")
Back to Journals » Infection and Drug Resistance » Volume 16
Chromosome-Mediated Colistin Resistance in Clinical Isolates of Klebsiella pneumoniae and Escherichia coli: Mutation Analysis in the Light of Genetic Background
Authors Riquelme MP, Martinez RW, Brito B, García P , Legarraga P, Wozniak A
Received 24 June 2023
Accepted for publication 10 August 2023
Published 28 September 2023 Volume 2023:16 Pages 6451—6462
DOI https://doi.org/10.2147/IDR.S427398
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor Suresh Antony
María Paz Riquelme,1 Rodrigo W Martinez,2,3 Bárbara Brito,4 Patricia García,1,3,5 Paulette Legarraga,1,5 Aniela Wozniak1,3,5
1Department of Clinical Laboratories - School of Medicine, Pontificia Universidad Católica de Chile, Santiago, Chile; 2Genomics & Resistant Microbes Group (Germ) - Instituto de Ciencias e Innovación en Medicina (ICIM); School of Medicine-Clínica Alemana, Universidad del Desarrollo, Santiago, Chile; 3Millennium Nucleus for Collaborative Research on Bacterial Resistance (MICROB-R), Santiago Chile; 4Australian Institute for Microbiology & Infection - Faculty of Science, University of Technology Sydney, Sydney, Australia; 5Clinical Laboratories Network, Red de Salud UC-CHRISTUS, Santiago, Chile
Correspondence: Aniela Wozniak, Laboratory of Microbiology, Pontificia Universidad Católica de Chile, Vicuña Mackenna 4686, 3rd Floor, Santiago, 7820436, Chile, Tel +562-23548573, Fax +562-23548571, Email [email protected]; [email protected]
Purpose: Colistin resistance mechanisms involving mutations in chromosomal genes associated with LPS modification are not completely understood. Mutations in genes coding for the MgrB regulator frequently account for colistin resistance in Klebsiella pneumoniae, whereas mutations in genes coding for PhoPQ and PmrAB are frequent in E. coli. Our aim was to perform a genetic analysis of chromosomal mutations in colistin-resistant (MIC ≥ 4 μg/mL) clinical isolates of K. pneumoniae (n = 8) and E. coli (n = 7) of different STs.
Methods: Isolates were obtained in a 3-year period in a university hospital in Santiago, Chile. Susceptibility to colistin, aminoglycosides, cephalosporins, carbapenems and ciprofloxacin was determined through broth microdilution. Whole genome sequencing was performed for all isolates and chromosomal gene sequences were compared with sequences of colistin-susceptible isolates of the same sequence types.
Results: None of the isolates carried mcr genes. Most of the isolates were susceptible to all the antibiotics analyzed. E. coli isolates were ST69, ST127, ST59, ST131 and ST14, and K. pneumoniae isolates were ST454, ST45, ST6293, ST380 and ST25. All the isolates had mutations in chromosomal genes analyzed. K. pneumoniae had mutations mainly in mgrB gene, whereas E. coli had mutations in pmrA, pmrB and pmrE genes. Most of the amino acid changes in LPS-modifying enzymes of colistin-resistant isolates were found in colistin-susceptible isolates of the same and/or different ST. Eleven of them were found only in colistin-resistant isolates.
Conclusion: Colistin resistance mechanisms depend on genetic background, and are due to chromosomal mutations, which implies a lower risk of transmission than plasmid-mediated genes. Colistin resistance is not associated with multidrug-resistance, nor to high-risk sequence types.
Keywords: sequence type, LPS-modifying enzymes, polymorphism vs potential mutation, MgrB regulator, PmrA-PmrB and PhoP-PhoQ three-component systems
Introduction
The increasing prevalence of multidrug-resistant Gram-negative bacteria, together with the lack of new antibiotics has led to the reintroduction of disused drugs such as colistin.1 For many years this antibiotic was used only as a topical treatment in human medicine, because of its nephrotoxicity. In contrast, it was massively used in veterinary medicine for the treatment and prevention of infectious diseases caused by Gram-negative bacteria.2 Despite its relatively recent reintroduction in clinical practice for systemic treatment, reports of colistin-resistant isolates are on the rise worldwide.3 The first reports about colistin resistance were about mutations in chromosomal genes involved in lipopolysaccharide (LPS) synthesis and modification through addition of cationic groups to lipid A that increase the positive charge of LPS leading to reduced interaction of the outer membrane with positively charged colistin.4 These LPS modifications are responsible for the acquisition of resistance to this antibiotic in Enterobacteriaceae.4 The first plasmid-mediated colistin resistance gene, mcr-1, was described in Escherichia coli in 2015;5 Mcr-1 catalyzes the modification of lipid A and captured attention because of its dissemination risk.6
Chromosomal genes associated with colistin resistance are of three classes. First, genes coding for enzymes that modify LPS through addition of positively charged groups: pmrC that adds phosphoethanolamine (PEtN) to LPS,7 pmrE and pmrHFIJKLM operon that catalyze the synthesis and union of 4-amino-4-deoxy-L-arabinose (L-Ara4N) to LPS.8 Addition of L-Ara4N confers increased levels of positive charge than addition of PEtN.9 Second, regulatory genes encoding PhoPQ and PmrAB, two-component systems (TCSs) that in normal conditions detect environmental stimuli and modify the LPS charge accordingly. PmrB is the sensor kinase that activates PmrA, and PmrA activates transcription of pmrC, pmrE and pmrHFIJKLM operon.10 Mutations in the pmrCAB operon have been described as the most frequent mechanism responsible for acquired resistance to colistin in E. coli.11 PhoQ is the sensor kinase that activates PhoP, and PhoP activates pmrHFIJKLM operon, pmrA and pmrD connector regulatory protein. Mutations in phoP and phoQ genes are also responsible for the acquired resistance to colistin in K. pneumoniae and E. coli.12 Mutated TCSs no longer respond to environmental stimuli and downstream activators become constitutively overexpressed. Third, genes encoding regulators of these TCSs, namely mgrB and crrAB.13 MgrB inhibits the phosphatase activity of PhoQ which consequently decreases PhoP phosphorylation. Alterations in mgrB gene lead to derepressed activity of the PhoPQ TCS and are responsible for colistin resistance in Klebsiella species.4 Nonsense mutations leading to a truncated MgrB protein, amino-acid substitutions, insertions, deletions, or even complete deletions of the mgrB locus, are the most frequently reported colistin resistance mechanism in Klebsiella,14 but not in E. coli. Other chromosomal genes associated with colistin resistance in K. pneumoniae are kpnE and kpnF,15 acrA and acrB,16 that code for efflux pump systems.
