Back to Journals » Infection and Drug Resistance » Volume 12
Characterization of NDM-5- and CTX-M-55-coproducing Escherichia coli GSH8M-2 isolated from the effluent of a wastewater treatment plant in Tokyo Bay
Authors Sekizuka T ![]() , Inamine Y, Segawa T
, Inamine Y, Segawa T ![]() , Kuroda M
, Kuroda M ![]()
Received 10 May 2019
Accepted for publication 17 June 2019
Published 23 July 2019 Volume 2019:12 Pages 2243—2249
DOI https://doi.org/10.2147/IDR.S215273
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Dr Joachim Wink
Tsuyoshi Sekizuka, Yuba Inamine, Takaya Segawa, Makoto Kuroda
Laboratory of Bacterial Genomics, Pathogen Genomics Center, National Institute of Infectious Diseases, Shinjuku, Tokyo, Japan
Purpose: New Delhi metallo-β-lactamase (NDM)-5-producing Enterobacteriaceae have been detected in rivers, sewage, and effluents from wastewater treatment plants (WWTPs). Environmental contamination due to discharged effluents is of particular concern as NDM variants may be released into waterways, thereby posing a risk to humans. In this study, we collected effluent samples from a WWTP discharged into a canal in Tokyo Bay, Japan.
Methods: Testing included the complete genome sequencing of Escherichia coli GSH8M-2 isolated from the effluent as well as a gene network analysis.
Results: The complete genome sequencing of GSH8M-2 revealed that it was an NDM-5-producing E. coli strain sequence type ST542, which carries multiple antimicrobial resistance genes for β-lactams, quinolone, tetracycline, trimethoprim-sulfamethoxazole, florfenicol/chloramphenicol, kanamycin, and fosfomycin. The blaNDM-5 gene was found in the IncX3 replicon plasmid pGSH8M-2-4. Gene network analysis using 142 IncX3 plasmid sequences suggested that pGSH8M-2-4 is related to both clinical isolates of E. coli and Klebsiella species in Eastern Asia. GSH8M-2 also carries the blaCTX-M-55 gene in IncX1 plasmid pGSH8M-2-3.
Conclusion: This is the first report of environmental NDM-5-producing E. coli isolated from a WWTP in Japan. NDM-5 detection is markedly increasing in veterinary and clinical settings, suggesting that dual β-lactamases, such as NDM-5 and CTX-M-55, might be acquired through multiple steps in environment settings. Environmental contamination through WWTP effluents that contain producers of NDM variants could be an emerging potential health hazard. Thus, regular monitoring of WWTP effluents is important for the detection of antimicrobial-resistant bacteria that may be released into the waterways and nearby communities.
Keywords: sewage, carbapenemase, NDM-5, IncX3, CTX-M-55, IncX1
Introduction
Antimicrobial resistance (AMR) is of global concern as the widespread detection of carbapenemase-producing Enterobacteriaceae (CPE) is an emerging issue with potentially serious public health implications.1 New Delhi metallo-β-lactamase (NDM) confers resistance to β-lactams with the exception of aztreonam, and the rapid global spread of NDM-1-producing strains has raised serious concerns that common infections with these organisms may soon be untreatable.2 Variants of NDM carbapenemase have been assigned to blaNDM-27 under curation by the National Database of Antibiotic Resistant Organisms (NDARO, https://www.ncbi.nlm.nih.gov/pathogens/isolates#/refgene/blaNDM). NDM-5 was first reported in 2011 in the United Kingdom and was isolated from a multidrug-resistant Escherichia coli ST648 clinical isolate from a patient with a recent history of hospitalization in India.3
NDM-5 exhibits expanded-spectrum cephalosporins and carbapenems degradation with two amino acid substitutions (Val88Leu and Met154Leu) in the NDM-1 amino acid sequence.3 NDM-5 has been characterized as exhibiting higher hydrolytic activity than other variants (NDM-1, NDM-4, NDM-6, and NDM-7).4
NDM-5-producing Enterobacteriaceae have been identified from many different sources in multiple countries. Thus far, it has been reported that blaNDM-5 appears to be predominant among all potential carbapenemase genes (including other NDM variants and KPC-2) in China.5 Moreover, diverse dissemination of the blaNDM-5 gene has been detected among non-clonal E. coli hosts (ST12, ST167, ST354, ST361, ST410, ST617, ST746, ST6335, and ST6395) via the IncX3 plasmid in China.6,7 To elucidate the dissemination of CPE including NDM variants, AMR monitoring of wastewater treatment plant (WWTP) effluent contributes to the detection of ongoing AMR bacterial dissemination in environment, and might provide early warning of potential dissemination in clinical settings and communities.
