Back to Journals » Infection and Drug Resistance » Volume 12
Characterization of the plasmid of incompatibility groups IncFIIpKF727591 and IncpKPHS1 from Enterobacteriaceae species
Authors Wang S, Dai E, Jiang X, Zeng L, Cheng Q, Jing Y, Hu L, Yin Z, Gao B, Wang J, Duan G, Cai X, Zhou D
Received 15 April 2019
Accepted for publication 8 August 2019
Published 6 September 2019 Volume 2019:12 Pages 2789—2797
DOI https://doi.org/10.2147/IDR.S212321
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Professor Suresh Antony
Shujie Wang1,*, Erhei Dai2,*, Xiaoyuan Jiang3, Lijun Zeng3, Qiaoxiang Cheng2,3, Ying Jing3, Lingfei Hu3, Zhe Yin3, Bo Gao3, Jinglin Wang3, Guixin Duan4, Xuehui Cai1, Dongsheng Zhou3
1State Key Laboratory of Veterinary Biotechnology, Harbin Veterinary Research Institute, Chinese Academy of Agricultural Sciences, Harbin 150069, People’s Republic of China; 2Department of Laboratory Medicine, The Fifth Hospital of Shijiazhuang, Hebei Medical University, Shijiazhuang, Hebei 050021, People’s Republic of China; 3State Key Laboratory of Pathogen and Biosecurity, Beijing Institute of Microbiology and Epidemiology, Beijing 100071, People’s Republic of China; 4Animal Science and Technology College, Heilongjiang Bayi Agricultural University, Daqing 163000, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Dongsheng Zhou
State Key Laboratory of Pathogen and Biosecurity, Beijing Institute of Microbiology and Epidemiology, No. 20 Dongdajie Street, Fengtai District, Beijing 100071, People’s Republic of China
Tel +86 106 694 8503
Email [email protected]
Xuehui Cai
State Key Laboratory of Veterinary Biotechnology, Harbin Veterinary Research Institute, Chinese Academy of Agricultural Sciences, No. 678 Haping Road, Xiangfang District, Harbin 150069, People’s Republic of China
Tel +86 4 515 105 1766
Email [email protected]
Background: Multiple incompatibility (Inc) groups of plasmids have been identified in Enterobacteriaceae species, but there are still quite a few sequenced plasmids that could not be assigned to any known Inc groups.
Methods: One IncFIIpKF727591β plasmid p205880-qnrS and two IncpKPHS1 plasmids p11219-CTXM and p205880-NR1 were fully sequenced in this work. Detailed genomic comparison was applied to all available sequenced plasmids of IncFIIpKF727591 or IncpKPHS1 group.
Results: p205880-qnrS carried a novel transposon Tn6396, which was an ISKpn19-compsite transposon and represented a prototype transposable element carrying a minimum core qnrS1 module. p11219-CTXM harbored a novel transposon Tn6559, which was generated from integration of a truncated IS903D–blaCTX-M-14–ISEcp1 unit into the Tn3-family cryptic unit transposon Tn1722. Two Inc groups, IncFIIpKF727591 and IncpKPHS1, of plasmids from Enterobacteriaceae species were proposed, and IncFIIpKF727591 was further grouped into two subgroups IncFIIpKF727591α and IncFIIpKF727591β. Each of the 11 IncFIIpKF727591 plasmids carried multiple accessory modules including at least one resistance module, and the relatively small IncFIIpKF727591 backbones could acquire a wealth of foreign genetic contents. The modular structures of plasmid backbones were conserved within each of IncFIIpKF727591α and IncFIIpKF727591β subgroups but dramatically different, although with similar gene organizations, between these two subgroups. The IncpKPHS1 backbones were conserved with respect to modular structures, and only four of the 14 IncpKPHS1 plasmids carried accessory modules, two of which contained resistance genes.
Conclusion: A genomic comparison of sequenced IncpKPHS1 or IncFIIpKF727591 plasmids provides insights into modular differences and genetic diversification of these plasmids, some of which carries antimicrobial resistance genes.
