Back to Journals » Drug Design, Development and Therapy » Volume 16
Chemical Chaperones to Inhibit Endoplasmic Reticulum Stress: Implications in Diseases
Authors Jeon JH, Im S, Kim HS, Lee D, Jeong K ![]() , Ku JM, Nam TG
, Ku JM, Nam TG ![]()
Received 26 October 2022
Accepted for publication 10 December 2022
Published 23 December 2022 Volume 2022:16 Pages 4385—4397
DOI https://doi.org/10.2147/DDDT.S393816
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Tuo Deng
Jae-Ho Jeon,1,* Somyoung Im,1,* Hyo Shin Kim,1 Dongyun Lee,1 Kwiwan Jeong,2 Jin-Mo Ku,2 Tae-Gyu Nam1
1Department of Pharmacy and Institute of Pharmaceutical Science and Technology, Hanyang University ERICA campus, Ansan, Gyeonggi-do, 15588, Republic of Korea; 2Gyeonggi Bio-Center, Gyeonggido Business and Science Accelerator, Suwon, Gyeonggi-do, 16229, Republic of Korea
*These authors contributed equally to this work
Correspondence: Tae-Gyu Nam, Tel +82-31-400-5807, Fax +82-31-400-5958, Email [email protected]
Abstract: The endoplasmic reticulum (ER) is responsible for structural transformation or folding of de novo proteins for transport to the Golgi. When the folding capacity of the ER is exceeded or excessive accumulation of misfolded proteins occurs, the ER enters a stressed condition (ER stress) and unfolded protein responses (UPR) are triggered in order to rescue cells from the stress. Recovery of ER proceeds toward either survival or cell apoptosis. ER stress is implicated in many pathologies, such as diabetes, cardiovascular diseases, inflammatory diseases, neurodegeneration, and lysosomal storage diseases. As a survival or adaptation mechanism, chaperone molecules are upregulated to manage ER stress. Chemical versions of chaperone have been developed in search of drug candidates for ER stress-related diseases. In this review, synthetic or semi-synthetic chemical chaperones are categorized according to potential therapeutic area and listed along with their chemical structure and activity. Although only a few chemical chaperones have been approved as pharmaceutical drugs, a dramatic increase in literatures over the recent decades indicates enormous amount of efforts paid by many researchers. The efforts warrant clearer understanding of ER stress and the related diseases and consequently will offer a promising drug discovery platform with chaperone activity.
Keywords: endoplasmic reticulum stress, unfolded protein response, chemical chaperone, drug discovery, diabetes, cardiovascular disease, neurodegeneration, lysosomal storage disease
Introduction
The endoplasmic reticulum (ER) is a cellular organelle in which folding of de novo synthesized proteins occurs. The ER plays an important role in homeostasis of proteins and calcium.1 Ribosomes on rough ER synthesize proteins based on the genetic information transferred by mRNA. Smooth ER does not have ribosomes but has neighboring Golgi bodies. The rough ER folds nascent proteins and transports them to the Golgi. Protein homeostasis is tightly controlled by various cellular mechanisms, and failure or error of these quality control systems results in cellular dysfunction. The ER enters a stressed condition when nascent proteins are misfolded or unfolded and abnormally accumulate in the lumen of the ER, leading to failure to transfer to the Golgi. Upon sensing misfolded proteins, a series of cellular events known as unfolded protein response (UPR) or ER stress response is triggered in order to adapt to the cellular damage caused by ER stress. The UPR consists of canonical cellular processes2 such as a decrease in translation to prevent further production of misfolded proteins, upregulation of chaperones to assist the folding process, ER-associated degradation (ERAD), and apoptosis. A successful rescue process can result in cell survival; a failed rescue can drive cells to apoptosis in order to reduce the risk of wasting precious amino acids and energy (Figure 1). ER stress is especially important in cells in which a high level of protein synthesis constantly occurs. In this context, insulin-secreting pancreatic β-cells and cancer cells could be more susceptible to ER stress than other cells.
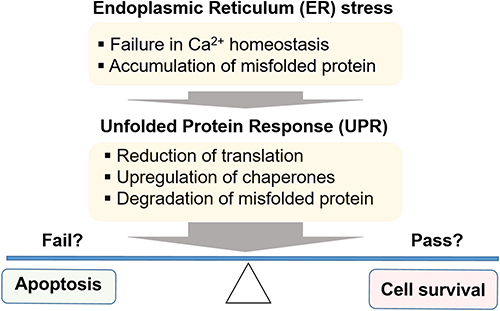 |
Figure 1 ER stress and UPR process to adapt stress condition: survival or cell death? |
ER stress is implicated in various diseases such as diabetes, β-cell apoptosis, diabetic neuropathy, inflammation, cardiovascular disease, neurodegeneration, and lysosomal storage diseases.3–6 A number of studies have suggested an intriguing role of ER stress to induce β-cell apoptosis for initiation and maintenance of diabetes.3,7 ER stress is detected by three well-conserved ER-resident sensor molecules: protein kinase RNA-activated (PKR)-like ER kinase (PERK), activating transcription factor 6 (ATF6), and inositol-requiring enzyme 1α (IRE1α).8 These factors relay complex and interrelated downstream signaling pathways that decide cell ‘survival’ (adaptive signaling) or “suicide” (apoptotic signaling). The activated PERK dimerizes and phosphorylates eukaryotic translation initiation factor 2α (eIF2α), which attenuates protein synthesis and increases the expression of ATF4 to upregulate apoptosis-inducing factors such as C/EBP homology protein (CHOP). It is noteworthy that ATF4 in general relays an adaptive signal to upregulate genes that promote ER homeostasis and survival. ATF6 is also involved in activating the transcription of many UPR mediators including an ER chaperone protein, glucose-regulated protein (GRP78).9 IRE1α processes X-box binding protein 1 (XBP1) mRNA to produce an active spliced form, which becomes a competent transcription factor for UPR-related genes. Chaperone molecules such as GRP78 (or BiP)10 play an important role in UPR to aid proteins in achieving a functional folding conformation. It is believed that chaperones partially or fully bind to the unfolded protein polypeptides, preventing aggregation or incorrect folding.11 Chemical versions of chaperones, chemical chaperones,12 have been identified and implicated as potential treatments for ER stress-related pathologies. The best known examples of chemical chaperones are shown in Figure 2. 4-phenylbutyric acid (4-PBA, 1) and tauro-ursodeoxycholic acid (t-UDCA, 2) provided therapeutic value for several ER stress-related conditions such as type 2 diabetes (T2D). Both compounds protected against ER stress. They suppressed tunicamycin (Tm)-induced phosphorylation of PERK and eIF2α and JNK activation in cells. XBP-1 mRNA was also markedly reduced by both of them. Their reduction of ER stress and recovery of insulin sensitivity in animals were able to strongly support correlation between ER-stress and T2D, suggesting ER stress being pathological cause of T2D and therapeutic alternative.13–15 Salubrinal (3) has also been found to protect cells from ER stress by protecting eIF2α from dephosphorylation, one of the hallmark events of UPR.16 More recently, high throughput screening (HTS) cell-based assays have been developed to identify novel chemical chaperones17–21 that prevent ER stress aggregation, proteotoxicity, and UPR. Consequently, several chemical chaperones such as compound 4 (IBT21) were identified.17 In contrast to chaperones that reduce ER stress to rescue cells, there is a group of chemicals that interfere or inhibit chaperone salvaging activity and induce cell death. Representative examples of this class include heat shock protein (HSP) inhibitors, which inhibit chaperone activity of HSP7022,23 and HSP 90,24–26 for use as anticancer agents.27,28
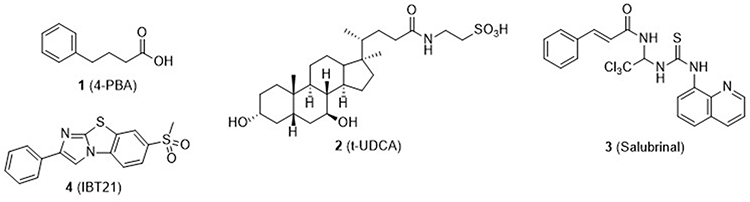 |
Figure 2 Structures of representative chemical chaperones found in literature. These compounds reduced ER stress and alleviated ER stress-related pathologies such as metabolic diseases (4-PBA, t-UDCA, and Salubrinal) and proteotoxicity induced by chemical (tunicamycin) and proteotoxin (mutant prion proteins) (IBT29). |
In this review, chemical chaperones related to pathologies caused by ER stress are described along with their structures and activities. A major part of the review covers synthetic compounds and synthetic derivatives of natural compounds as summarized in Table 1. It should be noted that Table 1 also displays the chaperone activity of those compounds to regulate ER stress signaling molecules.