Here, we isolated an NDM-5-producing E. coli strain GSH8M-2 from an effluent sampled at a WWTP in Tokyo Bay, Japan. Then, we determined the complete genome sequence of GSH8M-2, and characterized the genetic features of blaNDM-5- and blaCTX-M-55-positive plasmids.
Materials and methods
Bacterial strains and identification
The upper effluent flow from an urban WWTP was collected on August 6, 2018, in the canal (N35.631783, E139.7448151) at Tokyo Bay, Japan. The facilities of the WWTP process an influent by biological reactors and disinfection with chlorination. Average water qualities of pH, biochemical oxygen demand, and chemical oxygen demand are pH 6.8, 16, and 12 mg/L, respectively. The treated effluent sample in this study was collected within five consecutive days without recent rainfall to exclude the effect of the weather. Then, bacteria within the effluent sample were isolated as described previously for CPE.8 Briefly, 500 mL of effluent was filtered, and the membrane incubated in 20 mL of LB-broth supplemented with 1 mg/L meropenem at 37°C for 14 hrs, after which the culture was spread onto CHROMagar extended-spectrum β-lactamase (ESBL) plates (CHROMagar, Paris, France).
Antimicrobial susceptibility and CPE screening tests
Antimicrobial susceptibility was investigated by E test (bioMérieux, Marcy-l’Étoile, France) according to the European Committee on Antimicrobial Susceptibility Testing guideline v9.0. Carbapenemase production was assessed by performing a Carba NP test, as described previously.9
Whole-genome sequence analysis
Determination of the complete genome sequence was performed as described, previously.10 Briefly, the complete genome sequence was determined using an Illumina NextSeq 500 (2×150-mer, 350× median coverage) (Illumina, San Diego, CA, USA), followed by a PacBio Sequel sequencer for long-read sequencing (PacBio, Menlo Park, CA, USA). A de novo assembly was performed.10 Annotation was conducted in DFAST.11 Multilocus Sequence Typing was characterized at the pubMLST site (https://pubmlst.org/escherichia/). Circular representations of complete genome sequences were visualized using the GView server.12 AMR genes were identified by homology searching against the ResFinder database.13 Plasmid replicon typing was performed using the curated PlasmidFinder database at the CGE website (https://cge.cbs.dtu.dk//services/PlasmidFinder/).14
Comparative plasmid genome network analysis
Complete plasmid sequences were retrieved from NCBI “nt” and “RefSeq” database using a keyword search for the words “complete sequence” and “plasmid”, followed by gene prediction using the Prodigal program for normalization of gene finding. A plasmid network analysis was performed as previously described.15 In brief, amino acid sequences for the complete plasmids were clustered using the UCLUST program (version 6.0.307)16 with the following parameters: cluster_smallmem; id 1.0 (100% identity in amino acid sequence); target_cov 0.9, query_cov 0.9 (at least 90% coverage); minsl 0.9, minqt 0.9, maxqt 1.1 (less than 10% difference in length). Each plasmid, ranging from 30 to 60 kb in size, was connected to a network under the sharing threshold with at least 40 homologous genes, followed by community assignment using the multilevel community method in the R igraph library. In order to visualize the level of similarity for individual plasmid sequences in a dataset, non-metric multidimensional scaling analysis was performed with the distance matrix calculated with the Simpson’s index using the metaMDS program from Vegan R package. The Cytoscape version 3.2.0 was used to draw the plasmid network graph.17
Results
NDM-5-producing E. coli strain GSH8M-2
The isolated E. coli effluent strain GSH8M-2 was Carba NP test positive. MIC testing suggested that GSH8M-2 was highly resistant to the β-lactams, fluoroquinolone, trimethoprim-sulfamethoxazole, and chloramphenicol, kanamycin, and fosfomycin. However, it was still susceptible to colistin and aminoglycosides (amikacin, gentamicin, streptomycin, and tobramycin) except kanamycin (Table 1).
