Back to Journals » OncoTargets and Therapy » Volume 13
Cervical Cancer: Emerging Immune Landscape and Treatment
Received 24 May 2020
Accepted for publication 13 July 2020
Published 12 August 2020 Volume 2020:13 Pages 8037—8047
DOI https://doi.org/10.2147/OTT.S264312
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. Nicola Silvestris
Luopei Guo,1 Keqin Hua1,2
1Obstetrics and Gynecology Hospital of Fudan University, Shanghai 200011, People’s Republic of China; 2Shanghai Key Laboratory of Female Reproductive Endocrine Related Diseases, Shanghai 200011, People’s Republic of China
Correspondence: Keqin Hua
Obstetrics and Gynecology Hospital of Fudan University, 419 Fangxie Road, Shanghai 200011, People’s Republic of China
Email [email protected]
Abstract: Immune cells are essential for defending the body’s balance and have increasingly been implicated in controlling tumor growth. In cervical cancer (CC), the immune landscape is extensively connected with human papillomavirus (HPV) status. Recent insights from studies have revealed that as a result of infection with HPV, immune cell populations such as lymphocytes or monocytes change during carcinogenesis. Immune therapy, in particular checkpoint inhibitors, those targeting PD-1 or PD-L1, has shown promising efficacy. This article reviews the immune landscape and immunotherapy of CC.
Keywords: immune, TIL, macrophage, immune therapy, checkpoint blockade
Introduction
Cervical cancer (CC) is one of the most common gynecological malignancies worldwide, with nearly 570,000 new CC cases and more than 311,000 deaths every year.1 In 2019, there were 13,170 new CC cases and 4250 new deaths in America according to cancer statistics. In fact, CC is the second leading cause of cancer death in women aged 20–39 years, causing 9 deaths per week in this age group.2 Although vaccines and CC screenings, including human papillomavirus (HPV) test, cytologic test and colposcopy, have developed rapidly in recent years, effectively decreasing CC mortality,3,4 there is also an increasing need for cervical precancer screening and early-stage cervical cancer fertility preservation treatment, as well as chemotherapies such as paclitaxel.5
Hausen first postulated a possible role of HPV in CC in the 1970s and later put forward the structure of viral genes (such as E6 and E7) that were closely connected with CC.6 In fact, there are five genera of HPV, α, β, γ, μ and ν, including high-risk 16, 18, 31, 33, 35, 39, 45, 51, 52, 56, 58, 59 and 68 and low-risk 6, 11, 42 and 44. HPV prevalence is different according to genotype differences. Currently, it is known that HPVs lead to more than 90% of cervical lesions, especially HPV 16 and 18.7 In general, HPV is associated with approximately 70% of vaginal and vulvar cancers, 60% of penile cancers, 70% of oropharyngeal cancers and 10% of oral cavity cancers.8 In a worldwide study, HPV genotype prevalence rates with invasive cervical cancer were HPV16 (61%), HPV18 (10%), HPV31 (4%), HPV33 (4%), HPV35 (2%), HPV39 (2%), HPV45 (6%), HPV52 (3%), and HPV58 (2%), respectively.9,10 Then, HPV integrates into the host genome and mutates Retinoblastoma (Rb) or p53, which suppresses apoptosis and immortalizes epithelial cells, resulting in the development of low/high squamous intraepithelial lesions (LSIL/HSIL) or carcinogenesis.11,12
As HPV infection can be divided into transient and persistent infections, and 61% of LSIL cases regress spontaneously within 1 year and 91% regress within 2 years, there must exist an imbalance of the immune system in the context of persistent infection.13 Moreover, the immunosuppressed nature of the tumor microenvironment has long been appreciated, and CC evolution is well influenced by it.14,15 Therefore, this review attempts to focus on insights into the immune landscape and therapy of HPV-positive CC.
Cervical Cancer Genomics
A large number of studies have revealed that chromosome aberrations, DNA copies alterations, somatic mutations and methylations are related to the occurrence and development of CC. For instance, PIK3CA, TP53, KRAS and PTEN are all commonly mutated genes in CC patients. A recent study in Japan also showed that approximately one-third of Japanese women with CC developed a mutation of STK 11.16 In 2015, a team of academicians identified frequent HPV integrations in CC: POU5F1B, FHIT, KLF12, KLF5, LRP1B and LEPREL1, as well as HMGA2, DLG2 and SEMA3D, by conducting genome-wide sequencing and high-throughput viral integration detection.17 Recently, a group found that BRM270 could suppresses CC stem cells proliferation.18 Additionally, academicians found that host gene MYC disorder may also be the integration site of HPV.12 Researchers have begun to pay attention to the gene alterations in the tumor immune microenvironment, trying to predict the prognosis of patients. Researchers have also discovered 384 integrated gene sites related to T cell activation in the KEGG (gene ontology and Kyoto encyclopedia of genes and genomes) database.19 Therefore, this discovery elicits a comprehensive consideration for the study of immune genes in the tumor microenvironment and the prognosis of CC. As the majority of CCs are caused by HPV infection, the integration of E6 and E7 viral genes into the host genome causes excessive cell proliferation and ultimately leads to cancer. However, the mechanism of viral gene integration still needs further study.
Cervical Cancer and the Immune Landscape
On the one hand, the development of CC is directly linked to HPV infection. On the other hand, immune system defects play a significant role in cancer progress. It is believed that HPV infection triggers a primarily cell-mediated immune response, and there is evidence for T helper cell involvement in regressing lesions.20 One study suggested that Langerhans cells were increased in women who cleared HPV.21 Low- and high-risk HPV are stratified into different groups due to their oncogenic potential. Even though they stimulate a similar cellular environment and immune defense, it is interesting that they developed respective pathologies and cellular targets. One explanation is that low-risk HPV E7 protein has lower binding affinity, but the mechanism is not known.22 Tumors are recognized by the immune system and they may be attacked or prevented through a process called immunosurveillance. In the human body, mucosal immunity represents the first line of defense; cellular and humoral immunity also exert essential functions during carcinogenesis and disease progression. The peripheral blood of patients and the tumor microenvironment inspire us to explore cervical carcinoma’s connection with immunology.23
The Immune Cell Subpopulation in Cervical Cancer