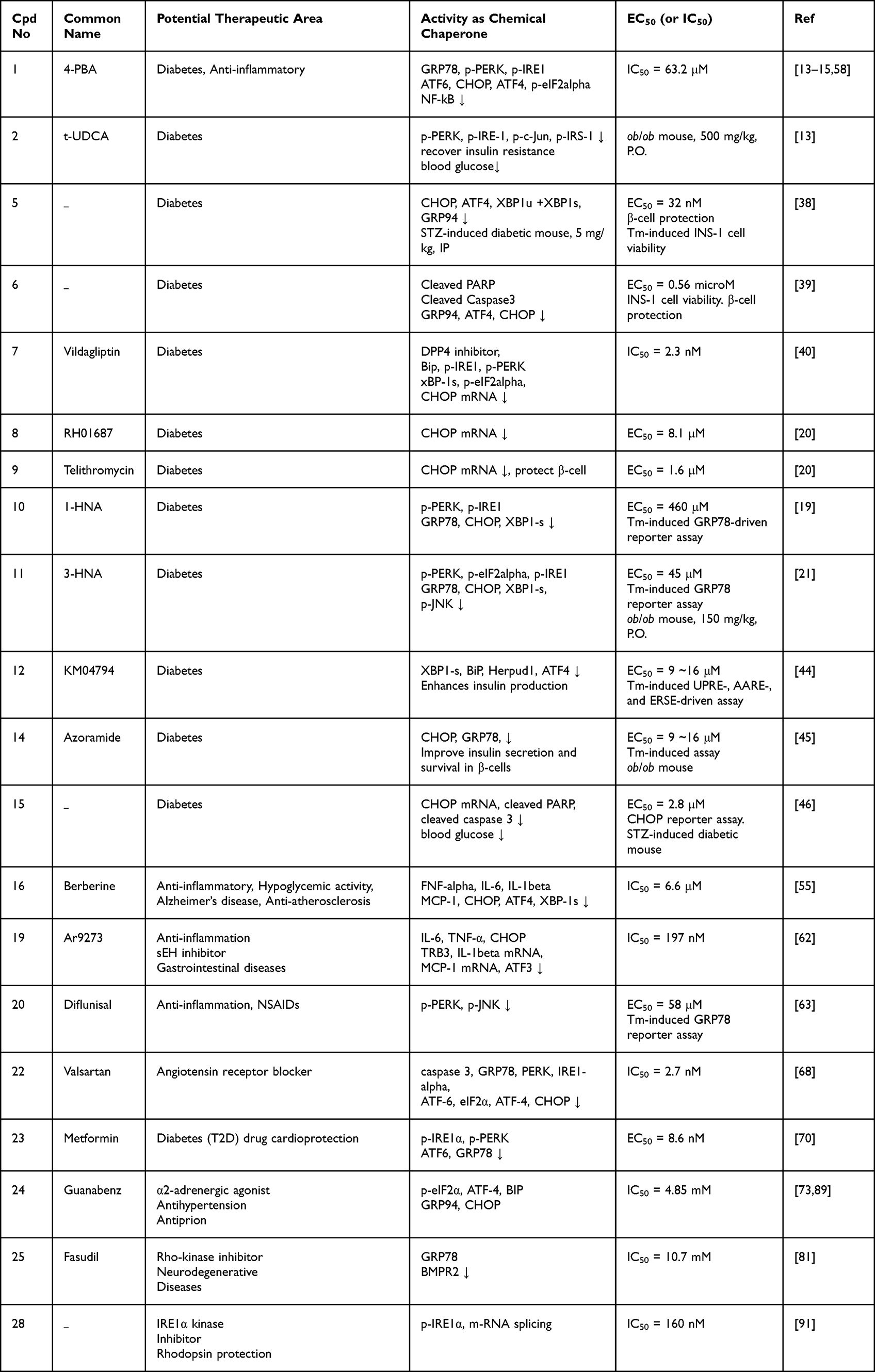 |
Table 1 Activity of Compounds as Chemical Chaperones in Various Pathologies |
Type 2 Diabetes (T2D)
One of the key features of T2D is increased insulin resistance. A high incidence of T2D is observed in the obese population whose elevated level of free fatty acids is attributed to the pathology. Although the precise mechanism is unclear, presence of free fatty acids in obesity patients is a well-established risk factor of ER stress and insulin resistance.10,29–33 These fatty acids cause the accumulation of misfolded proteins in the ER and induce UPR, leading to insulin resistance (Figure 3).
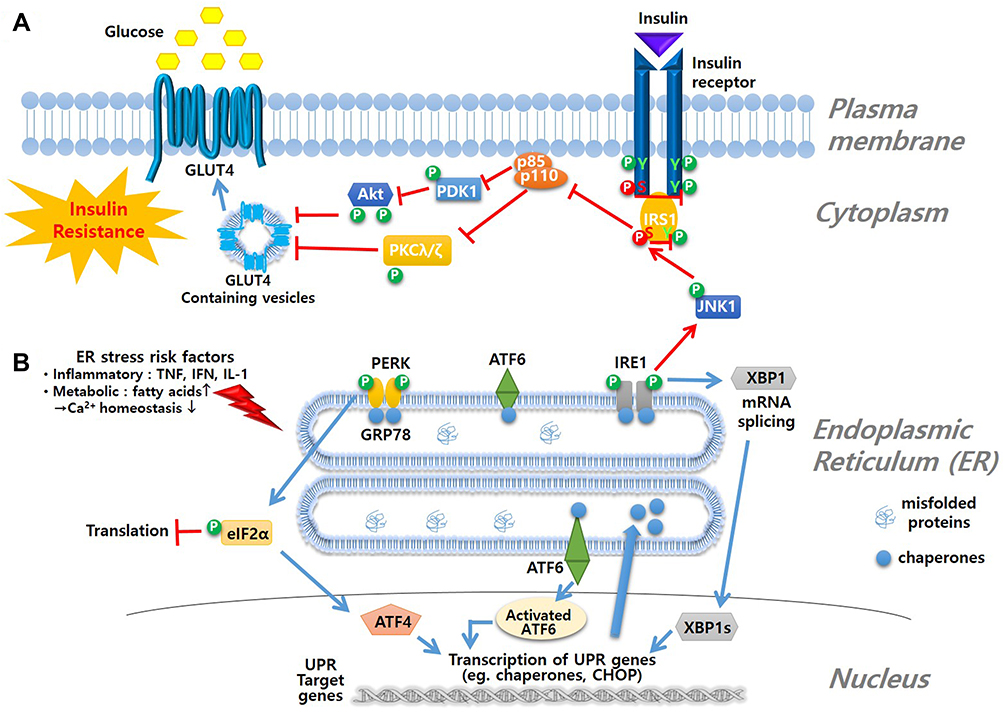 |
Figure 3 Mechanism of ER stress and insulin resistance. (A) insulin signal pathway is blocked due to ER stress-induced phosphorylation of JNK1 followed by phosphorylation of serine moiety of IRS1 to inhibit glucose influx as a result (B) ER stress is triggered by various risk factors that interrupt Ca2+ homeostasis. Accumulation of misfolded proteins in lumen of ER induces ER-resident membrane sensor molecules, such as PERK, ATF6 and IRE1, to initiate the UPR signal starting with dimerization and autophosphorylation of the sensing molecules. UPR signals lead to inhibition of translation, activation of UPR genes such as chaperones and CHOP, or degradation of misfolded protein by ER-associated protein degradation (ERAD) process. |
Under normal conditions, binding of insulin to its receptor initiates a signaling pathway via autophosphorylation of tyrosine residues of the receptor and consequent phosphorylation of tyrosine residues of the insulin receptor substrate (IRS1). These initial events result in relocation of cytosolic glucose transporter 4 (GLUT4) to the cell membrane where it transports plasma glucose into cells through complex signaling. Under ER stress, however, IRE1 phosphorylation caused by UPR induces phosphorylation of serine moieties of IRS1 via c-Jun N-terminal kinase 1 (JNK1) phosphorylation, which inhibits the phosphorylation of tyrosine residues of IRS1 (IRE1-JNK-IRS signaling axis). This sequence leads to blockage of the signaling pathway, causing failure of glucose influx, namely insulin resistance (Figure 3A). With increased insulin resistance, pancreatic β-cells produce additional insulin beyond the folding capacity of the ER, leading to ER stress. The pancreatic β-cell function is diminished in the T1D condition mainly via an autoimmune process, but ER stress is also implicated in deterioration of β-cell function, and some chemical chaperones have been shown to protect β-cells.8,14,34,35 There is a group of ER stress reducing chemicals that have shown anti-diabetic indications such as protection against insulin resistance and of β-cells (Figure 3).
