Back to Journals » Diabetes, Metabolic Syndrome and Obesity » Volume 18
Changes to the Haematological Parameters of Rat Offspring Born From High Fat High Carbohydrate (HFHC) Diet-Induced Prediabetic and Preeclamptic Sprague Dawley Rats: Assessing the Effects on Selected Haematological Markers
Authors Xulu N ![]() , Nkosi A, Khathi A, Sibiya NH
, Nkosi A, Khathi A, Sibiya NH ![]() , Ngubane PS
, Ngubane PS ![]()
Received 11 September 2023
Accepted for publication 21 December 2023
Published 21 March 2025 Volume 2025:18 Pages 831—845
DOI https://doi.org/10.2147/DMSO.S436001
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Konstantinos Tziomalos
Nombuso Xulu,1 Ayanda Nkosi,1 Andile Khathi,1 Ntethelelo H Sibiya,2 Phikelelani S Ngubane1
1Schools of Laboratory Medicine and Medical Sciences, College of Health Sciences, University of KwaZulu-Natal, Durban, South Africa; 2Pharmacology Division, Faculty of Pharmacy, Rhodes University, Makhanda, South Africa
Correspondence: Nombuso Xulu, Email [email protected]
Introduction: Infants delivered from preeclamptic pregnancies frequently exhibit developmental programming which leads to foetal growth restriction and foetal haematological abnormalities. Diabetes is recognised as a predisposing factor for preeclampsia (PE). Hyperglycaemia, a characteristic feature of pregestational type 2 diabetes, has been associated with the pathogenesis of intrauterine growth restriction (IUGR), a condition associated with disrupted foetal haematological pathways. Prediabetes pre-empts the onset of type 2 diabetes and is characterised by moderately elevated blood glucose levels, which have been shown in prediabetic models to induce erythrocyte dysfunction. However, the precise relationship between prediabetes and the development of preeclampsia or associated foetal complications remains to be fully elucidated. Accordingly, this study aims to investigate prediabetes as a risk factor for preeclampsia and its effects on selected haematological markers in Sprague Dawley rat pups.
Methods and Materials: Male and female pups born from normal, L-NAME preeclamptic and HFHC diet-induced prediabetic dams were immediately collected and weighed. The pups were then carefully returned to the dams for further development. On day 21, the pups were weaned and separated into males and females. Thereafter, the pups were sacrificed using a guillotine and blood and plasma was collected for haematological and biochemical analysis.
Results: Pups born from prediabetic and preeclamptic dams exhibited significantly lower birth weights than those born from normal pregnancies. Moreover, pups born from prediabetic and preeclamptic dams exhibited dysregulation of red blood cell (RBC) count, granulocyte colony-stimulating factor (G-CSF), erythropoietin (EPO) levels, glutathione peroxidase (GPx) and malondialdehyde (MDA) concentrations compared to those delivered from normal dams.
Conclusion: These findings suggest prediabetes caused dysregulation of haematological parameters in offspring and may be a predisposing factor for the development of preeclampsia in pregnancy. Therefore, strict monitoring of prediabetes during pregnancy may reduce the risk of preeclampsia and resultant foetal morbidity and mortality.
Keywords: pregestational diabetes, preeclampsia, intrauterine growth restriction, haematological parameters
Introduction
The development of the foetal blood cell lineage begins in the very first week of embryonic life.1 Blood cells are an essential component of maternofoetal circulation, with red blood cells playing a critical role in oxygen delivery to foetal tissues.1 The failure of foetal blood cellular development, maturation, and function can result in disastrous consequences for both the mother and child.1,2 Preeclampsia (PE) is a complex hypertensive disorder characterised by the development of de novo hypertension (140/90mmHg), proteinuria (300 mg/day), and oedema.2 PE is linked with oxidative stress, inflammation, and endothelial dysfunction, which mirrors the aetiology of diabetes.2 According to the World Health Organization (WHO), even with the provision of high-level tertiary care, hypertensive disorders of pregnancy or preeclamptic-eclamptic syndromes with underlying diabetic disorders are the primary causes of pregnancy-related maternal and infant deaths.2,3 Moreover, studies show that neonates born from preeclamptic women or women with diabetes are more susceptible to incur haematological permutations and this is further worsened when these disorders co-exist.3 In normal pregnancy, haematological pathways like erythropoiesis, leukopoiesis, and thrombopoiesis are narrowly regulated to ensure the proper growth of the foetus.3,4 However, in PE and diabetic pregnancies such as gestational diabetes mellitus (GDM) and pregestational diabetes, aberrant changes occur in foetal haematological pathways resulting in haematological derangements.5
Similar to PE, diabetic pregnancies cause intrauterine growth restriction (IUGR), which is implicated in the dysregulation of foetal haematological pathways leading to erythrocytosis via increased erythropoiesis (EPO) production, increased nucleated red blood cell (nRBC) production, increased mean cell volume (MCV), mean corpuscular haemoglobin (MCH), mean corpuscular haemoglobin (MCHC) and increased haemoglobin (Hgb) concentrations.6 Furthermore, IUGR has also been shown to cause complications which alter the production of mature myeloblasts via the dysregulation of granulocyte colony-stimulating factor (G-CSF) leading to leukocyte derangements such as neutropenia and platelet complications. These aberrant changes are not well elucidated; however, the literature implicates placental hypoxia observed during PE and pregestational diabetic pregnancies as the main cause.7,8
Placental hypoxia increases reactive oxygen species (ROS) which cause oxidative stress decreased proangiogenic factors as well as increases the unvetted release of inflammatory mediators.8 These constituents cross the maternoplacental environment during critical stages of embryonic development resulting in a plethora of complications.8 ROS and inflammatory mediators aid in foetal growth restriction which results in low infant birth weight and has been shown in the literature to negatively affect haematological pathways as it is directly linked to infant developmental milestones.9 Studies on haematological complications have been done in preeclamptic and pregestational type 2 diabetic pregnancies.9 However, limited research is available on the effects of pregestational prediabetes on foetal growth and development, specifically looking at its effects on haematological indices and how these changes relate to preeclampsia, such as prediabetes like type 2 diabetes, which may be a risk factor for PE.9
Prediabetes is a state where blood glucose concentrations are above the normal range but are not high enough to be classified as type 2 diabetes (T2D) mellitus.10 Prediabetes precedes T2D and is characterised by hyperglycaemia and insulin resistance resulting from the consumption of high carbohydrate and saturated fatty acid-based diets.10 In our laboratory, a high-fat high carbohydrate (HFHC) diet-induced prediabetic animal model has been established in female animals and its effects on glucose handling have been investigated.10 However, high-fat high carbohydrate (HFHC) diet-induced prediabetes as a risk factor for preeclampsia and its effect on haematological foetal outcomes are yet to be established.11 Therefore, the aim of this study is to investigate the effects of HFHC diet-induced prediabetes in relation to preeclampsia and the resulting adverse haematological outcomes in offspring-born female Sprague Dawley rats.
Materials and Methods
Drugs and Chemicals
All chemicals and reagents were sourced from standard pharmaceutical suppliers and were of analytical grade, including isofor (Safeline Pharmaceuticals (Pty) Ltd., Roodepoort, South Africa), liquid nitrogen (Chemistry Department UKZN, Westville, South Africa), ELISA kits (Elabscience, Mzansi Medical Laboratories, South Africa), metformin, phosphoric acid (Sigma, Durban, South Africa) thiobarbituric acid (Sigma, Durban, South Africa), hydrochloric acid (Sigma, Durban, South Africa), and butanol (Sigma, Durban, South Africa).