 |
Table 1 Antimicrobial susceptibility test in E. coli GSH8M-2 |
Genome features of E. coli strain GSH8M-2
The complete genome sequence was determined, as described in the materials and methods. GSH8M-2 carries multiple antimicrobial resistance genes (ARGs) for β-lactams (blaNDM-5, blaCTX-M-55, blaEC, and blaTEM-135), quinolone (qnrS1 and qnrS2), tetracycline (tet (A)), trimethoprim-sulfamethoxazole (dfrA14 and dfrA17), florfenicol/chloramphenicol (floR), kanamycin (aph (3ʹ)-Ia), and fosfomycin (fosA3).
Importantly, IncX3 plasmid pGSH8M-2–4 (46.1 kb) carries blaNDM-5 (Figure 1A), exhibiting an almost identical gene organization to pNDM-MGR194, which was first identified in IncX3 plasmids carrying blaNDM-5 in Klebsiella pneumoniae MGR-194 clinical isolates in India18 (Figure 1B). These two blaNDM-5-positive IncX3 plasmids showed partially distinct gene organization around the blaNDM-1 to IncX3 plasmid in p112298-NDM, which was first identified in 2013 in a Citrobacter freundii strain 112298 in China.19
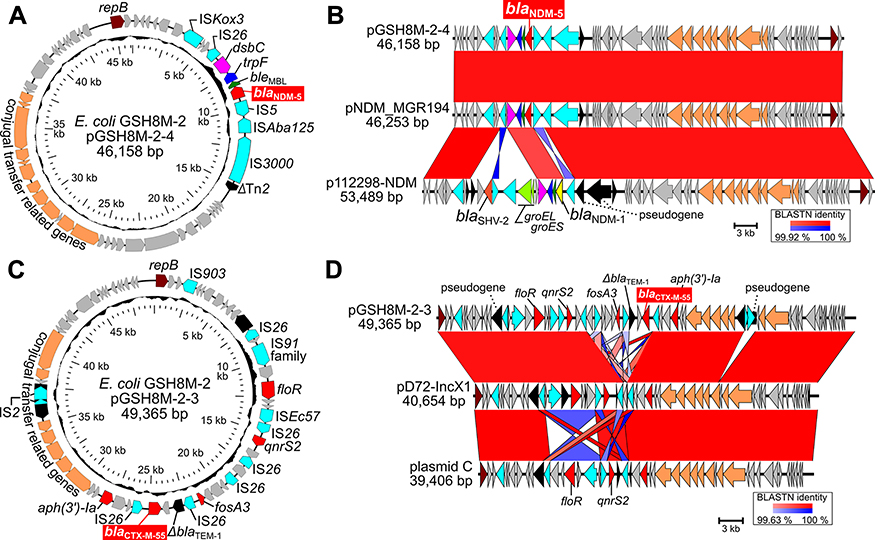 |
Figure 1 Plasmid analysis of pGSH8M-2–4 and pGSH8M-2–3. (A) Circular representation of pGSH8M-2-4. ORFs were painted with color by predicted gene function; blaNDM-5, red; replication, brown; insertion sequences, light blue; conjugal transfer related genes, orange. (B) Structural comparison of the IncX3 plasmids carrying blaNDM-positive plasmids. A pairwise nucleotide % identity is shown as red (forward orientation) and blue (complementary orientation) between plasmid sequences. pNDM-MGR194 (GenBank ID: KF220657) was first identified IncX3 plasmids carrying blaNDM-5 in Klebsiella pneumoniae in India. p112298-NDM (GenBnak ID: KP987216), IncX3 plasmid carrying blaNDM-1 instead of blaNDM-5, was identified in Citrobacter freundii strain 112298 in China in 2013. (C) Circular representation of pGSH8M-2-3. (D) Structural comparison of the IncX1 plasmids similar to pGSH8M-2-3. blaCTX-M-55 was found in pGSH8M-2-3, not but in pD72-IncX1 (GenBank ID: CP035315) in E. coli D72 isolated from pig in China, and plasmid C (GenBank ID: CP010155) in E. coli D9 isolated from dog. |