Keywords: plasmids, IncFIIpKF727591, IncpKPHS1, Tn6396, Tn6559
Introduction
Plasmid is a small DNA molecule within a bacterial cell and capable of replicating independently from the host’s chromosomal DNA. Plasmids are mobile genetic elements that commonly carry antimicrobial resistance genes and other genetic factors such as virulence genes. Plasmid-mediated transmission of antimicrobial resistance genes among Enterobacteriaceae and other bacteria imposes a major public health concern.
The original replicon-based scheme to classify plasmids into different incompatibility (Inc) groups was developed in 1970s, which is based on the experimental observations that plasmids with similar replication machinery are often unable to stably co-exist within the same host cell and thus the plasmid shows incompatibility with the same Inc group plasmid.1 Nowadays, Inc classification is always based on replication initiation protein (Rep) sequences, and it is not necessarily confirmed by conventional conjugation-based incompatibility experiments.1 At least 27 Inc groups have been identified in Enterobacteriaceae species,1 but there are quite a few sequenced plasmids that could not be assigned to any known Inc groups.
This study presented three sequenced plasmids (p205880-qnrS carrying a novel ISKpn19-compsite transposon Tn6396, p11219-CTXM harboring a novel Tn1722-derivated unit transposon Tn6559, and p205880-NR1 containing no resistance genes) and proposed two novel Inc groups (IncFIIpKF727591 and IncpKPHS1). p205880-qnrS belonged to IncpKPHS1, while p11219-CTXM and p205880-NR1 could be assigned to IncFIIpKF727591. Further detailed genomic comparison of all sequenced plasmids of IncpKPHS1 or IncFIIpKF727591 indicated considerable modular differences and genetic diversification of each group of plasmids.
Materials and methods
Bacterial strains and genome sequencing
Klebsiella pneumoniae 205880 and 11219 were recovered from the sputum specimens of two different patients with pneumonia in two different Chinese hospitals in 2012 and 2013, respectively. For each strain, genomic DNA isolation, genome sequencing, and sequence assembly and annotation were carried out as described previously.2 An unrooted neighbor-joining tree was generated from the aligned repA sequences of indicative plasmids.2 Plasmids p205880-qnrS, p11219-CTXM and p205880-NR1 had GenBank accession numbers MF190368, MF133442 and MF144193, respectively.
Phenotypic assays
Plasmid conjugal transfer was carried out, as described previously,2 with Escherichia coli EC600 as a recipient and the 205880 or 11219 isolates as a donor, for selecting an E. coli transconjugant that carried blaCTX-M-14 (p11219-CTXM) or qnrS1 (p205880-qnrS), respectively. Electroporation of plasmid p11219-CTXM from the 11219 isolate into E. coli TOP10 was performed, as described previously,2 to obtain an E. coli electroporant carrying blaCTX-M-14 (p11219-CTXM). Double-disk synergy test was performed to detect the activity of extended-spectrum β-lactamase (ESBL) in indicative bacterial strains.3 BioMérieux VITEK 2 was used to test bacterial antimicrobial susceptibility, which was interpreted as per the Clinical and Laboratory Standards Institute (CLSI) guidelines.4
Results and discussion
Diversification of IncFIIpKF727591 plasmids
One new plasmid p205880-qnrS was fully sequenced (Table 1) and could be transferred from the wild-type 205880 isolate into EC600, through conjugation, giving a qnrS-positive transconjugant p205880-qnrS-EC600. As expected, these two strains were resistant to ciprofloxacin and levofloxacin with minimum inhibitory concentration (MIC) values ≥4.
 |
Table 1 Major features of plasmids analyzed |
A collection of 11 plasmids including p205880-qnrS (Table S1), which had homologous repA (replication initiation) genes and similar backbone gene organizations, were assigned into a novel Inc group designated IncFIIpKF727591 (Increference plasmid), because all these RepA proteins had an IncFII super-family domain. The phylogenetic tree (Figure 1) based on repA sequences indicated that these 11 plasmids could be divided into two separately clustering subgroups IncFIIpKF727591α (n=8) and IncFIIpKF727591β (n=3). As shown by pairwise comparison of repA nucleotide sequences, plasmids within each subgroup showed 100% identity, while those from different subgroups displayed ≥79% identity (Table S2A). Predicted RepA-binding iterons were located from 245 bp to 365 bp downstream of repA for IncFIIpKF727591α plasmids, but upstream from 366 bp to 460 bp for IncFIIpKF727591β plasmids, and three copy numbers of iteron were found for all IncFIIpKF727591 plasmids (Table S1). Plasmids within each subgroup shared a conserved iteron motif, but those from different subgroups had dramatically different iteron motifs (Figure 1).