As mentioned above, the characteristics of precancer LSIL and HSIL progression are also tightly correlated with the immune system. Thus, we discuss immune alteration with cervical lesions and cervical cancer below. An abundance of various types of immune cells comprise the tumor microenvironment, including T cells, B cells, dendritic cells, NK (nature killing) cells and macrophages. Immune cells can be divided into immunoactive and immunosuppressive types according to their function (Table 1). Moreover, the presence of many tumor-infiltrating lymphocytes (TIL) is generally associated with CC disease progression.24
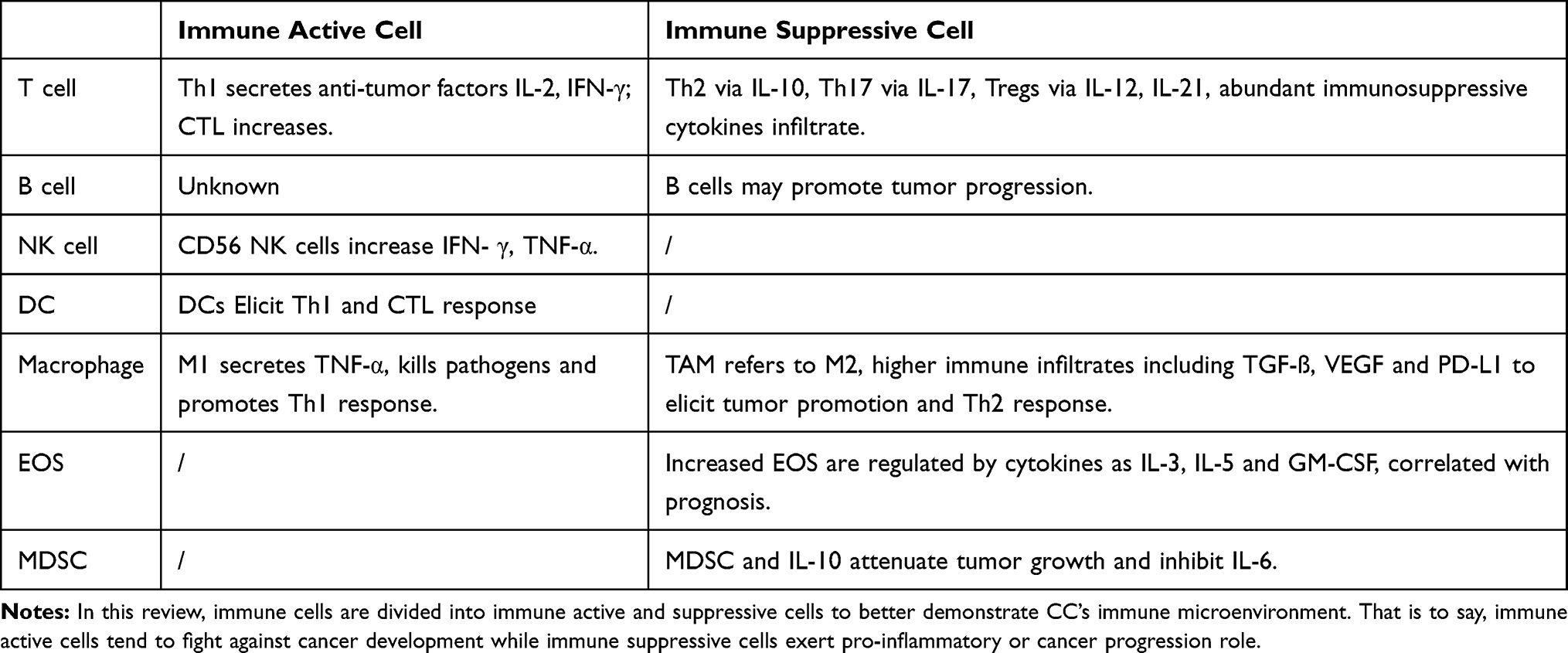 |
Table 1 Differences in Immunophenotype Between Active and Suppressive Cells for Cervical Cancer |
Tumor-Infiltrating Lymphocyte (TIL) T Cells
Analysis of tumor-infiltrating lymphocytes (TILs) is one of the cornerstones for the research of the tumor microenvironment. The interactions between tumors and the immune system are critical for tumor initiation, and TILs have attracted extensive attention in anti-tumor immunity. However, their functions during the tumor immunoediting processes are ambiguous. The cytotoxic immune response is characterized by CD4, CD8, antigen-presenting cells, and the infiltration of other lymphoid elements.25 CD4+ T cells are appointed to assist CD8+ T cells in exerting their effects. CD4+ T cells are subdivided into four major subsets: Th1, Th2, Th17 and regulatory T cells (Tregs).
Th1, Th2, Th17 and Tregs produce positive or negative effects in the maintenance of normal immune function by secreting various cytokines: interleukin-2 (IL-2) and interferon-γ (IFN-γ), the principal cytokines delivered by Th1cells, IL-12, a key cytokine involved in inducing and maintaining Th1cells and the IFN-γ responses, and IL-10, a potent modulator of cell-mediated immune responses secreted by Th2.26 IL-17 is a recently identified pro-inflammatory cytokine secreted by Th17 cells. According to a previous report, IL-17 promotes angiogenesis and cell proliferation and invasion in CC.27 Specifically, Th17 can modulate CC cell growth and apoptosis by the effect of miR-146a or miR155.28,29 A Th17/Treg imbalance often leads to infection, inflammatory response and autoimmune disorders.30 Th9 is a newly discovered T cell subset that is suspected to hamper tumor growth in several cancers, and it has been proved that Th9 can inhibit CC progression and immune evasion.31
Most importantly, Tregs and the production of TGF-ß are essential for immune homeostasis, and increasing evidence indicates that Tregs are present in human tumors and locally suppress antitumor T-cell responses.32 Moreover, Tregs express the transcription factor forkhead box protein 3 (Foxp3), which is important for maintaining self-tolerance and immune homeostasis, together with CD25.33 Additionally, Foxp3 Tregs’ prognostic value in different types of cancer is related to the tumor site.34 Studies also proved that elevated Foxp3 expression was associated with high-grade cervical disease and predicted poor overall survival (OS).35,36 Therefore, Foxp3 might be a useful biomarker for risk stratification in CC patients.37 Specifically, HLADRhi Tregs, a subset of Tregs, were associated with unfavorable outcomes in cervical squamous cell carcinoma. That is to say, HLADRhi Tregs were highly enriched in the tumor microenvironment and exhibited potent suppressive activity.38 In addition, IL-12 and IL-21 are both well-known agents, and a combination of them could fight against CC cells effectively by downregulating Tregs and Th17.39
CD8+ cytotoxic T lymphocytes (CTLs) are the major effector cells that kill tumors.40 One study observed that CD8+ cells predominantly infiltrated the epithelial layer in HPV+ normal cervical tissue, and the trend decreased with increasing cervical lesion size.41 Another report obtained the same result, showing that CD8+ cells increased after cisplatin treatment.42 Ultimately, CD8+ T cells downregulated macrophages or CD4+ cells and mainly exerted cytotoxic responses.43
B Cells
B cells are conventionally generated in germinal centers, which exert a strong and swift response to stimulation by secreting antibodies and cytokines. Recently, several teams discovered that B cells associated with tertiary lymphoid structures may influence the development of cancer progression.44–46 In 1984, a gynecologist reported that both T and B lymphocyte counts decreased in peripheral blood.47 Recently, a group of scientists discovered that B cells promoted HPV-mediated CC progression in a mouse model with the immunosuppressive cytokine IL-10.48 At the same time, another group found that B cells and IL-10 were increased in human CC samples, which may be significant in cancer progression.49 Most recently, B cells were proved to improve HPV-associated squamous CC, which may be activated by radiation and PD-1 blockade. Following data collection from over 800 head and neck squamous cell carcinoma (HNSCC) and CC patients, single-cell RNA-sequencing revealed dramatic increases in B-cell germinal center formation after PD-1 blockade and enhanced IgG and IgM antibody responses.50 However, the studies focusing on B cells in cervical cancer microenvironment are limited and might be a very much promising research direction. More reports are needed to support B cells’ role in CC.51
Dendritic Cells (DCs)
CD4+ and CD8+ T lymphocytes can be activated by specific antigen-presenting cells (APCs). Dendritic cells (DCs) are highly potent APCs and play an important role in the CC immune response, though the response is less focused on CC. Once DCs present antigen signals to elicit Th1 and CTL responses, anti-tumor immunity starts, inspiring us a novel immune therapeutic strategy.52,53 However, DCs tend to tolerate CC cells via RANKL secretion, a receptor activator of nuclear factor kappa-B ligand, a TNF family member. RANKL is a promising candidate for immune evasion together with Tregs in human CC.54 Furthermore, low DCs and high Tregs might be significantly associated with hrHPV persistence, suggesting that DCs gradually lose antigen-presenting ability gradually.55
Eosinophils
Eosinophils (EOSs) have long been known to infiltrate human tumors, such as gastric, colorectal, nasopharyngeal, oral, laryngeal and breast cancers. The cytokines IL-3, IL-5 and GM-CSF are critical in regulating EOS development. Several studies now focus on eosinophils’ function in CC.56 Researchers have proved that EOS infiltration increased with the progression of the CC. Thymic stromal lymphopoietin (TSLP) is a cytokine that can regulate EOS. Moreover, TSLP secreted by CC cells is able to promote tumor proliferation and invasion and may be correlated with a decline in microRNA-132 expression decline.57,58
Scientists have begun to focus their attention on eosinophils as a new CC prognostic biomarker. Recently, a new pre-treatment biomarker, the eosinophil-to-lymphocytes ratio (ELR), has been used to predict prognosis in CC patients. Studies have found out that higher values of ELR in peripheral blood are suspected to predict better overall survival (OS), while the ELR value in tumor is unclear.59 However, another report disagreed with the former retrospective study and concluded that higher eosinophil number are correlated with worse outcomes of cervical squamous cell cancer.60
