Back to Journals » Drug Design, Development and Therapy » Volume 20
Cellular and Molecular Mechanisms of Hyperglycemia-Induced Atherosclerosis and Intervention Strategies of Chinese Herbal Medicine
Authors Luo S ![]() , Zhu J, Zeng Q, Yang L
, Zhu J, Zeng Q, Yang L ![]() , Liu J, Deng Y
, Liu J, Deng Y
Received 9 February 2026
Accepted for publication 31 March 2026
Published 23 April 2026 Volume 2026:20 602453
DOI https://doi.org/10.2147/DDDT.S602453
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Solomon Tadesse Zeleke
Shengping Luo,1,2 Jiayu Zhu,3 Qianru Zeng,4 Le Yang,1,2 Jing Liu,1,2 Yihui Deng4
1School of Integrated Chinese and Western Medicine, Hunan University of Chinese Medicine, Changsha, People’s Republic of China; 2Hunan Province Key Laboratory of Cerebrovascular Disease Prevention and Treatment of Integrated Traditional Chinese and Western Medicine, Hunan University of Chinese Medicine, Changsha, People’s Republic of China; 3The First Clinical College of Chinese Medicine, Hunan University of Chinese Medicine, Changsha, People’s Republic of China; 4School of Traditional Chinese Medicine, Hunan University of Chinese Medicine, Changsha, People’s Republic of China
Correspondence: Yihui Deng, School of Traditional Chinese Medicine, Hunan University of Chinese Medicine, Changsha, People’s Republic of China, Email [email protected]
Abstract: Diabetes is a significant global public health issue, and atherosclerosis serves as the primary pathological basis for the occurrence and progression of cardiovascular and cerebrovascular events. Studies have shown that hyperglycemia can promote the occurrence and progression of atherosclerosis by damaging vascular endothelial cells, macrophages, and vascular smooth muscle cells. In recent years, Chinese herbal medicines and their active ingredients have demonstrated unique advantages in preventing and treating high-glucose-induced vascular cell damage. Their mechanisms of action primarily encompass two aspects: firstly, regulating blood glucose through multiple pathways such as modulating the insulin signaling pathway, enhancing peripheral glucose uptake and utilization, and delaying intestinal carbohydrate absorption; secondly, intervening in key pathological processes such as high-glucose-induced vascular cell inflammation, cell death, phenotypic transformation, and metabolic reprogramming. This, in turn, slows down the progression of diabetes-related atherosclerosis. This article systematically elucidates the molecular mechanisms underlying hyperglycemia-induced damage to vascular endothelial cells, macrophages, and vascular smooth muscle cells. It also summarizes the protective mechanisms of Chinese herbal medicine active ingredients (albiflorin, salvianolic acid B, scutellarin, vitexin, hydroxysafflor yellow A, ginsenoside Rb1, maslinic acid, paeonol, citronellal, matrine, Panax notoginseng saponins, astragalus polysaccharides, 6-gingerol, and sodium tanshinone IIA sulfonate) and traditional Chinese medicine compounds (buyang huanwu decoction, guanxining tablets, and fufang zhenzhu tiaozhi) against vascular endothelial cells, macrophages, and vascular smooth muscle cells. The aim is to provide a theoretical basis for the prevention and treatment of diabetes-related atherosclerosis, as well as the development of related therapeutic drugs.
