")
Back to Journals » Infection and Drug Resistance » Volume 16
Characterisation of a Novel Hybrid IncFIIpHN7A8:IncR:IncN Plasmid Co-Harboring blaNDM-5 and blaKPC-2 from a Clinical ST11 Carbapenem-Resistant Klebsiella pneumoniae Strain
Authors Sun L, Chen Y, Qu T, Shi K, Han X, Wu W, Jiang Y, Yu Y
Received 14 August 2023
Accepted for publication 16 November 2023
Published 12 December 2023 Volume 2023:16 Pages 7621—7628
DOI https://doi.org/10.2147/IDR.S435195
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. Héctor Mora-Montes
Lingyan Sun,1– 3,* Yan Chen,4– 6,* Tingting Qu,7,8 Keren Shi,4– 6 Xinhong Han,4– 6 Wenhao Wu,7,8 Yan Jiang,4– 6 Yunsong Yu4– 6
1Department of Laboratory Medicine, The First Affiliated Hospital, Zhejiang University School of Medicine, Hangzhou, People’s Republic of China; 2Key Laboratory of Clinical in Vitro Diagnostic Techniques of Zhejiang Province, Hangzhou, People’s Republic of China; 3Institute of Laboratory Medicine, Zhejiang University, Hangzhou, People’s Republic of China; 4Department of Infectious Diseases, Sir Run Run Shaw Hospital, Zhejiang University School of Medicine, Hangzhou, People’s Republic of China; 5Key Laboratory of Microbial Technology and Bioinformatics of Zhejiang Province, Hangzhou, People’s Republic of China; 6Regional Medical Center for National Institute of Respiratory Diseases, Sir Run Run Shaw Hospital, Zhejiang University School of Medicine, Hangzhou, People’s Republic of China; 7State Key Laboratory for Diagnosis and Treatment of Infectious Diseases, National Clinical Research Center for Infectious Diseases, Collaborative Innovation Center for Diagnosis and Treatment of Infectious Diseases, The First Affiliated Hospital, Zhejiang University School of Medicine, Hangzhou, People’s Republic of China; 8Infection Control Department, The First Affiliated Hospital, Zhejiang University School of Medicine, Hangzhou, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Yunsong Yu, Tel +86 138 0579 0432, Email [email protected]
Purpose: We aimed to characterize a novel blaNDM-5 and blaKPC-2 co-carrying hybrid plasmid from a clinical carbapenem-resistant Klebsiella pneumoniae (CRKP) strain.
Methods: Antimicrobial susceptibility was determined by the broth microdilution method. Plasmid size and localization were estimated using S1 nuclease pulsed-field gel electrophoresis (S1-PFGE) and Southern blotting. Plasmid transfer ability was evaluated by conjugation experiments. Whole genome sequencing (WGS) was performed using Illumina NovaSeq6000 and Oxford Nanopore MinION platforms. Genomic characteristics were analyzed using bioinformatics methods.
Results: Strain ZY27320 was a multidrug-resistant (MDR) clinical ST11 K. pneumoniae strain that confers high-level resistance to carbapenems (meropenem, MIC 128 mg/L; imipenem, MIC 64 mg/L) and ceftazidime/avibactam (MIC > 128/4 mg/L). Both S1-PFGE–Southern blotting and whole genome sequencing revealed that the carbapenemase genes blaKPC-2 and blaNDM-5 were carried by the same IncFIIpHN7A8:IncR:IncN hybrid plasmid (pKPC2_NDM5). Conjugation experiments indicated that pKPC2_NDM5 was a non-conjugative plasmid.
Conclusion: This is the first report of a hybrid plasmid carrying both carbapenemase genes blaNDM-5 and blaKPC-2 in a clinical K. pneumoniae ST11 isolate that confers resistance to both ceftazidime/avibactam and carbapenems, thereby presenting a serious threat to treatment in clinical practice.
