Back to Journals » Nature and Science of Sleep » Volume 17

Circadian Rhythm Dysfunction in Neurodegenerative Diseases: A Bidirectional Perspective and Therapeutic Potential
Received 25 August 2025
Accepted for publication 2 November 2025
Published 19 November 2025 Volume 2025:17 Pages 2969—2989
DOI https://doi.org/10.2147/NSS.S561326
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. Ahmed BaHammam
Dhondup Namgyal,1,2 Chae-Seok Lim1
1Department of Pharmacology, Institute of Wonkwang Medical Science, Wonkwang University School of Medicine, Iksan, Jeonbuk, 54538, Republic of Korea; 2Center for Craniofacial Molecular Biology, University of Southern California, Los Angeles, CA, 90033, USA
Correspondence: Chae-Seok Lim, Department of Pharmacology, Institute of Wonkwang Medical Science, Wonkwang University School of Medicine, Iksan, Jeonbuk, 54538, Republic of Korea, Tel +82 63 850 6765, Email [email protected]
Abstract: Disruption of circadian rhythms is a recognized hallmark of age-related neurodegenerative diseases such as Alzheimer’s disease (AD), Parkinson’s disease (PD), and Huntington’s disease (HD). Emerging evidence suggests these disruptions are not merely symptoms but potential causal factors that, in some cases, manifest prior to clinical onset. This points to a bidirectional relationship in which neurodegenerative processes and circadian dysfunction mutually exacerbate each other. Core clock genes, including BMAL1, PER, and CRY, regulate critical processes such as redox balance, mitochondrial function, and neuroinflammation, which are commonly disrupted in neurodegenerative conditions. Although molecular pathways involving altered protein homeostasis, immune dysregulation, and inflammatory processes are proposed, the precise mechanisms linking circadian rhythm disruptions to neurodegeneration remain unclear. This review provides an integrated overview of shared circadian rhythm disruptions observed in major neurodegenerative diseases and evidence on the underlying molecular mechanisms including oxidative stress and clock gene perturbation, and evaluates the temporal dynamics of circadian disruption relative to disease onset and progression. Furthermore, we discuss the translational potential of circadian-oriented interventions and highlight the limitations of current evidence. Understanding these interactions may help identify novel therapeutic strategies for stabilizing circadian rhythm to mitigate disease progression in neurodegenerative diseases.
Keywords: circadian rhythm, neurodegenerative diseases, suprachiasmatic nucleus, Circadian-oriented interventions
Introduction
The development of public healthcare systems has significantly increased global life expectancy. However, this advancement has coincided with the prevalence of neurodegenerative diseases, including Alzheimer’s disease (AD), Parkinson’s disease (PD), Huntington’s disease (HD) and other dementias. These disorders, which primarily affect older adults, are characterized by progressive loss of neuronal structure and function, resulting in cognitive, behavioral, and motor impairments. For instance, AD, the most prevalent form of dementia, is characterized by the accumulation of β-amyloid (Aβ) plaques and tau tangles, which disrupt synaptic function and lead to neuronal death.1 PD is caused by the degeneration of dopaminergic neurons in the substantia nigra and manifests as motor symptoms such as tremors, bradykinesia (slowness of movement), and muscle stiffness, together with non-motor symptoms like sleep disturbances, mood disorders, and cognitive impairment.2 HD is linked to a mutation in the HTT gene, which results in abnormal repetition of DNA sequences (CAG repeats). This mutation causes neuronal degeneration in the basal ganglia, leading to involuntary movements, cognitive decline, depression, and anxiety.3
Circadian rhythms are intrinsic 24-hour cycles that regulate physiological processes such as sleep-wake cycles, hormone release, body temperature, and cognitive function and are regulated by the suprachiasmatic nucleus (SCN) of the brain. The SCN, located anterior to the hypothalamus and dorsal to the optic chiasm, synchronizes peripheral clocks across the body through synaptic and diffusible signals. Light exposure signals are relayed to the SCN via the retinohypothalamic tract and synchronize the internal circadian rhythm with environmental stimuli.4–6 This coordination is critical for the maintenance of local circadian rhythm in peripheral organs such as the lungs, liver, and heart.7 Circadian rhythms also play a vital role in sustaining cognitive function and overall brain health. Disruptions in these rhythms, caused by irregular sleep patterns or exposure to artificial light at night, are associated with an increased risk of neurodegenerative diseases, mood disorders, and cognitive decline, particularly in aged populations.5,8
At the molecular level, the circadian rhythm is regulated by feedback loops involving core clock genes such as BMAL1, CLOCK, PERIOD (PER), and CRYPTOCHROME (CRY). BMAL1 and CLOCK heterodimers promote CRY and PER transcription, but the resulting PER-CRY complexes inhibit BMAL1-CLOCK activity through a feedback loop. Additional regulation occurs at the post-transcriptional level via kinases such as casein kinase 1ε/δ (CK1ε/δ).9 Furthermore, other clock genes modulate BMAL1 expression: RORα promotes its expression, while REV-ERBα represses it.10 The rhythmic expression of these clock genes influences various cellular processes, including cell division, metabolism, and oxidative stress11,12 (Figure 1).
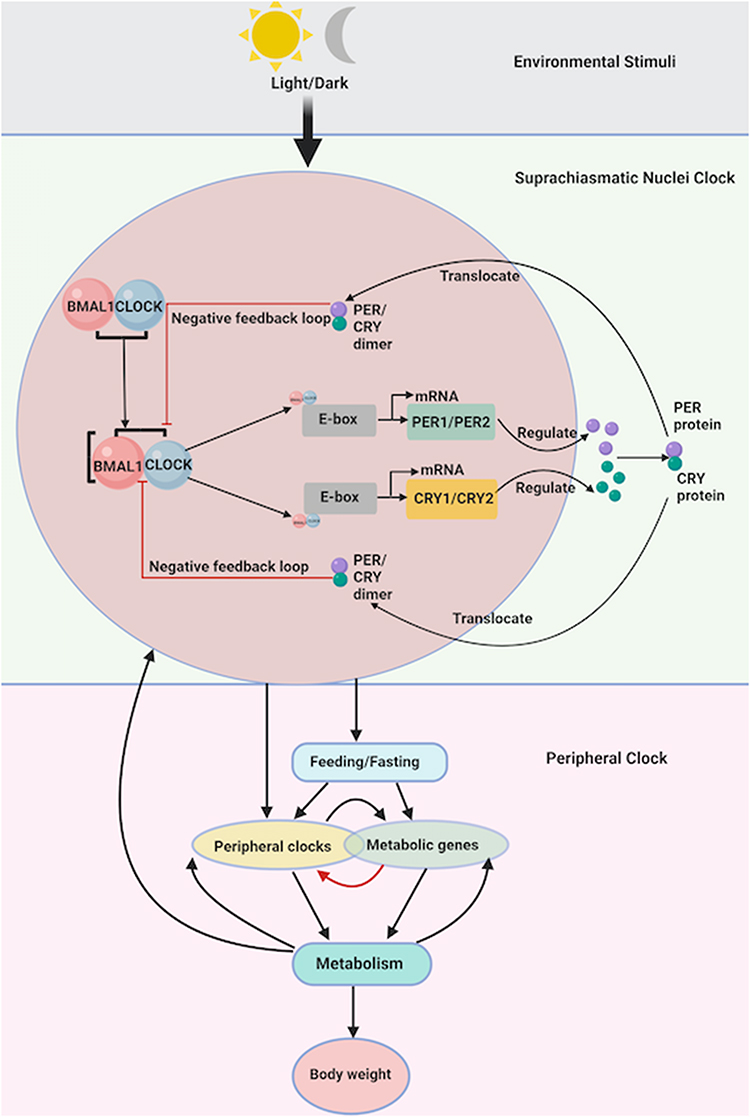 |
Figure 1 Molecular and systemic regulation of the circadian clock and its impact on metabolism and neurodegenerative disease. The circadian rhythm is orchestrated by the central pacemaker located in the suprachiasmatic nucleus (SCN) of the hypothalamus, which receives environmental light/dark cues via the retinohypothalamic tract. These cues regulate the expression of core clock genes including BMAL1, CLOCK, PER, and CRY within the SCN. BMAL1 and CLOCK form heterodimers that bind to E-box elements to initiate the transcription of PER and CRY genes. The translated PER and CRY proteins form dimers that translocate back into the nucleus and inhibit BMAL1/CLOCK activity, forming a transcription-translation feedback loop. This molecular clock drives rhythmic output signals that synchronize peripheral clocks located in organs such as the liver, lungs, and heart. Peripheral clocks are influenced by feeding/fasting cycles and regulate metabolic gene expression, impacting systemic metabolism and ultimately affecting body weight. Created in BioRender. Namgyal, (D) (2025). https://BioRender.com/zjhagzh. |
Disruption of circadian rhythms is linked to many health issues, including cancer, metabolic disorders, and mood disorders like depression and bipolar disorder.13–15 Many studies also have demonstrated the significant role of circadian disruptions in the development of motor and cognitive symptoms in neurodegenerative diseases.16–18 Mutations in Per and Bmal1 genes have been shown to accelerate aging and tissue degeneration in mice, resulting in disrupted cognitive behaviors and reduced lifespan,19,20 while circadian disruption in humans is associated with worsened cognitive and behavioral symptoms.
In this review, we summarize findings on the links between circadian rhythm disruptions and neurodegenerative diseases and explore circadian rhythm-oriented therapies such as bright light therapy, timed melatonin supplementation, time-restricted feeding and scheduled physical activity as promising non-pharmacological strategies to support brain health and delay disease progression.
Methodology
A systematic literature search was conducted in PubMed/MEDLINE, Scopus, Embase, and Web of Science covering studies published on neurodegenerative diseases and circadian rhythm disruptions/dysfunctions between January 2000 and September 2025. Search terms included combinations of circadian rhythm, sleep-wake cycle, clock genes, neurodegeneration, Alzheimer’s disease, Parkinson’s disease, Huntington’s disease, and therapeutics. Inclusion criteria were studies that examined circadian dysfunction in neurodegenerative diseases, described bidirectional mechanisms with disease pathology, or evaluated therapeutic strategies targeting circadian rhythms. Only peer-reviewed, English-language articles were included. Exclusion criteria were studies limited to non-neurodegenerative disorders, conference abstracts, editorials, commentaries, case reports without mechanistic or therapeutic relevance, doctoral dissertations without primary data, and papers lacking direct relevance to circadian biology. The literature selection process involved initial title and abstract screening, followed by full-text evaluation of potentially relevant papers. Disagreements between reviewers during the screening process were resolved through discussion and consensus.
Disrupted Circadian Rhythm in Neurodegenerative Diseases
Circadian rhythms regulate numerous physiological processes, including sleep-wake cycles, core body temperature (CBT), hormone secretion, and blood pressure, oscillating about a 24 hour period. These rhythms often become disrupted in neurodegenerative diseases, significantly impacting patient behavior and physiology.21 Such circadian disruptions are particularly prevalent in older people, coinciding with the increased incidence of age-related neurodegenerative disorders.22 While these associations are well documented, many of the underlying studies are based on relatively small cohorts and observational designs, limiting generalizability. In addition, contradictory findings have been reported across patient populations and disease stages, suggesting that circadian disruption may not manifest uniformly in all neurodegenerative conditions.
In mammals, systemic circadian rhythms are regulated by the suprachiasmatic nucleus (SCN), the master clock located in the anterior hypothalamus. At the cellular level, circadian rhythms are generated by transcriptional-translational feedback loops involving core clock genes such as BMAL1, CLOCK, PERIOD (PER), and CRYPTOCHROME (CRY). These genes regulate the rhythmic expression of other genes in a tissue-specific manner, influencing processes like metabolism, immune function, and cellular repair.23,24 Neurodegenerative diseases often disrupt this precise regulation, but detailed insights into specific alterations of clock gene and protein expression in these conditions remain incomplete. For example, while some studies in Alzheimer’s disease (AD) report loss of rhythmic BMAL1 expression,25,26 others find preserved but phase-shifted rhythms,27,28 reflecting heterogeneity and methodological variation across studies.
Given the overlapping features of circadian disruptions and neurodegenerative symptoms, it is essential to distinguish between circadian rhythm disruptions directly contributing to disease progression and those that are secondary outcomes. Failure to separate correlation (circadian disruption occurring alongside disease) from causation (circadian dysfunction actively driving disease pathology) has been a major limitation of much of the existing research. Current evidence remains predominantly correlational, and only a handful of longitudinal or interventional studies support causality. Such differentiation can aid in identifying biomarkers for disease diagnosis and designing therapeutic strategies. Common circadian disruptions in neurodegenerative diseases include severe disruptions in sleep-wake cycles, CBT regulation, hormone secretion, mood, and behavior, as well as aberrations in core clock gene expression.