IncX1 plasmid pGSH8M-2–3 (49.3 kb) carries multiple ARGs including blaCTX-M-55, aph(3ʹ)-Ia, floR, fosA3, and qnrS2 (Figure 1C). Blast homology search revealed that three publicly complete sequences (two plasmids and one chromosome DNA of E. coli strains isolated from a dog and pig) showed homology with >80% region coverage, for all three plasmids sharing the aph(3ʹ)-Ia, floR, and qnrS2 for three primarily veterinary use antimicrobials (aminoglycosides, florfenicol, and quinolone, respectively) (Figure 1D). However, pGSH8M-2–3 carries additional blaCTX-M-55 and fosA3 with IS26 mediated acquisition (Figure 1D).
Plasmid gene network analysis of IncX3 plasmids
To further elucidate the dissemination of blaNDM-5-positive IncX3 plasmids, gene clustering, and network analysis was performed using the IncX3 plasmids relevant to pGSH8M-2–4, resulting in 141 plasmid entries obtained from a public database (Table S1). Those 142 plasmid sequences were classified into four communities (Figure 2). The pGSH8M-2–4 strain originated from a community (C1) that shares blaNDM-5 with E. coli and Klebsiella clinical isolates in Eastern Asia. Of note, the community network analysis strongly suggested that blaNDM-5-positive IncX3 plasmids are of clonal lineage, but horizontally disseminated over multiple bacteria hosts (Escherichia, Klebsiella, Enterobacter, Citrobacter, and Kluyvera). On the other hand, other β-lactamases [blaNDM-1, blaKPC, blaOXA-181, and ESBL gene-positive IncX3 plasmids can be clustered into different communities from each other (C2, C3, and C4 in Figure 2), because of horizontal acquisition with extra adjacent genes around the ARGs (Figure 1B). Thus, each β-lactamase-positive IncX3 plasmid appears to have disseminated in an independent manner among Enterobacteriaceae.