The aim of this study was to perform a retrospective genomic analysis of colistin-resistant clinical isolates of K. pneumoniae and E. coli obtained in a university hospital in Santiago, Chile. None of the isolates harbored mcr genes, therefore the analysis was focused on chromosomal gene mutations and association with their genetic background.
Methods
Strains
Fifteen colistin-resistant Enterobacteriaceae isolates were analyzed: 8 Klebsiella pneumoniae (named KPN-1, KPN-3, KPN-5, KPN-6, KPN-7, KPN-10, KPN-11, KPN-20) from blood cultures, urine, peritoneal fluid, and oral mucosa, and 7 Escherichia coli (EC-4, EC-10, EC-11, EC-13, EC-15, EC-16, EC-19) all from urine (Table 1). They were collected from different outpatients at the Catholic University Hospital in Santiago, Chile between 2015 and 2018. Urine cultures were cultured in 5% sheep blood agar and chromogenic CPS agar. Blood specimens were inoculated in BacT/ALERT blood culture bottles (BioMerieux, France) and then subcultured on MacConkey and 5% sheep blood agar. Oral mucosa swabs and peritoneal fluid were cultured in 5% sheep blood agar, chocolate agar and McConkey agar plates. All agar plates were Biomerieux (Lètolile, France). Suspicious bacterial colonies were identified through Matrix Assisted Laser Desorption – Time of Flight Mass Spectrometry (MALDI–TOF) (Bruker-Daltonics, Bremen, Germany). All the isolates were negative for carbapenemase production as determined through CarbaNP test performed according to the guidelines of Clinical and Laboratory Standards Institute 2022.17 Two colistin-susceptible controls were used, Escherichia coli ATCC 25922 and Klebsiella pneumoniae ATCC BAA-1706. Colistin-susceptible isolates were obtained from the university hospital (BR1-E11, BR1-F11, BR1-C3, BR1-B1) and from the MICROB-R Network of 11 hospitals along Chile (SCL7471, SCL2906, SCL2922). Colistin-susceptible isolates of ST454, ST380 and ST127 types were obtained from PATRIC database and colistin susceptibility was predicted using Kleborate platform (for K. pneumoniae)18 and Resfinder platform (for E. coli).19 Approval from the Ethics Committee of Pontificia Universidad Católica de Chile for the use of strains isolated from human samples was obtained.
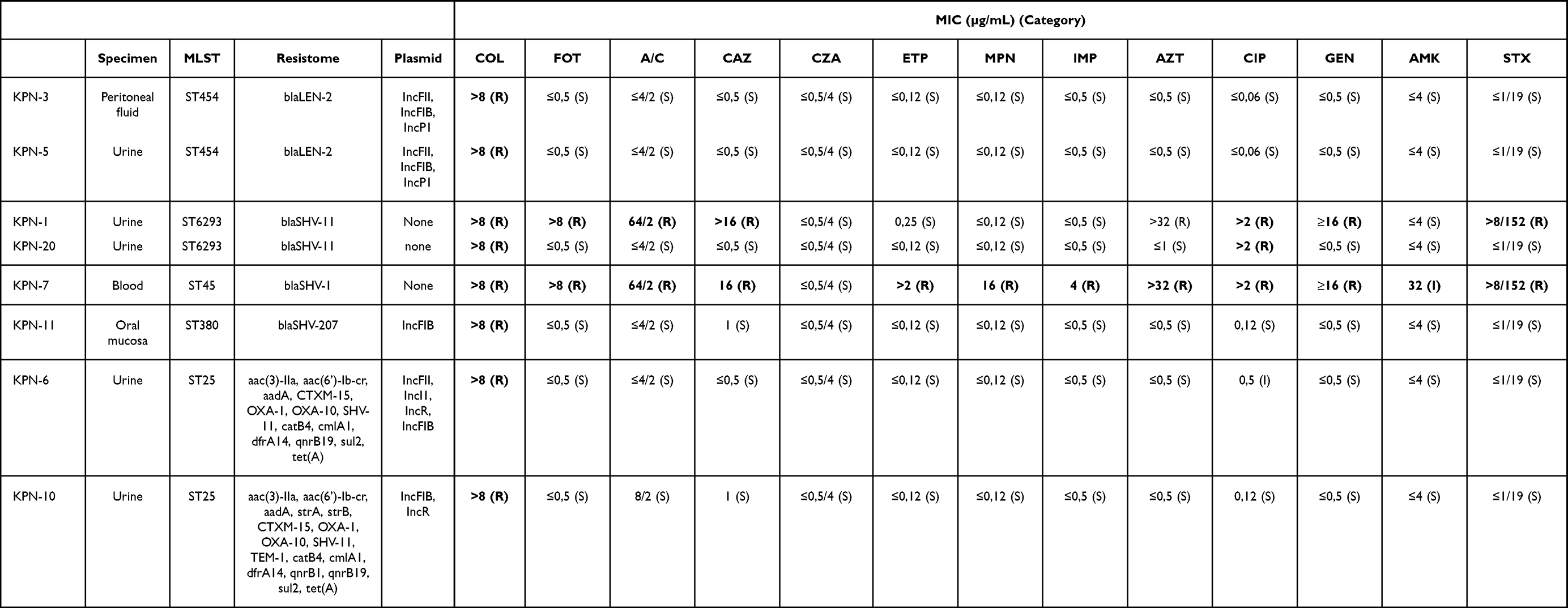 |
Table 1 Antimicrobial Susceptibility, Acquired Resistome and Plasmids Found in K. pneumoniae Isolates |
Antimicrobial Susceptibility Testing
Minimum inhibitory concentration (MIC) of colistin was determined through broth microdilution method using Sensititre Gram-Negative DKMGN Plates according to manufacturer’s instructions (ThermoFisher Scientific). Overnight-grown colonies in Mueller Hinton agar plates were resuspended in PBS to obtain a McFarland turbidity standard between 0.50 and 0.55. Plates were inoculated with 10 µL of adjusted bacterial suspension and incubated for 20 h before reading. Isolates with MIC ≥4 µg/mL were considered resistant and those with MIC ≤2 µg/mL were classified as intermediately resistant, according to CLSI 2022 guidelines. Susceptibility to amikacin, gentamicin, ertapenem, meropenem, imipenem, aztreonam, amoxicillin/clavulanate, cefotaxime, ceftazidime, ceftazidime/avibactam, trimethoprim/sulfamethoxazole and ciprofloxacin was also determined through the above-mentioned broth microdilution method using the breakpoints suggested by the Clinical Laboratory Standards Institute 2022 guidelines.17
Whole Genome Sequencing (WGS) Through Illumina Short Reads Method
Isolates were sequenced using short-read WGS (Illumina). A 350 bp insert DNA library was prepared using Illumina DNA Prep Kit (formerly Nextera Flex), following the Hackflex protocol.20 Sequencing was performed in an Illumina Platform PE150, at the University of Technology Sydney’s Bioscience Laboratory (Sydney, Australia). The Q30 obtained was >90% for all isolates. De novo assembly was performed using SPADES version 3.7 package.21 Genomic annotation of the recovered draft genomes was performed with Prokka tool 1.11.22 MLST analysis, plasmid types and resistome were determined using Kleborate platform18 and Resfinder platform for K. pneumoniae and E. coli respectively. Final visualization was made using Galaxy-Australia platform (https://usegalaxy.org.au/).
