whether CircPTK2 modulates miRNA biogenesis by inhibiting the miRNA biogenesis pathway needs to be further explored.
Conclusion
CircPTK2 is under-expressed in glioblastoma and might suppress cancer cell invasion and migration by inhibiting miR-23a maturation.
Availability of Supporting Data
The data that support the findings of this study are not publicly available due to their containing information that could compromise the privacy of research participants but available on request from the corresponding author: Minxue Lian, Department of Neurosurgery, First Affiliated Hospital of Xi’an Jiaotong University, No. 277 West Yanta Road, Xi’an City, Shaanxi Province, 710061, P. R. China. E-mail:[email protected].
Ethical Approval and Consent to Participate
Informed consent was obtained from all individual participants included in the study. All producers were approved by the Ethics Committee of the First Affiliated Hospital of Xi’an Jiaotong University. Procedures operated in this research were completed in keeping with the standards set out in the Announcement of Helsinki and laboratory guidelines of research in China.
Acknowledgments
We thank Shaanxi International Cooperation Project for approval.
Author Contributions
Wei Chen, Minxue Lian: study concepts, literature research, clinical studies, data analysis, experimental studies, manuscript writing and review; Ning Wang: study design, literature research, experimental studies, and manuscript editing. All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was supported by the Shaanxi International Cooperation Project (Grant No. S2017-ZDYF-GJHZ-MSXM-0115).
Disclosure
All authors have no conflicts of interest. We declare that we do not have any commercial or associative interest that represents a conflict of interest in connection with the work submitted.
References
1. Batash R, Asna N, Schaffer P, et al. Glioblastoma multiforme, diagnosis and treatment; recent literature review. Curr Med Chem. 2017;24(27):3002–3009. doi:10.2174/0929867324666170516123206
2. Raj R, Seppä K, Luostarinen T, et al. Disparities in glioblastoma survival by case volume: a nationwide observational study. J Neurooncol. 2020;147(2):361–370. doi:10.1007/s11060-020-03428-5
3. Delgado-López PD, Corrales-García EM. Survival in glioblastoma: a review on the impact of treatment modalities. Clin Transl Oncol. 2016;18(11):1062–1071. doi:10.1007/s12094-016-1497-x
4. Lee CY. Strategies of temozolomide in future glioblastoma treatment. Onco Targets Ther. 2017;10:265–270. doi:10.2147/OTT.S120662
5. Nam JY, De Groot JF. Treatment of glioblastoma. J Oncol Pract. 2017;13(10):629–638. doi:10.1200/JOP.2017.025536
6. Yan D, Hao C, Xiao-feng L, et al. Molecular mechanism of Notch signaling with special emphasis on microRNAs: implications for glioma. J Cell Physiol. 2019;234(1):158–170. doi:10.1002/jcp.26775
7. Wang X, Liu S, Shao Z, et al. Bioinformatic analysis of the potential molecular mechanism of PAK7 expression in glioblastoma. Mol Med Rep. 2020;22(2):1362–1372. doi:10.3892/mmr.2020.11206
8. Festuccia C, Biordi AL, Tombolini V, et al. Targeted molecular therapy in glioblastoma. J Oncol. 2020;2020:5104876. doi:10.1155/2020/5104876
9. Li X, Wu C, Chen N, et al. PI3K/Akt/mTOR signaling pathway and targeted therapy for glioblastoma. Oncotarget. 2016;7(22):33440–33450. doi:10.18632/oncotarget.7961
10. Le Rhun E, Preusser M, Roth P, et al. Molecular targeted therapy of glioblastoma. Cancer Treat Rev. 2019;80:101896. doi:10.1016/j.ctrv.2019.101896
11. Vo JN, Cieslik M, Zhang Y, et al. The landscape of circular RNA in cancer. Cell. 2019;176(4):869–881. e13. doi:10.1016/j.cell.2018.12.021
12. Zhao ZJ, Shen J. Circular RNA participates in the carcinogenesis and the malignant behavior of cancer. RNA Biol. 2017;14(5):514–521. doi:10.1080/15476286.2015.1122162
13. Wang L, Tong X, Zhou Z, et al. Circular RNA hsa_circ_0008305 (circPTK2) inhibits TGF-β-induced epithelial-mesenchymal transition and metastasis by controlling TIF1γ in non-small cell lung cancer. Mol Cancer. 2018;17(1):140. doi:10.1186/s12943-018-0889-7
14. Xu ZQ, Yang MG, Liu HJ, et al. Circular RNA hsa_circ_0003221 (circPTK2) promotes the proliferation and migration of bladder cancer cells. J Cell Biochem. 2018;119(4):3317–3325. doi:10.1002/jcb.26492
15. Yang H, Li X, Meng Q, et al. CircPTK2 (hsa_circ_0005273) as a novel therapeutic target for metastatic colorectal cancer. Mol Cancer. 2020;19(1):13. doi:10.1186/s12943-020-1139-3
16. Yachi K, Tsuda M, Kohsaka S, et al. miR-23a promotes invasion of glioblastoma via HOXD10-regulated glial-mesenchymal transition. Signal Transduct Target Ther. 2018;3:33. doi:10.1038/s41392-018-0033-6
17. He Y, Meng C, Shao Z, et al. MiR-23a functions as a tumor suppressor in osteosarcoma. Cell Physiol Biochem. 2014;34(5):1485–1496. doi:10.1159/000366353
18. Han K, Wang F, Cao C, et al. CircLONP2 enhances colorectal carcinoma invasion and metastasis through modulating the maturation and exosomal dissemination of microRNA-17. Mol Cancer. 2020;19(1):60. doi:10.1186/s12943-020-01184-8
 © 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.