Keywords: blaNDM-5, blaKPC-2, Klebsiella pneumoniae, hybrid plasmid, IS 26
Introduction
Carbapenem-resistant Klebsiella pneumoniae (CRKP) poses a global threat to clinical practice because of its ability to cause severe infections with high mortality rates.1,2 Carbapenemase production is the dominant cause of carbapenem resistance in carbapenem-resistant Enterobacteriaceae, with KPC and NDM being the most common. KPC-2 carbapenemase, encoded by blaKPC-2 is predominant in Klebsiella pneumoniae and can be potently inhibited by new β-lactamase inhibitors such as avibactam, vaborbactam, and relebactam. Thus, ceftazidime/avibactam is a valuable treatment option to combat CRKP-producing KPC carbapenemase.3 However, metallo-β-lactamase NDM encoded by blaNDM, which is most common in Escherichia coli in China,4,5 cannot be overcome using clinically available β-lactamase inhibitors, including ceftazidime/avibactam.6 CRKP, which produces both KPC and NDM, confers resistance to nearly all β-lactams and β-lactamase inhibitors, and presents a significant challenge in clinical treatment owing to extremely limited options. In recent years, the co-production of KPC and NDM (mainly KPC-2 and NDM-1) has become a severe threat to public health, with increasing reports worldwide,7–12 and has been documented in other Enterobacteriaceae.13–15 A nationwide survey in China revealed an increased prevalence of CRKP co-producing KPC-2 and NDM-1 in recent years.16 According to previous reports, blaKPC and blaNDM are usually located on separate plasmids that can be transferred to new recipient bacteria through conjugation. There are few reports on plasmids co-carrying blaKPC and blaNDM. Recently, an IncR plasmid known as pCF2075-1 carrying both blaKPC-2 and blaNDM-1 was identified in Citrobacter freundii.17 Additionally, an IncHI5 plasmid named pK254-KPC_NDM co-carrying blaKPC-2 and blaNDM-1 was discovered in Klebsiella michiganensis.18 Furthermore, a conjugative hybrid plasmid containing blaKPC-2 and blaIMP-4 that co-expressed serine- and metallo-carbapenemases was documented in a clinical Klebsiella quasipneumoniae isolate.19 Here, we describe the co-occurrence of NDM-5 as the most common metallo-carbapenemase and KPC-2 as the most prevalent serine β-lactamase on a chimeric plasmid from a clinical K. pneumoniae ST11 isolate.
Materials and Methods
Strain Identification and Antimicrobial Susceptibility Testing
Strain ZY27320 was obtained from an abdominal drainage sample of a man aged 61 with severe abdominal infection who was hospitalized in a tertiary hospital in February 2021. The isolate was identified as K. pneumoniae using a Bruker Microflex MALDI-TOF (Bruker, Germany).
The minimum inhibitory concentrations (MICs) of colistin, tigecycline, ceftazidime/avibactam, piperacillin/tazobactam, ceftolozane/tazobactam, imipenem/relebactam, meropenem/vaborbactam, meropenem, imipenem, ertapenem, levofloxacin, ciprofloxacin, amikacin, fosfomycin, and aztreonam were determined using the microbroth dilution method with E. coli ATCC 25922 as a quality control. The results were interpreted based on the MIC breakpoints of the Clinical and Laboratory Standards Institute (CLSI M100-S32, 2022). The MIC breakpoint of tigecycline was based on the standards of the US FDA and National Medical Products Administration (NMPA) in China.
S1 -PFGE and Southern Blot Hybridization
To clarify the plasmid location of blaKPC-2 and blaNDM-5, we performed S1-PFGE and Southern hybridization as described previously19 using digoxigenin-labelled blaKPC-2 and blaNDM-5-specific probes with an NBT/BCIP color detection kit (Roche, Mannheim, Germany).
Conjugation Assay
To assess plasmid transferability, we performed conjugation experiments by filter mating and broth mating using different recipient bacteria (E. coli J53, E. coli C600, and induced rifampin-resistant K. pneumoniae ATCC13883). E. coli J53 transconjugants were selected on MH agar plates containing 100 mg/L of sodium azide and 4 mg/L of meropenem. E. coli EC600 transconjugants were selected on MH agar plates containing 600 mg/L of rifampicin and 4 mg/L of meropenem. K. pneumoniae ATCC13883 transconjugants were selected on MH agar plates containing 300 mg/L of rifampicin and 4 mg/L of meropenem.
Whole Genome Sequencing and Analysis
The whole genome of ZY27320 was sequenced using the Illumina NovaSeq6000 (Illumina, San Diego, CA, USA) and Oxford Nanopore MinION (Nanopore, Oxford, UK) platforms (Digital-Micro Biotech Co., Ltd, Hangzhou, China). The hybrid assembly of Illumina and Nanopore reads was performed using Unicycler v0.4.8.20 Complete plasmid sequences were annotated using the RAST server.21 Multilocus sequence typing (MLST), resistance genes, virulence genes, and plasmid replicons were performed using MLST 2.0, ResFinder 4.1, VirulenceFinder 2.0, and PlasmidFinder 2.1 web databases (http://www.genomicepidemiology.org/services), respectively. Insertion sequence elements (ISs) were identified using the ISFinder.22 The self-transmissible mobile genetic elements (MGE) and origin of the transfer site were identified using the OriTfinder web server.23 Sequence alignment and visualization were performed using Proksee web server v1.1.4 and Easyfig v2.2.3.24
Accession Numbers
The complete chromosome and plasmid sequences of Klebsiella pneumoniae ZY27320 were deposited in GenBank under BioProject No. PRJNA975440; accession numbers CP128564-CP128565.