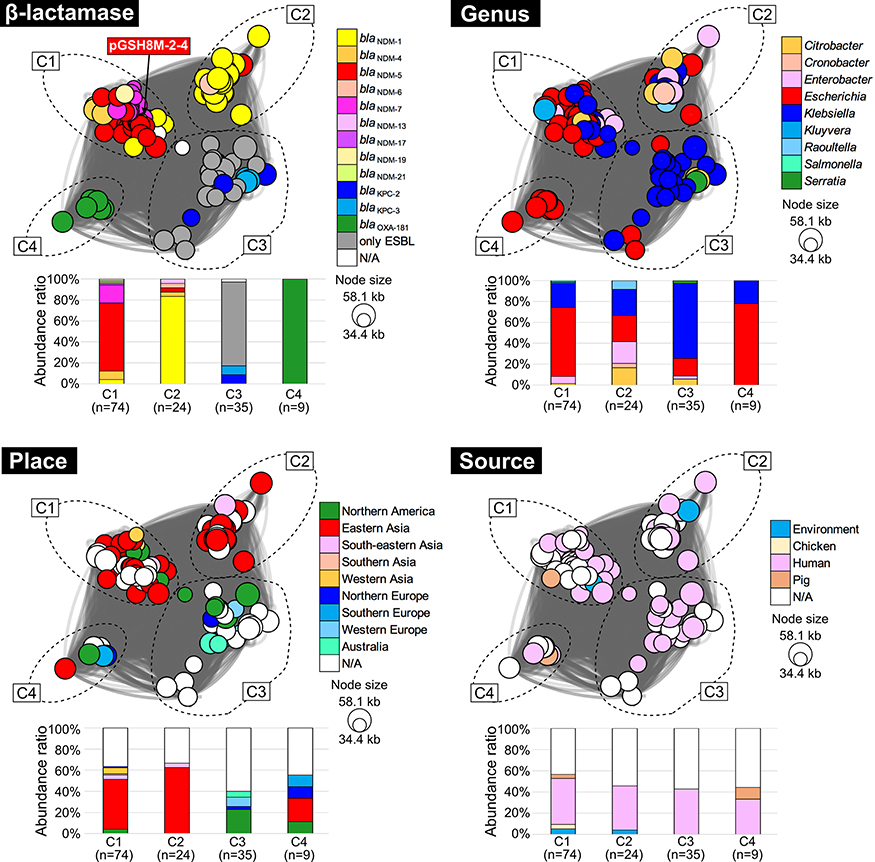 |
Figure 2 Plasmid gene network analysis using pGSH8M-2-4 related sequences. Using pGSH8M-2-4 sequence as query, 141 IncX3 plasmid sequences were obtained from the public database. In total, 142 plasmid sequences (see Table S1), each plasmid can be clustered into four communities (C1–C4). A circle indicates the clustering position of each plasmid, highlighted with the indicated color based on the type of β-lactamase, genus, place, and source of isolation. |
Discussion
In this study, we isolated the NDM-5-producing E. coli strain (GSH8M-2) from effluent of a WWTP in Tokyo Bay, Japan. This is the first report of an environmental NDM-5-producing E. coli isolate from a WWTP in Japan. Thus far, the detection of NDM variants in clinical settings is very rare and uncommon in Japan, a few reports suggested that NDM-5-producing E. coli and K. pneumoniae were identified in a Japanese individual traveling to Bangladesh in 2013 and also in a case in Indonesia in 2014. However, both of these were imported cases to Japan not a local occurrence.20 Thus, our findings in this study may indicate an emerging public health issue within the Japanese community.
In addition, previous environmental studies have suggested that NDM-5-producing Enterobacteriaceae have been detected in an urban river in Montpellier, France,21 hospital sewage water in India22 and Taiwan,23 and effluent from a WWTP in Switzerland.24 Although a few studies revealed dissemination of environmental NDM variants, WWTPs could be a primary source of most AMR bacteria in aquatic environments, because of the partial purification that occurs during the wastewater treatment processes. In particular, ongoing global dissemination could be archived by the rapid spread of the IncX3 replicon-based plasmid transmission among Enterobacteriaceae (Figure 2).
In addition to NDM-5, GSH8M-2 harbors CTX-M-55, which was first reported in 2006 in Thailand25 and is a variant of CTX-M-15 in the CTX-M-1 cluster, with only one amino acid substitution (Ala80Val). Over the past decade, CTX-M-55-producing E. coli isolated from animals and in clinical settings has been increasing.26 Moreover, the blaCTX-M-55-positive plasmid pGSH8M-2-3 is associated with other ARGs (aph(3ʹ)-Ia, floR, fosA3, and qnrS2), the co-localization of the florfenicol resistance gene floR is of marked concern because floR is detected in increasing amounts in both veterinary27 and clinical isolates28 within China. Therefore, CTX-M-55-positive E. coli could disseminate in other settings under highly selective pressure by co-selection of the veterinary antimicrobials (florfenicol) and clinical use (cephalosporin) and has become one of the most important emergent AMR bacteria in environment. These findings suggest that the dissemination of dual β-lactamase (carbapenemase and ESBL)-positive bacteria has become a global issue, and environmental contamination could act as a reservoir of NDM variants-producing AMR Enterobacteriaceae.