NK Cells
NK cells play an important role in cancer immunity through secreting various cytokines. In the tumor environment, NK cells are stimulated by IL-2. Likewise, NK cells release interferon gamma (IFN-γ) and tumor necrosis factor alpha (TNF-α) to fight against CC tumor proliferation.61 In other words, NK cells are the primary effector to recognize abnormal cells without an antigen presentation process. NKG2D, a type Ⅱ C-type lectin-like family of transmembrane proteins that is different from the NK cell biomarker CD56, works as a stimulatory receptor. Several studies have revealed its association with cancer immunosurveillance and HPV-induced cancers, indicating that the NKG2D gene may influence NK cell cytotoxicity and susceptibility to CC.62,63 While NKG2D, NKp46, and NKp30 are activating receptors, CD158a, CD158b, and NKG2A are inhibitory NK receptors. One study detected the upregulation of inhibitory NK receptors in CC, suggesting that Tregs may suppress NK function by the inhibition of TGF-ß64 In addition, recent studies have reported that NK cells could not only mediate immune clearance but also predict disease prognosis. One clinical trial indicated that after 4 cycles of chemotherapy on cervical squamous cell carcinoma at stages Ⅱb-шb, NK cells increased and tumor size reduced.65 Another study added evidence to this discovery, reporting that NK cells were significantly associated with improved prognosis.66
Tumor Associated Macrophage
Macrophages are derived from bone marrow, and their role is particularly prominent. At the same time the function of tumor-associated macrophage (TAMs) has long been discussed. Macrophages can be divided into two groups, namely the classical M1 and the alternative M2, which promote inflammation and tumor progression. First, M1 macrophages are activated by IFN-γ, lipopolysaccharides (LPS) through Toll-like receptors and granulocyte-monocyte colony-stimulating factor (GM-CSF). M1 macrophages highly express major histocompatibility complex class Ⅱ and co-stimulatory molecules such as CD86/CD80, IL-12, IL-13, TNF-α, and reactive nitrogen species. Their major functions are killing pathogens and promoting a Th1 immune response. M2 macrophages are stimulated by IL-4, IL-10, IL-13, IL-33 and IL-21 and release vascular endothelial growth factor (VEGF), transforming growth factor (TGF)-ß, indoleamine 2.3-dioxygenase, and programmed death ligand 1 (PD-L1) expression. M2 macrophages can take part in tumor promotion and the Th2 response.67 CD68 is a well-recognized maker of macrophages, while CD163 is regarded as a dominant marker for identifying M2 macrophages. As M2 macrophages mostly promote tumor inflammation, TAMs more likely refer to M2 macrophages.68
In 2007, a study first described the population of macrophages in cervical intraepithelial neoplasia (CIN) progression and its influence on CIN outcome. Not surprisingly, the macrophage percentage increased linearly with neoplasia progression.69 Later studies further revealed that M2 TAMs were associated with high-risk HPV infection and were positively correlated with cervical carcinogenesis. More importantly, higher FIGO stage and lymph node metastasis or lymphangiogenesis usually showed larger counts of M2 macrophages.70–72 M2 macrophages are usually associated with poor prognosis. CC cell lines possibly induce more monocytes toward an M2-like phenotype in TAMs, significantly maintaining a tumor immunosuppressive microenvironment and promoting angiogenesis and metastasis.67,73 Furthermore, one study suspected that CC cell lines could induces monocytes into M2 macrophages through lactate secretion.74 Another study reported that CC cell supernatants may shift LPS-induced M1 into M2 macrophages by increasing the production of TLR and nitric oxide (NO).75 Moreover, the hypoxic tumor microenvironment can modulate the activation of M2 macrophages via neuropilin-1 (Nrp-1), which may serve as a potential therapeutic marker.76
MDSCs
Myeloid derived suppressor cells (MDSCs) are closely linked with tumor staging, progression, clinical therapeutic efficacy and prognosis; thus, MDSCs play an immunosuppressive and tumor-promoting role.77,78 Using an HPV-mediated CC mouse model, researchers proved that MDSCs mediated an immunosuppressive activity via IL-6-JAK-STAT3 (signal transducer and activator of transcription 3) signaling.15 More specifically, IL-10 might attenuate tumor growth by inhibiting IL-6 release while activating STAT3 signaling in CC.79 MDSCs impaired CD8+ T cells cytotoxicity, activation of APCs and immune responses to foreign antigens, ultimately destroying the efficacy of immune therapy against HPV-mediated cancer.15
Immune Therapy
Immune therapy is the next generation treatment compared with traditional therapies such as tumor surgery, radiotherapy and chemical therapy. Immune therapy aims to evoke the body’s immune system and then enhance tumor-killing ability. In general, CC immune therapy can be divided into four groups: immune checkpoint blockade (ICB), adoptive cell transfer therapy, therapeutic vaccines and cytokine treatment.
Immune Checkpoint Blockade
Immune suppression and activation are two poles of the immune system. Cancer cells are able to escape from the immune balance by provoking an immune-suppressive state and tumor growth. Immune checkpoints often educate the tumor microenvironment into immune tolerance.80 There are eight well known immune checkpoints [programmed cell death protein 1 (PD-1), cytotoxic T-lymphocyte antigen-4 (CTLA-4), T-cell immunoglobulin and mucin-domain containing-3 (Tim-3), 2B4, killer cell lectin like receptor G1 (KLRG-1), TIGIT, B- and T-lymphocyte attenuator (BTLA) and CD160], which serve as the inhibitory signals of T cell activation and help maintain self-tolerance.81 Checkpoint blockades aim to break microenvironment immune suppression. PD-1 and CLTA-4 are the most promising immune checkpoints targeted in CC.
At present, PD-1/PD-L1 (PD-Ligand1) and CTLA-4 checkpoint inhibitors for CC being tested for clinical use (Table 2). Though immune checkpoint inhibitors have durable response rates, side effects such as autoimmune diseases and low response rates are still unavoidable challenges.82 The expression levels of PD-1/PD-L1 and CTLA-4 are high in CC, and DCs and T cells express high levels of PD-1 and PD-L1 in CIN samples.83,84 At the same time, the costimulatory molecules CD80 and CD86 and the anti-inflammatory cytokines IL-12 and IFN-γ are decreased with the increased expression of the immune-suppressive cytokine IL-10.85 This phenomenon suggests that the upregulation of the immune checkpoint contributes to immune evasion and tumor progression. One study observed that PD-1/PD-L1 blockade interrupted immune suppression in CC; they determined that a subset of T cells, CD8+FoxP3+CD25+T cells increased and immune-suppressive Tregs decreased.86 Therefore, ICB therapy intends to enhance the body’s immune defense and lessen immune surveillance. The application of ICB has rapidly become a promising treatment approach in this decade. In addition, the combination of PD-1/PD-L1 and CTLA-4 may enhance the therapeutic efficiency. Studies in animal models have determined that this combination prolonged the survival rate with fewer side effects.32,87 Several clinical trials have investigated the PD-1 checkpoint inhibitors nivolumab and pembrolizumab, the PD-L1 checkpoint inhibitors atezolizumab and durvalumab, or the CTLA-4 inhibitor ipilimumab; some trials are still ongoing, while others have come to their conclusion. These clinical trials proved the safety of ICB, even though many tolerable side effects occurred. However, we still need to pave the way to confirm ICB’s antitumor activity in CC.88–91