Animals and Induction of Prediabetes Mellitus
Animals
Female Sprague-Dawley rats (150–180g), which will be used for breeding, were housed in the Biomedical Research Unit of the University of KwaZulu-Natal and were used in the study. All animal procedures and housing conditions were approved by the Animal Research Ethics Committee of the University of KwaZulu-Natal, which conforms to the principles and guidelines of the Canadian Council on Animal Care. The Animal Research Ethics Committee of the University of of KwaZulu-Natal (AREC/032/020D) thoroughly assessed and granted approval for the conduct of animal experimentation.
Induction of Prediabetes
Experimental prediabetes was induced in the animals by using a previously described protocol by.10 The experimental animals will be exposed to a HCHF diet and 15% fructose in drinking water for 20 weeks. The control group will give standard chow for 20 weeks. The World Health Organisation criteria will be used for the diagnosis of prediabetes. Pre-diabetic symptoms including fasting blood glucose concentration of 6.1–7.1mmol/L, oral glucose tolerance test, two-hour glucose concentration of 7.1–8.1mmol/L and plasma triglycerides concentration of greater than 2 mmol/L will be considered pre-diabetic. Blood samples will be collected from the tail vein through the tail prick method and the blood glucose concentration levels will be measured using the Elite glucometer.
HFHC Diet
The high-fat high-carbohydrate diet (AVI food in Amanzimtoti, South Africa) is going to be formulated based on carbohydrate (55% Kcal/g), fats (30% Kcal/g), and protein (15% Kcal/g) composition.10 The drinking water of the experimental animals is going to be supplemented with 15% fructose. Procedures involving animal care will be conducted in conformity with the institutional guidelines for animal care of the University of KwaZulu-Natal.
Mating
Female Sprague-Dawley rats will be mated with males of the same strain. Briefly, two females will be housed in one cage to allow their oestrus cycle to become synchronised. The ovarian cycle of rats is between four to five days long. Daily vaginal smears will be taken using the pipette smear technique and when the female rats are in the pro-estrus stage of their cycle a male rat will be introduced into the cage. The following day, vaginal smears will be taken from the female rats and the presence of sperm will indicate successful mating and pregnancy will be confirmed by looking at cytology. The male rats will then be removed from the cage and male rats will be returned to the BRU. The day of fertilisation will be regarded as Gestational Day (GND) 0.
Induction of Preeclampsia
After pregnancy is confirmed by the visualisation of spermatozoa in a vaginal smear, the pregnant dams will be randomly divided into one rat per cage, which will be given 0.3g/L a day of L-NAME in drinking water from day 14.5 to day 20.5 of gestation and then blood pressure will be measured and the presence or absence of protein in the urine will be assessed. Preeclampsia will be confirmed by the measure of blood pressure which will be monitored every third day after administration. Animals presenting with a MAP of 140/90 mmHg and proteinuria of 300mg/day will be considered preeclamptic. The animals will be given ad libitum access to the drinking water containing L-NAME. Blood will be monitored using a non-invasive tail-cuff method with photoelectric sensors (IITC Model 31 Computerised Blood Pressure Monitor, Life Sciences, Woodland Hills, California, USA). Blood pressure will be measured on GND 0, 12, and 19. Urine samples will be collected over the night using metabolic cages on GND 0, 12, and 19.
Male and Female Pups Were Collected for the Study
Male and female pups born from normal, L-NAME preeclamptic3 and HFHC diet-induced prediabetic11 dams were immediately collected and weighed. The pups were then carefully returned to the dams and were placed in Makrolon polycarbonate cages and were given access to food and water. The animals were kept with their mothers for a period of 21 days under standard laboratory conditions (for temperature and humidity) in a 12hr day and 12hr night cycle. At day 21 the pups were sacrificed for terminal studies.
Study Design
The effect of prediabetes on haematological function was investigated, the pups were separated into the following groups, Group 1: Male and female pups born from normal pregnant control rats (NCON) which received normal standard chow and normal drinking water throughout the study (n = 12/group); Group 2: Male and female pups born from PD pregnant rats which received the HCHF diet+ 15% fructose throughout the study (n = 12/group); Group3: Male and female pups born from PE pregnant control rats which received normal standard chow (n = 12/group). The pups were immediately weighed after birth and then returned to the dams for further development. On postnatal day 21, animals were weaned and divided into pups born from normal control dams (PCON), pups born from preeclamptic dams (PPE), and pups born from prediabetic dams (PPD) (n=12/group).
Terminal Studies
Thereafter, the guillotine was used to decapitate pups for blood collection into EDTA tubes (Eppendorf centrifuge 5403, Germany) that were centrifuged at 4 °C, 503g for 15 minutes. The plasma was collected for red blood cell parameters, erythropoietin (EPO), Granulocyte Colony-stimulating factor analysis, (G-CSF), malondialdehyde (MDA), superoxide dismutase (SOD) and glutathione peroxidase (GPx) measurement.
Haematological Analysis
The erythrocyte markers namely, red blood cell count (RBC Count), haemoglobin (Hgb), mean cell volume (MCV), mean corpuscular haemoglobin (MCH) and mean corpuscular haemoglobin concentration (MCHC) and red blood cell distribution width (RDW) were measured together with the white blood cell and neutrophil counts Coulter Ac•T 5diff CP, USA).
Biochemical Analysis
Plasma EPO, G-CSF, SOD and GPx Analysis
Rat specific plasma EPO, G-CSF, SOD and GPx ELISA kits (Elabscience and Biotechnology, WuHan) were used following manufacturer’s instruction. Briefly, 100μL of standard or sample was added to the wells and incubated for 90 min at 37°C. The liquid was discarded, and 100μL of Biotinylated Detection Ab working solution was added to each well and incubated for 60 min at 37°C. Thereafter the plate was aspirated and washed 3 times. 100μL HRP conjugate working solution was added then the plate was incubated for 30 min at 37°C. The plate was then aspirated and washed 5 times. 90μL of Substrate Reagent was added and the plate was incubated for 15 min at 37°C. After-which 50μL of Stop Solution was added and the plate was read at 450nm immediately for the calculation of the results.
Malondialdehyde Measurement
Malondialdehyde was measured using an established protocol.10 Plasma was centrifuged at 400 g for 10 min. Thereafter, 400 mL of the plasma was supplemented with 400 mL 2% phosphoric acid and then separated into two glass tubes, each receiving equal volumes of the solution. Subsequently, 200 mL of 7% phosphoric acid was added into both glass tubes followed by the addition of 400 mL of thiobarbituric acid (TBA)/butylated hydroxytoluene (BHT) into one glass tube (sample test) and 400 mL of 3 mm hydrochloric acid (HCl) into the second glass tube (blank). To ensure an acidic pH of 1.5, 200 mL of 1M HCl was added to sample and blank test tubes. Both solutions were heated at 100 ◦C for 15 min and allowed to cool to room temperature. Butanol (1.5 mL) was added to the cooled solution; the sample was vortexed for 1 min to ensure rigorous mixing and allowed to settle until two phases could be distinguished. The butanol phase (top layer) was transferred to Eppendorf tubes and centrifuged at 13,200 g for 6 min. The samples were aliquoted into a 96-well microtiter plate in triplicate and the absorbance was read at 532 nm (reference 600 nm) on a BioTek mQuant spectrophotometer (Biotek, Johannesburg, South Africa). The absorbance from these wavelengths was used to calculate the concentration of MDA using Beer’s Law:

Statistical Analysis
All data were expressed as means ± standard error of means (SEM). A normality test was conducted after which the data was analysed using GraphPad Prism InStat Software (version 9.00, GraphPad Software, San Diego, California, USA). The terminal parameters were analysed using a one-way analysis of variance (ANOVA) followed by a Tukey Kramer post hoc test to determine the statistical differences between the control and experimental groups. Values of p<0.05 were considered statistically significant.