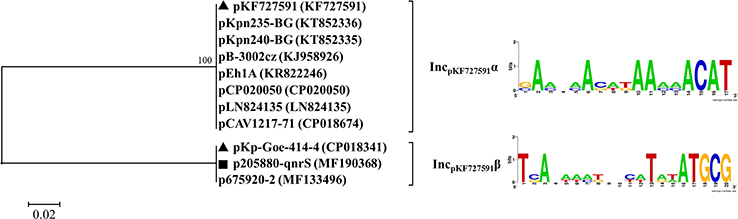 |
Figure 1 A neighbor-joining phylogenetic tree for IncFIIpKF727591 plasmids. The degree of support (percentage) for both cluster of associated taxa, as determined by bootstrap analysis, is shown next to each branch. The bar corresponds to the scale of sequence divergence. The triangles indicate the reference plasmids, while the square denotes the plasmid sequenced in this study. |
pKF727591 (the first sequenced IncFIIpKF727591 plasmid) and pKp_Goe_414-4 (the first sequenced IncFIIpKF727591β plasmid) were identified as the references for IncFIIpKF727591α and IncFIIpKF727591β, respectively. p205880-qnrS belonged to IncFIIpKF727591β.
The modular structure (Table 1 and Figure S1) of each plasmid could be divided into one or more accessory modules (defined as acquired DNA regions associated or bordered with mobile elements) and the remaining IncFIIpKF727591 backbone regions (responsible for plasmid replication, maintenance and conjugal transfer). The eight IncFIIpKF727591α plasmids shared ≥88% of their backbone sequences with ≥99% nucleotide identity, and the three IncFIIpKF727591β plasmids showed ≥99% nucleotide identity over ≥98% of their backbone sequences; by contrast, the backbones of IncFIIpKF727591α and IncFIIpKF727591β had ≤92% nucleotide identity across ≤70% of their backbone sequences (Table S2B). The modular structures of plasmid backbones were conserved within each of IncFIIpKF727591α and IncFIIpKF727591β subgroups but dramatically different between two subgroups.
Integration of accessory modules at various sites of IncFIIpKF727591 backbones led to the interruption of relevant backbone genes (eg, umuC), the disruption of the maintenance or conjugal transfer regions, or the deletion of surrounding backbone regions (eg, 5.3-kb deletion containing mtsM) (Figure 2). The IncFIIpKF727591 replicons and the conjugal transfer regions (encoding an F-type type IV secretion system) were found in all 11 plasmids and thus represented the core IncFIIpKF727591 backbone. An 11-kb maintenance region carrying parAB (partition) and ccdBA (toxin-antitoxin) was found in all IncFIIpKF727591α plasmids, while another distinct 15-kb maintenance region containing stbAB (mediator of plasmid stability) and resD (resolvase) in all IncFIIpKF727591β plasmids.