Genetic Analysis of Chromosomal Genes Associated with Colistin Resistance
Sequences were compared with wild-type (WT) gene sequences using Sequencher, BLAST, and ClustalW software. The protein functionality was analyzed with SIFT algorithm (Sorting Intolerant From Tolerant) allowing a prediction of functional impact of mutations as “deleterious” (score ≤ 0.05) or “neutral” (score >0.05).23 Genome sequences described in this paper have been deposited in GenBank database under Bioproject N° PRJNA991619.
Results
Susceptibility of Isolates to Colistin and Other Antimicrobials
Clinical isolates of colistin-resistant K. pneumoniae (8 isolates) and E. coli (7 isolates) were obtained in a 3-year period in the institutional university hospital. All K. pneumoniae isolates had a MIC of colistin >8 µg/mL, whereas E. coli isolates had a MIC of 4 (1 isolate), 8 (4 isolates) and >8 µg/mL (2 isolates) (Table 1 and 2).
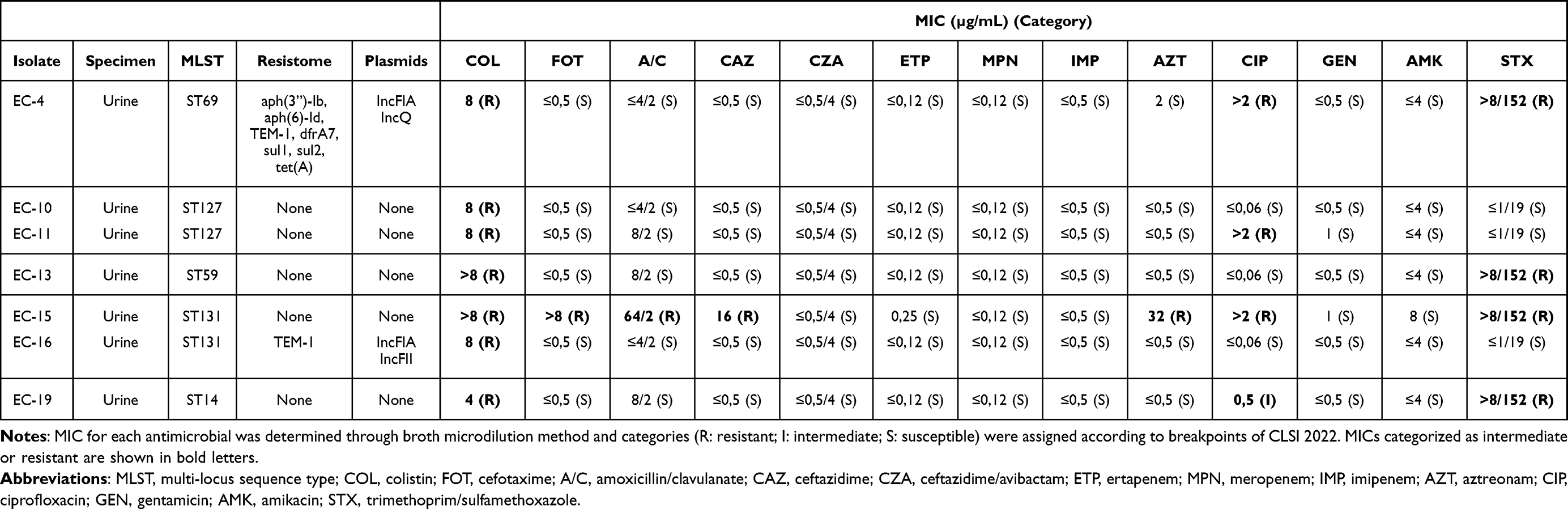 |
Table 2 Antimicrobial Susceptibility, Acquired Resistome and Plasmids Found in E. coli Isolates |
Among K. pneumoniae isolates, 6/8 were susceptible to all antimicrobials tested (except for isolate KPN-20 that was resistant to ciprofloxacin) and were classified as non-MDR according to Magiorakos criteria (MDR is defined as non-susceptibility to at least one agent in three or more antimicrobial categories).24 Two isolates were MDR: isolate KPN-7 was resistant to all antimicrobials tested except for ceftazidime/avibactam and intermediately resistant to amikacin, and isolate KPN-1 was resistant to cefotaxime, ceftazidime, amoxicillin/clavulanate, ciprofloxacin, gentamicin, and trimethoprim/sulfamethoxazole (Table 1). E. coli isolates were all susceptible to amikacin, gentamicin, meropenem, imipenem, ertapenem and ceftazidime/avibactam. Among them, 4/7 isolates were non-MDR and 3/7 were MDR. Isolate EC-15 was resistant to cefotaxime, amoxicillin/clavulanate, ceftazidime, aztreonam, ciprofloxacin and trimethoprim/sulfamethoxazole, and isolates EC-4 and EC-19 were resistant to ciprofloxacin and trimethoprim/sulfamethoxazole (Table 2). Overall, 10/15 colistin-resistant isolates were non-MDR.
Genomic Analysis of Colistin-Resistant Isolates: Sequence Types and Phylogenetic Analysis
Among K. pneumoniae isolates 2 of them were K. pneumoniae subsp. variicola and 6 were K. pneumoniae subsp. pneumoniae according to Kleborate analysis.18 The STs found were ST25 (2 isolates), ST45 (1 isolate), ST454 (2 K. pneumoniae subsp. variicola isolates), ST380 (1 isolate) and two isolates were assigned a new ST named ST6293. This new ST is evolutionarily related to ST12. A phylogenetic tree was constructed for each species based on the core genome. Publicly available genomes from PATRIC database of the same STs were included in the analysis. Chilean K. pneumoniae isolates clustered together with isolates of the same STs from different countries (Figure 1A). It can be observed that two clusters were formed, one with K. pneumoniae subsp. variicola isolates and the other one with K. pneumoniae subsp. pneumoniae isolates (Figure 1A). Isolates belonging to the novel ST6293 formed a cluster close to ST12 isolates. Genomes of ST12 type were included because of their relatedness to ST6293. STs found among E. coli isolates were ST131 (2 isolates), ST69 (1 isolate), ST59 (1 isolate), ST14 (1 isolate) and ST127 (2 isolates) according to genomic analysis made using Resfinder in the Galaxy-Australia platform.19 The phylogenetic tree constructed based on the core genome showed that Chilean E. coli isolates clustered together with isolates of the same STs from different countries (Figure 1B).