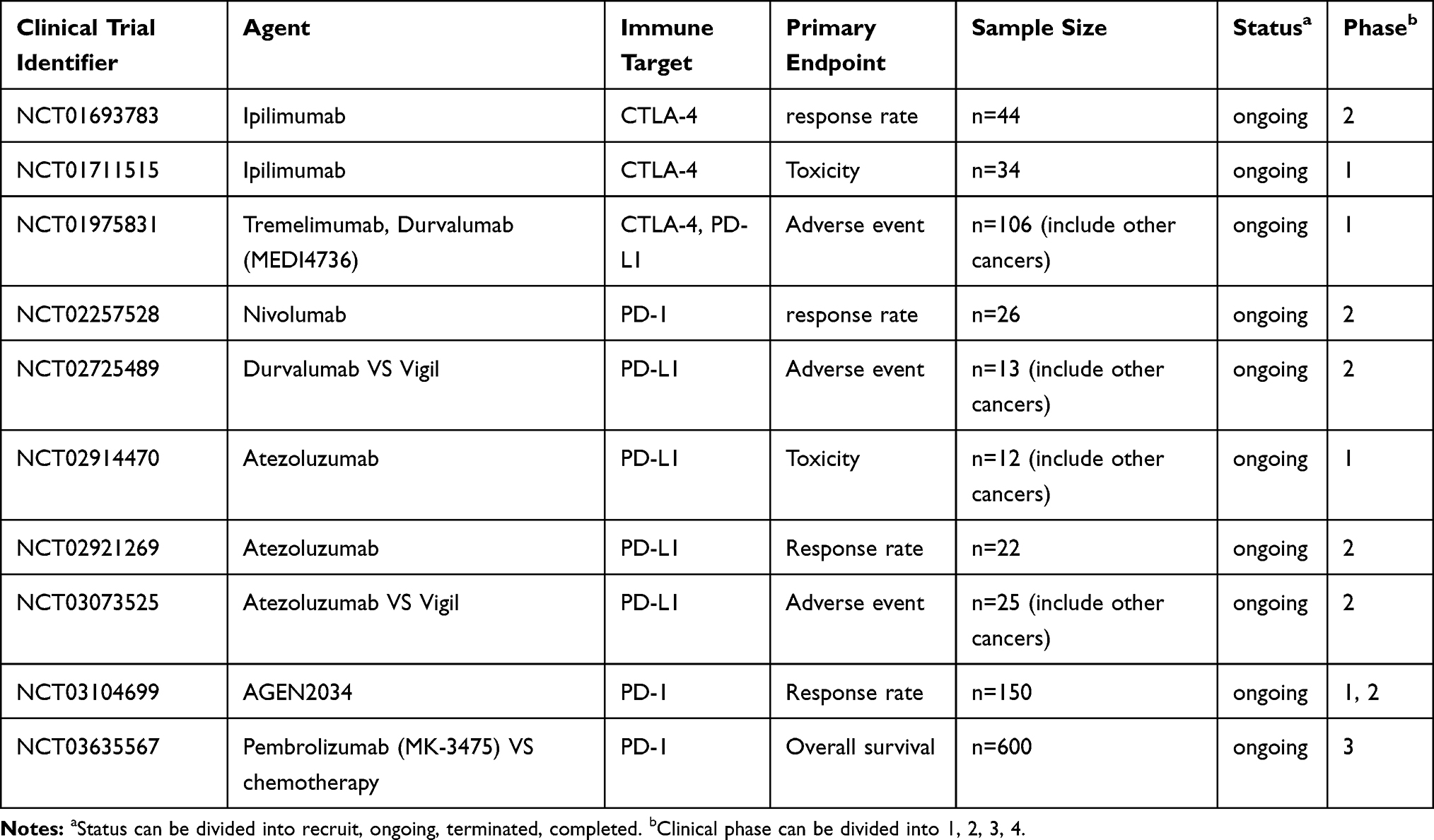 |
Table 2 Studies of Immune Checkpoint Inhibitor PD-1/PD-L1 and CTLA4 from Clinicaltrials.gov (Accessed 15th April 2020) |
Among gynecological malignancies, besides CC, blockading the PD-1/PD-L1 pathway may also be beneficial in ovarian cancer and vulvar cancer.92,93 Both ovarian cancer and vulvar cancer express high levels of PD-1/PD-L1, and in tumor cells and mouse models, their antitumor immunities were detected via CD8+ T cells and Tregs regulation.94,95 However, more clinical trials are needed to explore the effects of PD-1/PD-L1 ICB.
Adoptive Cell Transfer Therapy
Adoptive cell transfer therapy is a highly effective pharmacological option against cancers.96,97 T cells play an important role in CC immune surveillance; thus, antigen-specific T cell immunotherapy could be used to attack tumor evasion and may have many potential benefits. Scientists have observed that isolated T lymphocytes with HPV E6 and E7 specificity from CC patients expanded stably in vitro.98 However, the current methods to expand HPV specific T cells from healthy donors have failed to achieve expected results; by optimizing DC maturation and adding appropriate cytokines, it is possible to obtain oncoproteins E6 and E7-specific T cells.99 If get a good balance in clinical conditions and engineered cell projects is maintained, the potential use of HPV-specific T cells for CC is very promising. Chimeric antigen receptor T cells (CAR-T) have been approved by the Food and Drug Administration (FDA) and are successfully improving outcomes for hematological malignancies.100 Although the research studies involving CAR-T therapy in CC are rare, one study investigated the killing effect of mesothelin-CAR-T in CC cells and achieved positive results.101 A clinical trial combined autologous cytokine-induced killer (CIK) cell transfusion and radiochemotherapy in CC patients and showed that the application of CIK cells improved immune function and life quality.102 Moreover, NK cell transfusion may also improve CC status.103 In conclusion, adoptive cell transfer therapy should be given more attention in CC.
Therapeutic Vaccines and Cytokines
Virus-like particles (VLPs) of HPV are used as prophylactic vaccines against CC. Though CC is not rare, current treatments involving therapeutic vaccines are deficient. Among preclinical research studies, a therapeutic HPV16 E6/7 vaccines provided a highly effective immune response in murine models, evoking CD8+ and CD4+ T cells via targeting CD40.40 More Specifically, there is also crosstalk between DCs and innate immune NK cells that assists in the HPV defense effect via CD40 interaction and IL-12p70 secretion, resulting in the production of neutralizing antibodies and cellular immunity.104 Administration of Fc-fused IL7 could also play a role in modulating vaccines therapy through a CD8+ T cell response.105 Various therapeutic vaccines are being developed, including DNA, RNA and peptide vaccines. All of these vaccines types exerted antitumor effects through activating the T cell response, especially CD8+ cells.106–108 In addition, increased level of IFN- γ helped mediate cellular immunity.109 Scientists prefer to combine therapeutic vaccines and immune checkpoint inhibitors such as PL-L1 and CTLA-4 in trials.106,110 Several clinical trials have indicated that therapeutic vaccines plus immune checkpoint inhibitors or radiotherapy together induced an immune response in premalignant lesions and CC.111 Interestingly, another clinical trial even demonstrated that VGX-3100, a therapeutic DNA vaccine targeting HPV16/18, had histopathological regression for CIN.112 Thus, HPV-associated therapeutic vaccines for CC are a promising research trend that will require further study.113
Cytokine therapy has long been an exciting field for cancer treatment since its initial discovery due to its easy accessibility and construction. The release of cytokines has crucial effects in controlling immune responses, educating lymphocytes maturation and exerting biological activities. However, most cytokine-based therapy trials have not performed as expected, with the major obstacle being toxic reaction.114 In common, TNF-α, IL-6, IL-8, IL-1α are classical pro-inflammatory cytokines, and IL-10, IL-12, TGF-ß are anti-inflammatory cytokines. Their specific functions in CC are not clear. IL-2 and IL-6 confer protection to CC cells against apoptosis.115–117 In addition, immunoactive TNF- α and immunosuppressive IL-10 are associated with CC susceptibility.118 More efforts are needed to explore cytokine-based therapies in CC.
Perspective
Even with the recent advances in understanding the genomic and immune landscape of CC, there have been few clinically useful biomarkers developed except for HPV tests and immunohistochemistry (IHC). CC is closely connected with virus infection, which fights against the human immune system. It is commonly known that the tumor microenvironment is very different from normal tissue, and immune-suppressive cells assume a leadership role in immune cell population. Therefore, obtaining a thorough knowledge of tumor immunology will definitely promote CC treatment. Immune therapy is one of the most promising breakthroughs in cancer treatment. Moreover, PD-1/PD-L1 blockade seems to be a potential treatment approach for CC, though the toxicities of current immune therapy will require more effort. Additional research studies bridging the gap between HPV-positive cervical disease and the vaginal metabolome.119 These discoveries enlightened us of the importance of enhancing CC immune research with metabolism studies.
Author Contributions
All authors contributed to conception and design, drafting and revising the article, gave final approval of the version to be published and agreed to be accountable for all aspects of the work.
Disclosure
The authors report no conflicts of interest for this work. This project was supported by funding from the Research and Innovation Project of Shanghai Municipal Education Commission (No. 2019-01-07-00-07-E00050), and the Shanghai Science and Technology Natural Science Foundation (No. 19ZR1406900).