Figure 4 lists compounds with chaperone activity as well as anti-diabetic activity. Salubrinal (3) is a well-known ER stress inhibitor and eIF2α-dephosphorylation inhibitor16 and extends inhibition of translation. However, its activity is controversial and cell-type dependent. It was reported that 1 protects pheochromocytoma PC12 cells against ER stress-induced apoptosis but triggers apoptosis in pancreatic β-cells.27,36 Duan et al synthesized a series of benzamide derivatives and screened for potential ER stress inhibiting activity.37 Of the derivatives, a benzamide 5 was found to have EC50 = 32 nM in protecting INS-1 cells from Tm-induced ER stress. 5 showed remarkable activity to protect β-cells from Tm-induced ER stress and downregulated ER stress markers including ATF4, CHOP, XBP1s, and BiP. In addition, 5 significantly lowered blood glucose and increased β-cell survival in a streptozotocin (STZ)-induced diabetic mouse model. Another report by that group disclosed a 2,4-diaminoquinazoline 6 with an EC50 value in the micromolar range. 6 also downregulated ER stress markers such as ATF4, CHOP, XBP1s, and BiP and cleaved PARP and caspase 3.38 Compound 7 (Vildagliptin) is a marketed drug that acts as a dipeptidyl peptidase-4 (DPP-4) inhibitor in diabetes. It inhibits the DPP-4 enzyme, increases GLP-1 activity, and stimulates insulin release. Thapsigargin (Tg)-induced ER stress in the liver was alleviated by Vildagliptin.39 Chemical chaperones that protect pancreatic β-cells from ER stress-induced apoptosis were searched through an HTS campaign based on the viability of the mouse insulinoma β cell line (βTC6) upon treatment with Tm. Of 17,600 compounds screened, 8 (RH01687) and 9 (Telithromycin) were identified as active hits.20 Both compounds modulated ER stress induced by Tm and protected β-cells, with EC50 = 8.1 and 1.6 μM, respectively. Telithromycin (9) is an erythromycin analogue antibiotics known to be an effective treatment for pneumonia.40 Another HTS assay consists of the rLuc reporter driven by the human GRP78 promoter harboring three consecutive ER stress response elements. Using the cell-based assay system, Jeong et al identified a series of hydroxynaphthoic acids as chemical chaperones.19 Compound 10 (1-hydroxy-3-naphthoic acid, 1-HNA) inhibited Tm- and palmitate-induced ER stress, with EC50 = 460 μM and 60 μM, respectively. ER stress markers such as p-PERK, p-IRE1, GRP78, CHOP, and XBP1-s were decreased accordingly. The anti-diabetic activity of compound 11 (3-HNA) was further studied by Park et al.21 The compound showed Tm- and palmitate-induced ER stress inhibition, with EC50 = 570 μM and 45 μM, respectively, and downregulated ER stress markers. Oral administration of 11 (150 mg/kg) to ob/ob mice resulted in resolution of insulin resistance and protection of β-cells from apoptosis induced by glucolipotoxicity. t-UDCA (2) is a bile acid that has proven to have medicinal effects in T2D, various heart diseases, and neurodegenerative diseases (Alzheimer’s and Amyotrophic lateral sclerosis (ALS)).41–43 Compounds 1 (4-PBA) and 2 (t-UDCA) showed anti-diabetic activities such as i) recovery of insulin sensitivity and ii) decreases in blood glucose and insulin secretion levels after oral administration to ob/ob mice at a dose of 500 mg/kg.13–15
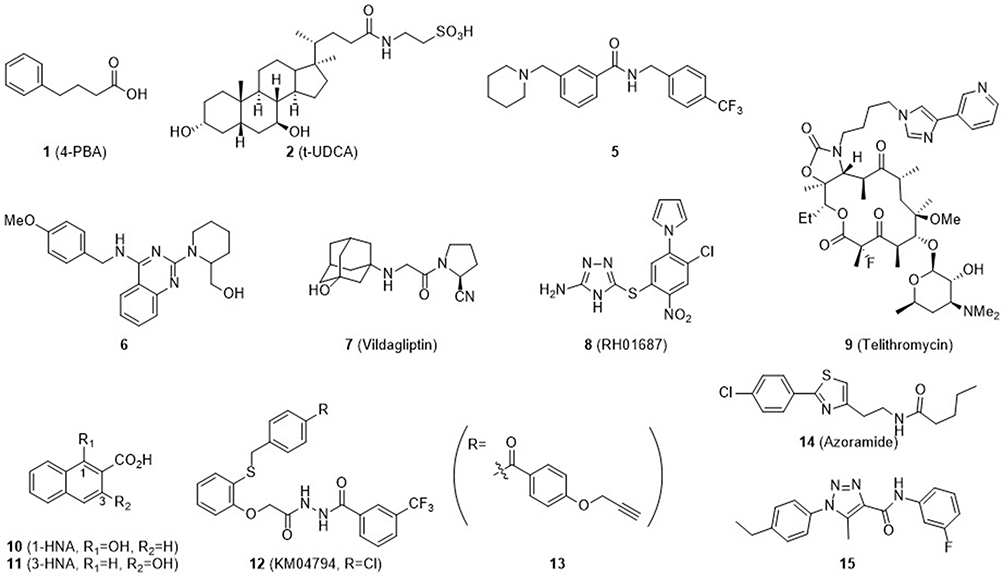 |
Figure 4 Chemical structures of compounds that attenuated ER stress as chemical chaperone and showed anti-diabetic activity. Many of them demonstrated anti-diabetic activity in vivo (4-PBA, t-UDCA, Azoramide, compound 5, 3-HNA, KM04794, compound 15). Note that Vildagliptin (7), a marketed anti-diabetic drug and DPP-4 inhibitor, showed chaperone activity to reduce ER stress. |
Miyake et al established a series of cell-based assays to identify ER stress inhibitors.44 They used ER stress response element (ERSE)-, unfolded protein response element (UPRE)-, and amino acid-response element (AARE)-based assay systems for parallel monitoring of three major ER stress signaling pathways: IRE1, PERK, and ATF6 pathways. Compound 12 (KM04794) was identified as an ER proteostasis modulator that inhibited UPR signaling caused by ER stress induced by diverse chemicals. KM04794 (12) alleviated protein aggregation and enhanced insulin production in pancreatic β-cells. In addition, compound 13 was prepared by replacing the terminal 4-chlorophenyl group of the parent compound, 12, with a pull-down moiety. Benzophenone and propynyl groups were tethered onto 13 as a photoaffinity tag and click reaction tag, respectively. Using 12 as a competitor molecule, a pull-down experiment was carried out and revealed BiP as the most probable binding partner of compound 13. BiP is one of the most important chaperones and seems to directly bind to 12 to produce an improvement of ER proteostasis. Another HTS assay consisting of cLuc activity driven by ATF6α identified 14 (Azoramide) as a modulator of UPR with anti-diabetic activity.45 14 decreased CHOP and GRP78 under Tm-induced ER stress. In ob/ob mice and high fat diet-induced obese mice, administration of 14 improved insulin secretion and survival of β-cells. Duan et al reported a 1,2,3-triazole derivative (15) that protected pancreatic β cells against endoplasmic reticulum stress-mediated dysfunction and death through inhibition of C/EBP-homologous protein expression.46
Inflammatory Diseases92
As shown in Figure 5, there are a number of compounds that possess chaperone activity as well as anti-inflammatory activity. Compound 16 (Berberine) is a naturally occurring compound that has various physiological effects such as hypoglycemic, anti-microbial, anti-inflammatory, anti-Alzheimer, and anti-atherosclerosis activities.47–52 Among its derivatives, 17 was reported to have anti-breast cancer activity, which also displayed ER stress inhibition.53 Study of ER stress reduction on mitochondrial dysfunction and inflammation have been reported.54 Berberine (16) inhibited palmitate (PA)- and lipopolysaccharide (LPS)–induced inflammation through modulation of ER stress.55
 |
Figure 5 Chemical chaperones to relieve the ER stress with anti-inflammatory activity. |