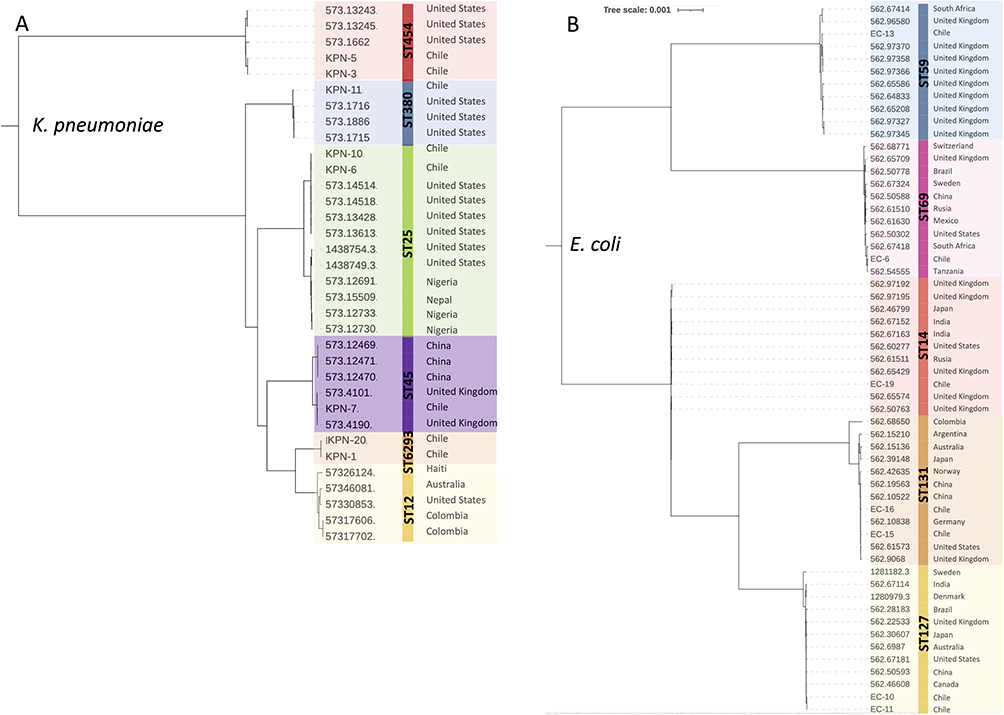 |
Figure 1 Phylogenetic tree based on the core genome of K. pneumoniae (A) and E. coli (B) isolates. Genomes of isolates of the same STs found in our work were randomly selected from PATRIC database and included in the phylogenetic tree. The genome ID and country of isolation are shown for each isolate. Multiple sequence alignment was performed using Clustal W and resultant output was generated with MegaX. |
Genomic Analysis: Resistome and Plasmids
mcr genes were not found in any of the isolates studied here. Analysis of the acquired resistome showed that K. pneumoniae subsp. variicola isolates had only the blaLEN-1 gene coding for the typical endogenous beta-lactamase whereas K. pneumoniae subsp. pneumoniae isolates had blaSHV genes (Table 1). ST25 isolates had additional antimicrobial resistance genes: aac(3)-IIa, aac(6’)-Ib-cr, aadA, CTXM-15, OXA-1, OXA-10, SHV-11, catB4, cmlA1, dfrA14, qnrB19, sul2, tet(A). All the K. pneumoniae isolates had plasmids belonging to incompatibility groups IncFII, IncFIB, IncP1 and IncR. In contrast, ST6293 and ST45 isolates had no plasmids according to Kleborate analysis.
Five of the 7 E. coli isolates had no resistance genes and no plasmids. Isolate EC-16 had only blaTEM-1 gene and plasmids IncFIA and IncQ1. Isolate EC-4 had aph(3”)-Ib, aph(6)-Id, blaTEM-1B, dfrA7, sul1, sul2 and tet(A) and plasmids IncFIA and IncFII (Table 2). There was no evident correlation between resistome and antimicrobial susceptibility, e.g. isolates EC-15 and KPN-7 were MDR but had no resistance genes and only blaSHV-1 respectively, conversely isolate KPN-6 had several resistance genes including extended-spectrum beta-lactamases but was susceptible to all antimicrobials tested except colistin.
Mutations in Chromosomal Genes Involved in LPS Modification
All the isolates had mutations in chromosomal genes involved in LPS modification with respect to the WT reference strains (K. pneumoniae ATCC BAA-1706 and E. coli ATCC 25922) (Table 3 and 4). To distinguish between gene polymorphisms associated with a particular ST and potential mutations, colistin-susceptible isolates of the same STs were included in the analysis. It was observed that most of the mutations were present in susceptible isolates of the same ST and different ST as well, meaning that they are most likely gene polymorphisms. Those mutations that were not present in susceptible isolates nor in the reference ATCC strains, are shown in bold underlined letters in Table 3 and 4. K. pneumoniae and E. coli isolates have a different mutation profile in LPS-modifying enzymes: most of K. pneumoniae isolates had mutations in mgrB, whereas E. coli had mutations in pmrA, pmrB and pmrE genes. Among K. pneumoniae isolates, 5/8 had alterations in the mgrB gene whereas 3/8 had mutations in pmr genes. Isolates KPN-3, KPN-6 and KPN-11 had a premature stop codon in the mgrB gene that produced a truncated MgrB protein of 2, 22, and 28 amino acids respectively. The mgrB gene was not found in isolate KPN-10, and isolate KPN-5 had a point mutation R33Q predicted as deleterious through SIFT algorithm (Table 3). Among K. pneumoniae subsp. pneumoniae isolates 11 amino acid changes were found with respect to the reference strain ATCC BAA-1706, but 8 of them were present in susceptible isolates of the same and/or different STs and are considered gene polymorphisms. Only three mutations were not found in susceptible isolates: Q331P in pmrE gene, D21A in pmrB gene and L716R in acrB gene. Isolates KPN-1 and KPN-20 (of the novel ST6293) had the mutation Q331P in their pmrE gene that was predicted as neutral through SIFT algorithm. Colistin-susceptible isolate of ST12 type had 21 amino acid changes with respect to ATCC BAA-1706 reference strain, and 20 of them were not present in isolates KPN-1 and KPN-20 and are most likely polymorphisms related to this novel ST6293. Isolate KPN-7 had D21A mutation in pmrB gene that was predicted as neutral though SIFT analysis, but also had a deletion of kpnE and pmrD genes. Isolate KPN-6 having a truncated MgrB protein, had also mutation L716R in its acrB gene, predicted as neutral through SIFT analysis. Isolates KPN-3, KPN-5 (K. pneumoniae subsp. variicola), KPN-7 and KPN-11 did not have crrA nor crrB genes. The colistin-susceptible isolates of the same STs did not have these genes either, therefore we consider the absence of these genes is not involved in colistin resistance.