Results
Strain ZY27320 was resistant to all β-lactamase inhibitors (ceftazidime/avibactam, piperacillin/tazobactam, ceftolozane/tazobactam, imipenem/relebactam, and meropenem/vaborbactam), carbapenems (ertapenem, meropenem, and imipenem), quinolones (ciprofloxacin and levofloxacin), aminoglycosides (amikacin), fosfomycin, and aztreonam; it was susceptible only to tigecycline and colistin (Supplementary Table 1).
S1-PFGE detected one plasmid approximately 167 kb in size in ZY27320 cells. Southern blotting with KPC and NDM probes revealed bands at the same location, suggesting that blaKPC-2 and blaNDM-5 were located on the same plasmid (Supplementary Figure 1).
To better understand the genetic environment of blaKPC-2 and blaNDM-5, the complete genome of ZY27320 was successfully sequenced by Illumina and Nanopore sequencing. The hybrid assembly using Unicycler revealed a circular chromosome of 5,578,590 bp (accession no. CP128564) with a GC content of 57.3%, and one circular plasmid of 167,277bp, designated pKPC2-NDM5 (accession no. CP128565) with a GC content of 53.0%. MLST typing of the assembled genome showed that ZY27320 belonged to the ST11. The chromosome of ZY27320 carries three antimicrobial resistance (AMR) genes (conferring resistance to fosfomycin (fosA), β-lactams (blaSHV-182), and aminoglycosides (aadA2)) and three virulence factors, including fyuA encoding a siderophore receptor, iutA encoding a ferric aerobactin receptor, and an irp2 gene.
pKPC2_NDM5 is a multiple drug resistance (MDR) plasmid that carries multiple AMR genes conferring resistance to β-lactams (blaKPC-2, blaNDM-5, blaCTX-M-65, and blaTEM-1B), fosfomycin (fosA3), streptomycin (aadA2), sulfamethoxazole (sul1), aminoglycosides (rmtB), and trimethoprim (dfrA12). In addition, plasmid pKPC2_NDM5 showed no sequence similarity to the virulence plasmid pLVPK25 and contained no virulence factors. We compared the sequence of pKPC2_NDM5 with the plasmid sequences co-carrying blaKPC-2 and blaNDM-1 registered in GenBank.17,18 BLASTN results showed that pKPC2_NDM5 exhibited low coverage (20%) with an IncR plasmid pCF2075-1 (accession no. CP119166) from an Citrobacter freundii strain, and shared 24% coverage with an IncHI5 plasmid pK254-KPC_NDM (accession no. OM938013) from an Klebsiella michiganensis strain. Three replicons, IncFIIpHN7A8, IncR, and IncN, were identified in pKPC2_NDM5 by using PlasmidFinder. IncFIIpHN7A8:IncR blaKPC-bearing plasmids are widely disseminated in K. pneumoniae strains from China and are typically associated with ST11 strains.26 Plasmid pKPC2_NDM5 had high coverage (73–90%) and sequence similarity (99.93– 99.97%) with several IncFIIpHN7A8:IncR:IncN blaKPC2-carrying plasmids of K. pneumoniae strain from Zhengzhou City, China, such as pKP18-41-KPC2 (accession no. CP082012) and pKP19-2196-KPC2 (accession no. CP082042) and pKP18-2172-KPC2 (accession no. CP082039) and pKP18-1-KPC2 (accession no. CP082003) and pKPN-hnqyy-kpc (accession no. CP074118).27 Subsequently, a series of blaNDM-5-carrying plasmids with high similarity (99.53–100%) appeared to carry complementary sequences to form the backbone of pKPC2_NDM5. A blaNDM-5-carrying plasmid pKPN-hnqyy-ndm (accession no. CP074117) exhibited 33% coverage and 100% identity, coincidentally originating from the same NDM-5 and KPC-2 co-producing K. pneumoniae isolate K9 as the blaKPC-2-carrying plasmid pKPN-hnqyy-kpc (accession no. CP074118).27 Interestingly, these two plasmids from a single NDM-5 and KPC-2 coproducing K. pneumoniae appeared to integrate into the backbone of pKPC2_NDM5 (Figure 1). Detailed sequence analysis revealed that pKPC2_NDM5 was a cointegrate consisting of four regions: an ~116 kb IncFIIpHN7A8:IncR:IncN plasmid-derived KPC-module (Region-1), an ~ 2.4 kb fosA carrying region (Region-2), an ~ 43.5kb blaNDM-5 carrying NDM-module (Region-3), and an ~ 4.6 kb blaTEM-1B and rmtB carrying region (Region-4). The Region-1 was perfectly aligned with the plasmid pKPN-hnqyy-kpc and Region-3 shares 100% sequence identity with that of plasmid pKPN-hnqyy-ndm. All four regions are bracketed by two IS26 elements (Figure 2). The formation of pKPC2_NDM5 appeared to be a recombination event in which an IS26-flanked NDM-fragment, IS26-bounded fosA-fragment and an IS26-bounded blaTEM-1B - rmtB-fragment were inserted into an IncFIIpHN7A8:IncR:IncN KPC-plasmid (plasmid pKPN-hnqyy-kpc) using IS26 as the insertion site.