References
1. Bray F, Ferlay J, Soerjomataram I, Siegel RL, Torre LA, Jemal A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2018;68(6):394–424. doi:10.3322/caac.21492
2. Siegel RL, Miller KD, Jemal A. Cancer statistics, 2019. CA: Cancer J Clin. 2019;69(1):7–34. doi:10.3322/caac.21551
3. Feng-Yi X, Feng X, Long S. Diagnostic accuracy of colposcopically directed biopsy and loop electrosurgical excision procedure for cervical lesions. Reprod Dev Med. 2018;2(3):137–141.
4. Qing W, Cai-Ying Z, Li-Mei C, et al. Clinical value of human papillomavirus E6/E7 mRNA testing in patients with atypical squamous cells of undetermined significance and low-grade squamous intraepithelial lesion. Reprod Dev Med. 2018;2(3):157–161.
5. Della Corte L, Barra F, Foreste V, et al. Advances in paclitaxel combinations for treating cervical cancer. Expert Opin Pharmacother. 2020;21(6):663–677. doi:10.1080/14656566.2020.1724284
6. Schwarz E, Freese UK, Gissmann L, et al. Structure and transcription of human papillomavirus sequences in cervical carcinoma cells. Nature. 1985;314(6006):111–114. doi:10.1038/314111a0
7. Li S, Hong X, Wei Z, et al. Ubiquitination of the HPV oncoprotein E6 is critical for E6/E6AP-mediated p53 degradation. Front Microbiol. 2019;10:2483. doi:10.3389/fmicb.2019.02483
8. Li Y, Xu C. Human papillomavirus-related cancers. Adv Exp Med Biol. 2017;1018:23–34.
9. de Sanjose S, Quint WG, Alemany L, et al. Human papillomavirus genotype attribution in invasive cervical cancer: a retrospective cross-sectional worldwide study. Lancet Oncol. 2010;11(11):1048–1056. doi:10.1016/S1470-2045(10)70230-8
10. Schiffman M, Kjaer SK. Chapter 2: natural history of anogenital human papillomavirus infection and neoplasia. J Natl Cancer Inst Monogr. 2003;2003(31):14–19. doi:10.1093/oxfordjournals.jncimonographs.a003476
11. Liu L, Ying C, Zhao Z, et al. Identification of reliable biomarkers of human papillomavirus 16 methylation in cervical lesions based on integration status using high-resolution melting analysis. Clin Epigenetics. 2018;10:10. doi:10.1186/s13148-018-0445-8
12. Zhang R, Shen C, Zhao L, et al. Dysregulation of host cellular genes targeted by human papillomavirus (HPV) integration contributes to HPV-related cervical carcinogenesis. Int J Cancer. 2016;138(5):1163–1174. doi:10.1002/ijc.29872
13. Moscicki AB, Shiboski S, Hills NK, et al. Regression of low-grade squamous intra-epithelial lesions in young women. Lancet. 2004;364(9446):1678–1683. doi:10.1016/S0140-6736(04)17354-6
14. Chun-Yan W, Yu-Qing Q, Yin-Yan H, Qing W, Xiao-Yong Z, Jun S. A retrospective review of 10 cases of villoglandular papillary adenocarcinoma of the uterine cervix including one with successful pregnancy. Reprod Dev Med. 2018;2(2):120–127.
15. Galliverti G, Wullschleger S, Tichet M, et al. Myeloid cells orchestrate systemic immunosuppression, impairing the efficacy of immunotherapy against HPV(+) cancers. Cancer Immunol Res. 2020;8(1):131–145. doi:10.1158/2326-6066.CIR-19-0315
16. Hirose S, Murakami N, Takahashi K, et al. Genomic alterations in STK11 can predict clinical outcomes in cervical cancer patients. Gynecol Oncol. 2020;156(1):203–210. doi:10.1016/j.ygyno.2019.10.022
17. Hu Z, Zhu D, Wang W, et al. Genome-wide profiling of HPV integration in cervical cancer identifies clustered genomic hot spots and a potential microhomology-mediated integration mechanism. Nat Genet. 2015;47(2):158–163. doi:10.1038/ng.3178
18. Chandimali N, Sun HN, Park YH, Kwon T. BRM270 suppresses cervical cancer stem cell characteristics and progression by inhibiting SOX2. In Vivo (Brooklyn). 2020;34(3):1085–1094. doi:10.21873/invivo.11879
19. Pan XB, Lu Y, Huang JL, Long Y, Yao DS. Prognostic genes in the tumor microenvironment in cervical squamous cell carcinoma. Aging (Albany NY). 2019;11(22):10154–10166. doi:10.18632/aging.102429
20. Burd EM. Human papillomavirus and cervical cancer. Clin Microbiol Rev. 2003;16(1):1–17. doi:10.1128/CMR.16.1.1-17.2003
21. Shannon B, Yi TJ, Perusini S, et al. Association of HPV infection and clearance with cervicovaginal immunology and the vaginal microbiota. Mucosal Immunol. 2017;10(5):1310–1319. doi:10.1038/mi.2016.129
22. Ivancic R, Iqbal H, deSilva B, Pan Q, Matrka L. Immunological tolerance of low-risk HPV in recurrent respiratory papillomatosis. Clin Exp Immunol. 2020;199(2):131–142. doi:10.1111/cei.13387
23. Foy JP, Bertolus C, Michallet MC, et al. The immune microenvironment of HPV-negative oral squamous cell carcinoma from never-smokers and never-drinkers patients suggests higher clinical benefit of IDO1 and PD1/PD-L1 blockade. Ann Oncol. 2017;28(8):1934–1941. doi:10.1093/annonc/mdx210
24. Huang H, Zhang M, Yao S, Zhang M, Peng J. Immune modulation of a lipid-soluble extract of Pinellia pedatisecta Schott in the tumor microenvironment of an HPV(+) tumor-burdened mouse model. J Ethnopharmacol. 2018;225:103–115. doi:10.1016/j.jep.2018.04.037
25. Qiao G, Wang X, Zhou X, et al. Immune correlates of clinical benefit in a Phase I study of hyperthermia with adoptive T cell immunotherapy in patients with solid tumors. Int J Hyperthermia. 2019;36(sup1):74–82. doi:10.1080/02656736.2019.1647350
26. Ma W, Wang K, Du J, Luan J, Lou G. Multi-dose parecoxib provides an immunoprotective effect by balancing T helper 1 (Th1), Th2, Th17 and regulatory T cytokines following laparoscopy in patients with cervical cancer. Mol Med Rep. 2015;11(4):2999–3008. doi:10.3892/mmr.2014.3003
27. Lv Q, Wu K, Liu F, Wu W, Chen Y, Zhang W. Interleukin17A and heparanase promote angiogenesis and cell proliferation and invasion in cervical cancer. Int J Oncol. 2018;53(4):1809–1817. doi:10.3892/ijo.2018.4503
28. Li T, Li M, Xu C, et al. miR146a regulates the function of Th17 cell differentiation to modulate cervical cancer cell growth and apoptosis through NFkappaB signaling by targeting TRAF6. Oncol Rep. 2019;41(5):2897–2908. doi:10.3892/or.2019.7046
29. Zhang Y, Wang ZC, Zhang ZS, Chen F. MicroRNA-155 regulates cervical cancer via inducing Th17/Treg imbalance. Eur Rev Med Pharmacol Sci. 2018;22(12):3719–3726. doi:10.26355/eurrev_201806_15251
30. Zhang Y, Ma D, Zhang Y, et al. The imbalance of Th17/Treg in patients with uterine cervical cancer. Clin Chim Acta. 2011;412(11–12):894–900. doi:10.1016/j.cca.2011.01.015
31. Chauhan SR, Singhal PG, Sharma U, Bandil K, Chakraborty K, Bharadwaj M. Th9 cytokines curb cervical cancer progression and immune evasion. Hum Immunol. 2019;80(12):1020–1025. doi:10.1016/j.humimm.2019.09.009
32. Tuve S, Chen BM, Liu Y, et al. Combination of tumor site-located CTL-associated antigen-4 blockade and systemic regulatory T-cell depletion induces tumor-destructive immune responses. Cancer Res. 2007;67(12):5929–5939. doi:10.1158/0008-5472.CAN-06-4296
33. Bloom DD, Reshetylo S, Nytes C, Goodsett CT, Hematti P. Blockade of BAFF receptor BR3 on T cells enhances their activation and cytotoxicity. J Immunother. 2018;41(5):213–223. doi:10.1097/CJI.0000000000000209
34. Shang B, Liu Y, Jiang SJ, Liu Y. Prognostic value of tumor-infiltrating FoxP3+ regulatory T cells in cancers: a systematic review and meta-analysis. Sci Rep. 2015;5:15179. doi:10.1038/srep15179
35. Scott ME, Ma Y, Kuzmich L, Moscicki AB. Diminished IFN-gamma and IL-10 and elevated Foxp3 mRNA expression in the cervix are associated with CIN 2 or 3. Int J Cancer. 2009;124(6):1379–1383. doi:10.1002/ijc.24117
36. Zeng C, Yao Y, Jie W, et al. Up-regulation of Foxp3 participates in progression of cervical cancer. Cancer Immunol Immunother. 2013;62(3):481–487. doi:10.1007/s00262-012-1348-8
37. Chen R, Gong Y, Zou D, Wang L, Yuan L, Zhou Q. Correlation between subsets of tumor-infiltrating immune cells and risk stratification in patients with cervical cancer. PeerJ. 2019;7:e7804. doi:10.7717/peerj.7804
38. Yang H, Ye S, Goswami S, et al. Highly immunosuppressive HLADR(hi) regulatory T cells are associated with unfavorable outcomes in cervical squamous cell carcinoma. Int J Cancer. 2019;146(7):1993–2006.