Conclusions
AMR monitoring of WWTP effluent may contribute to the detection of emerging AMR bacterial dissemination in clinical settings and communities. A comprehensive approach based on the complete determination of the genome sequence including the AMR plasmid will be required in order to fully understand AMR dissemination of these bacterial clones and relevant AMR plasmids within the environment.
Data availability
The complete, annotated genome sequence of E. coli GSH8M-2 was deposited in a public database as shown in Table 2. The short- and long-read sequences for DNA-Seq were deposited in the DNA Data Bank of Japan (BioProject PRJDB6962; BioSample SAMD00168435; DRA accession DRA008278).
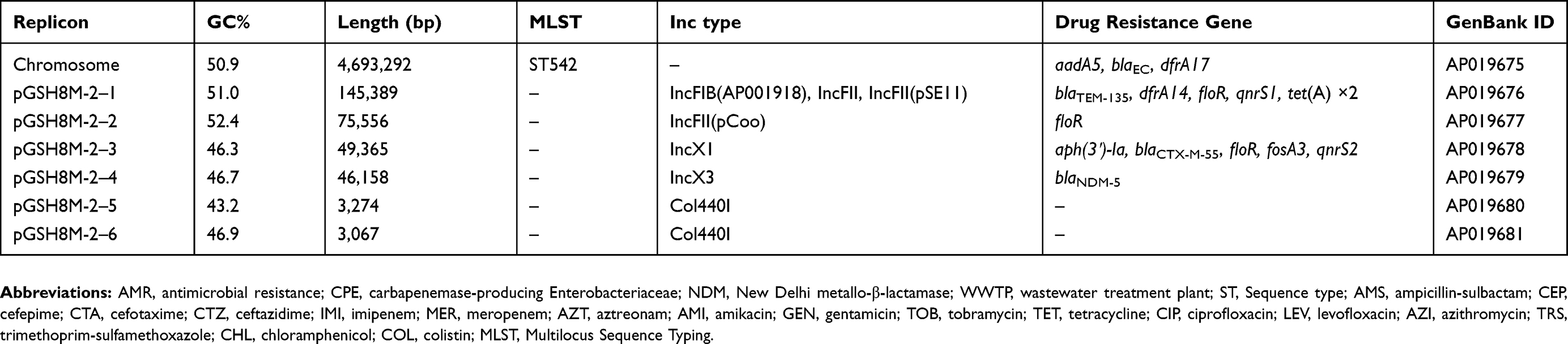 |
Table 2 Whole genome information of E. coli GSH8M-2 from WWTP effluent |
Acknowledgments
The authors would like to thank Rina Tanaka for technical assistance with the whole genome sequencing experiment. We would like to thank Editage for English language editing. This work was supported by the Research Program on Emerging and Re-emerging Infectious Diseases from the Japan Agency for Medical Research and Development, AMED (grant number JP19fk0108048). This work was also supported by a grant for Research on Emerging and Re-emerging Infectious Diseases and Immunization (H30 Shinkogyosei-Ippan-002) from the Ministry of Health, Labour, and Welfare, Japan. The funding agencies had no role in the study design, data collection and analysis, decision to publish, or manuscript preparation.