Dysregulated Sleep-Wake Cycles in Neurodegenerative Diseases
Humans spend approximately one-third of their lives sleeping, yet the importance of sleep for mental and physical health has only recently been widely recognized.29–31 Sleep can be distinguished from the awake state using electroencephalograms (EEGs), which measure brain activity. Based on these measurements, sleep is categorized into two distinct states: rapid eye movement (REM) sleep and non-rapid eye movement (NREM) sleep. In general, sleep cycles alternate between NREM and REM every 90–120 minutes, with NREM dominating earlier in the night and REM sleep becoming more prominent in the early morning.29,30
Disturbances in sleep-wake cycles are prominent circadian symptoms of neurodegenerative diseases.32,33 These conditions are often associated with fragmented nighttime sleep and increased daytime sleepiness.27,33–35 As neurodegenerative diseases progress, disruptions in sleep-wake cycles and activity levels between day and night become more pronounced. Sleep disorders in AD, for example, frequently emerge in the early stages of the disease and can be early predictors of β-amyloid (Aβ) plaque formation.36–39 However, most of these findings are derived from relatively small or cross-sectional studies, and others report inconsistent results,40,41 which makes it difficult to determine whether disrupted sleep is a causal factor of amyloid pathology or simply an early symptom. In PD, REM sleep behavior disorder and restless leg syndrome are highly prevalent.32,42 Although these disorders are strongly correlated with underlying α-synuclein pathology, the evidence for a direct causal role remains limited. Many studies lack adequate control for confounding factors such as dopaminergic medication use, disease stage, or comorbid psychiatric conditions,43–46 which complicates interpretation. Moreover, not all patients with REM sleep behavior disorder (RBD) develop PD,43,47 underscoring that correlation does not necessarily equal causation.
Sleep deprivation (SD) and sleep restriction (SR) have been widely studied in animal models, especially in rodents, and show detrimental effects on cognitive function. SD and SR impair various attentional processes, resulting in increased lapses during vigilance tests and slower reaction time.48,49 In modern society, SR and SD are increasingly common due to the prevalence of night shift work and the impact of artificial light exposure at night on sleep quality. Chronic SR, even at moderate levels, can mimic the cognitive and physiological effects of several days of acute SD, leading to significant consequences.50,51
Sleep is also crucial for memory encoding and consolidation.52 SD and SR, therefore, have profound adverse effects on learning and memory, particularly on emotional memories.53 Chronic REM sleep deprivation aggravates key pathological processes in neurodegeneration, such as the accumulation of Aβ plaques and tau protein tangles in the brain, which are critical in the pathogenesis of AD.54,55 These findings underscore the role of sleep disturbances as both contributors to and accelerators of neurodegenerative disease progression.56,57 Nevertheless, some clinical studies have failed to detect a consistent link between REM sleep disruption and tau accumulation,58,59 highlighting the importance of distinguishing correlation from causation.
Imbalanced Cortisol and Melatonin Secretion Rhythm in Neurodegenerative Diseases
The sympathetic nervous system (SNS) plays a crucial role in regulating melatonin secretion, a hormone associated with circadian rhythm and widely used to treat sleep disturbances.
Rhythmic melatonin secretion by the pineal gland is regulated by the suprachiasmatic nucleus (SCN) of the anterior hypothalamus. During nighttime, norepinephrine (NE) is released from the postganglionic sympathetic nerve terminals, stimulates arylalkylamine N-acetyltransferase (AANAT), leading to melatonin synthesis.60,61 Consequently, melatonin levels peak at night and decline in the early morning.60,62,63 Melatonin shifts endogenous circadian rhythms via a phase-response curve opposite to that of light exposure, promoting sleep, sedation, and the reduction of core body temperature (CBT).64–67
Cortisol is a glucocorticoid hormone synthesized by the adrenal glands and essential for maintaining homeostasis and responding to arousal events such as exercise, waking, and acute stress.68–70 The hypothalamic-pituitary-adrenal (HPA) axis regulates rhythmic cortisol secretion, which typically peaks in the early morning and declines throughout the day.70 Light exposure profoundly influences melatonin and cortisol levels (Figure 2A).
 |
Figure 2 Age- and disease-related alterations in circadian hormone rhythms. (A) Normal circadian rhythm of melatonin and cortisol secretion. In healthy individuals, melatonin levels rise at night, peaking during the dark phase under the control of the suprachiasmatic nucleus (SCN) and sympathetic nervous system via norepinephrine stimulation of the pineal gland. In contrast, cortisol levels—regulated by the hypothalamic-pituitary-adrenal (HPA) axis—peak in the early morning and gradually decline throughout the day. These opposing rhythms help maintain physiological homeostasis and sleep-wake regulation. (B) Disruption of circadian rhythms in aging and neurodegenerative diseases. With aging, melatonin secretion decreases and shifts to later hours, while basal cortisol levels increase and peak earlier, contributing to sleep-wake phase advances. In neurodegenerative diseases, melatonin rhythms are further dampened, and cortisol secretion becomes dysregulated. These changes exacerbate sleep disturbances and reflect broader circadian rhythm degeneration in these conditions. |
Internal circadian disruption is more common in elderly individuals due to progressive degeneration of molecular circadian mechanisms with age.71,72 Among the various circadian rhythms, melatonin and cortisol secretion rhythms exhibit significant changes in older people. Melatonin secretion decreases with age, and its peak shifts later into the night.73,74 Conversely, basal cortisol release increases, with the maximum secretion occurring earlier at night, contributing to advanced sleep-wake phases often seen in elderly people.75 Neurodegenerative diseases commonly feature disrupted melatonin secretion rhythms, with a reduced nighttime peak compared to age-matched healthy controls.76–81 In dementia patients, decreased nighttime melatonin levels correlate with increased daytime sleepiness, too.77,81,82 However, some small-scale clinical cohorts report preserved melatonin secretion in early Alzheimer’s disease (AD) stages,83,84 suggesting that hormonal disruption may be disease-stage dependent. In addition, variations in assay methods and limited sample sizes across studies may account for inconsistent findings. Alterations in cortisol rhythms vary among neurodegenerative diseases.
In healthy people, cortisol secretion peaks early in the morning and gradually declines toward the evening. While minimal rhythmic changes are observed in AD patients, alterations include increased total daily cortisol levels and altered secretion timing in Huntington’s disease (HD) and Parkinson’s disease (PD) patients33,82,85–87 (Figure 2B). Although some AD cohorts report no significant differences compared to controls,88–90 others find elevated baseline cortisol.91–93 This variability underscores methodological limitations, including heterogeneous patient populations, differing disease stages, and confounding medication effects.
Overall, most evidence linking melatonin and cortisol disruption with neurodegeneration remains correlational. While reduced melatonin is consistently associated with poor sleep quality and cognitive function, causation has not been definitively established.94–97 Similarly, altered cortisol patterns may reflect downstream stress responses rather than direct circadian dysfunction. Larger, longitudinal studies are needed to clarify whether these hormonal changes act as drivers, biomarkers, or consequences of neurodegeneration.
Core Body Temperature Rhythm Fluctuation in Neurodegenerative Diseases
Core body temperature (CBT) represents the physiological state at which internal organs and their functions operate optimally, playing a critical role in the body’s thermoregulation. Human CBT varies throughout the day, increasing during waking hours and peaking in the evening before declining at night to reach its lowest level in the early morning.98 These fluctuations are closely related to autonomic nervous system (ANS) activity and are regulated by circadian rhythms governing thermal production and cellular metabolic loss.99,100 During nighttime, CBT decreases as metabolic activity decreases, with melatonin playing a pivotal role in modulating CBT through the hypothalamic thermoregulatory center.101
CBT within the brain is homeostatically regulated and buffered against environmental heat fluxes by the skull, but it remains influenced by external conditions. This regulation is mediated through systematically organized neuronal mechanisms.102 In addition, projections from the rostral suprachiasmatic nucleus (SCN) circadian pacemaker to the preoptic area facilitate the modulation of CBT circadian rhythms.103 The SCN coordinates the daily light-dark cycle, influencing CBT rhythms.104 Light stimuli entering the retina relay information to the SCN, which transforms it into neuronal pulses to regulate CBT. These daily CBT fluctuations help synchronize cell metabolism and determine organ activity states.105
Many studies have shown the role of CBT in regulating three primary mechanisms of cellular damage: oxidative stress,106 oxygen demand overload,107 and inflammation.108 Oxidative stress is a leading cause of neurodegeneration and cellular injury.109 Notably, reduced CBT has been linked to anti-aging effects and extended lifespan, as demonstrated in caloric restricted rodent models.110 Lowering CBT may have protective effects against neurodegeneration.110 However, most of these findings stem from animal studies, often with small sample sizes and tightly controlled laboratory conditions. Whether similar protective effects apply to human populations remains unclear, and caution must be exercised when extrapolating these results.
In patients with Alzheimer’s disease (AD), a significant delay in peak CBT and reduced CBT amplitude are observed, distinguishing them from healthy age-matched controls.111,112 Parkinson’s disease (PD) patients also show reduced CBT amplitude, though the peak timing remains unaffected.113,114 Similarly, significant reductions in CBT amplitude are shown in Huntington’s disease (HD) rodent models compared to controls.115,116 However, it remains uncertain whether CBT alterations directly drive neurodegenerative processes or are secondary consequences of neuronal loss and circadian disruption. At present, evidence linking CBT fluctuations to neurodegeneration is predominantly correlational, and interventional studies directly testing whether CBT manipulation alters disease progression are lacking.
Dysregulation of Mood and Behavioral Rhythm in Neurodegenerative Diseases
From a psychological perspective, mood can be defined as an emotional state with a positive or negative valence. Mood disorders are characterized by significant fluctuations in an individual’s mood, which can adversely impact daily life activities. These disorders are primarily classified into three categories: major depressive disorder, manic disorder, and bipolar disorder.117,118 Major depressive disorder is marked by a persistently depressed mood, manic disorder by an elevated mood, and bipolar disorder by cycles of both depressive and manic states.
Alterations in circadian rhythmicity are consistently observed in mood disorders. Patients with major depressive disorder and bipolar disorder exhibit significant alterations in core body temperature (CBT), behavioral activity, and hormone secretion rhythms.118 Environmental factors such as jet lag, shift work, and seasonal changes in day length can contribute to mood disturbances.119,120 Many core clock genes are implicated in mood disorders. Treatments targeting these disorders, including social rhythm therapy, light/dark exposure therapy, and advanced sleep phase therapy, have shown effectiveness in managing symptoms.121,122 Consequently, addressing circadian disruptions may represent a crucial therapeutic approach for mood disorders.123,124 However, much of the available evidence comes from small or heterogeneous patient cohorts, and contradictory results have been reported regarding whether circadian rhythm interventions directly improve mood symptoms or act indirectly by stabilizing sleep.125,126 Furthermore, causal relationships remain unclear, as mood instability itself may disrupt circadian rhythms.118,127
As neurodegenerative diseases progress, significant mood changes and heightened emotional volatility often emerge. This phenomenon, referred to as “sundown syndrome”, is characterized by increased levels of agitation, aggression, and emotional instability, peaking in the late afternoon or early evening.128,129 Sundown syndrome is not yet formally recognized as a clinical mood disorder due to uncertainties about which behavioral features qualify and whether these disturbances occur consistently at specific times of day.130,131 Nonetheless, predictable daily patterns of behavioral and emotional disturbances have been reported in elderly people with dementia.132,133 While the underlying causes of sundown syndrome remain unclear, these emotional disturbances are not solely linked to sleep loss.134 Further investigation is required to elucidate the factors contributing to this phenomenon and to differentiate it from conventional mood disorders. Some studies attribute sundown syndrome primarily to circadian disruption,129,134,135 whereas others suggest environmental stressors, caregiver fatigue, or disease severity play larger roles.129,136,137 These contradictions emphasize the multifactorial nature of mood and behavioral dysregulation in dementia. Importantly, it remains unresolved whether circadian dysfunction causally drives sundowning, or whether both phenomena stem from advancing neurodegenerative pathology.
Molecular Mechanisms Linking Circadian Rhythms to Neurodegenerative Disease Progression
Disturbances in circadian rhythm are widely recognized in aging and across neurodegenerative disorders. However, the mechanisms linking these disturbances to disease progression remain incompletely understood. In Drosophila, the antioxidant glutathione exhibits circadian-like oscillations regulated by the Period (Per) gene. Loss of Per aggravates oxidative stress-induced damage, accelerates neurodegeneration, and shortens lifespan.138,139 Similarly, age-related reductions in Cryptochrome (Cry) gene expression, which modulates the light response, have been observed in Drosophila,140 suggesting that core clock genes like Per and Cry influence age-related neurodegenerative processes.21 Although these animal studies provide mechanistic insight, their applicability to human disease remains limited due to species-specific differences in circadian regulation. Larger comparative and translational studies are required to validate these mechanisms (Table 1).