Compound 1 (4-PBA) is an inhibitor of histone deacetylase (HDAC),56 which is involved in various diseases such as metabolic syndrome (obesity, T2D), misfolding diseases (cystic fibrosis), inflammatory disorder (diabetic nephropathy), neurological disorder (Parkinson’s), tissue diseases (fibrosis), and cancers (gastric carcinoma, colon cancer). HDAC is also known to inhibit platelet aggregation and is associated with β-globin disorders.57 ER stress suppression was examined in male C57BL/6J mouse hyperoxia-exposed lung epithelial cells. Hyperoxia-induced lung injury was attenuated by treatment with 4-PBA (1), which modulated inflammation-related markers (IkB-α, NF-kB).58 Soluble epoxide hydrogenase (sEH) converts epoxyeicosatrienoic acid (EET) to dihydroxytrienoic acids (DiHETrES), which triggers the inflammation cascade.59 Compound 18 (Ar9281)60,61 has a potent IC50 = 8.0 nM against sEH; however, due to its low water solubility, a more hydrophilic analogue was synthesized (19, AR9273). Although the IC50 of 19 was lower than that of 18 (197 nM), water solubility, microsomal stability, sEH enzyme selectivity, and blood-brain barrier (BBB) penetration were improved. ER stress reduction was observed in the Ce (cerulean)-induced pancreatitis AR42J cell model.62 In a continuing study on salicylate-related ER stress chaperones, our group found that 3-HNA (11), which contains a naphthoic acid scaffold, improves glucose lowering activity through ER stress amelioration.21 Compared to the parent salicylate (EC50=5.07 mM),19 a standard anti-inflammatory drug, extended aromaticity provided by the naphthalene moiety of 3-HNA (11) was credited with its remarkable anti-ER stress activity (EC50=0.57 mM).21 In addition, salicylate analogues with a biphenyl scaffold were synthesized. Of these, 20 (Diflunisal), an anti-inflammatory drug with a biphenyl ring harboring a fluorine substituent, exhibited the best anti-ER stress activity. The EC50 value (EC50 = 58 μM) of Diflunisal (20)63 was approximately 6- and 90-fold higher than 3-HNA (11) (EC50 = 328 μM) and t-UDCA (2) (EC50 = 5.2 mM),64 respectively. Moreover, Diflunisal (20) ameliorated palmitate-induced ER stress and decreased UPR markers.63
Cardiovascular Diseases65,66
Chemical chaperones that inhibit ER stress and provide cardioprotective activity are shown in Figure 6. Doxorubicin (Dox) is an anticancer drug, but it induces cardiotoxicity, resulting in apoptosis of cardiomyocytes, which has been attributed to ER stress.66,67 Compounds 21 (Sacubitril) and 22 (Valsartan) have been used as a combination therapy for treatment of heart failure. Sacubitril is a prodrug and a neprilysin inhibitor, and valsartan is an angiotensin II receptor blocker (ARB). Administration of the Sac/Val combination suppressed ER stress induced by treatment with Dox; UPR markers caspase 3, GRP78, PERK, IRE1α, ATF-6, eIF2α, ATF-4, and CHOP were all decreased.68 Another study investigated the anti-ER stress activity of Valsartan (22) in tubular epithelial cells of diabetic cardiomyopathy rats.69
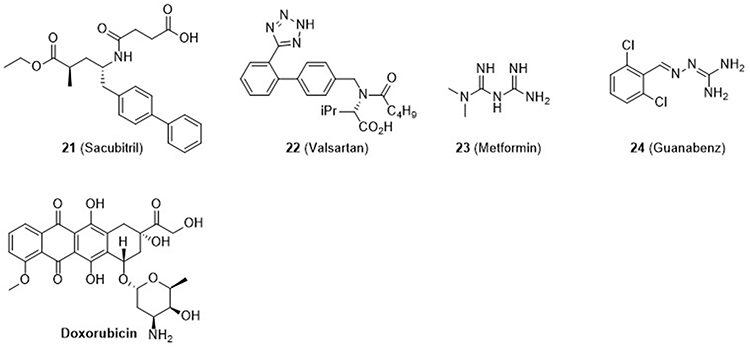 |
Figure 6 Chemical chaperones to relieve the ER stress with cardioprotection activity. |
Compound 23 (Metformin) is effective in T2D treatment, especially when co-administered with sulfonylureas, and is especially beneficial for obese diabetic patients. It has also been shown to be effective in the treatment of cardiovascular disease (CVD).70 The cardioprotective effect of Metformin (23) through ER stress reduction on human coronary artery endothelial cells (HCAEC) has been reported. Metformin (23) suppressed Tm- and high dextrose-induced ER stress by regulating p-IRE1α, p-PERK, and ATF6 activities, leading to cardioprotection.70 Compound 24 (Guanabenz) is an α-2 adrenergic receptor agonist that is used as an antihypertensive drug. Albeit somewhat controversial, it is believed that Guanabenz (24) interferes with dephosphorylation of elF2α-P’ by disrupting the PPPIR15-PP1 complex.71,72 Guanabenz (24) did not show any noticeable effect alone, but when used with ER stress inducer (tunicamycin), it provided lower levels of ER stress markers such as p-eIF2α, ATF4, BiP, GRP94, and CHOP and eventual cell protection.73
Neurodegeneration and Neuroprotection74,75
Figure 7 shows chemical chaperones that possess neuroprotective activity and anti-neurodegenerative activity. Compound 25 (Fasudil), a Rho-kinase inhibitor, is used as a treatment for cerebral vasospasm, pulmonary hypertension, cardiovascular diseases, age-related neurodegenerative memory loss, corneal neovascularization, and other conditions.75–80 It improves motor function and has been approved in Japan for treatment of cerebral vasospasm following subarachnoid hemorrhage.81 Several ER stress modulating functions of Fasudil (25) have been reported; i) ischemia/reperfusion injury through SERCA activity,82 ii) inhibition of vascular cellular adhesion molecule (VCAM-1) expression by modulating UPR,83 and iii) inhibition of leukocyte-endothelial interaction through modulation of GRP78 and BMPR2 expression.84 Compound 26 is an HSP70 agonist showing chaperone activity. It produced a clear reduction in α-synuclein aggregation in neuroglioma cells, which is a hallmark of Parkinson’s disease.85–87 Oxindole 27 suppressed protein aggregate accumulation in vitro and in hippocampal HT22 neuronal cells and prevented ER stress-induced cell death as a chemical chaperone.88 Guanabenz (24), a antihypertensive drug, promoted ovine PrPsc clearance in a cell-based assay, increasing the survival of treated mice significantly. Interestingly, other α2-adrenergic agonists did not show such anti-prion activity.89
 |
Figure 7 Chemical chaperones to inhibit ER stress with neuroprotection activity. |
Miscellaneous Diseases
In addition to the diseases discussed above, ER stress is involved in other pathologies, and compounds with chaperone activity have been developed and characterized (Figure 8). It would be rather appropriate to consider IRE1 kinase as a core UPR component whose inhibitors can make a huge subset collection of kinase inhibitors. It will be beyond the scope of this review to describe the vast list of IRE1 kinase inhibitors. We here would like to provide a few examples of IRE1 kinase inhibitors that showed ER stress inhibition. Inhibitors of IRE1α kinase, a major signaling molecule of UPR, were developed to treat neurodegenerative cancer, diabetes, lipidemia, and inflammatory diseases.8,90 Compound 28 was discovered through docking studies and has IC50 = 160 nM and 80 nM against IRE1α kinase and IRE1α RNAse, respectively. It is highly selective for IRE1α kinase over the IRE1β isoform, although interference of kinase activity by compound 28 was marginal in a kinome assay.91 ER stress can occur in photoreceptors as well. Ocular protein conformational diseases such as retinitis pigmentosa can be caused by misfolded or mistrafficked rhodopsin, a complex of opsin protein and retinal, which aggregate and accumulate in the ER, leading to photoreceptor cell death. Compound 29 binds to opsin, a visual pigment, as a chemical chaperone, assisting its proper folding and trafficking to the outer cell membrane and preventing the loss of photoreceptors under ER stress conditions.93