Author contributions
TS and MK contributed to the isolation of the NDM-5 producing strains. TS, YI, and MK performed the genome sequencing and the comparative genome analysis. TS and MK performed antimicrobial susceptibility testing. MK wrote the manuscript. All authors contributed to data analysis, drafting or revising the article, gave final approval of the version to be published, and agree to be accountable for all aspects of the work.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Logan LK, Weinstein RA. The epidemiology of carbapenem-resistant enterobacteriaceae: the impact and evolution of a global menace. J Infect Dis. 2017;215(suppl_1):S28–S36. doi:10.1093/infdis/jiw282
2. Walsh TR. Emerging carbapenemases: a global perspective. Int J Antimicrob Agents. 2010;36(Suppl 3):S8–S14. doi:10.1016/S0924-8579(10)70004-2
3. Hornsey M, Phee L, Wareham DW. A novel variant, NDM-5, of the New Delhi metallo-beta-lactamase in a multidrug-resistant Escherichia coli ST648 isolate recovered from a patient in the United Kingdom. Antimicrob Agents Chemother. 2011;55(12):5952–5954. doi:10.1128/AAC.05108-11
4. Ali A, Gupta D, Srivastava G, Sharma A, Khan AU. Molecular and computational approaches to understand resistance of New Delhi metallo beta- lactamase variants (NDM-1, NDM-4, NDM-5, NDM-6, NDM-7)-producing strains against carbapenems. J Biomol Struct Dyn. 2019;37(8):2061–2071. doi:10.1080/07391102.2018.1475261
5. Bi R, Kong Z, Qian H, et al. High prevalence of bla NDM variants among carbapenem-resistant Escherichia coli in Northern Jiangsu Province, China. Front Microbiol. 2018;9:2704. doi:10.3389/fmicb.2018.02704
6. Zhang F, Xie L, Wang X, et al. Further spread of bla NDM-5 in Enterobacteriaceae via IncX3 plasmids in Shanghai, China. Front Microbiol. 2016;7:424.
7. Li X, Fu Y, Shen M, et al. Dissemination of blaNDM-5 gene via an IncX3-type plasmid among non-clonal Escherichia coli in China. Antimicrob Resist Infect Control. 2018;7:59. doi:10.1186/s13756-018-0349-6
8. Sekizuka T, Yatsu K, Inamine Y, et al. Complete genome sequence of a blaKPC-2-positive Klebsiella pneumoniae strain isolated from the effluent of an Urban Sewage Treatment Plant in Japan. mSphere. 2018;3:5. doi:10.1128/mSphere.00314-18
9. Nordmann P, Poirel L, Dortet L. Rapid detection of carbapenemase-producing Enterobacteriaceae. Emerg Infect Dis. 2012;18(9):1503–1507. doi:10.3201/eid1809.120355
10. Sekizuka T, Matsui M, Takahashi T, et al. Complete genome sequence of bla IMP-6-positive Metakosakonia sp. MRY16-398 isolate from the ascites of a diverticulitis patient. Front Microbiol. 2018;9:2853. doi:10.3389/fmicb.2018.02853
11. Tanizawa Y, Fujisawa T, Nakamura Y. DFAST: a flexible prokaryotic genome annotation pipeline for faster genome publication. Bioinformatics. 2018;34(6):1037–1039. doi:10.1093/bioinformatics/btx713
12. Petkau A, Stuart-Edwards M, Stothard P, Van Domselaar G. Interactive microbial genome visualization with GView. Bioinformatics. 2010;26(24):3125–3126. doi:10.1093/bioinformatics/btq588
13. Zankari E, Hasman H, Cosentino S, et al. Identification of acquired antimicrobial resistance genes. J Antimicrob Chemother. 2012;67(11):2640–2644. doi:10.1093/jac/dks261
14. Carattoli A, Zankari E, Garcia-Fernandez A, et al. In silico detection and typing of plasmids using PlasmidFinder and plasmid multilocus sequence typing. Antimicrob Agents Chemother. 2014;58(7):3895–3903. doi:10.1128/AAC.02412-14