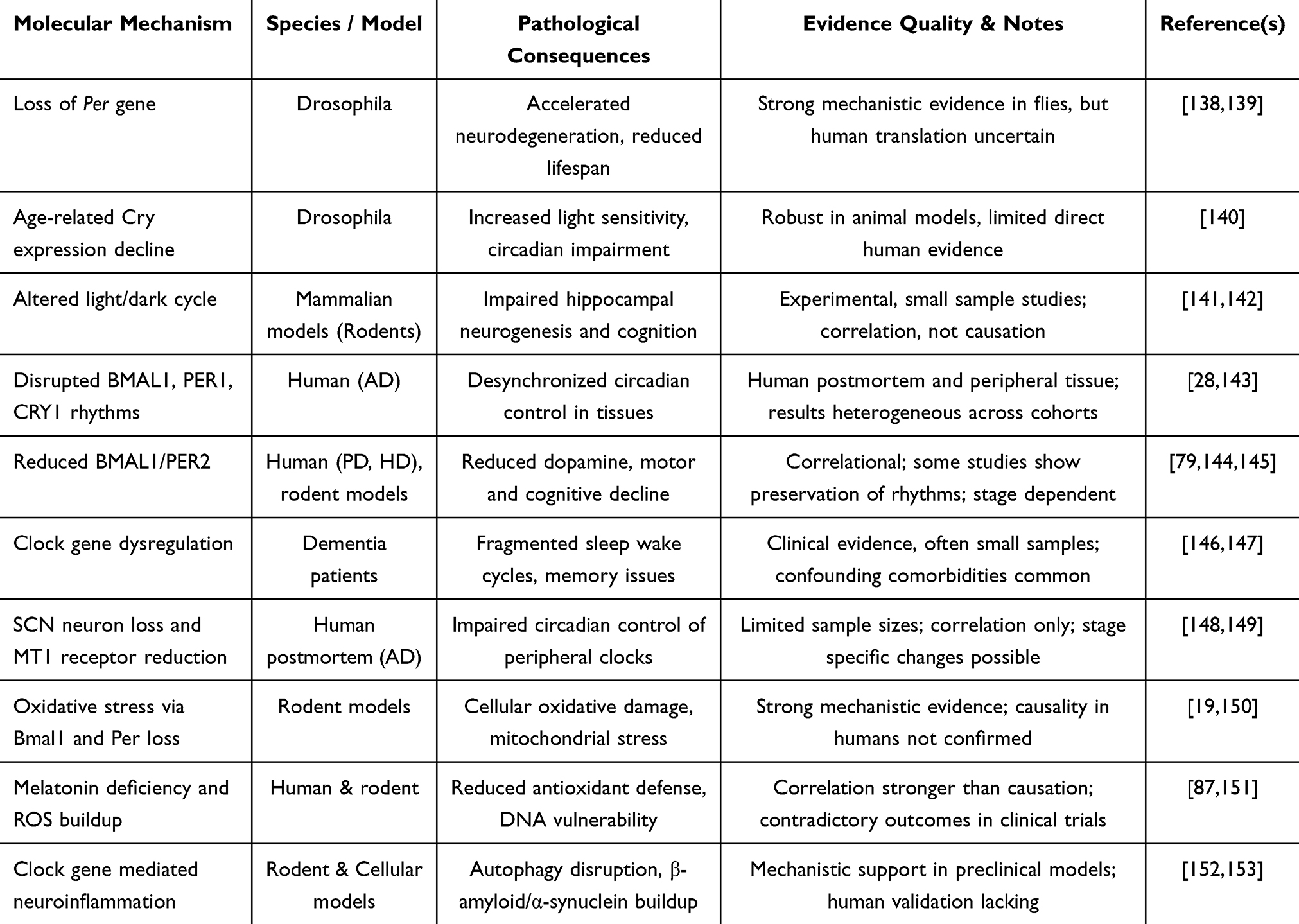 |
Table 1 Molecular Mechanisms Linking Circadian Disruption to Neurodegeneration |
In mammalian models, environmental perturbations such as altered light/dark cycles impair hippocampal neurogenesis and cognitive behaviors.141,142 Despite these insights, the precise pathways through which circadian dysfunction drives neurodegeneration remain to be elucidated.21,87
Clock gene abnormalities have been documented in patients with neurodegenerative diseases. For instance, in Alzheimer’s disease (AD), BMAL1 expression remains rhythmic in both brain and peripheral tissues but exhibits altered phase relationships.27,28 In the pineal gland of AD patients, the rhythmic expression of BMAL1, PER1, and CRY1 is absent.143 Peripheral BMAL1 transcript levels are significantly reduced,79,144 and Per2 expression is disrupted in brain areas such as the striatum, correlating with dopamine depletion145,154 in Parkinson’s disease (PD). Likewise, Huntington’s disease (HD) mouse models show disturbed Per2 expression across both the central and peripheral nervous systems.34,155 However, these findings are not entirely consistent; some studies report preserved or only mildly altered rhythmicity,156–158 suggesting that clock gene disruption may be stage-dependent or influenced by methodological variability such as tissue source, detection methods, and cohort characteristics.
Patients with dementia and neurodegenerative diseases often display excessive daytime sleepiness, fragmented activity patterns, and sleep behavior disorders.146,147,159–164 Clock genes such as BMAL1 and PER1 directly influence neurodegenerative processes, and their altered expression increases PD risk.165 Presenilin-2, which exhibits circadian rhythmicity, regulates intracellular β-amyloid levels, and disruptions in its expression are linked to AD pathology.166–168 Yet, much of this evidence remains correlational, and it is unclear whether gene disruptions are causal in driving pathology or represent downstream consequences of neurodegenerative processes.
Postmortem studies show loss of suprachiasmatic nucleus (SCN) neurons and reduced MT1 melatonin receptor expression in AD brains.148,149,169,170 In animal models, SCN dysfunction corresponds with reduced neuronal firing and impaired circadian control.115,171,172 As the SCN orchestrates circadian rhythms throughout the body, its degeneration may exacerbate neurodegenerative progression via downstream effects on peripheral oscillators.87,173 Nevertheless, postmortem studies are limited by small sample sizes and variability in disease staging, which restricts definitive conclusions about causality.
Oxidative stress—a central mechanism in neurodegeneration—is tightly coupled to circadian regulation. Core clock genes modulate intracellular levels of reactive oxygen species (ROS). For instance, Bmal1 controls antioxidant responses by transcriptional regulation of genes containing antioxidant response elements.19,152,153 Mouse models lacking Bmal1 or Per exhibit heightened oxidative damage and reduced resistance to ROS-induced stress.19,150 Melatonin, a circadian output with potent antioxidant properties, also plays a protective role by reducing free radicals and supporting mitochondrial integrity151 (Table 1). However, while animal and cellular studies strongly support this role, clinical evidence in patients remains sparse and sometimes contradictory, with trials on antioxidant benefits showing inconsistent outcomes.
Ultimately, clock gene disruption accelerates neurodegenerative pathology through interconnected pathways involving mitochondrial dysfunction, impaired autophagy, and neuroinflammation. Clock gene dysregulation disrupts redox homeostasis, leading to oxidative damage and pathological accumulation of misfolded proteins such as β-amyloid and α-synuclein.152,164 Neuroinflammatory processes, mediated by PARP1 hyperactivity and microglial activation, further exacerbate damage.138,153 Melatonin deficiency worsens this cycle by compromising antioxidant gene regulation and enhancing DNA vulnerability.87,151 These processes form a self-reinforcing feedback loop in which neurodegeneration further disrupts circadian rhythm, accelerating disease progression. While this bidirectional model is compelling, it remains largely theoretical, and empirical evidence to confirm causation in humans is limited. Novel mechanistic frameworks and integrative longitudinal studies are necessary to validate these pathways.
Circadian-Oriented Interventions in Neurodegenerative Diseases
Circadian-oriented interventions offer promising, non-invasive strategies for alleviating symptoms of neurodegenerative diseases. Approaches like bright light therapy, timed melatonin supplementation, time-restricted feeding, and scheduled physical activity have shown therapeutic potential87,155,174,175 (Figure 3 and Table 2).
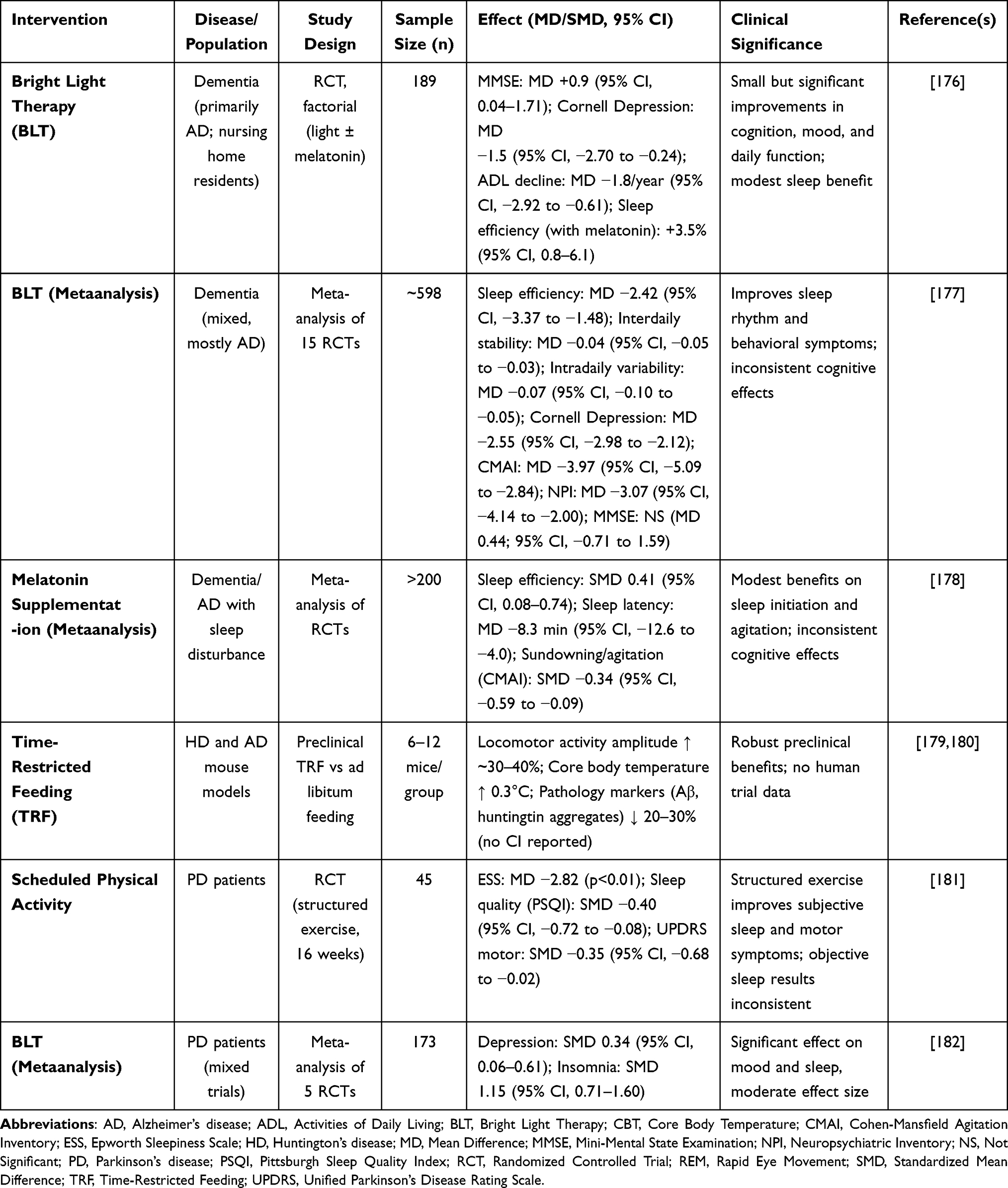 |
Table 2 Evidence Summary of Circadian-Oriented Interventions in Neurodegenerative Diseases |
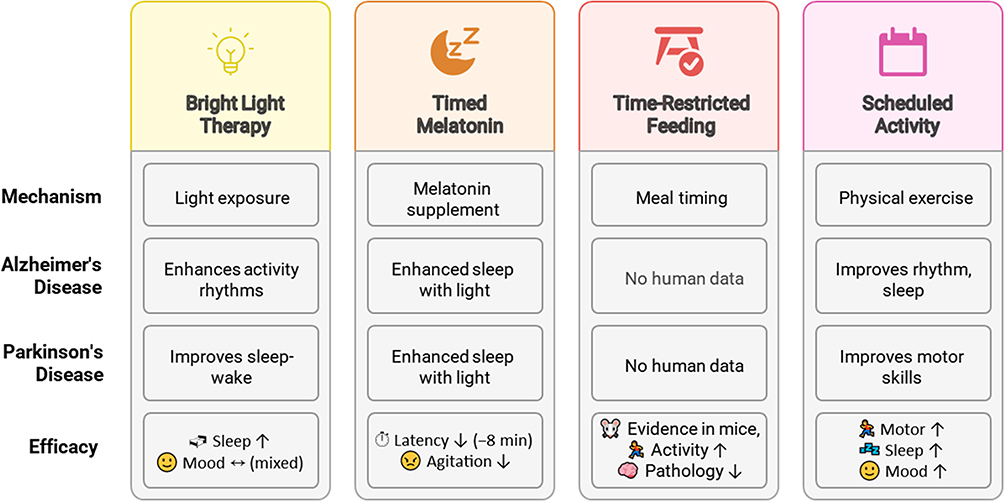 |
Figure 3 Circadian-based interventions for neurodegenerative diseases. This illustration summarizes four non-invasive strategies to improve circadian rhythm and alleviate symptoms in neurodegenerative diseases. Bright light therapy (BLT) improves daily activity rhythms in Alzheimer’s disease (AD) and stabilizes sleep-wake cycles in Parkinson’s disease (PD), with modest gains in cognition, mood, and agitation reduction, though results remain heterogeneous. Timed melatonin supplementation reduces sleep latency (−8 minutes), improves sleep efficiency, and alleviates sundowning symptoms, particularly when combined with BLT, but findings remain inconsistent across trials. Time-restricted feeding (TRF) shows benefits in Huntington’s disease (HD) mouse models by improving locomotor activity and clock gene expression yet lack human data. Scheduled physical activity functions as a non-photic zeitgeber, improving circadian amplitude, sleep quality, mood and motor performance in both animal models and PD patients. Each intervention’s effectiveness varies, and further research is needed to standardize protocols, validate long-term outcomes and clarify causal mechanisms. |
Bright Light Therapy
Bright light therapy is a commonly used circadian intervention for neurodegenerative diseases. Timed exposure to bright light has been shown to improve circadian rhythm regulation in patients.175 In AD patients, daily bright light therapy enhances the consolidation of daily activity rhythms,183,184 while in PD patients, it improves daily sleep-wake cycles.185 However, many studies are limited by small sample sizes, short follow-up periods, and variability in light intensity, duration, and timing.175,186,187 Some trials report significant improvements in sleep consolidation and mood, while others find no clear benefit, reflecting contradictory outcomes. Standardizing variables such as light intensity and timing could help establish the effectiveness of bright light therapy for neurodegenerative diseases.