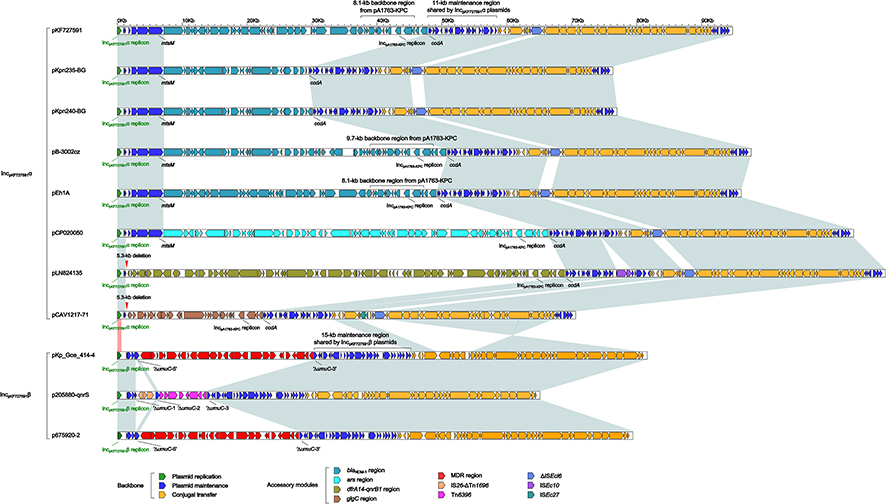 |
Figure 2 Linear comparison of complete sequences of IncFIIpKF727591 plasmids. Genes are denoted by arrows. Genes, mobile elements and other features are colored based on function classification. Shading regions denote homology of plasmid backbone regions (light blue: ≥90% nucleotide identity; light red: <90% nucleotide identity) but not accessory modules. |
All the three IncFIIpKF727591β plasmids contained a single IncFIIpKF727591β replicon, which six of the eight IncFIIpKF727591α plasmids contained a second IncpA1763-KPC replicon beside the master IncFIIpKF727591α replicon (Figure 2). Notably, the IncpA1763-KPC replicon was located within a 9.7- or 8.1-kb backbone region [carrying maintenance genes such as parA and resA; as observed in pA1763-KPC (GenBank accession number MH909340], which was a part of relevant accessory modules (see below; Figure 3). Two coexistent replicons IncFIIpKF727591α and IncpA1763-KPC, together with their supporting maintenance genes, will promote relevant plasmids to overcome incompatibility barrier with incoming plasmids. All the above replicons belonged to the iteron-regulated replicon, for which Rep monomers specifically bound to iterons.5
 |
Figure 3 Organization of selected accessory modules from IncFIIpKF727591α plasmids and comparison with related regions. Genes are denoted by arrows. Genes, mobile elements and other features are colored based on function classification. Shading denotes regions of homology (>95% nucleotide identity). Numbers in brackets indicate e nucleotide positions within corresponding plasmids. The accession numbers of Tn125,6 Tn625617 and Tn133118 for reference are JN872328, KP851978 and KC354802, respectively. |
Accessory resistance modules of IncFIIpKF727591 plasmids
A large accessory module was integrated at a site between the two maintenance genes mtsM and ccdA in each of the eight IncFIIpKF727591α plasmids (Figure 3). These eight modules had some common regions but showed considerable modular differences across the whole modules, indicating their sole evolutionary origin followed by parallel mosaic diversification. The 9.7- or 8.1-kb backbone region from IncpA1763-KPC (see above) was found in six IncFIIpKF727591α plasmids except for pKpn235-BG and pKpn240-BG.The accessory modules from seven IncFIIpKF727591α plasmids, except for pCAV1217-71, carried resistance loci (Figure 3 and Table S3): i) a truncated Tn125 transposon carrying blaNDM−16 was harbored in pB-3002cz, pKpn235-BG and pKpn240-BG, while another truncated version of Tn125 in pKF727591 and pEh1A; ii) the ars (arsenical resistance) locus was found in pKF727591, pEh1A and pCP020050 and, notably, the first two plasmids showed coexistence of blaNDM−1 and ars; and iii) qnrB1 and dfrA14-carrying In191 were identified in pLN824135.
A 24.7-kb MDR region, another 26.9-kb MDR region and Tn6396 (Figure 4) were inserted at the same site within the umuC gene of the three IncFIIpKF727591β plasmids p675920-2, pKp_Goe_414-4 and p205880-qnrS, respectively. The 24.7-kb MDR region, carrying multiple resistance genes (Table S3), was generated from integration of an IS26–∆Tn6346–∆GIsul2–IS26 unit7 into Tn1721,8 which was further connected with a truncated IS26–blaLAP-2–qnrS1–IS26 unit.7 The 26.9-kb MDR region was highly similar to the 24.7-kb MDR region but differed from it mainly by inversion of IS26–∆Tn6346–∆GIsul2–IS26 and further upstream insertion of an IS26–pdk–catA2–IS26 unit. Tn6396 was a novel ISKpn19-compsite transposon, which carried the qnrS1–ΔtnpR region and bracketed by 7-bp direct repeats (DRs: target site duplication signals for transposition) at both ends. Tn6396 represented a prototype transposable element carrying a minimum core qnrS1 module. Different Tn6396 derivatives with distinct terminal truncations were found in various plasmids including pKp_Goe_414-4 and p675920-2 (Figure S2).