39. Tian Y, Yuan C, Ma D, et al. IL-21 and IL-12 inhibit differentiation of Treg and TH17 cells and enhance cytotoxicity of peripheral blood mononuclear cells in patients with cervical cancer. Int J Gynecol Cancer. 2011;21(9):1672–1678. doi:10.1097/IGC.0b013e3182358955
40. Yin W, Duluc D, Joo H, et al. Therapeutic HPV cancer vaccine targeted to CD40 elicits effective CD8+ T-cell immunity. Cancer Immunol Res. 2016;4(10):823–834. doi:10.1158/2326-6066.CIR-16-0128
41. Maskey N, Thapa N, Maharjan M, et al. Infiltrating CD4 and CD8 lymphocytes in HPV infected uterine cervical milieu. Cancer Manag Res. 2019;11:7647–7655. doi:10.2147/CMAR.S217264
42. Heeren AM, van Luijk IF, Lakeman J, et al. Neoadjuvant cisplatin and paclitaxel modulate tumor-infiltrating T cells in patients with cervical cancer. Cancer Immunol Immunother. 2019;68(11):1759–1767. doi:10.1007/s00262-019-02412-x
43. de Vos van Steenwijk PJ, Ramwadhdoebe TH, Goedemans R, et al. Tumor-infiltrating CD14-positive myeloid cells and CD8-positive T-cells prolong survival in patients with cervical carcinoma. Int J Cancer. 2013;133(12):2884–2894. doi:10.1002/ijc.28309
44. Cabrita R, Lauss M, Sanna A, et al. Tertiary lymphoid structures improve immunotherapy and survival in melanoma. Nature. 2020;577(7791):561–565. doi:10.1038/s41586-019-1914-8
45. Helmink BA, Reddy SM, Gao J, et al. B cells and tertiary lymphoid structures promote immunotherapy response. Nature. 2020;577(7791):549–555. doi:10.1038/s41586-019-1922-8
46. Petitprez F, de Reynies A, Keung EZ, et al. B cells are associated with survival and immunotherapy response in sarcoma. Nature. 2020;577(7791):556–560. doi:10.1038/s41586-019-1906-8
47. Kietlińska Z. T and B lymphocyte counts and blast transformation in patients with Stage I cervical cancer. Gynecol Oncol. 1984;18(2):247–256. doi:10.1016/0090-8258(84)90033-7
48. Tang A, Dadaglio G, Oberkampf M, et al. B cells promote tumor progression in a mouse model of HPV-mediated cervical cancer. Int J Cancer. 2016;139(6):1358–1371. doi:10.1002/ijc.30169
49. Chen Z, Zhu Y, Du R, et al. Role of regulatory B cells in the progression of cervical cancer. Mediators Inflamm. 2019;2019:6519427. doi:10.1155/2019/6519427
50. Kim SS, Shen S, Miyauchi S, et al. B cells improve overall survival in HPV-associated squamous cell carcinomas and are activated by radiation and PD-1 blockade. Clin Cancer Res. 2020;26(13):3345–3359. doi:10.1158/1078-0432.CCR-19-3211
51. Lucena AA, Guimaraes MV, Michelin MA, et al. Evaluation of T, B and natural killer lymphocyte in the cervical stroma of HIV-positive and negative patients with cervical intraepithelial neoplasia. Immunol Lett. 2016;169:98–103. doi:10.1016/j.imlet.2015.10.016
52. Cao G, Cui R, Liu C, Zhang G, Zhang Z. MTBHsp70-exFPR1-pulsed dendritic cells enhance the immune response against cervical cancer. J Cancer. 2019;10(25):6364–6373. doi:10.7150/jca.29779
53. Wang Y, Huang H, Yao S, et al. A lipid-soluble extract of Pinellia pedatisecta Schott enhances antitumor T cell responses by restoring tumor-associated dendritic cell activation and maturation. J Ethnopharmacol. 2019;241:111980. doi:10.1016/j.jep.2019.111980
54. Demoulin SA, Somja J, Duray A, et al. Cervical (pre)neoplastic microenvironment promotes the emergence of tolerogenic dendritic cells via RANKL secretion. Oncoimmunology. 2015;4(6):e1008334. doi:10.1080/2162402X.2015.1008334
55. Strickler HD, Martinson J, Desai S, et al. The relation of plasmacytoid dendritic cells (pDCs) and regulatory T-cells (Tregs) with HPV persistence in HIV-infected and HIV-uninfected women. Viral Immunol. 2014;27(1):20–25. doi:10.1089/vim.2013.0097
56. Zhang B, Wei CY, Chang KK, et al. TSLP promotes angiogenesis of human umbilical vein endothelial cells by strengthening the crosstalk between cervical cancer cells and eosinophils. Oncol Lett. 2017;14(6):7483–7488. doi:10.3892/ol.2017.7121
57. Zhou WJ, Yang HL, Chang KK, et al. Human thymic stromal lymphopoietin promotes the proliferation and invasion of cervical cancer cells by downregulating microRNA-132 expression. Oncol Lett. 2017;14(6):7910–7916. doi:10.3892/ol.2017.7260
58. Xie F, Liu LB, Shang WQ, et al. The infiltration and functional regulation of eosinophils induced by TSLP promote the proliferation of cervical cancer cell. Cancer Lett. 2015;364(2):106–117. doi:10.1016/j.canlet.2015.04.029
59. Holub K, Biete A. Impact of systemic inflammation biomarkers on the survival outcomes of cervical cancer patients. Clin Transl Oncol. 2019;21(7):836–844. doi:10.1007/s12094-018-1991-4
60. Zhu J, Wang H, Gao MJ, et al. Prognostic values of lymphocyte and eosinophil counts in resectable cervical squamous cell carcinoma. Future Oncol. 2019;15(30):3467–3481. doi:10.2217/fon-2018-0879
61. Zhu SY, Wu QY, Zhang CX, et al. miR-20a inhibits the killing effect of natural killer cells to cervical cancer cells by downregulating RUNX1. Biochem Biophys Res Commun. 2018;505(1):309–316. doi:10.1016/j.bbrc.2018.09.102
62. Espinoza JL, Nguyen VH, Ichimura H, et al. A functional polymorphism in the NKG2D gene modulates NK-cell cytotoxicity and is associated with susceptibility to human papilloma virus-related cancers. Sci Rep. 2016;6:39231. doi:10.1038/srep39231
63. Escarra-Senmarti M, Bueno-Topete MR, Jave-Suarez LF, et al. Loss of CD28 within CD4(+) T cell subsets from cervical cancer patients is accompanied by the acquisition of intracellular perforin, and is further enhanced by NKG2D expression. Immunol Lett. 2017;182:30–38. doi:10.1016/j.imlet.2017.01.006
64. Chang WC, Li CH, Chu LH, Huang PS, Sheu BC, Huang SC. Regulatory T Cells Suppress Natural Killer Cell Immunity in Patients With Human Cervical Carcinoma. Int J Gynecol Cancer. 2016;26(1):156–162. doi:10.1097/IGC.0000000000000578
65. Saraswati W, Dahlan EG, Saputra K, Sutrisno TC. Effect of electroacupuncture on natural-killer cells and tumor size in patients with cervical squamous-cell carcinoma: a randomized controlled trial. Med Acupunct. 2019;31(1):29–36. doi:10.1089/acu.2018.1316