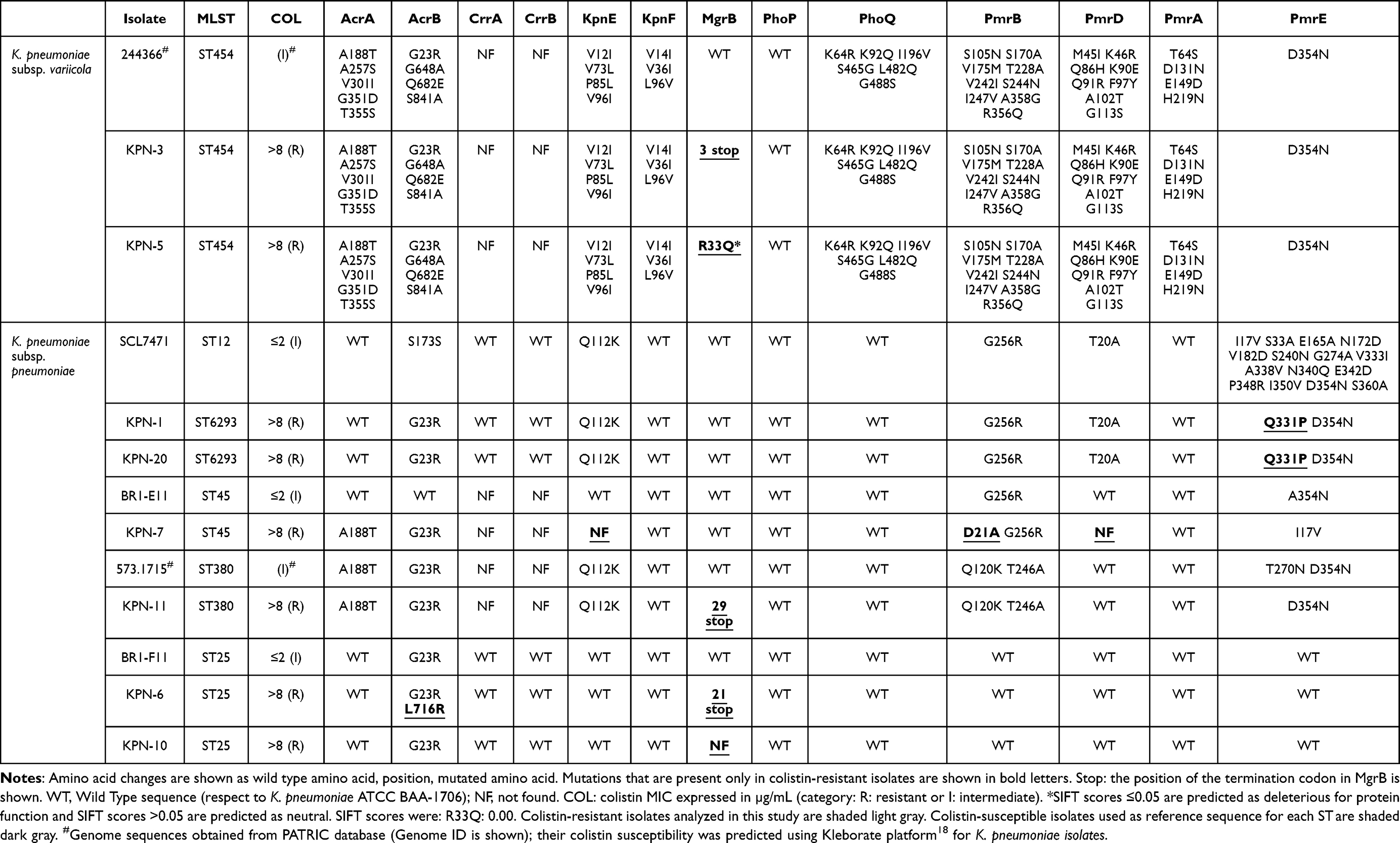 |
Table 3 Amino Acid Changes in Proteins Associated With Colistin Resistance With Respect to K. pneumoniae ATCC BAA-1706 |
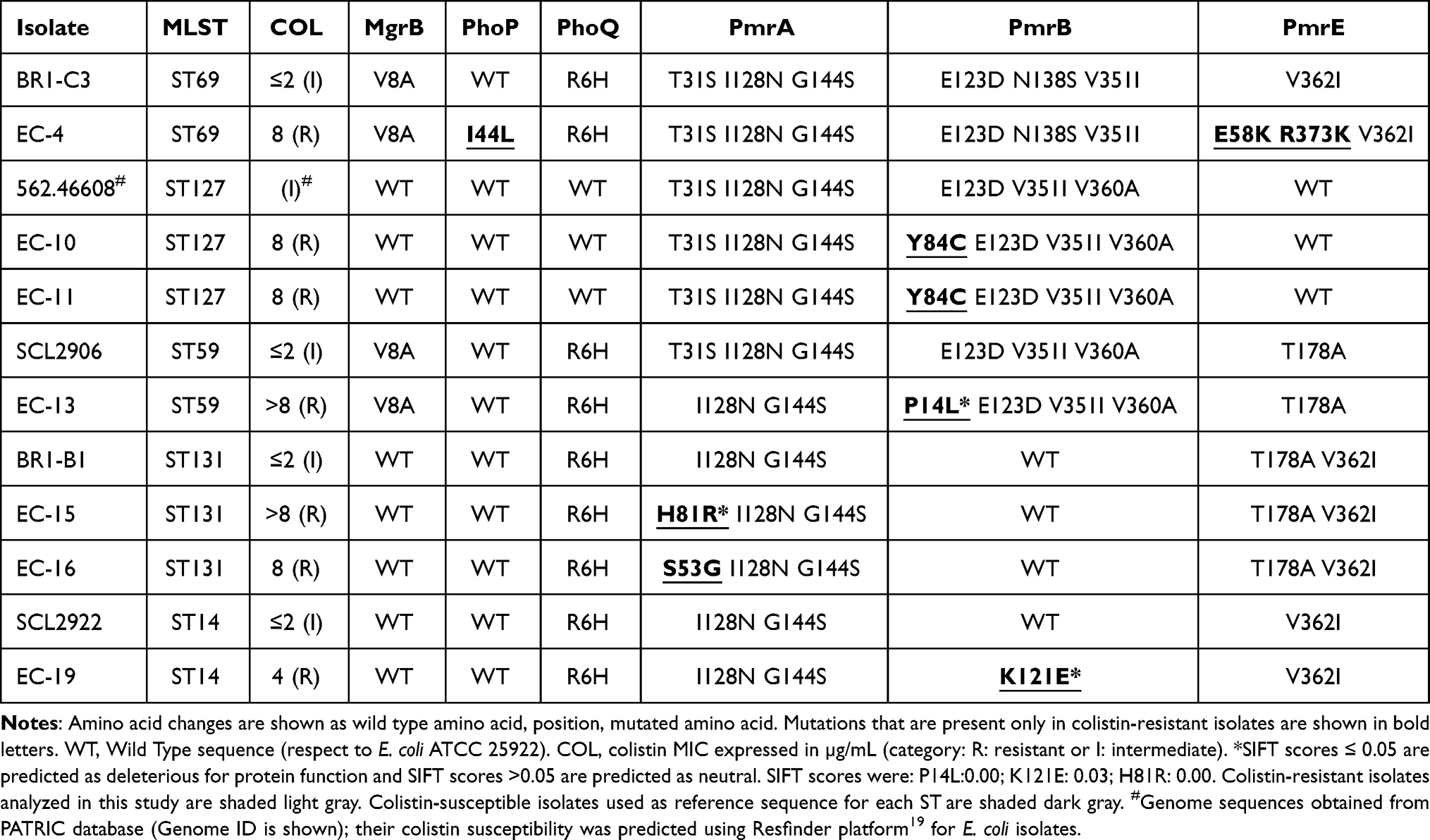 |
Table 4 Amino Acid Changes in Proteins Associated With Colistin-Resistance With Respect to E. coli ATCC 25922 |
K. pneumoniae subsp. variicola isolates KPN-3 and KPN-5 had 45 mutations with respect to the reference strain ATCC BAA-1706 (Table 3), but 44 of them were also present in the colistin-susceptible isolate of K. pneumoniae subsp. variicola of the same ST, meaning that they are most likely polymorphisms associated with this subspecies; only R33Q mutation was not found in susceptible isolates.
Among E. coli isolates, all of them (7/7) had a WT mgrB gene. All the E. coli isolates had mutations in pmrA, pmrB or pmrE genes. Overall, 8 mutations were found in LPS modifying enzymes of E. coli isolates and are shown in bold letters in Table 4. Isolates EC-15 and EC-16 each had a point mutation in pmrA gene, R81H and G53S respectively. Isolates EC-10 and EC-11 had a mutation in pmrB, Y84C. Isolates EC-13 and EC-19 had a point mutation in the pmrB gene, P14L and K121E respectively. Isolate EC-4 had two mutations in pmrE, K58E and K373R, and L44I in phoP gene. Only P14L, R81H and K121E were predicted as deleterious through SIFT analysis, the rest of them were predicted as neutral (Table 3 and 4).
Discussion
The present work corresponds to the first report of chromosomal mutations in colistin-resistant isolates of K. pneumoniae and E. coli from Chile. Several mutations were found in this work. However, most of them correspond to gene polymorphisms, and only a few of them can account for colistin resistance. MgrB is a crucial regulator and their mutations have been described as sufficient to increase the colistin MIC in K. pneumoniae.14 Hence, truncated or absent MgrB protein in 5/8 K. pneumoniae isolates most likely accounts for their colistin resistance; mutation R33Q in MgrB of isolate KPN-5 has been previously described in a colistin-resistant