 |
Figure 4 Organization of accessory resistance modules from IncFIIpKF727591β plasmids and comparison with related regions. Genes are denoted by arrows. Genes, mobile elements and other features are colored based on function classification. Shading denotes regions of homology (>95% nucleotide identity). Numbers in brackets indicate e nucleotide positions within corresponding plasmids. The accession numbers of GIsul2,19 Tn6346,20 Tn1728 and IS26–blaLAP-2–qnrS1–IS26 unit21 for reference are CP001918, EU696790, X61367 and HF545433, respectively. |
Characterization of IncpKPHS1 plasmids
Two additional new plasmids p11219-CTXM (carrying blaCTX-M-14) and p205880-NR1 (containing no resistance genes) were fully sequenced (Table 1 and Figure S1). p11219-CTXM could not be transferred from the wild-type 11219 isolate into EC600 through conjugation, but could be transferred into TOP10 through electroporation, generating a blaCTX-M-14-positive electroporant 11219-CTXM-TOP10. These two wild-type and electroporant strains had ESBL activity (data not shown) and were resistant to cefazolin, cefuroxime and ceftazidime with MIC values ≥64.
A total of 14 plasmids including p11219-CTXM and p205880-NR1 (Table S1), each of which carried a single repA gene with >96% nucleotide identity to repApKPHS1 (Table S4A) and had a backbone gene organization similar to pKPHS1 (Figure 5), were assigned into a novel Inc group named as IncpKPHS1 (Figure S3). These 14 RepA proteins did not any of known domain super-families. Four copy numbers of a conserved iteron motif (Figure S3) were found 48 bp to 218 bp downstream of repA for all IncpKPHS1 plasmids (Table S1). All plasmids carried a single iteron-regulated IncpKPHS1 replicon. pKPHS1, the first sequenced IncpKPHS1 plasmid, was identified as the IncpKPHS1 reference. These 14 plasmids had >96% nucleotide identity over >75% coverage of their backbone sequences (Table S4B). Modular differences were found at multiple sites of the maintenance regions (Figure 5). None of conjugal transfer genes was found in all plasmids, which was consistent to the non-conjugative nature of p11219-CTXM. Remarkably, all these plasmids carried ɸpKPHS1 regions resembling SSU5 phage.9
 |
Figure 5 Linear comparison of complete sequences of IncpKPHS1 plasmids. Genes are denoted by arrows. Genes, mobile elements and other features are colored based on function classification. Shading regions denote homology of plasmid backbone regions (≥90% nucleotide identity) but not accessory modules. |
Only four IncpKPHS1 plasmids had accessory modules, including the two resistance modules: Tn6558 from pKPHS1 and Tn6559 from p11219-CTXM (Table 1 and Figure 5). The highly similar Tn6558 and Tn6559 (Figure 6) were novel Tn3-family unit transposons generated from integration of truncated IS903D–blaCTX-M-14–ISEcp1 units (representing the master prototype blaCTX-M-14 genetic environments in China)10 into the mcp gene of cryptic Tn1722,8 and they slightly differed from one another by distinct truncations occurred within IS903D–blaCTX-M-14–ISEcp1 unit or mcp.