66. Wang J, Li Z, Gao A, Wen Q, Sun Y. The prognostic landscape of tumor-infiltrating immune cells in cervical cancer. Biomed Pharmacother. 2019;120:109444. doi:10.1016/j.biopha.2019.109444
67. Pedraza-Brindis EJ, Sanchez-Reyes K, Hernandez-Flores G, et al. Culture supernatants of cervical cancer cells induce an M2 phenotypic profile in THP-1 macrophages. Cell Immunol. 2016;310:42–52. doi:10.1016/j.cellimm.2016.07.001
68. Li Y, Huang G, Zhang S. Associations between intratumoral and peritumoral M2 macrophage counts and cervical squamous cell carcinoma invasion patterns. Int J Gynaecol Obstet. 2017;139(3):346–351. doi:10.1002/ijgo.12320
69. Hammes LS, Tekmal RR, Naud P, et al. Macrophages, inflammation and risk of cervical intraepithelial neoplasia (CIN) progression- Clinicopathological correlation. Gynecol Oncol. 2007;105(1):157–165. doi:10.1016/j.ygyno.2006.11.023
70. Ding H, Cai J, Mao M, et al. Tumor-associated macrophages induce lymphangiogenesis in cervical cancer via interaction with tumor cells. Apmis. 2014;122(11):1059–1069. doi:10.1111/apm.12257
71. Chen XJ, Han LF, Wu XG, et al. Clinical significance of CD163+ and CD68+ tumor-associated macrophages in high-risk HPV-related cervical cancer. J Cancer. 2017;8(18):3868–3875. doi:10.7150/jca.21444
72. Swangphon P, Pientong C, Sunthamala N, et al. Correlation of circulating CD64(+)/CD163(+) monocyte ratio and stroma/peri-tumoral CD163(+) monocyte density with human papillomavirus infected cervical lesion severity. Cancer Microenviron. 2017;10(1–3):77–85. doi:10.1007/s12307-017-0200-2
73. Jiang S, Yang Y, Fang M, Li X, Yuan X, Yuan J. Co-evolution of tumor-associated macrophages and tumor neo-vessels during cervical cancer invasion. Oncol Lett. 2016;12(4):2625–2631. doi:10.3892/ol.2016.5014
74. Stone SC, Rossetti RAM, Alvarez KLF, et al. Lactate secreted by cervical cancer cells modulates macrophage phenotype. J Leukoc Biol. 2019;105(5):1041–1054. doi:10.1002/JLB.3A0718-274RR
75. Sanchez-Reyes K, Bravo-Cuellar A, Hernandez-Flores G, et al. Cervical cancer cell supernatants induce a phenotypic switch from U937-derived macrophage-activated M1 state into M2-like suppressor phenotype with change in Toll-like receptor profile. Biomed Res Int. 2014;2014:683068. doi:10.1155/2014/683068
76. Chen XJ, Wu S, Yan RM, et al. The role of the hypoxia-Nrp-1 axis in the activation of M2-like tumor-associated macrophages in the tumor microenvironment of cervical cancer. Mol Carcinog. 2019;58(3):388–397. doi:10.1002/mc.22936
77. Liang Y, Lu B, Zhao P, Lu W. Increased circulating GrMyeloid-derived suppressor cells correlated with tumor burden and survival in locally advanced cervical cancer patient. J Cancer. 2019;10(6):1341–1348. doi:10.7150/jca.29647
78. Wu L, Liu H, Guo H, et al. Circulating and tumor-infiltrating myeloid-derived suppressor cells in cervical carcinoma patients. Oncol Lett. 2018;15(6):9507–9515. doi:10.3892/ol.2018.8532
79. Lee BR, Kwon BE, Hong EH, et al. Interleukin-10 attenuates tumour growth by inhibiting interleukin-6/signal transducer and activator of transcription 3 signalling in myeloid-derived suppressor cells. Cancer Lett. 2016;381(1):156–164. doi:10.1016/j.canlet.2016.07.012
80. Heeren AM, Koster BD, Samuels S, et al. High and interrelated rates of PD-L1+CD14+ antigen-presenting cells and regulatory T cells mark the microenvironment of metastatic lymph nodes from patients with cervical cancer. Cancer Immunol Res. 2015;3(1):48–58. doi:10.1158/2326-6066.CIR-14-0149
81. Li X, Wang R, Fan P, et al. A comprehensive analysis of key immune checkpoint receptors on tumor-infiltrating T cells from multiple types of cancer. Front Oncol. 2019;9:1066. doi:10.3389/fonc.2019.01066
82. Zhen S, Lu J, Liu YH, Chen W, Li X. Synergistic antitumor effect on cervical cancer by rational combination of PD1 blockade and CRISPR-Cas9-mediated HPV knockout. Cancer Gene Ther. 2019;27:1–11.
83. Hu S, Pu D, Xia X, Guo B, Zhang C. CTLA-4 rs5742909 polymorphism and cervical cancer risk: a meta-analysis. Medicine (Baltimore). 2020;99(11):e19433. doi:10.1097/MD.0000000000019433
84. Karpathiou G, Chauleur C, Mobarki M, Peoc’h M. The immune checkpoints CTLA-4 and PD-L1 in carcinomas of the uterine cervix. Pathol Res Pract. 2020;216(1):152782. doi:10.1016/j.prp.2019.152782
85. Yang W, Song Y, Lu YL, Sun JZ, Wang HW. Increased expression of programmed death (PD)-1 and its ligand PD-L1 correlates with impaired cell-mediated immunity in high-risk human papillomavirus-related cervical intraepithelial neoplasia. Immunology. 2013;139(4):513–522. doi:10.1111/imm.12101
86. Heeren AM, Rotman J, Stam AGM, et al. Efficacy of PD-1 blockade in cervical cancer is related to a CD8(+)FoxP3(+)CD25(+) T-cell subset with operational effector functions despite high immune checkpoint levels. J Immunother Cancer. 2019;7(1):43. doi:10.1186/s40425-019-0526-z
87. Dorta-Estremera S, Hegde VL, Slay RB, et al. Targeting interferon signaling and CTLA-4 enhance the therapeutic efficacy of anti-PD-1 immunotherapy in preclinical model of HPV(+) oral cancer. J Immunother Cancer. 2019;7(1):252. doi:10.1186/s40425-019-0728-4
88. Lheureux S, Butler MO, Clarke B, et al. Association of ipilimumab with safety and antitumor activity in women with metastatic or recurrent human papillomavirus-related cervical carcinoma. JAMA Oncol. 2018;4(7):e173776. doi:10.1001/jamaoncol.2017.3776
89. Rotman J, Mom CH, Jordanova ES, de Gruijl TD, Kenter GG. ‘DURVIT’: a phase-I trial of single low-dose durvalumab (Medi4736) IntraTumourally injected in cervical cancer: safety, toxicity and effect on the primary tumour- and lymph node microenvironment. BMC Cancer. 2018;18(1):888. doi:10.1186/s12885-018-4764-0
90. Jung KH, LoRusso P, Burris H, et al. Phase I study of the indoleamine 2,3-dioxygenase 1 (IDO1) inhibitor navoximod (GDC-0919) administered with PD-L1 inhibitor (atezolizumab) in advanced solid tumors. Clin Cancer Res. 2019;25(11):3220–3228. doi:10.1158/1078-0432.CCR-18-2740
91. Mayadev J, Zamarin D, Deng W, et al. Anti-PD-L1 (atezolizumab) as an immune primer and concurrently with extended-field chemoradiotherapy for node-positive locally advanced cervical cancer. Int J Gynecol Cancer. 2019;30.