Results
Pup Birth Weights
Figure 1 shows the birth weights of pups born from normal control dams (PCON), L-NAME-induced preeclamptic dams (PPE), and dams induced with prediabetes (PPD) at the end of the study. The pups born from preeclamptic dams and prediabetic dams showed significantly (p < 0.05) lower birth weights in comparison to the pups born from normal dams. However, pups born from preeclamptic dams showed no significant change, but a slight decrease in birth weight when compared to pups born from prediabetes-induced animals.
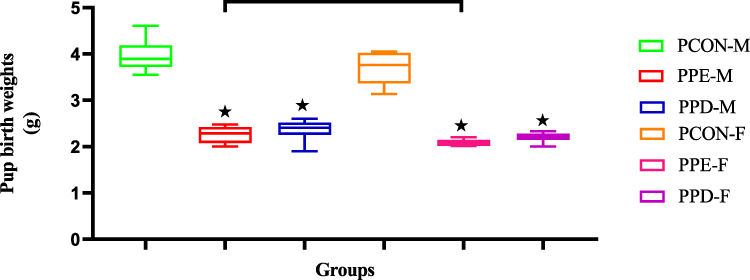 |
Figure 1 Shows the birth weights of pups born from normal control (PCON), preeclamptic (PPE), and prediabetic dams (PPD). Values are presented as means and standard error of means (SEM). *p < 0.05 by comparison with PCON and αp < 0.05 by comparison with PPE. |
Red Blood Cell Count (RBC) of Pups
Figure 2 shows the red blood cell count (RBC) of pups born from normal control dams (PCON), L-NAME induced preeclamptic dam (PPE), and dams induced with prediabetes (PPD) at the end of the study. The pups born from preeclamptic dams and prediabetic rats showed significantly (p < 0.05) increased RBC in comparison to the pups born from normal dams. As anticipated, pups born from preeclamptic dams showed significantly (p < 0.05) decreased RBC when compared to pups born from prediabetes-induced animals. Moreover, there was no significant change in the RBC of the male and female pups.
Haemoglobin (Hgb) Concentration of Pups
Figure 3 shows the haemoglobin (Hgb) of pups born from normal control dams (PCON), L-NAME-induced preeclamptic dam (PPE), and dams induced with prediabetes (PPD) at the end of the study. The pups born from preeclamptic dams and prediabetic dams showed significantly (p < 0.05) increased Hgb in comparison to pups born from normal dams. As expected, pups born from preeclamptic dams showed significantly (p < 0.05) decreased Hgb when compared to pups born from prediabetes-induced animals.
Red Blood Cell Parameters of Pups
Table 1 shows the red blood cell indices of pups born from normal control dams (PCON), L-NAME-induced preeclamptic dams (PPE), and dams induced with prediabetes (PPD). The pups born from preeclamptic dams showed a significant increase in the mean cell volume (MCV) and a decrease in (RDW) levels in comparison to the normal control animals. However, no significant changes were observed in the mean corpuscular haemoglobin (MCH) and mean corpuscular haemoglobin concentration (MCHC) concentrations. Furthermore, the pups born from prediabetic dams showed a significant (p < 0.05) decrease in MCV, MCH, and MCHC levels as well as a significant (p < 0.05) increase in RDW in comparison to the pups born from normal dams. Interestingly, pups born from preeclamptic dams showed significantly (p < 0.05) increased MCV, MCH, MCHC, and RDW levels when compared to pups born from prediabetic animals.
White Blood Cell (WBC), Neutrophil (NE), and Platelet (PLT) Counts of Pups
Table 2 shows the white blood cell (WBC), neutrophil (NE), and platelet counts (PLT) of pups born from normal control dams (PCON), L-NAME-induced preeclamptic dams (PPE) and dams induced with prediabetes (PPD). The pups born from preeclamptic dams and dams induced with prediabetes showed significantly (p < 0.05) decreased and increased WBC, respectively, as well as decreased and increased NE levels in comparison to the pups born from normal dams. Furthermore, the PLT of pups born from preeclamptic and prediabetic dams was significantly increased compared to pups born from normal dams. Interestingly, pups born from preeclamptic dams showed significantly (p < 0.05) decreased WBC, NE, and PLT when compared to pups born from prediabetes-induced animals.
Erythropoietin (EPO) Concentrations of Pups
Figure 4 shows the erythropoietin (EPO) of pups born from normal control dams (PCON), L-NAME-induced preeclamptic dam (PPE), and dams induced with prediabetes (PPD) at the end of the study. The pups born from preeclamptic dams and dams induced with prediabetes showed significantly (p < 0.05) increased EPO in comparison to the pups born from normal dams. As anticipated, pups born from preeclamptic dams showed significantly (p < 0.05) decreased EPO when compared to pups born from prediabetes-induced animals.
Granulocyte Colony-Stimulating Factor (G-CSF) Concentrations of Pups
Figure 5 shows the granulocyte colony-stimulating factor (G-CSF) of pups born from normal control dams (PCON), L-NAME-induced preeclamptic dam (PPE), and dams induced with prediabetes (PPD) at the end of the study. The pups born from preeclamptic dams and dams induced with prediabetes showed significantly (p < 0.05) decreased G-CSF in comparison to the pups born from normal dams. As anticipated, pups born from preeclamptic dams showed significantly (p < 0.05) increased G-CSF when compared to pups born from prediabetes-induced animals.
Malondialdehyde (MDA) Concentrations of Pups
Figure 6 shows the malondialdehyde (MDA) of pups born from normal control dams (PCON), L-NAME-induced preeclamptic dam (PPE), and dams induced with prediabetes (PPD) at the end of the study. The pups born from preeclamptic dams and prediabetic dams showed significantly (p < 0.05) increased MDA in comparison to the pups born from normal dams. As anticipated, pups born from preeclamptic dams showed significantly (p < 0.05) decreased MDA when compared to pups born from prediabetes-induced animals.
Superoxide Dismutase (SOD) Concentrations of Pups
Figure 7 shows the superoxide dismutase (SOD) of pups born from normal control dams (PCON), L-NAME-induced preeclamptic dam (PPE), and dams induced with prediabetes (PPD) at the end of the study. The pups born from preeclamptic dams and prediabetic dams showed significantly (p < 0.05) decreased SOD in comparison to the pups born from normal dams. Pups born from preeclamptic dams showed slightly increased SOD concentrations when compared to pups born from prediabetes-induced animals.
Glutathione Peroxidase (GPx) Concentrations of Pups
Figure 8 Glutathione Peroxidase (GPx) of pups born from normal control dams (PCON), L-NAME-induced preeclamptic dam (PPE), and dams induced with prediabetes (PPD) at the end of the study. The pups born from preeclamptic dams and prediabetic dams showed significantly (p < 0.05) decreased GPx in comparison to pups born from normal dams. Pups born from preeclamptic dams showed slightly increased GPx concentrations when compared to pups born from prediabetes-induced animals.