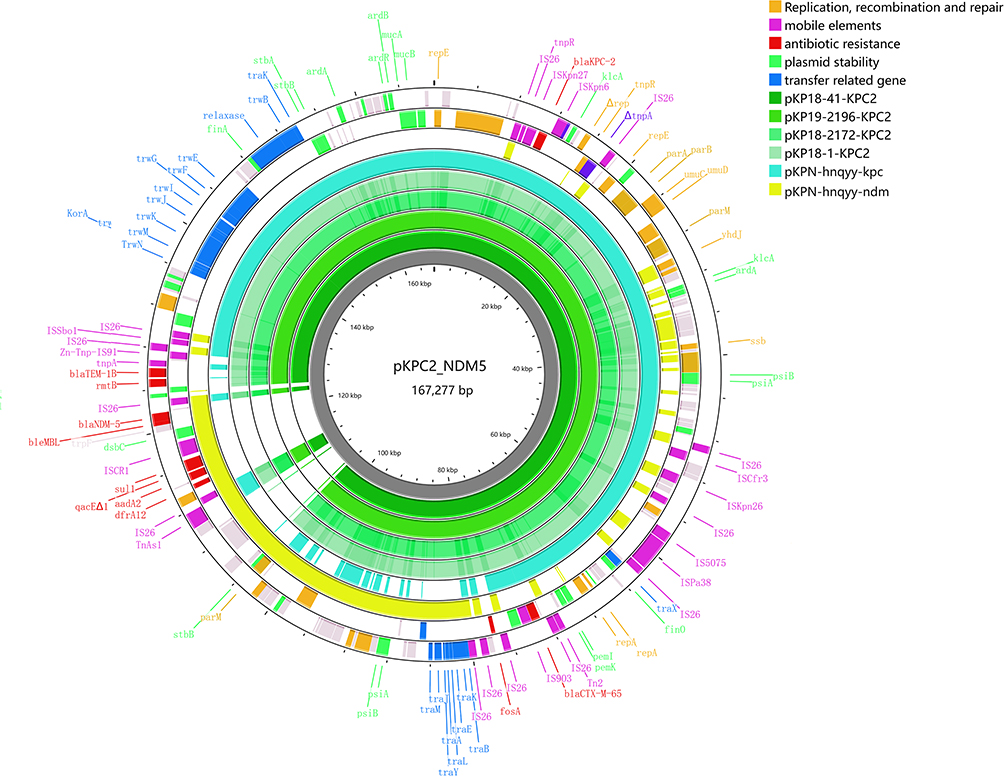 |
Figure 1 Schematic map of plasmid pKPC2_NDM5 and alignment with other similar plasmids from GenBank. The visualization map was generated by Proksee web server v1.1.4. |
 |
Figure 2 Sequence alignment of plasmid pKPC2_NDM5 with sequences of pKPN-hnqyy-ndm (GenBank accession no. CP074117), and pKPN-hnqyy-kpc (GenBank accession no. CP074118). The visualization map was created by EasyFig software. |
We further analyzed the environments of blaKPC-2 and blaNDM-5 (Figure 3). blaKPC-2 is located on a non-Tn4401 element with the following structures: IS26, ΔtnpR, ISKpn27, blaKPC-2, and ISKpn6-korC-klcA-ΔrepA-tnpR-ΔtnpA-IS26. The gene blaKPC has typically been found in classical Tn4401 elements,26 as well as non-Tn4401 elements (NTEKPC), including NTEKPC-I, NTEKPC-II, and NTEKPC-III.28,29 The blaKPC-bearing genetic elements in pKPC2-NDM5 were assigned to the NTEKPC-I group because ΔblaTEM or Tn5563/IS6100 were not inserted upstream of blaKPC-2.30 Compared with the blaKPC-carrying fragment with the NTEKPC-Ib structure in pKPHS2 (Accession no. CP003224), a large part of Tn1721 tnpA was absent and was replaced by an IS26 element in the blaKPC-2 genetic context of pKPC2_NDM5. In addition, the genetic environment of blaKPC-2 in pKPC2_NDM5 had 92% query coverage compared to that of the plasmid pKPC2_020009 (accession no. CP038003) carrying a truncated tnpA of Tn1721.