K. pneumoniae isolate.25,26 Isolates with WT mgrB gene have mutations or loss of pmr genes that may account for colistin resistance: Q331P in pmrE gene of KPN-1 and KPN-20, D21A in pmrB gene together with loss of pmrD gene in KPN-7. To our knowledge, these mutations have not been reported to date in colistin-resistant clinical isolates. It must be noted that mutation D21A in pmrB gene of KPN-5 is accompanied by loss of kpnE and pmrD genes. KpnE together with KpnF form an efflux-pump associated with resistance to several antibiotics including colistin: mutant K. pneumoniae lacking kpnEF genes reduced its MIC to colistin two-fold.15 PmrD is a transcriptional activator of the PmrAB TCS, and this activation is required for LPS modification. Although LPS modification can also be activated independently of PmrD,4 further experiments should be made to understand how the lack of these genes, which a priori contribute to LPS modification, affects colistin resistance. Isolate KPN-6 besides a truncated MgrB protein has the amino acid change L716R in acrB gene, that codes for an efflux-pump. Mutation Q331P in PmrE was the only mutation in isolates KPN-1 and KPN-20 that was not present in susceptible isolates. PmrE is the first enzyme in L-Ara4N synthesis and is required for LPS modification. Q331P is located in the binding site of NAD+ of PmrE enzyme and it could increase efficiency of PmrE enzyme.10 In K. pneumoniae genes crrA and crrB code for a regulator protein and a sensor kinase respectively.4 Although its role is not completely understood, inactivation of CrrB leads to overexpression of PmrAB operon and addition of positive charges to LPS.27 These genes were not found in K. pneumoniae subsp. variicola isolates, nor in the colistin-susceptible isolate. The same was observed in isolates of ST45 and ST380 types. It is known that all isolates of the ST258 type have crrAB genes, whereas other STs do not have these genes.27 It is possible that other, yet unknown genes, are involved in colistin resistance in isolates that lack crrAB genes.