Discussion
During pregnancy, various maternal adaptations occur which modify biological systems to benefit the mother and child.12 These adaptations alter respiration, metabolism, and cardiovascular systems to maintain a healthy balance between the mother and the foetus, not hindering foetal growth and development.13 The placenta is the main organ through which foetal oxygenation, nutrition, and immunity are achieved.13 It is a critical functional unit of pregnancy that ensures the well-being of the mother and fetus.14 Therefore, it is imperative for maternal adaptations which favour proper placental development to occur.15 These adaptations are critical in pregnancy-related systems as well as in vascular modulation that affects foetal perfusion.16,17 Adequate foetal perfusion plays a critical role in haematological pathways, mainly erythropoiesis, leukopoiesis, and thrombopoiesis, which are essential for foetal survival into adulthood.18,19 Furthermore, abnormal placental perfusion has been positively correlated with low infant birth weight.20 Placental hypoxia results in restricted oxygen and nutrient supply, altering foetal metabolism and growth resulting in low infant birth weight and intrauterine growth restriction (IUGR).20 IUGR has major effects on the developmental programming and haematological pathways and blood cellular components.20
Studies have shown that in pregnancy, infant birth weights are strictly monitored as a predictor for infant developmental milestones, as infant birth weights give insight to the health and developmental of the infant from neonatal age until adulthood.20,21 Infants born from pregnancies where preeclampsia and pregestational diabetes, like T2D, are reported have been shown to exhibit low infant birth weights.21,22 Low birth weights are a significant risk factor for poor foetal health outcomes in short- and long-term development.23,24 In the present study, Figure 1 displays the infant birth weights of both male and female preeclamptic pups (PPE) and prediabetic pups (PPD). The birth weights of the PPE and PPD groups were significantly lower than the offspring born from the control group. Furthermore, there was a significant increase in the infant birth weight of offspring born in the PPE group compared with the PPD group.25,26 This aligns with the existing literature suggesting that prenatal hypoxia in pregestational and preeclamptic pregnancies leads to IUGR which causes low birth weights.
The present study also divulges how PPD follows the same birth weight trend as other pregestational pregnancies. Although the mechanisms are not well elucidated, it is assumed that prenatal hypoxia impedes foetal growth via placental insufficiency, which impairs foetal oxygenation and nutrition.27,28 This is because, in healthy pregnancies, maternal spiral artery remodelling occurs via cytotrophoblast invasion.29,30 This process involves the conversion of the maternal spiral arteries, which are initially narrow and resistant vessels, into wider and more accommodating vessels which are capable of providing adequate placental perfusion to sustain foetal growth via oxygenation and nutrition.31,32 We speculate that in PPE, T2D as well as in the PPD group, the invasion of cytotrophoblast into the maternal spiral artery is unsuccessful.33 This leads to placental insufficiency resulting in foetal hypoxia and lowers both oxygenation and nutrition of the fetus resulting in lower birth weight and IUGR.34 Furthermore, the lower infant birth weight observed in the PPD group compared to the PE can be attributed to hyperglycemia added insult which causes placentomegaly and increases the severity of IUGR in pregestational pregnancies.35,36 Moreover, the significantly higher birth weight observed in the male compared to the female group was attributed to inherent physiological and genetic differences and did not indicate more severe foetal outcomes in females.37,38 Rather, studies show that females tend to experience better health outcomes, and this is largely due to the influence of sexual hormones such as oestrogen, which confer cellular protection.39,40
Moreover, low birth weight and IUGR due to prenatal hypoxia have also been implicated in the disruption of foetal blood cellular development in which erythrocyte production is altered.41–43 For effective maternofoetal oxygen transport to occur, foetal erythroid cells are formed by the embryo in the first week of gestation.44,45 Once matured, these erythrocytes play an imperative role in ensuring oxygen delivery to the newborn during gestation and postpartum.12,13 Erythrocyte markers such as RBC, Hgb, MCV, MCH, MCHC, and RDW denote the maturity and health of erythrocytes and are regulated within a narrow range.14,46 Deviations from this range can have detrimental effects on erythrocyte structure and function.3,47 In this study, Figures 2 and 3 show significantly elevated RBC and Hgb concentrations in PPE and PPD compared to normal control offspring. There was also a significant decrease in the RBC and Hgb levels of the PPE group in comparison to the PPD group. We speculate that changes in RBC, MCH, MCHC, and Hgb concentrations are a result of the prenatal hypoxia observed in the mothers of PPE and PPD.48 Hypoxia significantly increases erythropoietin (EPO) in PPE and PPD offspring in comparison to normal controls (Figure 4).49,50 This causes the rate of erythropoiesis in the liver and bone marrow of the foetus to surge, which expands the red cell mass resulting in elevated nucleated red blood cells/erythroblasts and thus increases RBC MCH, MCHC and Hgb levels.49,50 Moreover, the higher RBC and Hgb levels in the PPD group compared to the PPE group are a result of hyperglycemia which causes foetal polycythemia by stimulating excess erythropoietin production in the fetus.49,50 The elevated blood glucose concentration increases foetal blood viscosity and reduces blood flow, which can further contribute to the development of polycythemia.50 Also, the observed disparities in RBC and Hgb levels between male and female offspring are due to inherent differences between the two sexes.50 The main difference was significantly higher testosterone levels in males, which have been shown to stimulate erythropoiesis increasing erythrocyte production.50
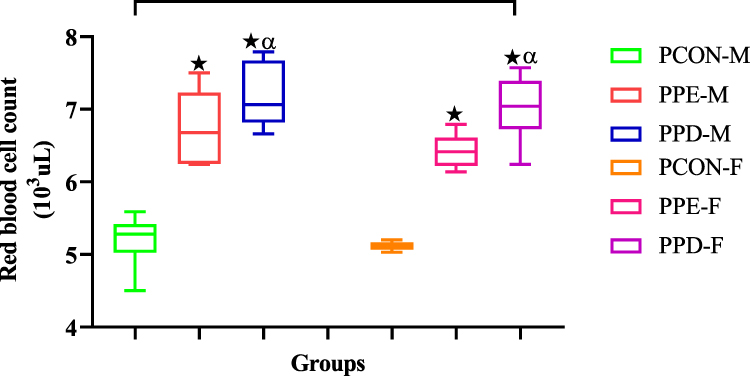 |
Figure 2 Shows the red blood cell counts of pups born from normal control (PCON), preeclamptic (PPE), and prediabetic dams (PPD). Values are presented as means and standard error of means (SEM). *p < 0.05 by comparison with PCON and αp < 0.05 by comparison with PPE. |
 |