92. Chinn Z, Stoler MH, Mills AM. PD-L1 and IDO expression in cervical and vulvar invasive and intraepithelial squamous neoplasias: implications for combination immunotherapy. Histopathology. 2019;74(2):256–268. doi:10.1111/his.13723
93. Hamanishi J, Mandai M, Ikeda T, et al. Safety and antitumor activity of anti-PD-1 antibody, nivolumab, in patients with platinum-resistant ovarian cancer. J Clin Oncol. 2015;33(34):4015–4022. doi:10.1200/JCO.2015.62.3397
94. Huang RY, Francois A, McGray AR, Miliotto A, Odunsi K. Compensatory upregulation of PD-1, LAG-3, and CTLA-4 limits the efficacy of single-agent checkpoint blockade in metastatic ovarian cancer. Oncoimmunology. 2017;6(1):e1249561. doi:10.1080/2162402X.2016.1249561
95. Thangarajah F, Morgenstern B, Pahmeyer C, et al. Clinical impact of PD-L1 and PD-1 expression in squamous cell cancer of the vulva. J Cancer Res Clin Oncol. 2019;145(6):1651–1660. doi:10.1007/s00432-019-02915-1
96. Zacharakis N, Chinnasamy H, Black M, et al. Immune recognition of somatic mutations leading to complete durable regression in metastatic breast cancer. Nat Med. 2018;24(6):724–730. doi:10.1038/s41591-018-0040-8
97. Zhang R, Zhang Z, Liu Z, et al. Adoptive cell transfer therapy for hepatocellular carcinoma. Front Med. 2019;13(1):3–11. doi:10.1007/s11684-019-0684-x
98. van Poelgeest MI, Visconti VV, Aghai Z, et al. Potential use of lymph node-derived HPV-specific T cells for adoptive cell therapy of cervical cancer. Cancer Immunol Immunother. 2016;65(12):1451–1463. doi:10.1007/s00262-016-1892-8
99. McCormack SE, Cruz CRY, Wright KE, et al. Human papilloma virus-specific T cells can be generated from naive T cells for use as an immunotherapeutic strategy for immunocompromised patients. Cytotherapy. 2018;20(3):385–393. doi:10.1016/j.jcyt.2017.11.010
100. Schuster SJ, Svoboda J, Chong EA, et al. Chimeric antigen receptor T cells in refractory B-cell lymphomas. N Engl J Med. 2017;377(26):2545–2554. doi:10.1056/NEJMoa1708566
101. He Y, Li XM, Yin CH, Wu YM. Killing cervical cancer cells by specific chimeric antigen receptor-modified T cells. J Reprod Immunol. 2020;139:103115. doi:10.1016/j.jri.2020.103115
102. Li N, Tian YW, Xu Y, et al. Combined treatment with autologous CIK cells, radiotherapy and chemotherapy in advanced cervical cancer. Pathol Oncol Res. 2019;25(2):691–696. doi:10.1007/s12253-018-0541-2
103. Veluchamy JP, Heeren AM, Spanholtz J, et al. High-efficiency lysis of cervical cancer by allogeneic NK cells derived from umbilical cord progenitors is independent of HLA status. Cancer Immunol Immunother. 2017;66(1):51–61. doi:10.1007/s00262-016-1919-1
104. Langers I, Renoux V, Reschner A, et al. Natural killer and dendritic cells collaborate in the immune response induced by the vaccine against uterine cervical cancer. Eur J Immunol. 2014;44(12):3585–3595. doi:10.1002/eji.201444594
105. Choi YW, Kang MC, Seo YB, et al. Intravaginal administration of Fc-fused IL7 suppresses the cervicovaginal tumor by recruiting HPV DNA vaccine-induced CD8 T cells. Clin Cancer Res. 2016;22(23):5898–5908. doi:10.1158/1078-0432.CCR-16-0423
106. Grunwitz C, Salomon N, Vascotto F, et al. HPV16 RNA-LPX vaccine mediates complete regression of aggressively growing HPV-positive mouse tumors and establishes protective T cell memory. Oncoimmunology. 2019;8(9):e1629259. doi:10.1080/2162402X.2019.1629259
107. Baghban Rahimi S, Mohebbi A, Vakilzadeh G, et al. Enhancement of therapeutic DNA vaccine potency by melatonin through inhibiting VEGF expression and induction of antitumor immunity mediated by CD8+ T cells. Arch Virol. 2018;163(3):587–597. doi:10.1007/s00705-017-3647-z
108. Atherton MJ, Stephenson KB, Nikota JK, et al. Preclinical development of peptide vaccination combined with oncolytic MG1-E6E7 for HPV-associated cancer. Vaccine. 2018;36(16):2181–2192. doi:10.1016/j.vaccine.2018.02.070
109. Yang Y, Che Y, Zhao Y, Wang X. Prevention and treatment of cervical cancer by a single administration of human papillomavirus peptide vaccine with CpG oligodeoxynucleotides as an adjuvant in vivo. Int Immunopharmacol. 2019;69:279–288. doi:10.1016/j.intimp.2019.01.024
110. Dorta-Estremera S, Chin RL, Sierra G, et al. Mucosal HPV E6/E7 peptide vaccination in combination with immune checkpoint modulation induces regression of HPV(+) oral cancers. Cancer Res. 2018;78(18):5327–5339. doi:10.1158/0008-5472.CAN-18-0892
111. Vici P, Pizzuti L, Mariani L, et al. Targeting immune response with therapeutic vaccines in premalignant lesions and cervical cancer: hope or reality from clinical studies. Expert Rev Vaccines. 2016;15(10):1327–1336. doi:10.1080/14760584.2016.1176533
112. Trimble CL, Morrow MP, Kraynyak KA, et al. Safety, efficacy, and immunogenicity of VGX-3100, a therapeutic synthetic DNA vaccine targeting human papillomavirus 16 and 18 E6 and E7 proteins for cervical intraepithelial neoplasia 2/3: a randomised, double-blind, placebo-controlled phase 2b trial. Lancet. 2015;386(10008):2078–2088. doi:10.1016/S0140-6736(15)00239-1
113. Barra F, Della Corte L, Noberasco G, et al. Advances in therapeutic vaccines for treating human papillomavirus-related cervical intraepithelial neoplasia. J Obstet Gynaecol Res. 2020;46(7):989–1006. doi:10.1111/jog.14276
114. Weber JS, Yang JC, Atkins MB, Disis ML. Toxicities of Immunotherapy for the Practitioner. J Clin Oncol. 2015;33(18):2092–2099. doi:10.1200/JCO.2014.60.0379
115. Lagunas-Cruz MDC, Valle-Mendiola A, Trejo-Huerta J, et al. IL-2 induces transient arrest in the G1 phase to protect cervical cancer cells from entering apoptosis. J Oncol. 2019;2019:7475295. doi:10.1155/2019/7475295
116. Morgan EL, Macdonald A, Galloway DA. Autocrine STAT3 activation in HPV positive cervical cancer through a virus-driven Rac1-NFκB-IL-6 signalling axis. PLoS Pathog. 2019;15(6):e1007835. doi:10.1371/journal.ppat.1007835
117. Rangel-Corona R, Corona-Ortega T, Soto-Cruz I, et al. Evidence that cervical cancer cells secrete IL-2, which becomes an autocrine growth factor. Cytokine. 2010;50(3):273–277. doi:10.1016/j.cyto.2010.02.013
118. Du GH, Wang JK, Richards JR, Wang JJ. Genetic polymorphisms in tumor necrosis factor alpha and interleukin-10 are associated with an increased risk of cervical cancer. Int Immunopharmacol. 2019;66:154–161. doi:10.1016/j.intimp.2018.11.015
119. Borgogna JC, Shardell MD, Santori EK, et al. The vaginal metabolome and microbiota of cervical HPV-positive and HPV-negative women: a cross-sectional analysis. Bjog. 2019;127:182–192.
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.