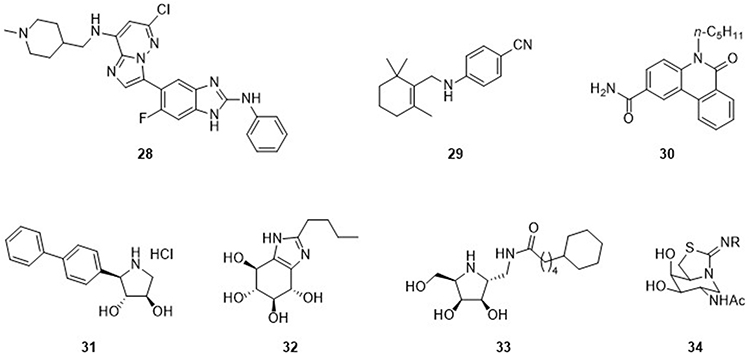 |
Figure 8 Chemical chaperones to inhibit ER stress with therapeutic implications in other diseases. |
Chemical chaperones have further been implicated in lysosomal storage disorders such as Gaucher disease,94 Fabry disease, and Tay-Sachs disease, in which trafficking of proteins or lipids is disrupted. Compound 25 was synthesized and tested in Niemann-Pick disease type C195 (NPC1), a disease characterized by abnormal accumulation of lipids and cholesterol in lysosomal and late endosomal compartments. Compound 30 alleviated the folding defect of the NPC1 protein I1061T mutant, resulting in transport of misfolded mutant NPC1 to late endosomes similar to normal NPC1. Compound 31 is an imino sugar analogue that inhibited β-glucosidase or β-glucocerebrosidase (GCase) that cleaves glucose from gluco(syl)ceramide (sphingolipid). Defects of this enzyme cause accumulation of glucoceramide, the genesis of Gaucher disease. Compound 31-assisted folding of mutant enzyme prevents its degradation by the ER proteostasis function.96 Compound 32 also strongly inhibited β-glucosidase.97 Similarly, a defect in lysosomal α-galactosidase A (α-Gal A) causes Fabry disease due to accumulation of neutral glycosphingolipids bearing a terminal α-galactosyl residue. Compound 33 showed a chaperone effect for several α-Gal A mutants in COS7 cells and lymphocytes of the N215S Fabry patient-derived cell line.98
In the late-onset form of Tay-Sachs disease, defects were found in N-acetyl-β-hexosaminidase A (HexA) that catalyzes the removal of terminal, non-reducing N-acetyl-β-D-glucosamine (GlcNAc) or galactosamine (GalNAc). Mutational defects impair folding of the enzyme, resulting in its normal trafficking to lysosome. Compound 34 was found to be a competitive inhibitor of HexA and rescued disease-causing mutant HexA, showing potential as a chemical chaperone. It increased the activity of lysosomal HexA in Tay-Sachs patient fibroblasts containing the G269S mutation, the most prevalent mutation in late-onset Tay-Sachs disease.99
Conclusion and Perspectives
ER stress is implicated in various diseases and pathologies. We listed in this review chemical versions of chaperone molecules and categorized them into potential therapeutic areas such as diabetes, inflammatory diseases, cardiovascular diseases, and neurodegeneration. In addition, lysosomal storage diseases have shed new light on the therapeutic value of chemical chaperones. Many compounds with chaperone activity have shown therapeutic potential in rescuing misfolded proteins caused by ER stress or genetic defects. Various HTS assays are currently available, which may inspire researchers to conduct phenotypic screening using large volume chemical libraries. Target-based medicinal chemistry efforts can also be undertaken against specific targets. ER stress sensing molecules such as PERK inhibitors have been developed and examined clinically.100,101 Recent studies have shown that PERK signaling plays a critical role in immunosuppression in macrophages, opening a new era into the role of chemical chaperone inhibitors in the immune system.102 It was also reported that STING-PERK signaling can be an alternative innate immune pathway, which plays a critical role in fibrotic diseases.103 Other prevalent targets of chemical chaperones include HSP’s as mentioned in this review. Inhibitors of HSP’s have mostly been pursued for their therapeutic value as anticancer agents. Targets including GRP78 or BiP could also be novel candidates. BiP was identified as a direct binding partner of a chemical chaperone during a target identification effort using the pull-down probe, compound 13. An increasing number of chemical chaperones is being developed, and we hope this review and concise list of currently identified structures provides structural inspiration for new chemical chaperones.
Acknowledgments
This work was supported by National Research Foundation of Korea grants NRF-2019R1F1A1060071, NRF-2020R1A6A1A03042854 (Center for Proteinopathy), and an Institute of Information & Communications Technology Planning & Evaluation (IITP) grant funded by the Korean government (MSIT) (No. 2020-0-01343, Artificial Intelligence Convergence Research Centre (Hanyang University ERICA)).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Sarvani C, Sireesh D, Ramkumar KM. Unraveling the role of ER stress inhibitors in the context of metabolic diseases. Pharmacol Res. 2017;119:412–421. doi:10.1016/j.phrs.2017.02.018
2. Hetz C, Chevet E, Harding HP. Targeting the unfolded protein response in disease. Nat Rev Drug Discov. 2013;12:703–719. doi:10.1038/nrd3976
3. Marciniak SJ, Chambers JE, Ron D. Pharmacological targeting of endoplasmic reticulum stress in disease. Nat Rev Drug Discov. 2022;21(2):115–140. doi:10.1038/s41573-021-00320-3
4. Wang M, Kaufman RJ. Protein misfolding in the endoplasmic reticulum as a conduit to human disease. Nature. 2016;529(7586):326–335. doi:10.1038/nature17041
5. Kim I, Xu W, Reed JC. Cell death and endoplasmic reticulum stress: disease relevance and therapeutic opportunities. Nat Rev Drug Discov. 2008;7(12):1013–1030.
6. Fan Y, Zhang J, Xiao W, et al. Rtn1a-mediated endoplasmic reticulum stress in podocyte injury and diabetic nephropathy. Sci Rep. 2017;7(1):323.
7. Lee JH, Lee J. Endoplasmic reticulum (ER) stress and its role in pancreatic b-cell dysfunction and senescence in type 2 diabetes. Int J Mol Sci. 2022;23(9):4843.
8. Ron D, Hubbard SR. How IRE1 reacts to ER stress. Cell. 2008;132:24–26.
9. Haze K, Yoshida H, Yanagi H, Yura T, Mori K. Mammalian transcription factor ATF6 is synthesized as a transmembrane protein and activated by proteolysis in response to endoplasmic reticulum stress. Mol Biol Cell. 1999;10:3787–3799.