Figure 3 Shows the haemoglobin (Hgb) of pups born from normal control (PCON), preeclamptic (PPE), and prediabetic dams (PPD). Values are presented as means and standard error of means (SEM). *p < 0.05 by comparison with PCON and αp < 0.05 by comparison with PPE. |
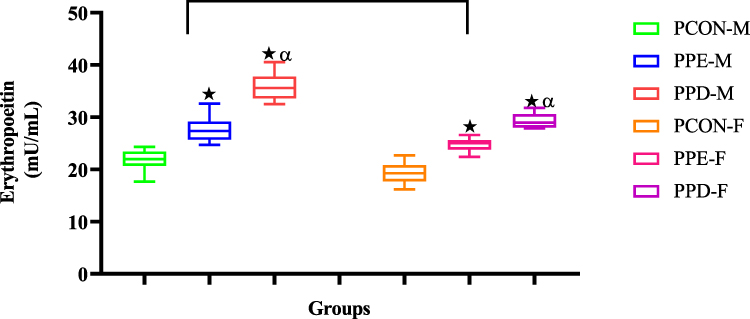 |
Figure 4 Shows the erythropoietin (EPO) of pups born from normal control (PCON), preeclamptic (PPE), and prediabetic dams (PPD). Values are presented as means and standard error of means (SEM). *p < 0.05 by comparison with PCON and αp < 0.05 by comparison with PPE. |
Oxidative stress is important in pregnancy for foetal blood cell production because it plays a critical role in the development of the foetal circulatory system. During pregnancy, the foetal blood cell production is a highly dynamic and complex process that requires a delicate balance of reactive oxygen species (ROS) and antioxidant defenses. However, excessive oxidative stress during pregnancy can have detrimental effects on foetal blood cell production. It can lead to the destruction of red blood cells and impair the development of the circulatory system, resulting in conditions such as fetal anaemia or intrauterine growth restriction. Therefore, maintaining a balance of oxidative stress is crucial for ensuring optimal foetal blood cell production and overall development.50 In this study, Figures 6–8 show changes in the antioxidant profile with elevated malondialdehyde (MDA) levels and decreased antioxidant enzymes superoxide dismutase (SOD) and glutathione peroxidase (GPx) observed in PPE and PPD groups in comparison to normal control. Furthermore, an increase in MDA is accompanied by further decreased antioxidant enzyme levels in PPD compared to the PPE group. This is due to prenatal hypoxia which favours reactive oxygen species (ROS) production overwhelming the antioxidant capacity.50 ROS such as peroxide (O2-) and hydrogen peroxide (H2O2) cross the placenta and oxidatively modify antioxidant enzymes, superoxide dismutase (SOD) and glutathione peroxidase (GPx), which decrease their availability in foetal circulation.51 ROS also induce DNA damage to erythroid progenitor cells leading to cell cycle arrest and increased immature erythroblast/ nucleated red blood cells (nRBC) in the circulation.52,53 Moreover, the antioxidant levels in the female offspring were higher than in the male offspring in all groups. This is due to the role of the sexual hormone estrogen in the form of 17 beta-estradiol, which influences the production of manganese superoxide dismutase (MnSOD) and extracellular superoxide dismutase (ecSOD) expression and activity providing vasoprotective effects to the female offspring.54
Furthermore, intrauterine growth restriction (IUGR) disrupts the nutritional supply to the fetus, leading to limited availability of essential components such as folate, vitamin B12, and iron.55 These components play a vital role in erythrocyte production, growth, and maturation, and their deficiency results in erythrocyte structural and functional abnormalities.56,57 Folate and vitamin B12 deficiencies lead to abnormal erythrocyte shape and size, impairing their oxygen-carrying capacity, while iron deficiency affects haemoglobin production and erythrocyte size, resulting in reduced circulating oxygen.58 In this study, Table 1 indicates a significant elevation in mean corpuscular volume (MCV) and red cell distribution width (RDW) concentrations in PPE compared to the normal control group. This effect may be due to the decreased supply of vitamin B12 and folate, leading to an increase in erythrocyte size (MCV) and size variation (RDW) in the circulation, leading to foetal megaloblastic anaemia.58,59 Conversely, Table 1 shows significantly decreased MCV and elevated RDW in PPD in comparison to the normal control group. This effect could be ascribed to hyperglycemia-induced prenatal hypoxia, and IUGR which causes iron deficiency anemia, resulting in hemoconcentration, decreased erythrocyte size (MCV) and size variation in circulation (RDW).59,60 Furthermore, the changes between PPE and PPD groups in Table 1 illustrate that offspring with PPE exhibit significantly higher MCV and RDW values than those from the PPD group. This effect can be a result of hyperglycaemia suppressing hepcidin expression. Low hepcidin levels are an early marker for iron deficiency anaemia.60 Low hepcidin levels have been shown to indicate exhausted iron stores, the lack of iron affects red blood cell structure and function leading to iron deficiency anaemia observed in the PPD group.60,61 Moreover, the significantly higher MCV and RDW values observed in males in comparison to females could be due to the influences of sexual hormones, testosterone, and oestrogen.62 Research has shown that testosterone increases erythrocyte size and haemoglobin production, which results in higher MCV and RDW values.62,63 Whereas oestrogen decreases erythrocyte size and increases heme production, which decreases the haemoglobin contained in each cell leading to lower MCV and RDW values.63
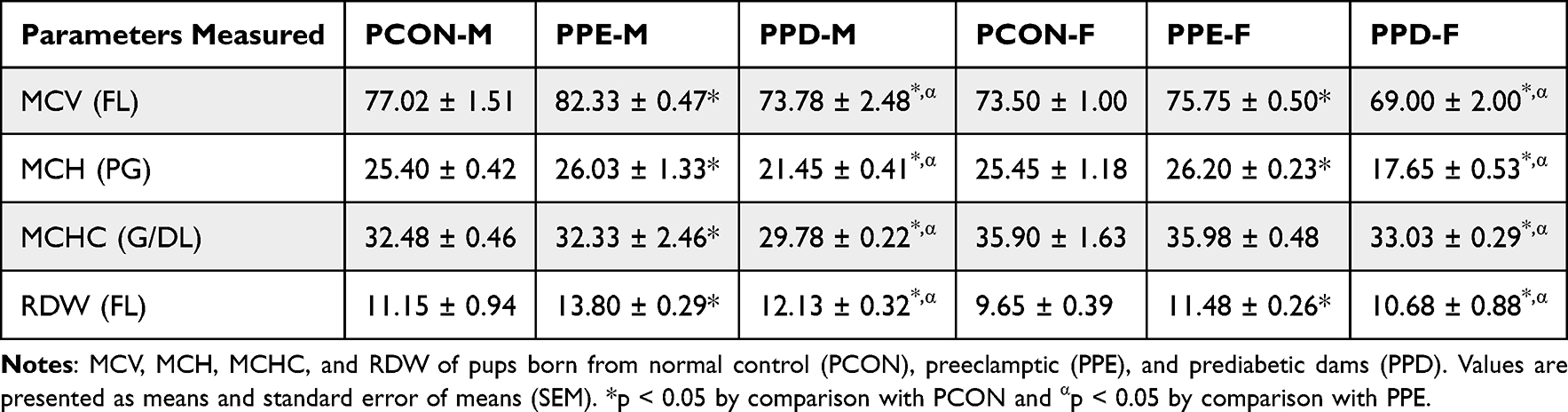 |
Table 1 Red Blood Cell Parameters of Pups |
Moreover, prenatal hypoxia can induce a decrement in the production of granulocyte colony-stimulating factor (G-CSF), which is a cytokine that holds a crucial role in the generation and differentiation of white blood cells, particularly neutrophils.64 Neutrophils are essential for fighting infections, especially in newborns as they have an immature immune system making them more susceptible to infection.64,65 In this study, Figure 5 and Table 2 show the G-CSF levels, neutrophil, and platelet counts in the PPE and PPD groups, which exhibited lower G-CSF, neutrophil and platelet levels than the offspring born from the control group. This could be in part due to prenatal hypoxia and IUGR that impairs foetal liver function, decreasing the production of GM-CSF.65 Low G-CSF levels contribute to the failure of downstream myeloid differentiation resulting in subsequent neutropenia and thrombocytopenia.65 Moreover, there was a significant increase in the G-CSF levels of offspring born in the PPE group in comparison to the PPD group. This could be attributed to severe or prolonged prenatal hypoxia brought about by the hyperglycaemia observed, which causes the dysregulation of several transcription factors and signalling pathways involved in G-CSF synthesis and secretion.66 Hyperglycaemia has been shown to activate the nuclear factor-kappa B(NF-KB) signalling pathway, which suppresses G-CSF expression.66 ROS have also been shown to further impair G-CSF expression by altering DNA sequences essential for G-sCSF gene expression, which lowers G-CSF levels, therefore disrupting neutrophil and platelet differentiation.66,67 The resulting decrease in neutrophil and platelet counts has detrimental consequences as it leaves the foetus vulnerable to infection and at an increased risk of bleeding or haemorrhage.67 Moreover, the G-CSF levels in the female offspring were significantly lower than in the male offspring in all groups.67 This is attributed to the difference in gene expression between sexes.67,68 Studies have shown that the promoter region of the G-CSF gene has oestrogen response elements (ERE) which are specific genetic sequences that are activated by oestrogen.68 Oestrogen binds to ERE and interferes with G-CSF transcription, subsequently decreasing the production and secretion of G-CSF resulting in neutropenia and thrombocytopenia.69 This makes female offspring more susceptible to immune dysfunction and haemorrhage and is further worsened by female offspring born with hyperglycaemic pregnancies.70