Timed Melatonin Supplementation
Melatonin supplementation is another circadian therapeutic approach. Daily administration of melatonin improves sleep quality, reduces latency, and alleviates behavioral impairments in patients with sundown syndrome.174,176 Combining melatonin supplementation with bright light therapy shows even better results, significantly enhancing sleep quality and total sleep time.176,188–190 Nonetheless, outcomes across clinical trials remain inconsistent. Some randomized controlled trials report clear improvements in sleep efficiency and reduced nighttime agitation, while others find minimal or no benefit.87,191 These discrepancies may stem from differences in dosage, timing of administration, patient age, or disease stage. Current evidence supports melatonin as a potentially useful adjunct but not a definitive standalone therapy.
Time-Restricted Feeding
Time-restricted feeding (TRF) has shown therapeutic promise in mouse models of neurodegenerative diseases. Meal restriction during the light phase improves locomotor activity and core clock gene expression in Huntington’s disease (HD) mouse models.155 Similarly, restricting meals during the dark phase delays the onset of HD phenotypes and increases core body temperature (CBT) in HD mice.87,192 While these findings are robust in animal models, human clinical validation is lacking. No large-scale trials in neurodegenerative patients have yet confirmed the benefit of TRF, and translation from rodent models to human care should be cautiously approached.
Scheduled Physical Activity
Scheduled physical activity is also recognized as a circadian-aligned intervention with therapeutic potential for neurodegenerative diseases. Exercise serves as a potent non-photic zeitgeber, which is able to reinforce and realign disrupted circadian rhythms. When performed consistently at fixed times of day, physical activity can restore behavioral and physiological cycles in both animal models and humans.193–196 In rodent models of Alzheimer’s disease (AD), long-term physical activity has been shown to improve circadian rhythm amplitude and sleep quality, likely through enhanced hippocampal neurogenesis and synaptic plasticity. Similar effects were found in Parkinson’s disease (PD) models, where scheduled treadmill exercise normalized circadian locomotor activity and improved motor performance.72,196–200
In clinical studies, PD patients participating in structured exercise programs experienced improved motor performance, better sleep quality, and reduced depressive symptoms.200,201 These benefits are likely mediated by increased brain-derived neurotrophic factor (BDNF) levels, improved mitochondrial dynamics, and reduced neuroinflammatory cytokine expression-all of which intersect with circadian regulation pathways.194–196,198,202,203
While the optimal parameters-such as time of day, intensity, and duration of exercise for maximum circadian benefit-remain to be standardized, the cumulative evidence supports the role of scheduled physical activity as a cost-effective, non-invasive, and accessible therapeutic strategy. Tailoring exercise timing to individual chronotype–people’s tendencies to wake up and fall asleep at certain times of the day or disease stage may further enhance its efficacy in supporting circadian health and mitigating neurodegenerative progression.194,202
In summary, circadian-oriented interventions show promise but remain preliminary. Current evidence is often correlational, and while short-term improvements in sleep and behavior are reported, definitive proof of disease-modifying effects is lacking. Well-designed, large-scale randomized controlled trials are essential before these therapies can be considered reliable clinical strategies.
Some circadian-oriented interventions were summarized in Table 2. Across randomized controlled trials (RCTs) of bright light therapy and timed melatonin supplementation, benefits on sleep consolidation were small-to-moderate and inconsistent across cognition and motor outcomes. Scheduled physical activity appears to act as a robust non-photic zeitgeber, improving circadian amplitude and motor performance in PD, while evidence for time-restricted feeding (TRF) is strong in animal models but remains preliminary in humans. These findings highlight both the translational potential and the methodological weaknesses of current studies, underscoring the need for adequately powered, standardized RCTs with harmonized outcome measures.
A Bidirectional Model of Circadian–Neurodegeneration
Circadian disruption and neurodegenerative pathology appear to reinforce one another in a reciprocal cycle, as mentioned in the “Molecular Mechanisms” section. Loss of suprachiasmatic nucleus (SCN) neurons, reduced clock gene expression, and peripheral desynchrony can heighten oxidative stress, mitochondrial dysfunction, impaired autophagy, and neuroinflammation, thereby accelerating protein aggregation. Conversely, early pathological changes–including SCN degeneration, and downregulation of melatonin signaling–further weaken circadian amplitude and stability. This reciprocal deterioration contributes to sleep fragmentation, sundowning, REM sleep behavior disorder, and mood disturbances (Figure 4A).
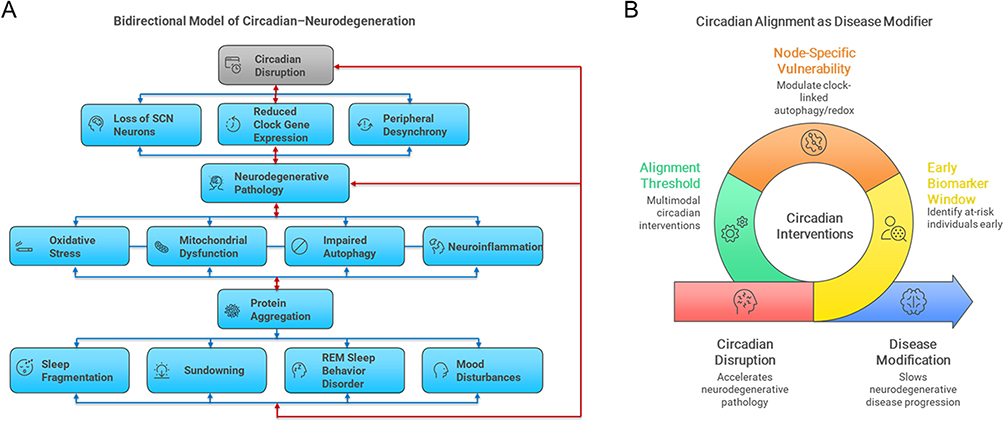 |
Figure 4 A bidirectional model of circadian–neurodegeneration. (A) Circadian disruption accelerates neurodegenerative pathology via suprachiasmatic nucleus (SCN) neuronal loss, weakened clock gene expression, and peripheral desynchrony, which heightens oxidative stress, mitochondrial dysfunction, impaired autophagy, and neuroinflammation, leading to pathological protein aggregation. In turn, early pathological changes–including SCN degeneration and impaired melatonin signaling–further weaken circadian amplitude and stability, creating a reciprocal cycle that contributes to sleep fragmentation, sundowning, REM sleep behavior disorder, and mood disturbances. (B) The model also highlights three testable predictions: (1) reduced circadian amplitude as an early biomarker window; (2) node-specific vulnerability through modulation of clock-linked autophagy and redox genes; and (3) an alignment threshold whereby multimodal interventions (light, activity, feeding) may exert supra-additive benefits. Together, panels A and B underscore circadian alignment as both a symptomatic and potentially disease-modifying strategy. |
From this conceptual framework, several testable predictions emerge (Figure 4B):
- Early biomarker window–Reduced circadian amplitude (via actigraphy or melatonin profiles) may precede clinical symptom onset in at-risk individuals.
- Node-specific vulnerability–Genetic or pharmacologic modulation of clock-linked autophagy and redox pathways could alter protein aggregation burden.
- Alignment threshold–Multimodal circadian interventions (light, activity, feeding) may produce greater benefits than single approaches.
This model highlights circadian alignment not only as a symptomatic treatment but also as a potential disease-modifying strategy, warranting further validation in larger, standardized clinical studies.
Conclusion and Perspectives
An expanding body of evidence underscores the connection between circadian rhythm disruption and the pathogenesis of neurodegenerative diseases such as Alzheimer’s disease (AD), Parkinson’s disease (PD), and Huntington’s disease (HD). Although circadian dysregulation is frequently reported in neurodegenerative diseases, whether it represents a driver or a consequence of pathology is still uncertain. In some cases, circadian disturbances may precede clinical manifestations, implying a contributory role.21,87 However, current findings are largely associative, and inconsistent outcomes across cohorts underscore the need for cautious interpretation. Recent studies point toward a reciprocal relationship between circadian dysfunction and neurodegeneration, suggesting that each can amplify the other’s progression. This bidirectional model, while conceptually strong, requires further mechanistic and clinical validation to establish its predictive and therapeutic value.
From a clinical perspective, circadian-oriented interventions, including bright light therapy, melatonin supplementation, time-restricted feeding, and scheduled physical activity, offer noninvasive therapeutic avenues for improving circadian rhythm and potentially delaying symptom progression in neurodegenerative diseases.163,175,176 However, claims regarding their disease-modifying potential must be moderated. Effect sizes across trials are often small, contradictory results have been reported, and methodological limitations (sample size, follow up duration, intervention protocols) constrain firm conclusions.
Practice Points
- Circadian-oriented interventions (bright light therapy, melatonin, time-restricted feeding, scheduled exercise) can support management of sleep, mood, and behavioral symptoms in neurodegenerative diseases.
- Simple clinical tools such as actigraphy, sleep diaries, and core body temperature monitoring can be used to assess circadian health and guide personalized care.
- Incorporating circadian strategies into routine neurology and sleep practice should emphasize individualized schedules aligned with patient chronotypes.
Future Directions
- Large, rigorously designed clinical trials are required to define optimal timing, intensity, and multimodal combinations of circadian interventions.
- Development of circadian biomarkers (eg, melatonin, cortisol, clock gene expression) may enable earlier diagnosis and more precise monitoring of treatment response.
- Mechanistic studies, including genetic, molecular, and metabolic approaches, are essential to establish causal pathways linking circadian disruption and neurodegeneration.
- Personalized circadian therapies should consider genetic variation, chronotype, and lifestyle factors to maximize efficacy and disease-modifying potential.
Abbreviations
AD, Alzheimer’s disease; PD, Parkinson’s disease; HD, Huntington’s disease; SCN, Suprachiasmatic nucleus; BMAL1, Brain and muscle ARNT-like protein 1; PER, Period gene; CRY, Cryptochrome; CLOCK, Circadian locomotor output cycles kaput; REM, Rapid eye movement; NREM, Non-rapid eye movement; ROS, Reactive oxygen species; BDNF, Brain-derived neurotrophic factor; Aβ, Amyloid beta.
Data Sharing Statement
Data sharing is not applicable to this article as no data was created or analyzed in this study.
Author Contributions
Dhondup Namgyal: conceptualization, investigation, writing - original draft; Chae-Seok Lim: conceptualization, investigation, writing - review and editing, supervision. All authors have reviewed and approved the final version of the paper; agreed on the journal this paper was submitted; and agree to be responsible for the contents of this paper.
Funding
This work was supported by the National Research Foundation of Korea (NRF) grants funded by the Korea government (MSIT) (RS-2023-NR077176, 2021R1A5A8029876) to C-SL.