10. Lièvremont JP, Rizzuto R, Hendershot L, Meldolesi J. BiP, a major chaperone protein of the endoplasmic reticulum lumen, plays a direct and important role in the storage of the rapidly exchanging pool of Ca2+. J Biol Chem. 1997;272(49):30873–30879. doi:10.1074/jbc.272.49.30873
11. Sucec I, Bersch B, Schanda P. How do chaperones bind (partly) unfolded client proteins? Front Mol Biosci. 2021;8:762005. doi:10.3389/fmolb.2021.762005
12. Papp E, Csermely P. Chemical chaperones: mechanisms of action and potential use. In: Starke K, Gaestel M, editors. Molecular Chaperones in Health and Disease. Handbook of Experimental Pharmacology. Vol. 172. Berlin, Heidelberg: Springer; 2006:405–416.
13. Ozcan U, Yilmaz E, Ozcan L, et al. Chemical chaperones reduce ER stress and restore glucose homeostasis in a mouse model of type 2 diabetes. Science. 2006;313(5790):1137–1140. doi:10.1126/science.1128294
14. Engin F, Yermalovich A, Nguyen T, et al. Restoration of the unfolded protein response in pancreatic β cells protects mice against type 1 diabetes. Sci Transl Med. 2013;5(211):211ra156. doi:10.1126/scitranslmed.3006534
15. Xiao C, Giacca A, Lewis GF. Sodium phenylbutyrate, a drug with known capacity to reduce endoplasmic reticulum stress, partially alleviates lipid-induced insulin resistance and beta-cell dysfunction in humans. Diabetes. 2011;60(3):918–924. doi:10.2337/db10-1433
16. Boyce M, Bryant KF, Jousse C, et al. A selective inhibitor of eIF2a dephosphorylation protects cells from ER stress. Science. 2005;307:935–939. doi:10.1126/science.1101902
17. Kitakaze K, Taniuchi S, Kawano E, et al. Cell-based HTS identifies a chemical chaperone for preventing ER protein aggregation and proteotoxicity. Elife. 2019;8:e43302. doi:10.7554/eLife.43302
18. Ghafouri M, Gauss CB, Fribley AM. HTS identification of activators and inhibitors of endoplasmic reticulum (ER) stress and the unfolded protein response (UPR). Methods Mol Biol. 2022;2378:317–327.
19. Jeong KW, Ku JM, Park MW, Park SM, Yang JE, Nam TG. Hydroxynaphthoic acids in a high throughput screening potently ameliorate endoplasmic reticulum stress as novel chemical chaperones. Chem Pharm Bull. 2013;6:740–746. doi:10.1248/cpb.c13-00251
20. Tran K, Li Y, Duan H, Arora D, Lim HY, Wang W. Identification of small molecules that protect pancreatic β cells against endoplasmic reticulum stress-induced cell death. ACS Chem Biol. 2014;9:2796–2806. doi:10.1021/cb500740d
21. Park SM, Choi J, Nam TG, Ku JM, Jeong K. Anti-adiabetic effect of 3-hydroxy-2-naphthoic acid, an endoplasmic reticulum stress-reducing chemical chaperone. Eur J Pharmacol. 2016;779:157–167. doi:10.1016/j.ejphar.2016.03.023
22. Huang Q, Johnson TW, Bailey S, et al. Design of potent and selective inhibitors to overcome clinical anaplastic lymphoma kinase mutations resistant to crizotinib. J Med Chem. 2014;57(4):1170–1187. doi:10.1021/jm401805h
23. Taldone T, Kang Y, Patel HJ, et al. Heat shock protein 70 inhibitors. 2. 2,5’-thiodipyrimidines, 5-(phenylthio)pyrimidines, 2-(pyridin-3-ylthio)pyrimidines, and 3-(phenylthio)pyridines as reversible binders to an allosteric site on heat shock protein 70. J Med Chem. 2014;57(4):1208–1224. doi:10.1021/jm401552y
24. Jiang F, Wang HJ, Jin YH, et al. Novel tetrahydropyrido[4,3-d]pyrimidines as potent inhibitors of chaperone heat shock protein 90. J Med Chem. 2016;59(23):10498–10519. doi:10.1021/acs.jmedchem.6b00912
25. Uno T, Kawai Y, Yamashita S, et al. Discovery of 3-ethyl-4-(3-isopropyl-4-(4-(1-methyl-1 H-pyrazol-4-yl)-1H-imidazol-1-yl)-1H-pyrazolo[3,4- b]pyridin-1-yl)benzamide (Tas-116) as a potent, selective, and orally Available HSP90 Inhibitor. J Med Chem. 2019;62(2):531–551. doi:10.1021/acs.jmedchem.8b01085
26. Huang DS, LeBlanc EV, Shekhar-Guturja T, et al. Design and synthesis of fungal-selective resorcylate aminopyrazole Hsp90 inhibitors. J Med Chem. 2020;63(5):2139–2180. doi:10.1021/acs.jmedchem.9b00826
27. Chevet E, Hetz C, Samali A. Endoplasmic reticulum stress-activated cell reprogramming in oncogenesis. Cancer Discov. 2015;5(6):586–597. doi:10.1158/2159-8290.CD-14-1490
28. Ma Y, Hendershot LM. The role of the unfolded protein response in tumour development: friend or foe? Nat Rev Cancer. 2004;4(12):966–977. doi:10.1038/nrc1505
29. Meyerovich K, Ortis F, Allagnat F, Cadozo AK. Endoplasmic reticulum stress and the unfolded protein response in pancreatic islet inflammation. J Mol Endocrinol. 2016;57(1):R1–R17. doi:10.1530/JME-15-0306
30. Hasnain SZ, Prins JB, McGuckin MA. Oxidative and endoplasmic reticulum stress in β-cell dysfunction in diabetes. J Mol Endocrinol. 2016;56(2):R33–R54. doi:10.1530/JME-15-0232
31. Yong J, Johnson JD, Arvan P, et al. Therapeutic opportunities for pancreatic β-cell ER stress in diabetes mellitus. Nat Rev Endocrinol. 2021;17(8):455–467. doi:10.1038/s41574-021-00510-4
32. Lemmer IL, Willemsen N, Hilal N, Bartelt A. A guide to understanding endoplasmic reticulum stress in metabolic disorders. Mol Metab. 2021;47:101169. doi:10.1016/j.molmet.2021.101169
33. Bilekova S, Sachs S, Lickert H. Pharmacological targeting of endoplasmic reticulum stress in pancreatic beta cells. Trends Pharmacol Sci. 2021;42(2):85–95. doi:10.1016/j.tips.2020.11.011
34. Eizirik DL, Cardozo AK, Cnop M. The role for endoplasmic reticulum stress in diabetes mellitus. Endocr Rev. 2008;29:42–61. doi:10.1210/er.2007-0015
35. Tersey SA, Nishiki Y, Templin AT, et al. Islet beta-cell endoplasmic reticulum stress precedes the onset of type 1 diabetes in the nonobese diabetic mouse model. Diabetes. 2012;61:818–827. doi:10.2337/db11-1293
36. Cnop M, Ladriere L, Hekerman P, et al. Selective inhibition of eukaryotic translation initiation factor 2 dephosphorylation potentiates fatty acid-induced endoplasmic reticulum stress and causes pancreatic cell dysfunction and apoptosis. J Biol Chem. 2007;282(6):3989–3997. doi:10.1074/jbc.M607627200
37. Duan H, Li Y, Arora D, Xu D, Lim HY, Wang W. Discovery of a benzamide derivative that protects pancreatic β-cells against endoplasmic reticulum stress. J Med Chem. 2017;60:6191–6204. doi:10.1021/acs.jmedchem.7b00435
38. Duan H, Lee JW, Moon SW, et al. Discovery, synthesis, and evaluation of 2,4-diaminoquinazolines as a novel class of pancreatic β-cell-protective agents against endoplasmic reticulum (ER) stress. J Med Chem. 2016;59:7783–7800. doi:10.1021/acs.jmedchem.6b00041
39. Ma X, Du W, Shao S, Yu C, Zhou L, Jing F. Vildagliptin can alleviate endoplasmic reticulum stress in the liver induced by a high fat diet. Biomed Res Int. 2018;2018:5045182. doi:10.1155/2018/5045182
40. Serafini M, Pirali T, Tron GC. Click 1,2,3-triazoles in drug discovery and development: from the flask to the clinic? Adv Heterocycl Chem. 2021;134:101–148.