 |
Figure 3 (A) Linear comparison of genetic environment of blaNDM-5 of pKPC2_NDM5 and that of pNDM_2_P21_SE1_04.20. (B) Linear comparison of the genetic environment of blaKPC-2 among pKP048, pKPHS2, pKPC2_NDM5, and pKPC2_020009. The plasmid comparison figure was generated by EasyFig software. Regions with identical sequences are shown in gray shadow between the different plasmids. |
The blaNDM-5 gene is located in a 16.7-kb multi-resistance module carrying dfrA12, aadA2, sul1, blaNDM-5, rmtB, and blaTEM-1B flanked by IS26 elements. The blaNDM-bearing element of pKPC2_NDM5 carries the common feature of ΔISAba125-blaNDM-bleMBL6 and has the same genes as classical Tn125 including trpF and dsbC downstream of bleMBL. In contrast to Tn125, the blaNDM-carrying element in pKPC2_NDM5 lost several genes, including cutA1, groES-groEL, and ISCR27, but had one additional IS26 element upstream of ΔISAba125 and carried an additional AGR gene cluster (ISCR1-sul1-aadA2-dfrA12-intI1-IS26) downstream of dsbC. BLAST analysis revealed that the blaNDM-bearing element in pKPC2_NDM5 was highly similar (99.99% identity and 94% coverage) to that of pNDM_2_P21_SE1_04.20 (accession no. CP085076.1) of one clinical Escherichia coli isolate from the United Kingdom (Figure 3) and many other plasmids, such as pM217_FII from Myanmar (not shown).
To understand the transmission capability of pKPC2_NDM5, we performed conjugation assays. We attempted filter mating and broth mating with different recipient bacteria (K. pneumoniae ATCC13883, E. coli strains EC600 and ECJ53) and tested various donor: recipient ratios. We did not obtain a transconjugant under any conditions. Plasmid extraction and electroporation were both unsuccessful. These results suggest that pKPC2_NDM5 is nontransmissible. To test this hypothesis, we submitted the plasmid sequence to the OriTfinder web server.23,31 The plasmid pKPC2_NDM5 sequence contained an oriT region (153,968–154,068 bp) with a conserved nick region (154,025–154,034 bp), relaxase-encoding gene, type IV coupling protein-encoding gene trwB, and two T4SS gene clusters: traB-traK-traE-traL-traA-traY-traM (80,324–85,114 bp) and trwN-trwM-trwK-trwJ-trwI-trwG-trwF-trwE (135,941–153,401 bp). The conjugative region of the self-transmissible MGE seems to be intact, but pKPC2_NDM5 lacks transfer ability which still presents a puzzle.
Discussion
blaKPC-2 is the predominant carbapenemase gene widely spread among Klebsiella pneumoniae ST11 and is usually carried by IncFII-like plasmids in China.32 In our study, the backbone of pKPC2-NDM5 displayed a multi-replicon IncFIIpHN7A8:IncR:IncN chimera structure, which may have evolved from several recombination events as described in previous study.27 NDM-5, as a variant of NDM-1, was found to exhibit enhanced carbapenemase activity and hydrolysis ability of ceftazidime/avibactam,33 and was carried by plasmids with diverse replicon types, including IncX3, IncFII, IncC, IncN, and IncR. Co-harboring blaNDM-5 and blaKPC-2 on a multidrug-resistant plasmid confers high-level resistance to both carbapenems and ceftazidime/avibactam and therefore becomes a significant challenge in clinical treatment. The plasmids co-carrying blaKPC-2 and blaNDM-1 were recently reported in Citrobacter freundii and Klebsiella michiganensis.17,18 The emergence of this kind of plasmid co-carrying blaKPC-2 and blaNDM-5 in the most prevalent ST clone of K. pneumoniae, poses a significant public health threat and deserves significant attention.