Disclosure
The authors report no conflict of interest.
References
1. Rajmohan R, Reddy PH. Amyloid-beta and phosphorylated tau accumulations cause abnormalities at synapses of Alzheimer’s disease neurons. J Alzheimers Dis. 2017;57(4):975–999. doi:10.3233/jad-160612
2. DeMaagd G, Philip A. Parkinson’s disease and its management: part 1: disease entity, risk factors, pathophysiology, clinical presentation, and diagnosis. Pharm Ther. 2015;40(8):50432.
3. Irfan Z, Khanam S, Karmakar V. Pathogenesis of huntington’s disease: an emphasis on molecular pathways and prevention by natural remedies. Brain Sci. 2022;12(10). doi:10.3390/brainsci12101389
4. Welsh DK, Takahashi JS, Kay SA. Suprachiasmatic nucleus: cell autonomy and network properties. Annu Rev Physiol. 2010;72:551–577. doi:10.1146/annurev-physiol-021909135919
5. Parsons MJ, Moffitt TE, Gregory AM, et al. Social jetlag, obesity and metabolic disorder: investigation in a cohort study. Int J Obes. 2015;39(5):842–848. doi:10.1038/ijo.2014.201
6. Hastings MH, Reddy AB, Maywood ES. A clockwork web: circadian timing in brain and periphery, in health and disease. Nat Rev Neurosci. 2003;4(8):649–661. doi:10.1038/nrn1177
7. Mohawk JA, Green CB, Takahashi JS. Central and peripheral circadian clocks in mammals. Annu Rev Neurosci. 2012;35:445–462. doi:10.1146/annurev-neuro-060909-153128
8. Fishbein AB, Knutson KL, Zee PC. Circadian disruption and human health. J Clin Invest. 2021;131(19). doi:10.1172/jci148286
9. Akashi M, Tsuchiya Y, Yoshino T, Nishida E. Control of intracellular dynamics of mammalian period proteins by casein kinase I epsilon (CKIepsilon) and CKIdelta in cultured cells. Mol Cell Biol. 2002;22(6):1693–1703. doi:10.1128/mcb.22.6.1693-1703.2002
10. Ramakrishnan SN, Muscat GE. The orphan Rev-erb nuclear receptors: a link between metabolism, circadian rhythm and inflammation? Nucl Recept Signal. 2006;4:e009. doi:10.1621/nrs.04009
11. Duffield GE. DNA microarray analyses of circadian timing: the genomic basis of biological time. J Neuroendocrinol. 2003;15(10):991–1002. doi:10.1046/j.1365-2826.2003.01082.x
12. Wilking M, Ndiaye M, Mukhtar H, Ahmad N. Circadian rhythm connections to oxidative stress: implications for human health. Antioxid Redox Signal. 2013;19(2):192–208. doi:10.1089/ars.2012.4889
13. Roybal K, Theobold D, Graham A, et al. Mania-like behavior induced by disruption of CLOCK. Proc Natl Acad Sci U S A. 2007;104(15):6406–6411. doi:10.1073/pnas.0609625104
14. Lucassen EA, Coomans CP, van Putten M, et al. Environmental 24-hr cycles are essential for health. Curr Biol. 2016;26(14):1843–1853. doi:10.1016/j.cub.2016.05.038
15. Morris CJ, Purvis TE, Hu K, Scheer FA. Circadian misalignment increases cardiovascular disease risk factors in humans. Proc Natl Acad Sci U S A. 2016;113(10):E1402–11. doi:10.1073/pnas.1516953113
16. Hastings MH, Goedert M. Circadian clocks and neurodegenerative diseases: time to aggregate? Curr Opin Neurobiol. 2013;23(5):880–887. doi:10.1016/j.conb.2013.05.004
17. Videnovic A, Lazar AS, Barker RA, Overeem S. ‘The clocks that time us’--circadian rhythms in neurodegenerative disorders. Nat Rev Neurol. 2014;10(12):683–693. doi:10.1038/nrneurol.2014.206
18. Abbott SM, Videnovic A. Chronic sleep disturbance and neural injury: links to neurodegenerative disease. Nat Sci Sleep. 2016;8:55–61. doi:10.2147/nss.s78947
19. Kondratov RV, Kondratova AA, Gorbacheva VY, Vykhovanets OV, Antoch MP. Early aging and age-related pathologies in mice deficient in BMAL1, the core componentof the circadian clock. Genes Dev. 2006;20(14):1868–1873. doi:10.1101/gad.1432206
20. Krishnan N, Kretzschmar D, Rakshit K, Chow E, Giebultowicz JM. The circadian clock gene period extends healthspan in aging Drosophila melanogaster. Aging. 2009;1(11):937–948. doi:10.18632/aging.100103
21. Musiek ES. Circadian clock disruption in neurodegenerative diseases: cause and effect? Front Pharmacol. 2015;6:29. doi:10.3389/fphar.2015.00029
22. Ju YE, Lucey BP, Holtzman DM. Sleep and Alzheimer disease pathology--a bidirectional relationship. Nat Rev Neurol. 2014;10(2):115–119. doi:10.1038/nrneurol.2013.269
23. Bass J, Takahashi JS. Circadian integration of metabolism and energetics. Science. 2010;330(6009):1349–1354. doi:10.1126/science.1195027
24. Evans JA, Davidson AJ. Health consequences of circadian disruption in humans and animal models. Prog Mol Biol Transl Sci. 2013;119:283–323. doi:10.1016/b978-0-12-3969712.00010-5
25. Song H, Moon M, Choe HK, et al. Aβ-induced degradation of BMAL1 and CBP leads to circadian rhythm disruption in Alzheimer’s disease. Mol Neurodegener. 2015;10:13. doi:10.1186/s13024-015-0007-x
26. Wang Y, Lv D, Liu W, et al. Disruption of the circadian clock alters antioxidative defense via the SIRT1-BMAL1 pathway in 6-OHDA-induced models of Parkinson’s disease. Oxid Med Cell Longev. 2018;2018:4854732. doi:10.1155/2018/4854732
27. Weissová K, Bartoš A, Sládek M, Nováková M, Sumová A. Moderate changes in the circadian system of Alzheimer’s disease patients detected in their home environment. PLoS One. 2016;11(1):e0146200. doi:10.1371/journal.pone.0146200
28. Cermakian N, Lamont EW, Boudreau P, Boivin DB. Circadian clock gene expression in brain regions of Alzheimer ‘s disease patients and control subjects. J Biol Rhythms. 2011;26(2):160–170. doi:10.1177/0748730410395732
29. Fuller PM, Gooley JJ, Saper CB. Neurobiology of the sleep-wake cycle: sleep architecture, circadian regulation, and regulatory feedback. J Biol Rhythms. 2006;21(6):482–493. doi:10.1177/0748730406294627
30. Iber C, Ancoli-Israel S, Chesson AL, Quan S. The AASM manual for the scoring of sleep and associated events: rules, terminology and technical specifications. American Academy of Sleep Medicine. 2007;9(2):165–171. doi:10.1016/j.sleep.2007.02.009
31. Borbély AA, Daan S, Wirz-Justice A, Deboer T. The two-process model of sleep regulation: a reappraisal. J Sleep Res. 2016;25(2):131–143. doi:10.1111/jsr.12371
32. Barone P, Antonini A, Colosimo C, et al. The PRIAMO study: a multicenter assessment of nonmotor symptoms and their impact on quality of life in Parkinson’s disease. Mov Disord. 2009;24(11):1641–1649. doi:10.1002/mds.22643
33. Hatfield CF, Herbert J, van Someren EJ, Hodges JR, Hastings MH. Disrupted daily activity/rest cycles in relation to daily cortisol rhythms of home-dwelling patients with early Alzheimer’s dementia. Brain. 2004;127(Pt 5):1061–1074. doi:10.1093/brain/awh129
34. Morton AJ, Wood NI, Hastings MH, Hurelbrink C, Barker RA, Maywood ES. Disintegration of the sleep-wake cycle and circadian timing in Huntington’s disease. J Neurosci. 2005;25(1):157–163. doi:10.1523/jneurosci.3842-04.2005
35. Merlino G, Piani A, Gigli GL, et al. Daytime sleepiness is associated with dementia and cognitive decline in older Italian adults: a population-based study. Sleep Med. 2010;11(4):372–377. doi:10.1016/j.sleep.2009.07.018
36. Spira AP, Gamaldo AA, An Y, et al. Self-reported sleep and β-amyloid deposition in community-dwelling older adults. JAMA Neurol. 2013;70(12):1537–1543. doi:10.1001/jamaneurol.2013.4258
37. Nguyen Ho PT, Hoepel SJW, Rodriguez-Ayllon M, Luik AI, Vernooij MW, Neitzel J. Sleep, 24-hour activity rhythms, and subsequent amyloid-β pathology. JAMA Neurol. 2024;81(8):824–834. doi:10.1001/jamaneurol.2024.1755
38. Winer JR, Mander BA, Kumar S, et al. Sleep disturbance forecasts β-amyloid +accumulation across subsequent years. Curr Biol. 2020;30(21):4291–4298.e3. doi:10.1016/j.cub.2020.08.017
39. Lee MJ, Keum M, Kim JW, Suh GH, Choe YM. Relationship between disturbed sleep and longitudinal changes in cerebral beta‐amyloid accumulation in cognitively normal elderly adults. Alzheimers Dement. 2024;20(Suppl 3). doi:10.1002/alz.089892
40. Insel PS, Mohlenhoff BS, Neylan TC, Krystal AD, Mackin RS. Association of sleep and β-amyloid pathology among older cognitively unimpaired adults. JAMA Netw Open. 2021;4(7):e2117573. doi:10.1001/jamanetworkopen.2021.17573
41. Moon C, Schneider A, Cho YE, Zhang M, Dang H, Vu K. Sleep duration, sleep efficiency, and amyloid β among cognitively healthy later-life adults: a systematic review and meta-analysis. BMC Geriatr. 2024;24(1):408. doi:10.1186/s12877-024-05010-4
42. Comella CL. Sleep disorders in Parkinson’s disease: an overview. Mov Disord. 2007;22(Suppl 17):S367–73. doi:10.1002/mds.21682
43. Stefani A, Antelmi E, Arnaldi D, et al. From mechanisms to future therapy: a synopsis of isolated REM sleep behavior disorder as early synuclein-related disease. Mol Neurodegener. 2025;20(1):19. doi:10.1186/s13024-025-00809-0
44. Boeve BF, Ferman TJ. REM sleep behaviour disorder and α-synuclein. Lancet Neurol. 2024;23(12):1176–1178. doi:10.1016/s1474-4422(24)00442-3
45. Ferini-Strambi L, Carli G, Casoni F, Galbiati A. Restless legs syndrome and parkinson disease: a causal relationship between the two disorders? Front Neurol. 2018;9:551. doi:10.3389/fneur.2018.00551
46. Moccia M, Erro R, Picillo M, et al. A four-year longitudinal study on restless legs syndrome in parkinson disease. Sleep. 2016;39(2):405–412. doi:10.5665/sleep.5452
47. Xu Z, Anderson KN, Pavese N. Longitudinal studies of sleep disturbances in parkinson’s disease. Curr Neurol Neurosci Rep. 2022;22(10):635–655. doi:10.1007/s11910-022-01223-5
48. Pollak CP, Perlick D. Sleep problems and institutionalization of the elderly. J Geriatr Psychiatry Neurol. 1991;4(4):204–210. doi:10.1177/089198879100400405
49. García A, Angel JD, Borrani J, Ramirez C, Valdez P. Sleep deprivation effects on basic cognitive processes: which components of attention, working memory, and executive functions are more susceptible to the lack of sleep? Sleep Sci. 2021;14(2):107–118. doi:10.5935/1984-0063.20200049
50. Csipo T, Lipecz A, Owens C, et al. Sleep deprivation impairs cognitive performance, alters task-associated cerebral blood flow and decreases cortical neurovascular couplingrelated hemodynamic responses. Sci Rep. 2021;11(1):20994. doi:10.1038/s41598-02100188-8
51. Smith MG, Wusk GC, Nasrini J, et al. Effects of six weeks of chronic sleep restriction with weekend recovery on cognitive performance and wellbeing in high-performing adults. Sleep. 2021;44(8). doi:10.1093/sleep/zsab051
52. Klinzing JG, Niethard N, Born J. Mechanisms of systems memory consolidation during sleep. Nature Neuroscience. 2019;22(10):1598–1610. doi:10.1038/s41593-019-0467-3
53. Kim T, Kim S, Kang J, Kwon M, Lee SH. The common effects of sleep deprivation on human long-term memory and cognitive control processes. Front Neurosci. 2022;16:883848. doi:10.3389/fnins.2022.883848
54. Xiong X, Hu T, Yin Z, Zhang Y, Chen F, Lei P. Research advances in the study of sleep disorders, circadian rhythm disturbances and Alzheimer’s disease. Front Aging Neurosci. 2022;14:944283. doi:10.3389/fnagi.2022.944283
55. Kim DY, Kim SM, Han IO. Chronic rapid eye movement sleep deprivation aggravates the pathogenesis of Alzheimer’s disease by decreasing brain O-GlcNAc cycling in mice. J Neuroinflammation. 2024;21(1):180. doi:10.1186/s12974-024-03179-4
56. Niu L, Zhang F, Xu X, et al. Chronic sleep deprivation altered the expression of circadian clock genes and aggravated Alzheimer’s disease neuropathology. Brain Pathol. 2022;32(3):e13028. doi:10.1111/bpa.13028
57. Eide PK, Lashkarivand A, Pripp AH, Valnes LM, Hovd M. Mechanisms behind changes of neurodegeneration biomarkers in plasma induced by sleep deprivation. Brain Commun. 2023;5(6):fcad343. doi:10.1093/braincomms/fcad343
58. Stankeviciute L, Chhatwal JP, Levin R, et al. Amyloid beta-independent sleep markers associated with early regional tau burden and cortical thinning. Alzheimers Dement. 2024;16(3):e12616. doi:10.1002/dad2.12616
59. Li XY, Tang G, Lu J, et al. Self-reported REM Sleep behavior disorder in patients with progressive supranuclear palsy: clinical and (18)F-florzolotau PET imaging findings. Neurology. 2025;104(5):e213376. doi:10.1212/wnl.0000000000213376
60. Reiter RJ. The role of light and age in determining melatonin production in the pineal gland. In: Axelrod J, Fraschini F, Velo GP, editors. The Pineal Gland and Its Endocrine Role. Springer US; 1983:227–241.