41. Cadavez L, Montane J, Alcarraz-Vizan G, et al. Chaperones ameliorate beta cell dysfunction associated with human islet amyloid polypeptide overexpression. PLoS One. 2014;9:e101797. doi:10.1371/journal.pone.0101797
42. Purkayastha S, Zhang H, Zhang G, Ahmed Z, Wang Y, Cai D. Neutral dysregulation of peripheral insulin action and blood pressure by brain endoplasmic reticulum stress. PNAS USA. 2011;108:2939–2944. doi:10.1073/pnas.1006875108
43. Yoon YM, Lee JH, Yun SP, et al. Taurosodeoxycholic acid reduces ER stress by regulating of Akt-dependent cellular prion protein. Sci Rep. 2016;6:1–6. doi:10.1038/srep39838
44. Miyake M, Sobajima M, Kurahashi K, et al. Identification of an endoplasmic reticulum proteostasis modulator that enhances insulin production in pancreatic β cells. Cell Chem Biol. 2022;29:996–1009. doi:10.1016/j.chembiol.2022.01.002
45. Fu S, Yalcin A, Lee GY, et al. Phenotypic assays identify a small molecule modulator of the unfolded protein response with anti-diabetic activity. Sci Transl Med. 2015;7(292):292ra98. doi:10.1126/scitranslmed.aaa9134
46. Duan H, Arora D, Li Y, et al. Identification of 1,2,3-triazole derivatives that protect pancreatic β cells against endoplasmic reticulum stress-mediated dysfunction and death through the inhibition of C/EBP-homologous protein expression. Bioorg Med Chem. 2016;24(12):2621–2630. doi:10.1016/j.bmc.2016.03.057
47. Kong Y, Yi YJ, Liu XQ, Yu P, Zhao LG, Li -D-D. Discovery and structural optimization of 9-O-phenylsulfonyl-beberines as new lipid-lowering agents. Bioorg Chem. 2022;121:105665. doi:10.1016/j.bioorg.2022.105665
48. Li R, Wu J, He Y, Hai L, Wu Y. Synthesis and in vitro evaluation of 12-(substituted aminomethyl)berberrubine derivatives as anti-diabetics. Bioorg Med Chem Lett. 2014;24:1762–1765.
49. Zhang S, Wang X, Yin W, et al. Synthesis and hypoglycermic activity of 9-O-(lipophilic group substituted)berberine derivatives. Bioorg Med Chem Lett. 2016;26:4799–4803.
50. Huang L, Shi A, He F, Li X. Synthesis, biological evaluation, and molecular modelling of berberine derivatives as potent acetylcholinesterase inhibitors. Bioorg Me Chem. 2010;18:1244–1251.
51. Huang L, Luo Z, He F, Lu J, Li X. Synthesis and biological evaluation of a new series of berberine derivatives as dual inhibitors of acetylcholinesterase and butylcholinesterase. Bioorg Med Chem. 2010;18:4475–4484.
52. Xiao D, Liu Z, Zhang S, et al. Berberine derivatives with different pharmacological activities via structural modifications. Mini Rev Med Chem. 2018;18:1424–1441.
53. Jin H, Ko YS, Park SW, Chang KC, Kim HJ. 13-Ethylberberine induces apoptosis through the mitochondria-related apoptoic pathway in radiotherapy-resistant breast cancer cells. Molecules. 2019;24:2448.
54. Eom KS, Kim HJ, So HS, Park R, Kim TY. Berberine-induced apoptosis in human glioblastoma T98G cells is mediated by endioplasmic reticulum stress accompanying reactive oxygen species and mitochondrial dysfunction. Biol Pharm Bull. 2010;33:1644–1649.
55. Wang Y, Zhou X, Zhao D, et al. Berberine inhibits free fatty acid and LPS-induced inflammation via modulating ER stress response in macrophages and hepatocytes. PLoS One. 2020;15:e0232630.
56. Tischler JL, Abuaita B, Cuthpert SC, et al. Simple inhibitors of histone deacetylase activity that combine features of short-chain fatty acid and hydroxamic acid inhibitors. J Enzyme Inhib Med Chem. 2008;23:549–555.
57. Kolb PS, Ayaub EA, Zhou W, Yum V, Dickout JG, Ask K. The therapeutic effects of 4-phenylbutyric acid in maintaining proteostasis. Int J Biochem Cell Biol. 2015;61:45–52.
58. Pio HP, Liao WI, Tang SE, Wu SY, Huang KL, Chu SJ. Suppression of endoplasmic reticulum stress by 4-PBA protects against hyperoxia-induced acute lung injury via up-regulating claudin-4 expression. Front Immunol. 2021;12:674316.
59. Hashimoto K. Role of soluble epoxide hydrolase in metabolism of PUFAs in psychiatric and neurological disorders. Front Pharmacol. 2019;10:36.
60. Anandan SK, Webb HK, Chen D, et al. 1-(1-acetyl-piperidin-4-yl)-3-adamantan-1-yl-urea (AR9281) as a potent, selective, and orally available soluble epoxide hydrolase inhibitor with efficacy in rodent models of hypertension and dysglycemia. Bioorg Med Chem Lett. 2011;21(3):983–988.
61. Codony S, Calvó-Tusell C, Valverde E, et al. From the design to the in vivo evaluation of benzohomoadamantane-derived soluble epoxide hydrolase inhibitors for the treatment of acute pancreatitis. J Med Chem. 2021;64(9):5429–5446.
62. Codony S, Pujol E, Pizarro J, et al. 2-Oxaadamant-1-yl ureas as soluble epoxide hydrolase inhibitors: in vivo evaluation in a murine model of acute pancreatitis. J Med Chem. 2020;63:9237–9257.
63. Kim YE, Kim DH, Choi A, et al. Bi-aryl analogues of salicylic acids: design, synthesis and SAR study to ameliorate endoplasmic reticulum stress. Drug Des Devel Ther. 2021;15:3593–3604.
64. Choi H, Yun W, Lee JH, et al. Synthesis and anti-endoplasmic reticulum stress activity of N-substituted-2-arylcarbonylhydrazinecarbothioamides. Med Chem Res. 2019;28:2142–2152.
65. Choy KW, Murugan D, Mustafa MR. Natural products targeting ER stress pathway for the treatment of cardiovascular disease. Pharmacol Res. 2018;132:119–129.
66. Mooradian AD, Haas MJ. Endoplasmic reticulum stress: a common pharmacologic target of cardioprotective drugs. Eur J Pharmacol. 2022;931:175221.
67. Hu J, Wu Q, Wang Z, et al. Inhibition of CACNA1H attenuates doxorubicin-induced acute cardiotoxicity by affecting endoplasmic reticulum stress. Biomed Pharmacother. 2019;120:109475.
68. Kim BS, Park IH, Lee AH, Kim HJ, Lim YH, Shin JH. Sacubitril/valsartan reduces endoplasmic reticulum stress in a rat model of doxorubicin-induced cardiotoxicity. Arch Toxicol. 2022;96:1065–1074.
69. Peng PA, Wang L, Ma Q, et al. Valsartan protects HK-2 cells from contrast media-induced apoptosis by inhibiting endoplasmic reticulum stress. Cell Biol Int. 2015;39:1408–1417.
70. Kapadia P, Bikkina P, Landicho MA, Parekh S, Haas MJ, Mooradian AD. Effect of anti-hypergycemic drugs on endoplasmic reticulum (ER) stress in human coronary artery endothelial cells. Eur J Pharmacol. 2021;15:174249.