Plasmid pKPC2-NDM5 appeared to be a hybrid plasmid generated by the movement of an IS26-associated translatable unit. The high copy number of IS26 in pKPC2_NDM5 and its presence in each composition module suggest that IS26 plays an essential role in the mobilization and recombination of AMR genes.34,35 However, we did not recognize matching pairs of target site duplications (TSD) at the junction of each IS26, which are usually considered as the “tracers” of transposition. The lack of flanking TSDs was not uncommon, as reported previously, perhaps because of homologous recombination following IS26 transposition.36 Another explanation is that IS26 transposes through replicative transposition, which can generate IS26 copies without two-sided TSDs.34
pKPC2-NDM5 was non-conjugative, although it carried conjugal modules of the self-transmissible MGE. Interestingly, as the backbone of pKPC2_NDM5, the plasmid pKPN-hnqyy-kpc was proved to have high transfer ability to E. coli J53, however, plasmid pKPN-hnqyy-ndm was a non-conjugative plasmid.27 Previous studies have shown that the expression of conjugative systems is tightly controlled, because the conjugation process is energetically expensive.37 Although we could not measure the fitness cost of this plasmid, we suspected that the loss of horizontal transfer ability was a manifestation of fitness cost. Furthermore, although there have been many reports on the concurrence of the blaNDM-plasmid and blaKPC-plasmid in the same K. pneumoniae isolate,9,16,38 the same plasmid co-harboring blaNDM and blaKPC in K. pneumoniae has not yet been reported. However, with the widespread use of ceftazidime/avibactam and frequent combined with carbapenems in clinical practice, plasmids co-carrying these two critical carbapenemase genes might gradually gain a selection advantage during continuous evolution. Therefore, KPC-NDM-plasmid could become a great challenge in the treatment of CRE infections and is worth strengthening monitoring in the future.
Conclusion
In this study, we characterized a novel hybrid MDR plasmid co-harboring blaKPC-2 and blaNDM-5 obtained from an ST11 clinical K. pneumoniae isolate. This chimeric plasmid is likely a consequence of a recombination event involving the insertion of a blaNDM-5 fragment into an IncFIIpHN7A8:IncR:IncN blaKPC2-carrying plasmid, probably mediated by IS26 elements. Despite the lack of plasmid self-transmissibility, integration of these two critical carbapenemase genes, blaNDM-5 and blaKPC-2, into the same plasmid in K. pneumoniae presents great challenges for clinical treatment and requires stringent surveillance.
Ethical Approval
This study was approved by the Ethics Committee of the First Affiliated Hospital of Zhejiang University School of Medicine with the research ethics approval code of IIT20230943A. The study was conducted in accordance with the principles of the Declaration of Helsinki.
Funding
This work was supported by grants from the National Natural Science Foundation of China (grant numbers: 82002169, 81830069, and 32141001).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Ben-David D, Kordevani R, Keller N, et al. Outcome of carbapenem resistant Klebsiella pneumoniae bloodstream infections. Clin Microbiol Infect. 2012;18(1):54–60. doi:10.1111/j.1469-0691.2011.03478.x
2. Tamma PD, Goodman KE, Harris AD, et al. Comparing the outcomes of patients with carbapenemase-producing and non-carbapenemase-producing carbapenem-resistant Enterobacteriaceae bacteremia. Clin Infect Dis. 2017;64(3):257–264. doi:10.1093/cid/ciw741
3. Yahav D, Giske CG, Gramatniece A, Abodakpi H, Tam VH, Leibovici L. New beta-lactam-beta-lactamase inhibitor combinations. Clin Microbiol Rev. 2020;34(1). doi:10.1128/CMR.00115-20
4. Wang Q, Wang X, Wang J, et al. Phenotypic and genotypic characterization of carbapenem-resistant Enterobacteriaceae: data from a longitudinal large-scale CRE Study in China (2012–2016). Clin Infect Dis. 2018;67(suppl_2):S196–S205. doi:10.1093/cid/ciy660