61. Schulz P, Steimer T. Neurobiology of circadian systems. CNS Drugs. 2009;23 Suppl 2:3–13. doi:10.2165/11318620-000000000-00000
62. Brown GM. Light, melatonin and the sleep-wake cycle. J Psychiatry Neurosci. 1994;19(5):345–353.
63. Vasey C, McBride J, Penta K. Circadian rhythm dysregulation and restoration: the role of melatonin. Nutrients. 2021;13(10):3480. doi:10.3390/nu13103480
64. Lewy AJ, Ahmed S, Jackson JM, Sack RL. Melatonin shifts human circadian rhythms according to a phase-response curve. Chronobiol Int. 1992;9(5):380–392. doi:10.3109/07420529209064550
65. Doghramji K. Melatonin and its receptors: a new class of sleep-promoting agents. J Clin Sleep Med. 2007;3(5 Suppl):S17–23. doi:10.5664/jcsm.26932
66. Stone BM, Turner C, Mills SL, Nicholson AN. Hypnotic activity of melatonin. Sleep. 2000;23(5):663–669. doi:10.1093/sleep/23.5.1i
67. Kräuchi K, Wirz-Justice A. Circadian clues to sleep onset mechanisms. Neuropsychopharmacology. 2001;25(5 Suppl):S92–6. doi:10.1016/s0893-133x(01)00315-3
68. Pertovaara A, Huopaniemi T, Virtanen A, Johansson G. The influence of exercise on dental pain thresholds and the release of stress hormones. Physiol Behav. 1984;33(6):9236. doi:10.1016/0031-9384(84)90230-0
69. Payne JD, Nadel L. Sleep, dreams, and memory consolidation: the role of the stress hormone cortisol. Learn Mem. 2004;11(6):671–678. doi:10.1101/lm.77104
70. Sirianni R, Rehman KS, Carr BR, Parker CR Jr, Rainey WE. Corticotropin-releasing hormone directly stimulates cortisol and the cortisol biosynthetic pathway in human fetal adrenal cells. J Clin Endocrinol Metab. 2005;90(1):279–285. doi:10.1210/jc.2004-0865
71. Wever RA, Polásek J, Wildgruber CM. Bright light affects human circadian rhythms. Pflugers Arch. 1983;396(1):85–87. doi:10.1007/bf00584704
72. Hood S, Amir S. The aging clock: circadian rhythms and later life. J Clin Invest. 2017;127(2):437–446. doi:10.1172/jci90328
73. Sharma M, Palacios-Bois J, Schwartz G, et al. Circadian rhythms of melatonin and cortisol in aging. Biol Psychiatry. 1989;25(3):305–319. doi:10.1016/0006-3223(89)90178-9
74. Haimov I, Laudon M, Zisapel N, et al. Sleep disorders and melatonin rhythms in elderly people. BMJ. 1994;309(6948):167. doi:10.1136/bmj.309.6948.167
75. Van Cauter E, Leproult R, Plat L. Age-related changes in slow wave sleep and REM sleep and relationship with growth hormone and cortisol levels in healthy men. JAMA. 2000;284(7):861–868. doi:10.1001/jama.284.7.861
76. Mishima K, Tozawa T, Satoh K, Matsumoto Y, Hishikawa Y, Okawa M. Melatonin secretion rhythm disorders in patients with senile dementia of Alzheimer’s type with disturbed sleep-waking. Biol Psychiatry. 1999;45(4):417–421. doi:10.1016/s0006-3223(97)005106
77. Wu YH, Feenstra MG, Zhou JN, et al. Molecular changes underlying reduced pineal melatonin levels in Alzheimer disease: alterations in preclinical and clinical stages. J Clin Endocrinol Metab. 2003;88(12):5898–5906. doi:10.1210/jc.2003-030833
78. Aziz NA, Pijl H, Frölich M, et al. Delayed onset of the diurnal melatonin rise in patients with Huntington’s disease. J Neurol. 2009;256(12):1961–1965. doi:10.1007/s00415009-5196-1
79. Breen DP, Vuono R, Nawarathna U, et al. Sleep and circadian rhythm regulation in early Parkinson disease. JAMA Neurol. 2014;71(5):589–595. doi:10.1001/jamaneurol.2014.65
80. Kalliolia E, Silajdžić E, Nambron R, et al. Plasma melatonin is reduced in Huntington’s disease. Mov Disord. 2014;29(12):1511–1515. doi:10.1002/mds.26003
81. Videnovic A, Noble C, Reid KJ, et al. Circadian melatonin rhythm and excessive daytime sleepiness in Parkinson disease. JAMA Neurol. 2014;71(4):463–469. doi:10.1001/jamaneurol.2013.6239
82. Waller KL, Mortensen EL, Avlund K, et al. Melatonin and cortisol profiles in late midlife and their association with age-related changes in cognition. Nat Sci Sleep. 2016;8:47–53. doi:10.2147/nss.s75946
83. Lin CH, Chiu CC, Lane HY. Trough melatonin levels differ between early and late phases of Alzheimer disease. Clin Psychopharmacol Neurosci. 2021;19(1):135–144. doi:10.9758/cpn.2021.19.1.135
84. Nous A, Engelborghs S, Smolders I. Melatonin levels in the Alzheimer’s disease continuum: a systematic review. Alzheimers Res Ther. 2021;13(1):52. doi:10.1186/s13195-021-00788-6
85. Hartmann A, Veldhuis JD, Deuschle M, Standhardt H, Heuser I. Twenty-four hour cortisol release profiles in patients with Alzheimer’s and Parkinson’s disease compared to normal controls: ultradian secretory pulsatility and diurnal variation. Neurobiol Aging. 1997;18(3):285–289. doi:10.1016/s0197-4580(97)80309-0
86. Giubilei F, Patacchioli FR, Antonini G, et al. Altered circadian cortisol secretion in Alzheimer’s disease: clinical and neuroradiological aspects. J Neurosci Res. 2001;66(2):262–265. doi:10.1002/jnr.1219
87. Hood S, Amir S. Neurodegeneration and the circadian clock. Front Aging Neurosci. 2017;9:170. doi:10.3389/fnagi.2017.00170
88. Yao Z, Liu B, Wang Y, Dong X. High cortisol and the risk of Alzheimer disease: a protocol for systematic review and meta-analysis. Medicine. 2021;100(39):e27319. doi:10.1097/md.0000000000027319
89. Daniilidou M, Holleman J, Hagman G, et al. Neuroinflammation, cerebrovascular dysfunction and diurnal cortisol biomarkers in a memory clinic cohort: findings from the CoSTAR study. Transl Psychiatry. 2024;14(1):364. doi:10.1038/s41398-024-03072-x
90. Balietti M, Galeazzi R, Giacconi R, Santillo E, Giuli C. Early benefits with potential long-term risks of a comprehensive intervention on serum cortisol levels and cognitive performance in patients with Alzheimer’s disease. J Alzheimers Dis Rep. 2023;7(1):1445–1453. doi:10.3233/adr-230125
91. Ennis GE, An Y, Resnick SM, Ferrucci L, O’Brien RJ, Moffat SD. Long-term cortisol measures predict Alzheimer disease risk. Neurology. 2017;88(4):371–378. doi:10.1212/wnl.0000000000003537
92. Dronse J, Ohndorf A, Richter N, et al. Serum cortisol is negatively related to hippocampal volume, brain structure, and memory performance in healthy aging and Alzheimer’s disease. Front Aging Neurosci. 2023;15:1154112. doi:10.3389/fnagi.2023.1154112
93. Holleman J, Adagunodo S, Kåreholt I, et al. Cortisol, cognition and Alzheimer’s disease biomarkers among memory clinic patients. BMJ Neurol Open. 2022;4(2):e000344. doi:10.1136/bmjno-2022-000344
94. Cardinali DP. Melatonin: clinical Perspectives in Neurodegeneration. Front Endocrinol. 2019;10:480. doi:10.3389/fendo.2019.00480
95. Knezevic E, Nenic K, Milanovic V, Knezevic NN. The role of cortisol in chronic stress, neurodegenerative diseases, and psychological disorders. Cells. 2023;12(23):2726. doi:10.3390/cells12232726
96. Sumsuzzman DM, Choi J, Jin Y, Hong Y. Neurocognitive effects of melatonin treatment in healthy adults and individuals with Alzheimer’s disease and insomnia: a systematic review and meta-analysis of randomized controlled trials. Neurosci Biobehav Rev. 2021;127:459–473. doi:10.1016/j.neubiorev.2021.04.034
97. Fifel K, Videnovic A. Circadian and sleep dysfunctions in neurodegenerative disorders-an update. Front Neurosci. 2020;14:627330. doi:10.3389/fnins.2020.627330
98. Van Someren EJ. More than a marker: interaction between the circadian regulation of temperature and sleep, age-related changes, and treatment possibilities. Chronobiol Int. 2000;17(3):313–354. doi:10.1081/cbi-100101050
99. Johnson RH. The autonomic nervous system and body temperature. Proc R Soc Med. 1966;59(5):463–466. doi:10.1177/003591576605900538
100. Riganello F, Prada V, Soddu A, Di Perri C, Sannita WG. Circadian rhythms and measures of CNS/autonomic interaction. Int J Environ Res Public Health. 2019;16(13):2336. doi:10.3390/ijerph16132336
101. Zanghi BM, Gardner C, Araujo J, Milgram NW. Diurnal changes in core body temperature, day/night locomotor activity patterns, and actigraphy-generated behavioral sleep in aged canines with varying levels of cognitive dysfunction. Neurobiol Sleep Circadian Rhythms. 2016;1(1):8–18. doi:10.1016/j.nbscr.2016.07.001
102. McIlvoy L. Comparison of brain temperature to core temperature: a review of the literature. J Neurosci Nurs. 2004;36(1):23–31. doi:10.1097/01376517-200402000-00004