15. Yamashita A, Sekizuka T, Kuroda M. Characterization of antimicrobial resistance dissemination across plasmid communities classified by network analysis. Pathogens. 2014;3(2):356–376. doi:10.3390/pathogens3020356
16. Edgar RC. Search and clustering orders of magnitude faster than BLAST. Bioinformatics. 2010;26(19):2460–2461. doi:10.1093/bioinformatics/btq461
17. Shannon P, Markiel A, Ozier O, et al. Cytoscape: a software environment for integrated models of biomolecular interaction networks. Genome Res. 2003;13(11):2498–2504. doi:10.1101/gr.1239303
18. Krishnaraju M, Kamatchi C, Jha AK, et al. Complete sequencing of an IncX3 plasmid carrying blaNDM-5 allele reveals an early stage in the dissemination of the blaNDM gene. Indian J Med Microbiol. 2015;33(1):30–38. doi:10.4103/0255-0857.148373
19. Feng J, Qiu Y, Yin Z, et al. Coexistence of a novel KPC-2-encoding MDR plasmid and an NDM-1-encoding pNDM-HN380-like plasmid in a clinical isolate of Citrobacter freundii. J Antimicrob Chemother. 2015;70(11):2987–2991. doi:10.1093/jac/dkv232
20. Nakano R, Nakano A, Hikosaka K, et al. First report of metallo-beta-lactamase NDM-5-producing Escherichia coli in Japan. Antimicrob Agents Chemother. 2014;58(12):7611–7612. doi:10.1128/AAC.04265-14
21. Almakki A, Maure A, Pantel A, et al. NDM-5-producing Escherichia coli in an urban river in Montpellier, France. Int J Antimicrob Agents. 2017;50(1):123–124. doi:10.1016/j.ijantimicag.2017.04.003
22. Parvez S, Khan AU. Hospital sewage water: a reservoir for variants of New Delhi metallo-beta-lactamase (NDM)- and extended-spectrum beta-lactamase (ESBL)-producing Enterobacteriaceae. Int J Antimicrob Agents. 2018;51(1):82–88. doi:10.1016/j.ijantimicag.2017.08.032
23. Gomi R, Matsuda T, Yamamoto M, et al. Characteristics of carbapenemase-producing enterobacteriaceae in wastewater revealed by genomic analysis. Antimicrob Agents Chemother. 2018;62:5. doi:10.1128/AAC.02501-17
24. Zurfluh K, Stevens MJA, Stephan R, Nuesch-Inderbinen M. Complete and assembled genome sequence of an NDM-5- and CTX-M-15-producing Escherichia coli sequence type 617 isolated from wastewater in Switzerland. J Glob Antimicrob Resist. 2018;15:105–106. doi:10.1016/j.jgar.2018.08.015
25. Kiratisin P, Apisarnthanarak A, Saifon P, Laesripa C, Kitphati R, Mundy LM. The emergence of a novel ceftazidime-resistant CTX-M extended-spectrum beta-lactamase, CTX-M-55, in both community-onset and hospital-acquired infections in Thailand. Diagn Microbiol Infect Dis. 2007;58(3):349–355. doi:10.1016/j.diagmicrobio.2007.02.005
26. Hu X, Gou J, Guo X, et al. Genetic contexts related to the diffusion of plasmid-mediated CTX-M-55 extended-spectrum beta-lactamase isolated from Enterobacteriaceae in China. Ann Clin Microbiol Antimicrob. 2018;17(1):12. doi:10.1186/s12941-018-0265-x
27. Wang YH, Li XN, Chen C, Zhang J, Wang GQ. Detection of floR gene and active efflux mechanism of Escherichia coli in Ningxia, China. Microb Pathog. 2018;117:310–314. doi:10.1016/j.micpath.2018.02.042
28. Lu J, Zhang J, Xu L, et al. Spread of the florfenicol resistance floR gene among clinical Klebsiella pneumoniae isolates in China. Antimicrob Resist Infect Control. 2018;7:127. doi:10.1186/s13756-018-0415-0
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.