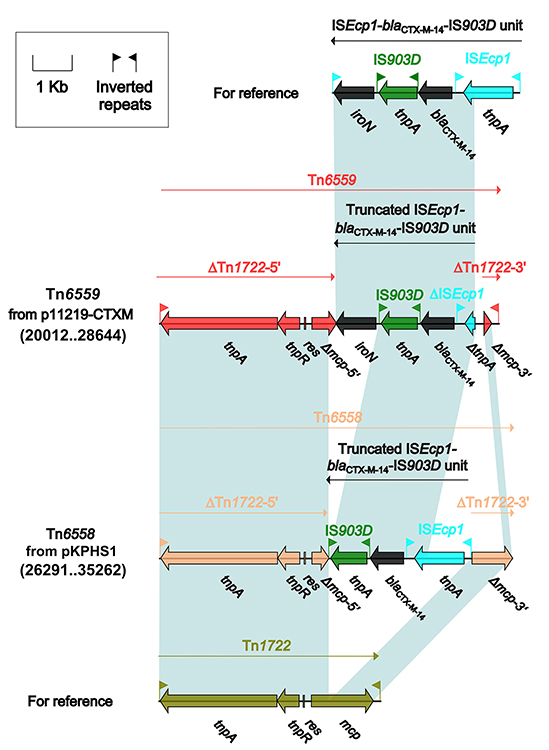 |
Figure 6 Organization of Tn6558 and Tn6559 and comparison with related regions. Genes are denoted by arrows. Genes, mobile elements and other features are colored based on function classification. Shading denotes regions of homology (>95% nucleotide identity). Numbers in brackets indicate the nucleotide positions within the corresponding plasmids. The accession numbers of ISEcp1–blaCTX-M-14–IS903D unit22 and Tn17228 are KX646543 and X61367, respectively. |
Ethics approval and informed consent
This study needs not to be reviewed or approved by the ethics committee of the hospitals, because the bacterial isolate involved in this study was part of the routine hospital laboratory procedure. The research involving biohazards and all related procedures were approved by the Biosafety Committee of the Beijing Institute of Microbiology and Epidemiology.
Acknowledgment
This work was supported by the National Key R&D Program (2018YFC1200100) of China and the Local Social Science Project (2018QD0031) of Heilongjiang Province.
Author contributions
All authors contributed toward data analysis, drafting and revising the paper, gave final approval of the version to be published and agree to be accountable for all aspects of the work.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Shintani M, Sanchez ZK, Kimbara K. Genomics of microbial plasmids: classification and identification based on replication and transfer systems and host taxonomy. Front Microbiol. 2015;6:242. doi:10.3389/fmicb.2015.00242
2. Zhan Z, Hu L, Jiang X, et al. Plasmid and chromosomal integration of four novel blaIMP-carrying transposons from Pseudomonas aeruginosa, Klebsiella pneumoniae and an Enterobacter sp. J Antimicrob Chemother. 2018;73(11):3005–3015. doi:10.1093/jac/dky288
3. CLSI. Performance Standards for Antimicrobial Susceptibility Testing: Twenty-Fifth Informational Supplement M100-S25. Wayne (PA): Clinical and Laboratory Standards Institute; 2015.
4. Wayne. Performance Standards for Antimicrobial Susceptibility Testing: Twenty-seventh Informational Supplement M100-S27. CLSI; 2017.
5. Pilla G, Tang CM. Going around in circles: virulence plasmids in enteric pathogens. Nat Rev Microbiol. 2018;16(8):484–495. doi:10.1038/s41579-018-0031-2
6. Poirel L, Bonnin RA, Boulanger A, Schrenzel J, Kaase M, Nordmann P. Tn125-related acquisition of blaNDM-like genes in Acinetobacter baumannii. Antimicrob Agents Chemother. 2012;56(2):1087–1089. doi:10.1128/AAC.05620-11
7. Feng J, Yin Z, Zhan Z, et al. Structure genomics of two chimera plasmids p675920-1 and p675920-2 coexisting in a multi-drug resistant Klebsiella pneumoniae isolate. Oncotarget. 2018. doi:10.18632/oncotarget.24235
8. Allmeier H, Cresnar B, Greck M, Schmitt R. Complete nucleotide sequence of Tn1721: gene organization and a novel gene product with features of a chemotaxis protein. Gene. 1992;111(1):11–20. doi:10.1016/0378-1119(92)90597-i
9. Kim M, Kim S, Ryu S. Complete genome sequence of bacteriophage SSU5 specific for Salmonella enterica serovar Typhimurium rough strains. J Virol. 2012;86(19):10894. doi:10.1128/JVI.01796-12
10. Poirel L, Lartigue MF, Decousser JW, Nordmann P. ISEcp1B-mediated transposition of blaCTX-M in Escherichia coli. Antimicrob Agents Chemother. 2005;49(1):447–450. doi:10.1128/AAC.49.1.447-450.2005