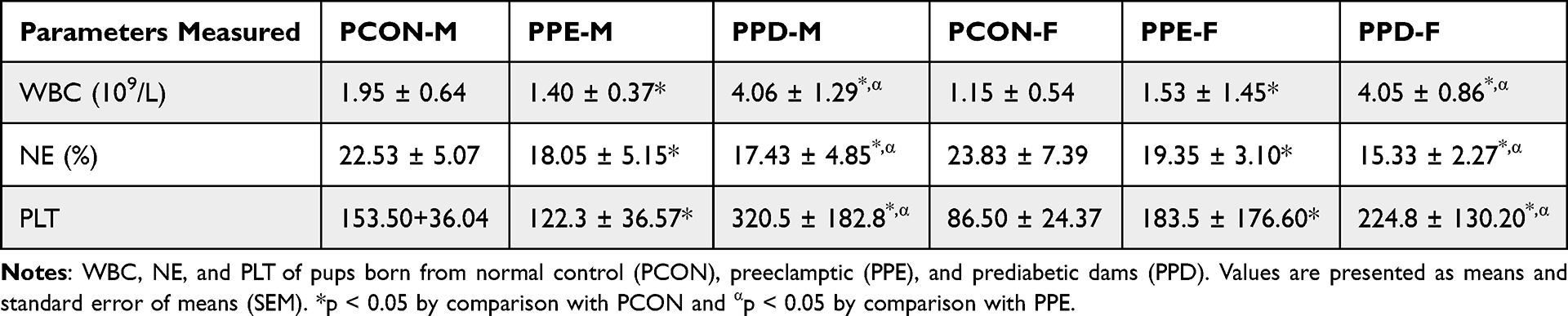 |
Table 2 White Blood Cell (WBC), Neutrophil (NE), and Platelet (PLT) Counts of Pups |
 |
Figure 5 Shows the granulocyte colony-stimulating factor (G-CSF) of pups born from normal control (PCON), preeclamptic (PPE), and prediabetic dams (PPD). Values are presented as means and standard error of means (SEM). *p < 0.05 by comparison with PCON and αp < 0.05 by comparison with PPE. |
 |
Figure 6 Shows the malondialdehyde (MDA) of pups born from normal control (PCON), preeclamptic (PPE), and prediabetic dams (PPD). Values are presented as means and standard error of means (SEM). *p < 0.05 by comparison with PCON and αp < 0.05 by comparison with PPE. |
 |
Figure 7 Shows the superoxide dismutase (SOD) of pups born from normal control (PCON), preeclamptic (PPE), and prediabetic dams (PPD). Values are presented as means and standard error of means (SEM). *p < 0.05 by comparison with PCON. |
 |
Figure 8 Shows the glutathione peroxidase (GPx) of pups born from normal control (PCON), preeclamptic (PPE), and prediabetic dams (PPD). Values are presented as means and standard error of means (SEM). *p < 0.05 by comparison with PCON. |
Conclusion
Preeclamptic and pregestational pregnancies, particularly those of T2D, have debilitating effects on both mothers and children and pose a financial burden to the healthcare industry.52,71,72 Research on these conditions can help identify effective prevention and treatment measures to improve outcomes for the mother and children.54 Prediabetes is a precursor of T2D and in this study, PD has been potentiated as a risk factor for PE.73 This study has shown that prediabetes can lead to haematological complications in both male and female offspring as supported by changes to erythrocyte parameters, RBC, Hgb, MCV and RDW as well as changes to WBC, neutrophils and platelet counts. These haematological complications give rise to cardiovascular disease and increase the risk of incurring infections.74 Therefore, early screening and diagnosis of prediabetes can offer an opportunity for early intervention and prevent the onset of type 2 diabetes, gestational diabetes mellitus, as well as preeclampsia.75 Moreover, research on therapies tailored for both males and females is necessary due to the notable differences in hormonal profiles, metabolic rates, and other genetic factors.75 This will eliminate gender bias in medical research and allow for safe and effective medical care.75
Data Sharing Statement
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Ethics Approval and Consent to Participate
All animal experimentation was reviewed and approved by the Animal Research Ethics Committee of the University of KwaZulu-Natal (AREC/054/017D). The animals were monitored for pain, discomfort and distress using the criteria listed in the university’s Animal Research Ethics Committee’s humane endpoint document.
Acknowledgments
The authors would like to thank biomedical research unit personnel for their technical assistance. This study was partly funded by NRF South Africa and the University of KwaZulu- Natal, college of health sciences.
Author Contributions
All authors made substantial contributions to conception and design, acquisition of data, or analysis and interpretation of data; took part in drafting the article or revising it critically for important intellectual content; gave final approval of the version to be published; and agree to be accountable for all aspects of the work.
Funding
NRF Funded- Funding was used to acquire animals (Sprague Dawley rats), equipment and machinery to perform analytical techniques (Blood analysis and ELISA’s).
Disclosure
The authors declare that they have no competing interests.
References
1. Yang H, Zhu C, Ma Q, et al. Variations of blood cells in prediction of gestational diabetes mellitus. J Perinat Med. 2015;43:89–93. doi:10.1515/jpm-2014-0007
2. Franco Robert S. The measurement and importance of red cell survival. Am J Hematol. 2009;84:109–114. doi:10.1002/ajh.21298
3. Backes CH, Markham K, Moorehead P, Cordero L, Nankervis CA, Giannone PJ. Maternal preeclampsia and neonatal outcomes. J Pregnancy. 2011;2011:01–07. doi:10.1155/2011/214365
4. Shokry E, Marchioro L, Uhl O, et al. Impact of maternal BMI and gestational diabetes mellitus on maternal and cord blood metabolome: results from the PREOBE cohort study. Acta Diabetol. 2019;56(4):421–430. doi:10.1007/s00592-019-01291-z
5. Krishnan S, Aston CE, Fields DA, Teague AM, Lyons TJ, Chernausek SD. Bone mass accrual in first six months of life: impact of maternal diabetes, infant adiposity, and cord blood adipokines. Calcif Tissue Int. 2022;111(3):1–8.
6. Lee YJ, Shin YH, Kim JK, et al. Metabolic syndrome and its association with white blood cell count in children and adolescents in Korea: the 2005 Korean National Health and Nutrition Examination Survey. Nutr Metab Cardiovasc Dis. 2010;20:165–172. doi:10.1016/j.numecd.2009.03.017
7. Erdogan S, Ozdemir O, Dogan HO, et al. Liver enzymes, mean platelet volume, and red cell distribution width in gestational diabetes. Turk J Med Sci. 2014;44:121–125. doi:10.3906/sag-1301-41
8. English E, Idris I, Smith G, et al. The effect of anaemia and abnormalities of erythrocyte indices on HbA1c analysis: a systematic review. Diabetologia. 2015;58:1409–1421. doi:10.1007/s00125-015-3599-3
9. Stevens gretchen A, Finucane Mariel M, De-Regil Luz M, et al. Global, regional, and national trends in haemoglobin concentration and prevalence of total and severe anaemia in children and pregnant and non-pregnant women for 1995–2011: a systematic analysis of population-representative data. Lancet Glob Heal. 2013;1:16–25. doi:10.1016/S2214-109X(13)70001-9
10. Luvuno M, Mabandla M, Khathi A. Voluntary ingestion of a high-fat high-carbohydrate diet: a model for prediabetes. Ponte Int Sci Res J. 2018;74.