71. Crespillo-Casado A, Chambers JE, Fischer PM, Marciniak SJ, Ron D. PPP1R15A-mediated dephosphorylation of eIF2α is unaffected by Sephin1 or Guanabenz. Elife. 2017;6:e26109.
72. Das I, Krzyzosiak A, Schneider K, et al. Preventing proteostasis diseases by selective inhibition of a phosphatase regulatory subunit. Science. 2015;348:239–242.
73. Neuber C, Uebeler J, Schulze T, Sotoud H, El-Armouche A, Eschenhagen T. Guanabenz interferes with ER stress and exerts productive effects in cardiac myocytes. PLoS One. 2014;9:e98893.
74. Bobori C, Theocharopoulou G, Vlamos P. Molecular chaperones in neurodegenerative diseases: a short review. Adv Exp Med Biol. 2017;987:219–231.
75. Gao X, Xu Y. Therapeutic effects of natural compounds and small molecule inhibitors targeting endoplasmic reticulum stress in Alzheimer’s disease. Front Cell Dev Biol. 2021;9:745011.
76. Zhang J, Wang S, He Y, Yao B, Zhang Y. Regulation of matrix metalloproteinases 2 and 9 in corneal neovascularization. Chem Biol Drug Des. 2020;95:485–492.
77. Sousa GR, Viera GM, Chagas PF, Pezuk JA, Brassesco MS. Should we keep rocking? Portraits from targeting Rho kinases in cancer. Pharmacol Res. 2020;160:105093.
78. Abedi F, Hayes AW, Reiter R, Karima G. Acute lung injury: the therapeutic role of Rho kinase inhibitors. Pharmacol Res. 2020;155:104736.
79. Yang X, Liu Y, Zong Z, Tian D. The Rho kinase inhibitor fasudil inhibits the migratory behavior of 95-D lung carcinoma cells. Biomed Pharmacother. 2010;64:58–62.
80. Wu DJ, Xu JZ, Wu YJ, et al. Effects of fasudil on early atherosclerotic plaque formation and established lesion progression in apolipoprotein E-knockout mice. Atherosclerosis. 2009;207:68–73.
81. Tanaka K, Minami H, Kota M, Kuwamura K, Kohmura E. Treatment of cerebral vasospasm with intra-arterial fasudil hydrochloride. Neurosurgery. 2005;56:214–223.
82. Li Y, Zhu W, Tao J, et al. Fasudil protects the heart against ischemia-reperfusion injury by attenuating endoplasmic reticulum stress and modulating SERCA activity: the differential role for PI3K/Akt and JAK2/STAT3 signalling pathways. PLoS One. 2012;7:e48115.
83. Kawanami D, Matoba K, Okada R, et al. Fasudil inbibits ER stress-induced VCAM-1 expression by modulating unfolded protein in endothelial cells. Biochem Biophys Res Commun. 2013;435:171–175.
84. Wang J, Xu J, Zhao X, Xie W, Wang H, Kong H. Fasudil inhibits neutrophil-endothelial cell interfactions by regulating the expressions of GRP78 and BMPR2. Exp Cell Res. 2018;365:97–105.
85. Chiang AN, Liang M, Dominguez-Meijide A, et al. Synthesis and evaluation of esterified Hsp70 agonists in cellular models of protein aggregation and folding. Bioorg Med Chem. 2019;27(1):79–91.
86. Wisen S, Bertelsen EB, Thompson AD, et al. Binding of a small molecule at a protein-protein interface regulates the chaperone activity of hsp70-hsp40. ACS Chem Biol. 2010;5(6):611–622.
87. Kilpatrick K, Novoa JA, Hancock T, et al. Chemical induction of Hsp70 reduces alpha-synuclein aggregation in neuroglioma cells. ACS Chem Biol. 2013;8(7):1460–1468.
88. Hasegawa Y, Motoyama M, Hamamoto A, et al. Identification of novel oxindole compounds that suppress ER Stress-induced cell death as chemical chaperones. ACS Chem Neurosci. 2022;13(7):1055–1064.
89. Tribouillard-Tranvier D, Béringue V, Desban N, et al. Antihypersensitive drug guanabenz is active in vivo against both yeast and mammalian prions. PLoS One. 2008;23:e1981.
90. Riccio G, D’Avino C, Raines RT, Lorenzo CD. A novel fully human antitumor immunoRNase resistant to the RNase inhibitor. Protein Eng Des Sel. 2013;26:243–248.
91. Colombano G, Caldwell JJ, Matthews TP, et al. Binding to an unusual kinase conformation by highly selective inhibitors of innositol-requiring enzyme 1α kinase-endoribonuclease. J Med Chem. 2019;62:2447–2465.
92. Zhang K, Kaufman RJ. From endoplasmic-reticulum stress to the inflammatory response. Nature. 2008;454(7203):455–462. doi:10.1038/nature07203
93. Pasqualetto G, Pileggi E, Schepelmann M, et al. Ligand-based rational design, synthesis and evaluation of novel potential chemical chaperones for opsin. Eur J Med Chem. 2021;226:113841.
94. Compain P, Martin OR, Boucheron C, et al. Design and synthesis of highly potent and selective pharmacological chaperones for the treatment of Gaucher’s disease. Chembiochem. 2006;7:1356–1359.
95. Fukuda H, Karaki F, Dodo K, et al. Phenanthridin-6-one derivatives as the first class of non-steroidal pharmacological chaperones for Niemann-Pick disease type C1 protein. Bioorg Med Chem Lett. 2017;27(12):2781–2787.
96. Wang JZ, Shimadate Y, Kise M, et al. trans, trans-2-C-Aryl-3,4-dihydroxypyrrolidines as potent and selective β-glucosidase inhibitors: pharmacological chaperones for Gaucher disease. Eur J Med Chem. 2022;238:114499.
97. Schröder SP, Wu L, Artola M, et al. Gluco-1H-imidazole: a new class of azole-type β-glucosidase inhibitor. J Am Chem Soc. 2018;140(15):5045–5048.
98. Cheng WC, Wang JH, Yun WY, et al. Rapid preparation of (3R,4S,5R) polyhydroxylated pyrrolidine-based libraries to discover a pharmacological chaperone for treatment of Fabry disease. Eur J Med Chem. 2017;126:1–6.
99. González-Cuesta M, Herrera-González I, García-Moreno MI, et al. sp2-Iminosugars targeting human lysosomal β-hexosaminidase as pharmacological chaperone candidates for late-onset Tay-Sachs disease. J Enzyme Inhib Med Chem. 2022;37(1):1364–1374. doi:10.1080/14756366.2022.2073444
100. Axten JM, Medina JR, Feng Y, et al. Discovery of 7-methyl-5-(1-{[3-(trifluoromethyl)phenyl]acetyl}-2,3-dihydro-1H-indol-5-yl)-7H-pyrrolo[2,3-d]pyrimidin-4-amine (GSK2606414), a potent and selective first-in-class inhibitor of protein kinase R (PKR)-like endoplasmic reticulum kinase (PERK). J Med Chem. 2012;55(16):7193–7207. doi:10.1021/jm300713s
101. Smith AL, Andrews KL, Beckmann H, et al. Discovery of 1H-pyrazol-3(2H)-ones as potent and selective inhibitors of protein kinase R-like endoplasmic reticulum kinase (PERK). J Med Chem. 2015;58(3):1426–1441. doi:10.1021/jm5017494
102. Raines LN, Zhao H, Wang Y, et al. PERK is a critical metabolic hub for immunosuppressive function in macrophages. Nat Immunol. 2022;23(3):431–445. doi:10.1038/s41590-022-01145-x
103. Zhang D, Liu Y, Zhu Y, et al. A non-canonical cGAS-STING-PERK pathway facilitates the translational program critical for senescence and organ fibrosis. Nat Cell Biol. 2022;24(5):766–782. doi:10.1038/s41556-022-00894-z
 © 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.