The two-component system PmrAB is commonly the most affected in colistin-resistant E. coli isolates.4,9 Mutations H81R and S53G in PmrA were not reported to date. However, similar mutations have been described in colistin-resistant Salmonella, C81R and E53G,28 and in K. pneumoniae, G81R,29 and S81R.30 Mutations H81R and S53G are located close to D51 in the active site of phosphate receiver domain and mutations in these positions were shown to increase colistin MIC.28 Amino acid changes P14L and K121E in PmrB were not reported to date, however, similar mutations, S14L, F14L and A121E were described in colistin-resistant Salmonella isolates and were genetically confirmed to be sufficient to confer colistin resistance.28 These mutations are in trans-membrane-1 and linker domains of PmrB, and mutation analysis with PmrB homologs in Salmonella showed that amino acid changes in these domains increase kinase activity of PmrB.28
Most of the amino acid changes found in this work are considered polymorphisms owing to their presence in colistin-susceptible isolates. Some of these polymorphisms, such as G256R were already reported in colistin-susceptible isolates,31 and in colistin-resistant isolates.32 Moreover, in a recent report, 6 of the 21 polymorphisms that we found here in PmrE were reported to be associated with colistin resistance.33 These data highlight the importance of performing point mutation analysis of resistant isolates together with colistin-susceptible isolates of the same ST, to distinguish between gene polymorphisms and potential mutations.
The association of ST with diverse colistin resistance mechanisms in K. pneumoniae was previously reported.26,27 Azam and coworkers reported mutations in MDR and PDR isolates obtained from hospitalized patients that received colistin antibiotic therapy,26 in contrast to isolates reported in this work that are mostly non-MDR and were obtained from outpatients. None of the mutations found by Azam and coworkers were found in our work and vice versa, and isolates belonged to STs other than those found here. A similar finding was observed with STs and mutations reported by Wright and coworkers.27 The results found here further support the association of ST with a colistin-resistance mechanism. To understand and clarify this association a higher number of isolates should be analyzed, and site-directed mutagenesis should be performed to confirm the contribution of each novel mutation to the resistant phenotype.
Colistin resistance is rising worldwide3 and in our setting too (unpublished data of our laboratory). Nevertheless, we can be optimistic about three aspects of the results found here. First, colistin resistance is associated with chromosomal mutations rather than plasmid-mediated mcr genes, lowering the risk of widespread dissemination through horizontal gene transfer. Second, colistin resistance is not associated with high-risk clones of E. coli and K. pneumoniae. Successful E. coli clones associated with community and hospital-acquired infections are ST131, ST410, ST38, ST73, ST405 and ST648.34 Among E. coli isolates studied here 3/7 belong to high-risk clones (ST131 and ST69), and none of the K. pneumoniae isolates belong to the globally distributed high-risk clones, namely ST258, ST11, ST512, ST14 and ST15 among others.35 Third, colistin resistance is not associated with multidrug-resistance. Most of the isolates reported here (10/15) are non-MDR and are susceptible to all the antibiotics tested, except for colistin. In contrast to the frequently reported carbapenem+colistin resistant clinical isolates for which few or no treatment options exist,26,33,36 antibiotic therapies are still available for isolates like the ones reported here.
Conclusion
We report here several mutations in genes coding for LPS-modifying enzymes, mgrB, pmrAB, phoPQ, and in other genes such as pmrE, pmrD, acrB and kpnE, whose actual contribution to colistin resistance requires analysis with isogenic mutants. Genomic analysis and results previously reported support the idea that colistin resistance mechanisms depend on genetic background. Colistin resistance in our setting is due to chromosomal mutations, which implies a lower risk of transmission than plasmid-mediated genes. Additionally, colistin resistance is not associated with multidrug-resistance since most of the isolates were susceptible to all antibiotics tested, and nor is it associated with high-risk STs.
Acknowledgments
The authors acknowledge the expert technical assistance of the staff of Laboratory of Microbiology from Pontificia Universidad Católica de Chile for their help in technical aspects of this work.
Funding
This work was supported by research funds from SENTRY (Antimicrobial Resistance Surveillance Program), the Red de Salud UC-Christus and the Department of Clinical Laboratories at the School of Medicine of Pontificia Universidad Católica de Chile.
Disclosure
The authors declare no conflicts of interest in this work.
References
1. Cassir N, Rolain J-M, Brouqui P. A new strategy to fight antimicrobial resistance: the revival of old antibiotics.Front Microbiol. 2014;5:551. doi:10.3389/fmicb.2014.00551
2. Catry B, Cavaleri M, Baptiste K, et al. Colistin resistance mechanisms in Klebsiella pneumoniae strains from Taiwan. Antimicrob Agents Chemother. 2015;59(5):2909–2913. doi:10.1128/AAC.04763-14
3. Binsker U, Käsbohrer A, Hammerl J. A Global colistin use: a review of the emergence of resistant Enterobacterales and the impact on their genetic basis, FEMS Microbiology. Reviews. 2022;46(1):fuab049. doi:10.1093/femsre/fuab049
4. Poirel L, Jayol A, Nordmann P. Polymyxins: antibacterial Activity, Susceptibility Testing, and Resistance Mechanisms Encoded by Plasmids or Chromosomes. Clin Microbiol Rev. 2017;30(2):557–596. doi:10.1128/cmr.00064-16
5. Liu YY, Wang Y, Walsh E, et al. Emergence of plasmid-mediated colistin resistance mechanism MCR-1 in animals and human beings in China: a microbiological and molecular biological study. Lancet Infect Dis. 2016;16(2):161–168. doi:10.1016/S1473-3099(15)00424-7
6. Hinchliffe P, Yang QE, Portal E, et al. Insights into the Mechanistic Basis of Plasmid-Mediated Colistin Resistance from Crystal Structures of the Catalytic Domain of MCR-1. Sci Rep. 2017;7(1):39392. doi:10.1038/srep39392
7. Jaidane N, Bonnin RA, Mansour W, et al. Genomic Insights into Colistin-Resistant Klebsiella pneumoniae from a Tunisian Teaching Hospital. Antimicrob Agents Chemother. 2018;62(2):e01601–17. doi:10.1128/AAC.01601-17
8. Yan A, Guan Z, Raetz CR. An undecaprenyl phosphate- aminoarabinose flippase required for polymyxin resistance in Escherichia coli. J Biol Chem. 2007;282(49):36077–36089. doi:10.1074/jbc.M706172200
9. Olaitan AO, Morand S, Rolain JM. Mechanisms of polymyxin resistance: acquired and intrinsic resistance in bacteria. Front Microbiol. 2014;5:643. doi:10.3389/fmicb.2014.00643
10. Gunn JS. The Salmonella PmrAB regulon: lipopolysaccharide modifications, antimicrobial peptide resistance and more. Trends Microbiol. 2008;16(6):284–290. doi:10.1016/j.tim.2008.03.007
11. Janssen AB, van Schaik W. Harder, better, faster, stronger: colistin resistance mechanisms in Escherichia coli. PLoS Genet. 2021;17(1):e1009262. doi:10.1371/journal.pgen.1009262
12. Jayol A, Nordmann P, Brink A, et al. Heteroresistance to colistin in Klebsiella pneumoniae associated with alterations in the PhoPQ regulatory system. Antimicrob Agents Chemother. 2015;59(5):2780–2784. doi:10.1128/AAC.05055-14
13. McConville TH, Annavajhala MK, Giddins G, et al. CrrB Positively Regulates High-Level Polymyxin Resistance and Virulence in Klebsiella pneumoniae. Cell Rep. 2020;33(4):108313. doi:10.1016/j.celrep.2020.108313
14. Cannatelli A, Giani T, D’Andrea M, et al. MgrB inactivation is a common mechanism of colistin resistance in KPC-producing Klebsiella pneumoniae of clinical origin. Antimicrob Agents Chemother. 2014;58(10):5696–5703. doi:10.1128/AAC.03110-14
15. Srinivasan VB, Rajamohan G. KpnEF, a new member of the Klebsiella pneumoniae cell envelope stress response regulon, is an SMR-type efflux pump involved in broad-spectrum antimicrobial resistance. Antimicrob Agents Chemother. 2013;57(9):4449–4462. doi:10.1128/AAC.02284-12
16. Razavi S, Mirnejad R, Babapour E. Involvement of AcrAB and OqxAB Efflux Pumps in Antimicrobial Resistance of Clinical Isolates of Klebsiella pneumonia. J Appl Biotechnol Rep. 2020;7(4):251–257. doi:10.30491/jabr.2020.120179