103. Honma K. Circadian rhythms in body temperature and sleep. Nihon Rinsho. 2013;71(12):2076–2081.
104. Drouyer E, Rieux C, Hut RA, Cooper HM. Responses of suprachiasmatic nucleus neurons to light and dark adaptation: relative contributions of melanopsin and rod-cone inputs. J Neurosci. 2007;27(36):9623–9631. doi:10.1523/jneurosci.1391-07.2007
105. Powell EW, Halberg F, Pasley JN, Lubanovic W, Ernsberger P, Scheving LE. Suprachiasmatic nucleus and circadian core temperature rhythm in the rat. Journal of Thermal Biology. 1980;5(4):189–196. doi:10.1016/0306-4565(80)90020-0
106. Mestre-Alfaro A, Ferrer MD, Banquells M, et al. Body temperature modulates the antioxidant and acute immune responses to exercise. Free Radic Res. 2012;46(6):799–808. doi:10.3109/10715762.2012.680193
107. Carlisle HJ, Ingram DL. The influence of body core temperature and peripheral temperatures on oxygen consumption in the pig. J Physiol. 1973;231(2):341–352. doi:10.1113/jphysiol.1973.sp010236
108. Tesch T, Bannert E, Kluess J, et al. Relationships between body temperatures and inflammation indicators under physiological and pathophysiological conditions in pigs exposed to systemic lipopolysaccharide and dietary deoxynivalenol. J Anim Physiol Anim Nutr. 2018;102(1):241–251. doi:10.1111/jpn.12684
109. Kim GH, Kim JE, Rhie SJ, Yoon S. The role of oxidative stress in neurodegenerative diseases. Exp Neurobiol. 2015;24(4):325–340. doi:10.5607/en.2015.24.4.325
110. Keil G, Cummings E, de Magalhães JP. Being cool: how body temperature influences ageing and longevity. Biogerontology. 2015;16(4):383–397. doi:10.1007/s10522-0159571-2
111. Satlin A, Volicer L, Stopa EG, Harper D. Circadian locomotor activity and core-body temperature rhythms in Alzheimer’s disease. Neurobiol Aging. 1995;16(5):765–771. doi:10.1016/0197-4580(95)00059-n
112. Harper DG, Volicer L, Stopa EG, McKee AC, Nitta M, Satlin A. Disturbance of endogenous circadian rhythm in aging and Alzheimer disease. Am J Geriatr Psychiatry. 2005;13(5):359–368. doi:10.1176/appi.ajgp.13.5.359
113. Pierangeli G, Provini F, Maltoni P, et al. Nocturnal body core temperature falls in Parkinson’s disease but not in multiple-system atrophy. Mov Disord. 2001;16(2):226–232. doi:10.1002/mds.1039
114. Zhong G, Bolitho S, Grunstein R, Naismith SL, Lewis SJ. The relationship between thermoregulation and REM sleep behaviour disorder in Parkinson’s disease. PLoS One. 2013;8(8):e72661. doi:10.1371/journal.pone.0072661
115. Kudo T, Schroeder A, Loh DH, et al. Dysfunctions in circadian behavior and physiology in mouse models of Huntington’s disease. Exp Neurol. 2011;228(1):80–90. doi:10.1016/j.expneurol.2010.12.011
116. Fisher SP, Black SW, Schwartz MD, et al. Longitudinal analysis of the electroencephalogram and sleep phenotype in the R6/2 mouse model of Huntington’s disease. Brain. 2013;136(Pt 7):2159–2172. doi:10.1093/brain/awt132
117. Andreasen NC. The relationship between creativity and mood disorders. Dialogues Clin Neurosci. 2008;10(2):251–255. doi:10.31887/DCNS.2008.10.2/ncandreasen
118. Walker WH 2nd, Walton JC. Circadian rhythm disruption and mental health. Transl Psychiatry. 2020;10(1):28. doi:10.1038/s41398-020-0694-0
119. Young DM. Psychiatric morbidity in travelers to Honolulu, Hawaii. Comprehensive Psychiatry. 1995;36(3):224–228. doi:10.1016/0010-440X(95)90086-B
120. Scott AJ, Monk TH, Brink LL. Shiftwork as a risk factor for depression: a pilot study. Int J Occup Environ Health. 1997;3(Supplement 2):S2–s9.
121. Benedetti F, Serretti A, Colombo C, et al. Influence of CLOCK gene polymorphism on circadian mood fluctuation and illness recurrence in bipolar depression. Am J Med Genet B Neuropsychiatr Genet. 2003;123b(1):23–26. doi:10.1002/ajmg.b.20038
122. McGrath CL, Glatt SJ, Sklar P, et al. Evidence for genetic association of RORB with bipolar disorder. BMC Psychiatry. 2009;9:70. doi:10.1186/1471-244x-9-70
123. Terman M, Terman JS. Light therapy for seasonal and nonseasonal depression: efficacy, protocol, safety, and side effects. CNS Spectr. 2005;10(8):647–63;quiz672. doi:10.1017/s1092852900019611
124. Benedetti F, Barbini B, Colombo C, Smeraldi E. Chronotherapeutics in a psychiatric ward. Sleep Med Rev. 2007;11(6):509–522. doi:10.1016/j.smrv.2007.06.004
125. Fong KN, Ge X, Ting KH, Wei M, Cheung H. The effects of light therapy on sleep, agitation and depression in people with dementia: a systematic review and meta-analysis of randomized controlled trials. Am J Alzheimers Dis Other Demen. 2023;38:15333175231160682. doi:10.1177/15333175231160682
126. Kolberg E, Hjetland GJ, Thun E, et al. The effects of bright light treatment on affective symptoms in people with dementia: a 24-week cluster randomized controlled trial. BMC Psychiatry. 2021;21(1):377. doi:10.1186/s12888-021-03376-y
127. Abbott SM, Malkani RG, Zee PC. Circadian disruption and human health: a bidirectional relationship. Eur J Neurosci. 2020;51(1):567–583. doi:10.1111/ejn.14298
128. Bachman D, Rabins P. “Sundowning” and other temporally associated agitation states in dementia patients. Annu Rev Med. 2006;57:499–511. doi:10.1146/annurev.med.57.071604.141451
129. Khachiyants N, Trinkle D, Son SJ, Kim KY. Sundown syndrome in persons with dementia: an update. Psychiatry Investig. Dec. 2011;8(4):275–287. doi:10.4306/pi.2011.8.4.275
130. Bliwise DL, Carroll JS, Lee KA, Nekich JC, Dement WC. Sleep and “sundowning” in nursing home patients with dementia. Psychiatry Res. 1993;48(3):277–292. doi:10.1016/0165-1781(93)90078-u
131. Yesavage JA, Friedman L, Ancoli-Israel S, et al. Development of diagnostic criteria for defining sleep disturbance in Alzheimer’s disease. J Geriatr Psychiatry Neurol. 2003;16(3):131–139. doi:10.1177/0891988703255684
132. Gallagher-Thompson D, Brooks JO 3rd, Bliwise D, Leader J, Yesavage JA. The relations among caregiver stress, “sundowning” symptoms, and cognitive decline inAlzheimer’s disease. J Am Geriatr Soc. 1992;40(8):807–810. doi:10.1111/j.15325415.1992.tb01853.x
133. Martin J, Marler M, Shochat T, Ancoli-Israel S. Circadian rhythms of agitation in institutionalized patients with Alzheimer’s disease. Chronobiol Int. 2000;17(3):405–418. doi:10.1081/cbi-100101054
134. Volicer L, Harper DG, Manning BC, Goldstein R, Satlin A. Sundowning and circadian rhythms in Alzheimer’s disease. Am J Psychiatry. 2001;158(5):704–711. doi:10.1176/appi.ajp.158.5.704
135. Reimus M, Siemiński M. Sundowning syndrome in dementia: mechanisms, diagnosis, and treatment. J Clin Med. 2025;14(4):1158. doi:10.3390/jcm14041158
136. Canevelli M, Valletta M, Trebbastoni A, et al. Sundowning in dementia: clinical relevance, pathophysiological determinants, and therapeutic approaches. Front Med. 2016;3:73. doi:10.3389/fmed.2016.00073
137. Xu Q, Lin FV, Liu Y, Zhao G. Bridging Gaps in Sundown Syndrome Research: a Scoping Review and Roadmap for Future multimodal approaches. Arch Clin Neuropsychol. 2025. doi:10.1093/arclin/acaf062
138. Krishnan N, Davis AJ, Giebultowicz JM. Circadian regulation of response to oxidative stress in Drosophila melanogaster. Biochem Biophys Res Commun. 2008;374(2):299–303. doi:10.1016/j.bbrc.2008.07.011
139. Beaver LM, Klichko VI, Chow ES, et al. Circadian regulation of glutathione levels and biosynthesis in Drosophila melanogaster. PLoS One. 2012;7(11):e50454. doi:10.1371/journal.pone.0050454
140. Rakshit K, Giebultowicz JM. Cryptochrome restores dampened circadian rhythms and promotes healthspan in aging Drosophila. Aging Cell. 2013;12(5):752–762. doi:10.1111/acel.12100
141. Gibson EM, Wang C, Tjho S, Khattar N, Kriegsfeld LJ. Experimental ‘jet lag’ inhibits adult neurogenesis and produces long-term cognitive deficits in female hamsters. PLoS One. 2010;5(12):e15267. doi:10.1371/journal.pone.0015267
142. Karatsoreos IN, Bhagat S, Bloss EB, Morrison JH, McEwen BS. Disruption of circadian clocks has ramifications for metabolism, brain, and behavior. Proc Natl Acad Sci U S A. 2011;108(4):1657–1662. doi:10.1073/pnas.1018375108
143. Wu YH, Fischer DF, Kalsbeek A, et al. Pineal clock gene oscillation is disturbed in Alzheimer’s disease, due to functional disconnection from the “master clock”. FASEB J. 2006;20(11):1874–1876. doi:10.1096/fj.05-4446fje
144. Cai Y, Liu S, Sothern RB, Xu S, Chan P. Expression of clock genes Per1 and Bmal1 in total leukocytes in health and Parkinson’s disease. Eur J Neurol. 2010;17(4):550–554. doi:10.1111/j.1468-1331.2009.02848.x
145. Hood S, Cassidy P, Cossette MP, et al. Endogenous dopamine regulates the rhythm of expression of the clock protein PER2 in the rat dorsal striatum via daily activation of D2 dopamine receptors. J Neurosci. 2010;30(42):14046–14058. doi:10.1523/jneurosci.212810.2010
146. Abbott RD, Ross GW, White LR, et al. Excessive daytime sleepiness and subsequent development of Parkinson disease. Neurology. 2005;65(9):1442–1446. doi:10.1212/01.wnl.0000183056.89590.0d
147. Schenck CH, Boeve BF, Mahowald MW. Delayed emergence of a parkinsonian disorder or dementia in 81% of older men initially diagnosed with idiopathic rapid eye movement sleep behavior disorder: a 16-year update on a previously reported series. Sleep Med. 2013;14(8):744–748. doi:10.1016/j.sleep.2012.10.009
148. Swaab DF, Fliers E, Partiman TS. The suprachiasmatic nucleus of the human brain in relation to sex, age and senile dementia. Brain Res. 1985;342(1):37–44. doi:10.1016/0006-8993(85)91350-2
149. Wang JL, Lim AS, Chiang WY, et al. Suprachiasmatic neuron numbers and restactivity circadian rhythms in older humans. Ann Neurol. 2015;78(2):317–322. doi:10.1002/ana.24432
150. Jang YS, Lee MH, Lee SH, Bae K. Cu/Zn superoxide dismutase is differentially regulated in period gene-mutant mice. Biochem Biophys Res Commun. 2011;409(1):22–27. doi:10.1016/j.bbrc.2011.04.099
151. Reiter RJ, Tan DX, Manchester LC, El-Sawi MR. Melatonin reduces oxidant damage and promotes mitochondrial respiration: implications for aging. Ann N Y Acad Sci. 2002;959:238–250. doi:10.1111/j.1749-6632.2002.tb02096.x
152. Kondratova AA, Kondratov RV. The circadian clock and pathology of the ageing brain. Nat Rev Neurosci. 2012;13(5):325–335. doi:10.1038/nrn3208
153. Lee J, Moulik M, Fang Z, et al. Bmal1 and β-cell clock are required for adaptation to circadian disruption, and their loss of function leads to oxidative stress-induced β-cell failure in mice. Mol Cell Biol. 2013;33(11):2327–2338. doi:10.1128/mcb.01421-12
154. Gravotta L, Gavrila AM, Hood S, Amir S. Global depletion of dopamine using intracerebroventricular 6-hydroxydopamine injection disrupts normal circadian wheel-running patterns and PERIOD2 expression in the rat forebrain. J Mol Neurosci. 2011;45(2):162. doi:10.1007/s12031-011-9520-8
155. Maywood ES, Fraenkel E, McAllister CJ, et al. Disruption of peripheral circadian timekeeping in a mouse model of Huntington’s disease and its restoration by temporally scheduled feeding. J Neurosci. 2010;30(30):10199–10204. doi:10.1523/jneurosci.169410.2010
156. Hollis HC, Sharma A, Sheehan PW, et al. Reconstructed cell-type-specific rhythms in human brain link Alzheimer’s pathology, circadian stress, and ribosomal disruption. Neuron. 2025;113(17):2822–2838.e7. doi:10.1016/j.neuron.2025.07.010
157. Cronin P, McCarthy MJ, Lim ASP, et al. Circadian alterations during early stages of Alzheimer’s disease are associated with aberrant cycles of DNA methylation in BMAL1. Alzheimers Dement. 2017;13(6):689–700. doi:10.1016/j.jalz.2016.10.003