11. Al-Bahadily A-K, Al-Omrani A, Mohammed M. The effect of pregnancy induced hypertension on complete blood count of newborn. J Ped Perspect. 2017;5(9):5667–5676.
12. Takahashi M, Makino S, Oguma K, et al. Fetal growth restriction as the initial finding of preeclampsia is a clinical predictor of maternal and neonatal prognoses: a single-center retrospective study. BMC Pregnancy Childbirth. 2021;21:1–8. doi:10.1186/s12884-021-04152-2
13. Sharifzadeh Mahalati F, Moeini Chaghervand M, Hashemi Dizaji S, Chamani M. Evaluation of the relationship between prenatal Doppler findings and hematological profile in neonates with intrauterine growth restriction at 32 to 36 weeks of gestation. J Obstet Gynecol Cancer Res. 2022;4(2):81–85.
14. Palis J, Yoder MC. Yolk-Sac hematopoiesis: the first blood cells of mouse and man. Exp Hematol. 2001;29(8):927–936. doi:10.1016/S0301-472X(01)00669-5
15. Prefumo F, Fichera A, Fratelli N, Sartori E. Fetal anemia: diagnosis and management. Best Pract Res Clin Obstet Gynaecol. 2019;58:2–14. doi:10.1016/j.bpobgyn.2019.01.001
16. Garzon S, Cacciato PM, Certelli C, Salvaggio C, Magliarditi M, Rizzo G. Iron deficiency anemia in pregnancy: novel approaches for an old problem. Oman Med Journal. 2020;35(5):e166. doi:10.5001/omj.2020.108
17. Moore LG. Fetal growth restriction and maternal oxygen transport during high altitude pregnancy. High Alt Med Biol. 2003;4(2):141–156. doi:10.1089/152702903322022767
18. Kalteren WS, Bos AF, Bergman KA, van Oeveren W, Hulscher JB, Kooi EM. The short-term effects of rbc transfusions on intestinal injury in preterm infants. Pediatr Res. 2022;93:1–7. doi:10.1038/s41390-022-02168-8
19. Chien S. Red cell deformability and its relevance to blood flow. Annu Rev Physiol. 1987;49(1):177–192. doi:10.1146/annurev.ph.49.030187.001141
20. McMahon TJ. Red blood cell deformability, vasoactive mediators, and adhesion. Front Physiol. 2019;10:1417. doi:10.3389/fphys.2019.01417
21. Brun J-F, Varlet-Marie E, Myzia J, Raynaud de Mauverger E, Pretorius E. Metabolic influences modulating erythrocyte deformability and eryptosis. Metabolites. 2022;12(1):4. doi:10.3390/metabo12010004
22. Zegarra RR, Dall’Asta A, Ghi T. Mechanisms of fetal adaptation to chronic hypoxia following placental insufficiency: a review. Fetal Diagn Ther. 2022;49(5–6):279–292. doi:10.1159/000525717
23. Bahr TM, Albertine KH, Christensen RD, et al. Nucleated red blood cell emergence-time in newborn lambs following a dose of darbepoetin alfa. Curr Pediatr Rev. 2023;19(4):425–428. doi:10.2174/1573396319666221219153902
24. Bahr TM, Ohls RK, Baserga MC, Lawrence SM, Winter SL, Christensen RD. Implications of an elevated nucleated red blood cell count in neonates with moderate to severe hypoxic-ischemic encephalopathy. J Pediatr. 2022;246:12–8.e2. doi:10.1016/j.jpeds.2022.04.015
25. Chatzakis C, Sotiriadis A, Gerede A, Dinas K, Nicolaides KH. Fetal Complications. In: Comprehensive Clinical Approach to Diabetes During Pregnancy. Springer; 2022:291–310.
26. Calvo C, Capellini-Suárez A, Hernandez A. Diabetic Fetopathy. Med Clin Res Open Access. 2021;2(4):1–7.
27. Chen Y-H, Chen W-Y, Chang C-Y, et al. Association between maternal factors and fetal macrosomia in full-term singleton births. J Chin Med Assoc. 2023;86(3):324–329. doi:10.1097/JCMA.0000000000000871
28. Desta SA, Damte A, Hailu T. Maternal factors associated with low birth weight in public hospitals of Mekelle city, Ethiopia: a case-control study. Ital J Pediatr. 2020;46:1–9. doi:10.1186/s13052-020-00890-9
29. KC A, Basel PL, Singh S. Low birth weight and its associated risk factors: health facility-based case-control study. PLoS One. 2020;15(6):e0234907. doi:10.1371/journal.pone.0234907
30. Malaza N, Masete M, Adam S, Dias S, Nyawo T, Pheiffer C. A systematic review to compare adverse pregnancy outcomes in women with pregestational diabetes and gestational diabetes. Int J Environ Res Public Health. 2022;19(17):10846. doi:10.3390/ijerph191710846
31. Bar J, Weiner E, Levy M, Gilboa Y. The thrifty phenotype hypothesis: the association between ultrasound and Doppler studies in fetal growth restriction and the development of adult disease. Am j Obstet Gynecol MFM. 2021;3(6):100473. doi:10.1016/j.ajogmf.2021.100473
32. Yang L, Feng L, Huang L, et al. Maternal factors for intrauterine growth retardation: systematic review and meta-analysis of observational studies. Reprod Sci. 2023:1–9.
33. Zhou H, Zhao C, Wang P, Yang W, Zhu H, Zhang S. Regulators involved in trophoblast syncytialization in the placenta of intrauterine growth restriction. Front Endocrinol. 2023;14.
34. Haram K, Mortensen JH, Myking O, Roald B, Magann EF, Morrison JC. Early development of the human placenta and pregnancy complications. J Matern Fetal Neonatal Med. 2020;33(20):3538–3545. doi:10.1080/14767058.2019.1578745
35. Sato Y. Endovascular trophoblast and spiral artery remodeling. Mol Cell Endocrinol. 2020;503:110699. doi:10.1016/j.mce.2019.110699
36. Quasimi H, Ray A, Iqbal Alam M. The role of nitric oxide in the etiopathogenesis of preeclampsia. In: Nitric Oxide: From Research to Therapeutics. Springer; 2023:391–409.
37. Phoswa WN, Khaliq OP. The role of oxidative stress in hypertensive disorders of pregnancy (Preeclampsia, Gestational Hypertension) and metabolic disorder of pregnancy (Gestational Diabetes Mellitus). Oxid Med Cell Longev. 2021;2021:1–10. doi:10.1155/2021/5581570
38. Sergi CM, Sergi CM. Placenta, abnormal conception, and prematurity. Pathol Child Adolesc. 2020:1409–1569.
39. Bhojani PS. Smart Study Series: Obstetrics & Gynecology. Elsevier Health Sciences; 2020.
40. Hoftiezer L, Hof MH, Dijs-Elsinga J, Hogeveen M, Hukkelhoven CW, van Lingen RA. From population reference to national standard: new and improved birthweight charts. Am J Clin Exp Obstet Gynecol. 2019;220(4):383.e1–e17. doi:10.1016/j.ajog.2018.12.023
41. de Mendonça ELSS, de Lima Macêna M, Bueno NB, de Oliveira ACM, Mello CS. Premature birth, low birth weight, small for gestational age and chronic non-communicable diseases in adult life: a systematic review with meta-analysis. Early Human Dev. 2020;149:105154. doi:10.1016/j.earlhumdev.2020.105154
42. Klein SL, Morgan R. The impact of sex and gender on immunotherapy outcomes. Biol Sex Differ. 2020;11(1):1–10. doi:10.1186/s13293-020-00301-y
43. Bath KG. Synthesizing views to understand sex differences in response to early life adversity. Trends Neurosci. 2020;43(5):300–310. doi:10.1016/j.tins.2020.02.004
44. Lateef OM, Akintubosun MO. Sleep and reproductive health. J Circadian Rhythms. 2020;18.
45. Sciatti E, Orabona R. A window of opportunity on cardiovascular prevention: pre-eclampsia and fetal growth restriction. Eur J Prev Cardiol. 2021;28(16):e17–e8. doi:10.1177/2047487320925646
46. Catarino C, Rebelo I, Belo L, et al. Erythrocyte damage/aging/removal are enhanched in both mother and foetus, in preeclampsia. Actas Bioq. 2008;9:63–67.