5. Han R, Shi Q, Wu S, et al. Dissemination of carbapenemases (KPC, NDM, OXA-48, IMP, and VIM) among carbapenem-resistant Enterobacteriaceae isolated from adult and children patients in China. Front Cell Infect Mi. 2020;10:314. doi:10.3389/fcimb.2020.00314
6. Wu W, Feng Y, Tang G, Qiao F, McNally A, Zong Z. NDM metallo-beta-lactamases and their bacterial producers in health care settings. Clin Microbiol Rev. 2019;32(2). doi:10.1128/CMR.00115-18
7. Vasquez-Ponce F, Dantas K, Becerra J, et al. Detecting KPC-2 and NDM-1 coexpression in Klebsiella pneumoniae complex from human and animal hosts in South America. Microbiol Spectr. 2022;10(5):e115922. doi:10.1128/spectrum.01159-22
8. Tekeli A, Dolapci I, Evren E, Oguzman E, Karahan ZC. Characterization of Klebsiella pneumoniae coproducing KPC and NDM-1 Carbapenemases from Turkey. Microb Drug Resist. 2020;26(2):118–125. doi:10.1089/mdr.2019.0086
9. Hu R, Li Q, Zhang F, Ding M, Liu J, Zhou Y. Characterisation of bla (NDM-5) and bla (KPC-2) co-occurrence in K64-ST11 carbapenem-resistant Klebsiella pneumoniae. J Glob Antimicrob Resist. 2021;27:63–66. doi:10.1016/j.jgar.2021.08.009
10. Tang M, Li J, Liu Z, et al. Clonal transmission of polymyxin B-resistant hypervirulent Klebsiella pneumoniae isolates coharboring bla (NDM-1) and bla (KPC-2) in a tertiary hospital in China. BMC Microbiol. 2023;23(1):64. doi:10.1186/s12866-023-02808-x
11. Hao J, Zhang B, Deng J, Wei Y, Xiao X, Liu J. Emergence of a hypervirulent tigecycline-resistant Klebsiella pneumoniae strain co-producing bla (NDM-1) and bla (KPC-2) with an uncommon sequence type ST464 in Southwestern China. Front Microbiol. 2022;13:868705. doi:10.3389/fmicb.2022.868705
12. Ahmed M, Yang Y, Yang Y, et al. Emergence of hypervirulent carbapenem-resistant Klebsiella pneumoniae coharboring a bla (NDM-1)-carrying virulent plasmid and a bla (KPC-2)-carrying plasmid in an Egyptian hospital. Msphere. 2021;6(3). doi:10.1128/mSphere.00088-21
13. Fu L, Wang S, Zhang Z, et al. Co-carrying of KPC-2, NDM-5, CTX-M-3 and CTX-M-65 in three plasmids with serotype O89: H10 Escherichia coli strain belonging to the ST2 clone in China. Microb Pathogenesis. 2019;128:1–6. doi:10.1016/j.micpath.2018.12.033
14. Li Y, Fang C, Qiu Y, Dai X, Zhang L. Genomic characterization of a carbapenem-resistant Citrobacter freundii cocarrying bla (KPC-2) and bla (NDM-1). J Glob Antimicrob Resist. 2022;29:289–292. doi:10.1016/j.jgar.2022.04.014
15. Bes T, Nagano D, Martins R, et al. Bloodstream infections caused by Klebsiella pneumoniae and Serratia marcescens isolates co-harboring NDM-1 and KPC-2. Ann Clin Microbiol Antimicrob. 2021;20(1):57. doi:10.1186/s12941-021-00464-5
16. Gao H, Liu Y, Wang R, Wang Q, Jin L, Wang H. The transferability and evolution of NDM-1 and KPC-2 co-producing Klebsiella pneumoniae from clinical settings. Ebiomedicine. 2020;51:102599. doi:10.1016/j.ebiom.2019.102599
17. Zhang F, Li Z, Liu X, et al. Carbapenem-resistant Citrobacter freundii harboring bla (KPC-2) and bla (NDM-1): a study on their transferability and potential dissemination via generating a transferrable hybrid plasmid mediated by IS6100. Front Microbiol. 2023;14:1239538. doi:10.3389/fmicb.2023.1239538
18. Luo X, Dong F, Dai P, et al. Coexistence of blaKPC-2 and blaNDM-1 in one IncHI5 plasmid confers transferable carbapenem resistance from a clinical isolate of Klebsiella michiganensis in China. J Glob Antimicrob Resist. 2023;35:104–109. doi:10.1016/j.jgar.2023.09.006
19. Quan J, Dai H, Liao W, et al. Etiology and prevalence of ESBLs in adult community-onset urinary tract infections in East China: a prospective multicenter study. J Infection. 2021;83(2):175–181. doi:10.1016/j.jinf.2021.06.004
20. Wick RR, Judd LM, Gorrie CL, Holt KE, Phillippy AM. Unicycler: resolving bacterial genome assemblies from short and long sequencing reads. PLOS Comput Biol. 2017;13(6):e1005595. doi:10.1371/journal.pcbi.1005595