158. Bellanti F, Iannelli G, Blonda M, et al. Alterations of clock gene RNA expression in brain regions of a triple transgenic model of Alzheimer’s disease. J Alzheimers Dis. 2017;59(2):615–631. doi:10.3233/jad-160942
159. Bonanni E, Maestri M, Tognoni G, et al. Daytime sleepiness in mild and moderate Alzheimer’s disease and its relationship with cognitive impairment. J Sleep Res. 2005;14(3):311–317. doi:10.1111/j.1365-2869.2005.00462.x
160. Tranah GJ, Blackwell T, Stone KL, et al. Circadian activity rhythms and risk of incident dementia and mild cognitive impairment in older women. Ann Neurol. 2011;70(5):722–732. doi:10.1002/ana.22468
161. Iranzo A, Tolosa E, Gelpi E, et al. Neurodegenerative disease status and post-mortem pathology in idiopathic rapid-eye-movement sleep behaviour disorder: an observational cohort study. Lancet Neurol. 2013;12(5):443–453. doi:10.1016/s1474-4422(13)70056-5
162. Postuma RB, Gagnon JF, Bertrand JA, Génier Marchand D, Montplaisir JY. Parkinson risk in idiopathic REM sleep behavior disorder: preparing for neuroprotective trials. Neurology. 2015;84(11):1104–1113. doi:10.1212/wnl.0000000000001364
163. Videnovic A, Zee PC. Consequences of Circadian Disruption on Neurologic Health. Sleep Med Clin. 2015;10(4):469–480. doi:10.1016/j.jsmc.2015.08.004
164. Mattis J, Sehgal A. Circadian rhythms, sleep, and disorders of aging. Trends Endocrinol Metab. 2016;27(4):192–203. doi:10.1016/j.tem.2016.02.003
165. Gu Z, Wang B, Zhang YB, et al. Association of ARNTL and PER1 genes with Parkinson’s disease: a case-control study of Han Chinese. Sci Rep. 2015;5:15891. doi:10.1038/srep15891
166. Wolfe MS. Presenilin and gamma-secretase: structure meets function. J Neurochem. 2001;76(6):1615–1620. doi:10.1046/j.1471-4159.2001.00245.x
167. Panda S, Antoch MP, Miller BH, et al. Coordinated transcription of key pathways in the mouse by the circadian clock. Cell. 2002;109(3):307–320. doi:10.1016/s0092-8674(02)00722-5
168. Giri M, Zhang M, Y L. Genes associated with Alzheimer’s disease: an overview and current status. Clin Interv Aging. 2016;11:665–681. doi:10.2147/cia.s105769
169. Stopa EG, Volicer L, Kuo-Leblanc V, et al. Pathologic evaluation of the human suprachiasmatic nucleus in severe dementia. J Neuropathol Exp Neurol. 1999;58(1):29–39. doi:10.1097/00005072-199901000-00004
170. Wu YH, Zhou JN, Van Heerikhuize J, Jockers R, Swaab DF. Decreased MT1 melatonin receptor expression in the suprachiasmatic nucleus in aging and Alzheimer’s disease. Neurobiol Aging. 2007;28(8):1239–1247. doi:10.1016/j.neurobiolaging.2006.06.002
171. Fahrenkrug J, Popovic N, Georg B, Brundin P, Hannibal J. Decreased VIP and VPAC2 receptor expression in the biological clock of the R6/2 Huntington’s disease mouse. J Mol Neurosci. 2007;31(2):139–148. doi:10.1385/jmn/31:02:139
172. Pallier PN, Maywood ES, Zheng Z, et al. Pharmacological imposition of sleep slows cognitive decline and reverses dysregulation of circadian gene expression in a transgenic mouse model of Huntington’s disease. J Neurosci. 2007;27(29):7869–7878. doi:10.1523/jneurosci.0649-07.2007
173. Ju YE, McLeland JS, Toedebusch CD, et al. Sleep quality and preclinical Alzheimer disease. JAMA Neurol. 2013;70(5):587–593. doi:10.1001/jamaneurol.2013.2334
174. de Jonghe A, Korevaar JC, van Munster BC, de Rooij SE. Effectiveness of melatonin treatment on circadian rhythm disturbances in dementia. Are there implications for delirium? A systematic review. Int J Geriatr Psychiatry. 2010;25(12):1201–1208. doi:10.1002/gps.2454176
175. Forbes D, Blake CM, Thiessen EJ, Peacock S, Hawranik P. Light therapy for improving cognition, activities of daily living, sleep, challenging behaviour, and psychiatric disturbances in dementia. Cochrane Database Syst Rev. 2014;2:Cd003946. doi:10.1002/14651858.CD003946.pub4
176. Riemersma-van der Lek RF, Swaab DF, Twisk J, Hol EM, Hoogendijk WJ, Van Someren EJ. Effect of bright light and melatonin on cognitive and noncognitive function in elderly residents of group care facilities: a randomized controlled trial. JAMA. 2008;299(22):2642–2655. doi:10.1001/jama.299.22.2642
177. Zang L, Liu X, Li Y, et al. The effect of light therapy on sleep disorders and psychobehavioral symptoms in patients with Alzheimer’s disease: A meta-analysis. PLoS One. 2023;18(12):e029397. doi:10.1371/journal.pone.0293977
178. Xu J, Wang LL, Dammer EB, et al. Melatonin for sleep disorders and cognition in dementia: a meta-analysis of randomized controlled trials. Am J Alzheimers Dis Other Demen. 2015;30(5):439–447. doi:10.1177/1533317514568005
179. Wang HB, Loh DH, Whittaker DS, Cutler T, Howland D, Colwell CS. Time-restricted feeding improves circadian dysfunction as well as motor symptoms in the Q175 mouse model of Huntington’s disease. eNeuro. 2018;5(1):ENEURO.0431–17.2017. doi:10.1523/eneuro.0431-17.2017
180. Whittaker DS, Akhmetova L, Carlin D, et al. Circadian modulation by time-restricted feeding rescues brain pathology and improves memory in mouse models of Alzheimer’s disease. Cell Metab. 2023;35(10):1704–1721.e6. doi:10.1016/j.cmet.2023.07.014
181. Amara AW, Wood KH, Joop A, et al. Randomized, controlled trial of exercise on objective and subjective sleep in Parkinson’s disease. Mov Disord. 2020;35(6):947–958. doi:10.1002/mds.28009
182. Lin F, Su Y, Weng Y, et al. The effects of bright light therapy on depression and sleep disturbances in patients with Parkinson’s disease: a systematic review and meta-analysis of randomized controlled trials. Sleep Med. 2021;83:280–289. doi:10.1016/j.sleep.2021.03.035
183. Ancoli-Israel S, Martin JL, Kripke DF, Marler M, Klauber MR. Effect of light treatment on sleep and circadian rhythms in demented nursing home patients. J Am Geriatr Soc. 2002;50(2):282–289. doi:10.1046/j.1532-5415.2002.50060.x
184. McCurry SM, Pike KC, Vitiello MV, Logsdon RG, Larson EB, Teri L. Increasing walking and bright light exposure to improve sleep in community-dwelling persons with Alzheimer’s disease: results of a randomized, controlled trial. J Am Geriatr Soc. 2011;59(8):1393–1402. doi:10.1111/j.1532-5415.2011.03519.x
185. Videnovic A, Klerman EB, Wang W, Marconi A, Kuhta T, Zee PC. Timed light therapy for sleep and daytime sleepiness associated with parkinson disease: a randomized clinical trial. JAMA Neurol. 2017;74(4):411–418. doi:10.1001/jamaneurol.2016.5192180
186. Paus S, Schmitz-Hübsch T, Wüllner U, Vogel A, Klockgether T, Abele M. Bright light therapy in Parkinson’s disease: a pilot study. Mov Disord. 2007;22(10):1495–1498. doi:10.1002/mds.21542
187. Rubiño JA, Gamundí A, Akaarir M, Canellas F, Rial R, Nicolau MC. Bright light therapy and circadian cycles in institutionalized elders. Front Neurosci. 2020;14:359. doi:10.3389/fnins.2020.00359
188. Asayama K, Yamadera H, Ito T, Suzuki H, Kudo Y, Endo S. Double blind study of melatonin effects on the sleep-wake rhythm, cognitive and non-cognitive functions in Alzheimer type dementia. J Nippon Med Sch. 2003;70(4):334–341. doi:10.1272/jnms.70.334
189. Medeiros CA, Carvalhedo de Bruin PF, Lopes LA, Magalhães MC, de Lourdes Seabra M, de Bruin VM. Effect of exogenous melatonin on sleep and motor dysfunction in Parkinson’s disease. A randomized, double blind, placebo-controlled study. J Neurol. 2007;254(4):459–464. doi:10.1007/s00415-006-0390-x
190. Chen Y, Hong W, Fang Y. Role of biological rhythm dysfunction in the development and management of bipolar disorders: a review. Gen Psychiatr. 2020;33(1):e100127. doi:10.1136/gpsych-2019-100127
191. Urrestarazu E, Iriarte J. Clinical management of sleep disturbances in Alzheimer’s disease: current and emerging strategies. Nat Sci Sleep. 2016;8:21–33. doi:10.2147/nss.s76706187
192. Skillings EA, Wood NI, Morton AJ. Beneficial effects of environmental enrichment and food entrainment in the R6/2 mouse model of Huntington’s disease. Brain Behav. 2014;4(5):675–686. doi:10.1002/brb3.235
193. Yamanaka Y, Hashimoto S, Tanahashi Y, Nishide SY, Honma S, Honma K. Physical exercise accelerates reentrainment of human sleep-wake cycle but not of plasma melatonin rhythm to 8-h phase-advanced sleep schedule. Am J Physiol Regul Integr Comp Physiol. 2010;298(3):R681–91. doi:10.1152/ajpregu.00345.2009
194. Memon AA, Coleman JJ, Amara AW. Effects of exercise on sleep in neurodegenerative disease. Neurobiol Dis. 2020;140:104859. doi:10.1016/j.nbd.2020.104859
195. Chen YC, Wang WS, Lewis SJG, Wu SL. Fighting against the clock: circadian disruption and Parkinson’s disease. J Mov Disord. 2024;17(1):1–14. doi:10.14802/jmd.23216
196. Weinert D, Gubin D. The impact of physical activity on the circadian system: benefits for health, performance and wellbeing. Applied Sciences. 2022;12(18):9220. doi:10.3390/app12189220
197. Hu Y, Niu L, Chen Y, et al. Voluntary wheel running exercise improves sleep disorder, circadian rhythm disturbance, and neuropathology in an animal model of Alzheimer’s disease. Alzheimers Dement. 2025;21(6):e70314. doi:10.1002/alz.70314
198. Jaberi S, Fahnestock M. Mechanisms of the beneficial effects of exercise on brainderived neurotrophic factor expression in Alzheimer’s disease. Biomolecules. 2023;13(11):1577. doi:10.3390/biom13111577
199. Hunt J, Coulson EJ, Rajnarayanan R, Oster H, Videnovic A, Rawashdeh O. Sleep and circadian rhythms in Parkinson’s disease and preclinical models. Mol Neurodegener. 2022;17(1):2. doi:10.1186/s13024-021-00504-w
200. Yalçin M, Grande V, Outeiro TF, Relόgio A. Circadian clock dysfunction in Parkinson’s disease: mechanisms, consequences, and therapeutic strategy. NPJ Parkinsons Dis. 2025;11(1):213. doi:10.1038/s41531-025-01009-9
201. Homolak J, Mudrovčić M, Vukić B, Toljan K. Circadian rhythm and Alzheimer’s disease. Med Sci. 2018;6(3). doi:10.3390/medsci6030052
202. Padilha C, Souza R, Grossl FS, Gauer APM, de Sá CA, Rodrigues-Junior SA. Physical exercise and its effects on people with Parkinson’s disease: umbrella review. PLoS One. 2023;18(11):e0293826. doi:10.1371/journal.pone.0293826
203. Hitrec T, Petit C, Cryer E, et al. Timed exercise stabilizes behavioral rhythms but not molecular programs in the brain’s suprachiasmatic clock. iScience. 2023;26(2):106002. doi:10.1016/j.isci.2023.106002
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Targeted Liposomal Co-Delivery Dopamine with 3-n-Butylphthalide for Effective Against Parkinson’s Disease in Mice Model

Liang Y, Feng L, Zheng Y, Gao Y, Shi R, Zhang Z, Ying X, Zeng Y
International Journal of Nanomedicine 2024, 19:12851-12870
Published Date: 30 November 2024
GPX3 as a Novel and Potential Therapeutic Target in the Shared Molecular Mechanisms of Traumatic Brain Injury and Parkinson’s Disease
Wang Y, Fang J, Yuan Q, Yu J, Hu J
Journal of Inflammation Research 2025, 18:1911-1928
Published Date: 7 February 2025
Exploring Potential Causal Links: How Parkinson’s Disease Brain Damage Impacts Liver Health–A Mendelian Randomization and Transcriptomic Study
Liu T, Chen Q, Wu H, Li J, Xian M, Lu K, Zhu C, Wei J
Journal of Inflammation Research 2025, 18:8173-8197
Published Date: 21 June 2025
Memory, Sleep, Dreams, and Consciousness: A Perspective Based on the Memory Theory of Consciousness
Budson AE, Paller KA
Nature and Science of Sleep 2025, 17:1957-1972
Published Date: 26 August 2025
GLP-1 Receptor Agonists in Neurological Disorders: From Mechanisms to Clinical Translation
Li P, Gao Y, Liu W
Drug Design, Development and Therapy 2026, 20:613616
Published Date: 12 June 2026