17. CLSI. Performance Standards for Antimicrobial Susceptibility Testing.
18. Lam MMC, Wick RR, Watts SC, et al. A genomic surveillance framework and genotyping tool for Klebsiella pneumoniae and its related species complex. Nat Commun. 2021;12(1):4188. doi:10.1038/s41467-021-24448-3
19. Florensa AF, Kaas RS, Clausen PTLC, et al. ResFinder - an open online resource for identification of antimicrobial resistance genes in next-generation sequencing data and prediction of phenotypes from genotypes. Microb Genom. 2022;8(1):000748. doi:10.1099/mgen.0.000748
20. Gaio D, Anantanawat K, To J, et al. Hackflex: low-cost, high-throughput, Illumina Nextera Flex library construction. Microb Genom. 2022;8(1):000744. doi:10.1099/mgen.0.000744
21. Bankevich A, Nurk S, Antipov D, et al. SPAdes: a New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J Comput Biol. 2012;19(5):455–477. doi:10.1089/cmb.2012.0021
22. Seemann T. Prokka: rapid prokaryotic genome annotation. Bioinformatics. 2014;30(14):2068–2069. doi:10.1093/bioinformatics/btu153
23. Vaser R, Adusumalli S, Leng SN, et al. SIFT missense predictions for genomes. Nat Protocols. 2016;11(1):1–9. doi:10.1038/nprot.2015.123
24. Magiorakos AP, Srinivasan A, Carey R, et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: an international expert proposal for interim standard definitions for acquired resistance. Clin Microbiol Infect. 2012;18(3):268–281. doi:10.1111/j.1469-0691.2011.03570.x
25. Delannoy S, Le Devendec L, Jouy E, et al. Characterization of Colistin-Resistant Escherichia coli Isolated from Diseased Pigs in France. Front Microbiol. 2017;8:2278. doi:10.3389/fmicb.2017.02278
26. Azam M, Gaind R, Yadav G, et al. Colistin Resistance Among Multiple Sequence Types of Klebsiella pneumoniae Is Associated with Diverse Resistance Mechanisms: a Report From India. Front Microbiol. 2021;12:609840. doi:10.3389/fmicb.2021.609840
27. Wright MS, Suzuki Y, Jones MB, et al. Genomic and transcriptomic analyses of colistin-resistant clinical isolates of Klebsiella pneumoniae reveal multiple pathways of resistance. Antimicrob Agents Chemother. 2015;59(1):536–543. doi:10.1128/AAC.04037-14
28. Sun S, Negrea A, Rhen M, et al. Genetic Analysis of Colistin Resistance in Salmonella enterica Serovar Typhimurium. Antimicrob Agents Chemother. 2009;53(6):2298–2305. doi:10.1128/AAC.01016-08
29. Mills J, Rojas L, Marshall SH, et al. Risk Factors for and Mechanisms of Colistin Resistance Among Enterobacterales: getting at the CORE of the Issue. Open Forum Infect Dis. 2021;8(7):ofab145. doi:10.1093/ofid/ofab145
30. Quesada A, Porrero MC, Tellez S, et al. Polymorphism of genes encoding PmrAB in colistin-resistant strains of Escherichia coli and Salmonella enterica isolated from poultry and swine. J Antimicrob Chemother. 2015;70(1):71–74. doi:10.1093/jac/dku320
31. Cheng YH, Lin TL, Pan YJ, et al. Colistin resistance mechanisms in Klebsiella pneumoniae strains from Taiwan. Antimicrob Agents Chemother. 2015;59(5):2909–2913. doi:10.1128/AAC.04763-14
32. Rodrigues ACS, Santos ICO, Campos CC, et al. Non-clonal occurrence of pmrB mutations associated with polymyxin resistance in carbapenem-resistant Klebsiella pneumoniae in Brazil. Mem Inst Oswaldo Cruz. 2019;114:e180555. doi:10.1590/0074-02760180555
33. Hee Lee T, Cho M, Lee J, et al. Molecular Characterization of Carbapenem-resistant, Colistin-resistant Klebsiella pneumoniae Isolates from a Tertiary Hospital in Jeonbuk, Korea. J Bacteriol Virol. 2021;51(3):120–127. doi:10.4167/jbv.2021.51.3.120
34. Mazumder R, Hussain A, Abdullah A, et al. International High-Risk Clones Among Extended-Spectrum β-Lactamase-Producing Escherichia coli in Dhaka, Bangladesh. Front Microbiol. 2021;4(12):736464. doi:10.3389/fmicb.2021.736464
35. Wyres KL, Lam MC, Holt KE. Population genomics of Klebsiella pneumoniae. Nat Rev Microbiol. 2020;18(6):344–359. doi:10.1038/s41579-019-0315-1
36. Di Tella D, Tamburro M, Guerrizio G, et al. Molecular Epidemiological Insights into Colistin-Resistant and Carbapenemases Producing Clinical Klebsiella pneumoniae Isolates. Infect Drug Resist. 2019;12:3783–3795. doi:10.2147/IDR.S226416
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.