21. Aziz RK, Bartels D, Best AA, et al. The RAST server: rapid annotations using subsystems technology. BMC Genomics. 2008;9(1):75. doi:10.1186/1471-2164-9-75
22. Siguier P, Perochon J, Lestrade L, Mahillon J, Chandler M. ISfinder: the reference centre for bacterial insertion sequences. Nucleic Acids Res. 2006;34(Database issue):D32–D36. doi:10.1093/nar/gkj014
23. Li X, Xie Y, Liu M, et al. oriTfinder: a web-based tool for the identification of origin of transfers in DNA sequences of bacterial mobile genetic elements. Nucleic Acids Res. 2018;46(W1):W229–W234. doi:10.1093/nar/gky352
24. Sullivan MJ, Petty NK, Beatson SA. Easyfig: a genome comparison visualizer. Bioinformatics. 2011;27(7):1009–1010. doi:10.1093/bioinformatics/btr039
25. Chen YT, Chang HY, Lai YC, Pan CC, Tsai SF, Peng HL. Sequencing and analysis of the large virulence plasmid pLVPK of Klebsiella pneumoniae CG43. Gene. 2004;337:189–198. doi:10.1016/j.gene.2004.05.008
26. Yang X, Dong N, Chan EW, Zhang R, Chen S. Carbapenem resistance-encoding and virulence-encoding conjugative plasmids in Klebsiella pneumoniae. Trends Microbiol. 2021;29(1):65–83. doi:10.1016/j.tim.2020.04.012
27. Xiao W, Wang X, Qu Y, et al. Identification of two novel carbapenemase-encoding hybrid plasmids harboring bla (NDM-5) and bla (KPC-2) in a clinical ST11-KL47 Klebsiella pneumoniae. Infect Drug Resist. 2023;16:4073–4081. doi:10.2147/IDR.S408824
28. Cerdeira LT, Lam M, Wyres KL, et al. Small IncQ1 and col-like plasmids harboring bla (KPC-2) and non-Tn4401 elements (NTE (KPC)-IId) in high-risk lineages of Klebsiella pneumoniae CG258. Antimicrob Agents Chemother. 2019;63(3). doi:10.1128/AAC.02140-18
29. Octavia S, Kalisvar M, Venkatachalam I, et al. Klebsiella pneumoniae and Klebsiella quasipneumoniae define the population structure of blaKPC-2Klebsiella: a 5 year retrospective genomic study in Singapore. J Antimicrob Chemoth. 2019;74(11):3205–3210. doi:10.1093/jac/dkz332
30. Chen L, Mathema B, Chavda KD, DeLeo FR, Bonomo RA, Kreiswirth BN. Carbapenemase-producing Klebsiella pneumoniae: molecular and genetic decoding. Trends Microbiol. 2014;22(12):686–696. doi:10.1016/j.tim.2014.09.003
31. de la Cruz F, Frost LS, Meyer RJ, Zechner EL. Conjugative DNA metabolism in gram-negative bacteria. Fems Microbiol Rev. 2010;34(1):18–40. doi:10.1111/j.1574-6976.2009.00195.x
32. Fu P, Tang Y, Li G, Yu L, Wang Y, Jiang X. Pandemic spread of bla((KPC-2)) among Klebsiella pneumoniae ST11 in China is associated with horizontal transfer mediated by IncFII-like plasmids. Int J Antimicrob Ag. 2019;54(2):117–124. doi:10.1016/j.ijantimicag.2019.03.014
33. Hornsey M, Phee L, Wareham DW. A novel variant, NDM-5, of the New Delhi metallo-β-lactamase in a multidrug-resistant Escherichia coli ST648 isolate recovered from a patient in the United Kingdom. Antimicrob Agents Chemother. 2011;55(12):5952–5954. doi:10.1128/AAC.05108-11
34. He S, Hickman AB, Varani AM, et al. Insertion sequence IS26 reorganizes plasmids in clinically isolated multidrug-resistant bacteria by replicative transposition. Mbio. 2015;6(3):e762. doi:10.1128/mBio.00762-15
35. Zhao QY, Zhu JH, Cai RM, et al. IS26 is responsible for the evolution and transmission of bla (NDM)-harboring plasmids in Escherichia coli of Poultry Origin in China. Msystems. 2021;6(4):e64621. doi:10.1128/mSystems.00646-21
36. Hudson CM, Bent ZW, Meagher RJ, Williams KP, Hall R. Resistance determinants and mobile genetic elements of an NDM-1-encoding Klebsiella pneumoniae strain. PLoS One. 2014;9(6):e99209. doi:10.1371/journal.pone.0099209
37. Koraimann G, Wagner MA. Social behavior and decision making in bacterial conjugation. Front Cell Infect Mi. 2014;4:54. doi:10.3389/fcimb.2014.00054
38. Boralli C, Paganini JA, Meneses RS, et al. Characterization of bla (KPC-2) and bla (NDM-1) plasmids of a K. pneumoniae ST11 outbreak clone. Antibiotics-Basel. 2023;12(5). doi:10.3390/antibiotics12050926
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.