47. Bolat A, Gursel O, Kurekci E, Atay A, Ozcan O. Blood parameters changes in cord blood of newborns of hypertensive mothers. Eur J Pediatr. 2013;172:1501–1509. doi:10.1007/s00431-013-2072-x
48. Goften EN, Capewel V, Natale R, Gratton RJ. Obstetrical intervention rates and maternal and neonatal outcomes of women with gestational hypertension. AM J Obstet Gynecol. 2001;185:798–803. doi:10.1067/mob.2001.117314
49. Ed Kurlat I, Sola A. Neonatal polycythemia in appropriately grown infants of hypertensive mothers. Acta Paediatr. 1992;81:662–664. doi:10.1111/j.1651-2227.1992.tb12328.x
50. Saarinen UM, Siimes MA. Developmental changes in red blood cells and indices of infants after exclusion of iron deficiency by laboratory criteria and continuous iron supplementation. J Pediatr. 1978;92:412–416. doi:10.1016/S0022-3476(78)80429-6
51. Shivkumar S, Bhat BV, Badhe BA. Effect of pregnancy induced hypertension on mothers and their babies. Indian J Pediatr. 2007;74(7):623–625. doi:10.1007/s12098-007-0110-2
52. Salaia CM, Ghidini A, Pezzullo JC, Rosenkrantz TS. Early neonatal nucleated erythrocyte counts in preterm deliveries: clinical and pathologic correlations. J Soc Gynecol Invest. 1997;4(3):138–143. doi:10.1177/107155769700400305
53. Hebbar S, Misha M, Rai L. Significance of maternal and cord blood nucleated red blood cell count in pregnancies complicated by preeclampsia. J Pregnancy. 2014;2014:01–07. doi:10.1155/2014/496416
54. Cristina C, Irene R, Luís B. Fetal and maternal angiogenic/ anti-angiogenic factors in normal and preeclamptic pregnancy. Growth Fact. 2009;27(6):345–351. doi:10.3109/08977190903184670
55. Troeger C, Holzgreve W, Cadewig A, Zhang XY, Hahn S. Examination of maternal plasma erythropoietin and activin A concentrations with regard to circulatory erythroblast levels in normal and pre-eclamptic pregnancies. Fetal Diagn Ther. 2006;21:56–60. doi:10.1159/000089068
56. Julaksiz H, Fein E, Redecker P, Stremmel W, Adler G, Cetin Y. Pancreatic beta-cells express hepcidin, an iron-uptake regulatory peptide. J Endocrinol. 2008;197:241–249.
57. Aigner E, Felder TK, Oberkofler H, et al. Glucose acts as a regulator of serum iron by increasing serum hepcidin concentrations. J Nutr Biochem. 2013;24:24112–24117. doi:10.1016/j.jnutbio.2012.02.017
58. Backe MB, Moen IW, Ellervik C, Hansen JB, Mandrup-Poulsen T. Iron regulation of pancreatic beta-cell functions and oxidative stress. Annu Rev Nutr. 2016;36:241–273. doi:10.1146/annurev-nutr-071715-050939
59. Fernandez-Real JM, McClain D. Manco M.Mechanisms linking glucose homeostasis and iron metabolism toward the onset and progression of Type 2 diabetes. Diabetes Care. 2015;38:382169–382176. doi:10.2337/dc14-3082
60. Aregbesola A, Voutilainen S, Virtanen JK, Aregbesola A, Tuomainen TP. Serum hepcidin concentrations and type 2 diabetes. World J Diabetes. 2015;6:978–982. doi:10.4239/wjd.v6.i7.978
61. Simcox JA, McClain DA. Iron and diabetes risk. Cell Metab. 2013;17:329–341. doi:10.1016/j.cmet.2013.02.007
62. Semerad CL, Liu F, Gregory AD, Stumpf K, Link DC. G-CSF is an essential regulator of neutrophil trafficking from the bone marrow to the blood. Immunity. 2002;17:413–423. doi:10.1016/S1074-7613(02)00424-7
63. McDermott DH, Liu Q, Velez D, et al. A Phase 1 clinical trial of long-term, low-dose treatment of WHIM syndrome with the CXCR4 antagonist plerixafor. Blood. 2014;123:2308–2316. doi:10.1182/blood-2013-09-527226
64. Greenbaum AM, Link DC. Mechanisms of G-CSF-mediated hematopoietic stem and progenitor mobilization. Leukemia. 2011;25:211–217. doi:10.1038/leu.2010.248
65. To LB, Levesque JP, Herbert KE. How I treat patients who mobilize hematopoietic stem cells poorly. Blood. 2011;118:4530–4540. doi:10.1182/blood-2011-06-318220
66. Scheinberg P, Young NS. How I treat acquired aplastic anemia. Blood. 2012;120:1185–1196. doi:10.1182/blood-2011-12-274019
67. Tichelli A, Schrezenmeier H, Socie G, et al. A randomized controlled study in patients with newly diagnosed severe aplastic anemia receiving antithymocyte globulin (ATG), cyclosporine, with or without G-CSF: a study of the SAA Working Party of the European Group for Blood and Marrow Transplantation. Blood. 2011;117:4434–4441. doi:10.1182/blood-2010-08-304071
68. Jeng MR, Naidu PE, Rieman MD, et al. Granulocyte-macrophage colony stimulating factor and immunosuppression in the treatment of pediatric acquired severe aplastic anemia. Pediatr Blood Cancer. 2005;45:170–175. doi:10.1002/pbc.20278
69. Kojima S. Use of hematopoietic growth factors for treatment of aplastic anemia. Bone Marrow Transplant. 1996;18(Suppl 3):S36–S38.
70. Rätsep MT, Hickman AF, Maser B, et al. Impact of preeclampsia on cognitive function in the offspring. Behav Brain Res. 2016;302:175–181. doi:10.1016/j.bbr.2016.01.030
71. Pennington KA, Schlitt JM, Jackson DL, Schulz LC, Schust DJ. Preeclampsia: multiple approaches for a multifactorial disease. Dis Models Mech. 2012;5(1):9–18. doi:10.1242/dmm.008516
72. Khalil RA, Granger JP. Vascular mechanisms of increased arterial pressure in preeclampsia: lessons from animal models. Am J Physiol Regul Integr Comp Physiol. 2002;283(1).
73. Al-Gazali L, Valian V, Barnes B, et al. Scientists of the world speak up for equality. Nature. 2013;495:35–38.
74. Bargh JA, Chartrand TL. The mind in the middle: a practical guide to priming and automaticity research. In: Reis HT, Judd CM, editors. Handbook of Research Methods in Social and Personality Psychology. New York: Cambridge University Press; 2000:253–285.
75. Case KA. Raising male privilege awareness and reducing sexism: an evaluation of diversity courses. Psychol Women Q. 2007;31:426–435. doi:10.1111/j.1471-6402.2007.00391.x
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.