11. Studentova V, Dobiasova H, Hedlova D, Dolejska M, Papagiannitsis CC, Hrabak J. Complete nucleotide sequences of two NDM-1-encoding plasmids from the same sequence type 11 Klebsiella pneumoniae strain. Antimicrob Agents Chemother. 2015;59(2):1325–1328. doi:10.1128/AAC.04095-14
12. Papagiannitsis CC, Malli E, Florou Z, et al. Emergence of sequence type 11 Klebsiella pneumoniae coproducing NDM-1 and VIM-1 metallo-beta-lactamases in a Greek hospital. Diagn Microbiol Infect Dis. 2017;87(3):295–297. doi:10.1016/j.diagmicrobio.2016.12.008
13. Campos JC, Da Silva MJ, Dos Santos PR, et al. Characterization of Tn3000, a transposon responsible for blaNDM-1 dissemination among enterobacteriaceae in Brazil, Nepal, Morocco, and India. Antimicrob Agents Chemother. 2015;59(12):7387–7395. doi:10.1128/AAC.01458-15
14. Liu P, Li P, Jiang X, et al. Complete genome sequence of Klebsiella pneumoniae subsp. pneumoniae HS11286, a multidrug-resistant strain isolated from human sputum. J Bacteriol. 2012;194(7):1841–1842. doi:10.1128/JB.00043-12
15. Stoesser N, Giess A, Batty EM, et al. Genome sequencing of an extended series of NDM-producing Klebsiella pneumoniae isolates from neonatal infections in a Nepali hospital characterizes the extent of community- versus hospital-associated transmission in an endemic setting. Antimicrob Agents Chemother. 2014;58(12):7347–7357. doi:10.1128/AAC.03900-14
16. Khong WX, Marimuthu K, Teo J, et al. Tracking inter-institutional spread of NDM and identification of a novel NDM-positive plasmid, pSg1-NDM, using next-generation sequencing approaches. J Antimicrob Chemother. 2016;71(11):3081–3089. doi:10.1093/jac/dkw277
17. Antonelli A, D’Andrea MM, Vaggelli G, Docquier JD, Rossolini GM. OXA-372, a novel carbapenem-hydrolysing class D beta-lactamase from a Citrobacter freundii isolated from a hospital wastewater plant. J Antimicrob Chemother. 2015;70(10):2749–2756. doi:10.1093/jac/dkv181
18. Espedido BA, Steen JA, Ziochos H, et al. Whole genome sequence analysis of the first Australian OXA-48-producing outbreak-associated Klebsiella pneumoniae isolates: the resistome and in vivo evolution. PLoS One. 2013;8(3):e59920. doi:10.1371/journal.pone.0059920
19. Nigro SJ, Hall RM. GIsul2, a genomic island carrying the sul2 sulphonamide resistance gene and the small mobile element CR2 found in the Enterobacter cloacae subspecies cloacae type strain ATCC 13047 from 1890, Shigella flexneri ATCC 700930 from 1954 and Acinetobacter baumannii ATCC 17978 from 1951. J Antimicrob Chemother. 2011;66(9):2175–2176. doi:10.1093/jac/dkr230
20. Ng SP, Davis B, Palombo EA, Bhave M. A Tn5051-like mer-containing transposon identified in a heavy metal tolerant strain Achromobacter sp. AO22. BMC Res Notes. 2009;2:38. doi:10.1186/1756-0500-2-38
21. Le V, Nhu NT, Cerdeno-Tarraga A, et al. Genetic characterization of three qnrS1-harbouring multidrug-resistance plasmids and qnrS1-containing transposons circulating in Ho Chi Minh City, Vietnam. J Med Microbiol. 2015;64(8):869–878. doi:10.1099/jmm.0.000100
22. Wang L, Liu L, Liu D, et al. The first report of a fully sequenced resistance plasmid from Shigella boydii. Front Microbiol. 2016;7:1579. doi:10.3389/fmicb.2016.01579
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.