Keywords: hyperglycemia, atherosclerosis, Chinese herbal medicine, endothelial cells, macrophages, vascular smooth muscle cells, molecular mechanisms, protective effect
Introduction
Diabetes mellitus is a metabolic disease characterized by chronic hyperglycemia, caused by defects in insulin secretion or action. Global disease burden data show that there were 536 million people with diabetes in 2021, and this is expected to rise to 783 million by 2045.1,2 Due to long-term hyperglycemic states and chronic inflammation, diabetes can lead to numerous complications. Among these, diabetic cardiovascular and cerebrovascular complicatio46ns are the leading cause of death in patients with type 2 diabetes, and about 90% of patients with diabetic cardiovascular and cerebrovascular diseases suffer from atherosclerotic cardiovascular disease.3 Atherosclerosis is a chronic inflammatory disease characterized by local lipid deposition, fibrous tissue proliferation, and calcium deposition, forming plaques, reducing vascular elasticity, and leading to vascular wall hardening.4 Research has found that compared to the general population, patients with diabetes exhibit earlier and more rapid progression of atherosclerosis; furthermore, plaques in diabetic patients often possess larger necrotic cores and more pronounced inflammation, leading to plaque instability and an increased risk of plaque rupture, which in turn can trigger acute clinical complications of atherosclerosis.5,6
The mechanism by which hyperglycemia exacerbates atherosclerosis is relatively complex, encompassing inflammatory response, oxidative stress, vascular proliferation, stromal changes, and vascular cell damage.7–10 The clinical efficacy of chinese herbal medicines in treating diabetes-associated atherosclerosis has been confirmed.11 Animal and cell experiments further reveal that its effects are attributed to a dual mechanism: on the one hand, it regulates glucose metabolism by modulating the insulin signaling pathway, enhancing peripheral glucose uptake and utilization, and delaying intestinal carbohydrate absorption, thereby eliminating hyperglycemia as the fundamental cause;12 on the other hand, it intervenes in the core pathological process of hyperglycemia-induced vascular cell damage, blocking the cascade reaction of “hyperglycemia-vascular damage atherosclerosis progression”13 For example, albiflorin, an active ingredient extracted from white Baishao (Paeonia lactiflora Pall), can promote glucose uptake in skeletal muscle cells and regulate insulin signaling, thereby reducing blood glucose levels and eliminating the damaging factor of hyperglycemia. Additionally, it can inhibit the release of inflammatory factors induced by hyperglycemia, reduce endothelial cell apoptosis, and thus mitigate hyperglycemia-induced damage to human umbilical vein endothelial cells, thereby delaying the progression of diabetes-associated atherosclerosis.13,14 Therefore, this article focuses on the damage to three types of vascular cells: endothelial cells, macrophages, and smooth muscle cells, summarizing the molecular mechanisms by which hyperglycemia exacerbates atherosclerosis. Furthermore, this article systematically reviews the mechanisms by which Chinese herbal medicine and its active ingredients regulate blood glucose and hyperglycemia-induced vascular cell damage, thereby improving diabetes-associated atherosclerosis. This may provide new directions for future research and aid in the further development of candidate drugs.
Review Methodology
We employed a systematic literature search methodology to identify high-quality studies, with coverage spanning authoritative academic platforms such as PubMed and Web of Science, to track research progress in this field comprehensively. The specific retrieval formula as follows: literature search strategy=(“Hyperglycemia” OR “Diabetes Mellitus” OR “High glucose”) AND (“Atherosclerosis” OR “Arteriosclerosis” OR “Plaque, Atherosclerotic” OR “vascular plaque”) AND (“Endothelial Cell*” OR “Vascular Endothelial Cell*” OR “Macrophage*” OR “Muscle, Smooth, Vascular” OR “vascular smooth muscle cell*”) AND (“Medicine, Chinese Traditional” OR “Chinese Traditional Medicine*” OR “Traditional Chinese Medicine*” OR “Chinese herbal medicine*” OR “active compound*” OR “metabolite*” OR “active component*” OR “Natural product*”). This search strategy focused on mechanistic studies investigating chinese herbal medicines and their active components in intervening against high-glucose-induced injury to endothelial cells, macrophages, and vascular smooth muscle cells to prevent and treat diabetes-associated atherosclerosis. No time restrictions were applied to publication selection, and publications with low relevance and non-English-language publications were excluded. To strictly control selection bias and ensure objectivity in screening results, two authors independently conducted the initial literature screening following systematic review protocols, with cross-checking and verification performed upon completion. Any disagreements were resolved through third-party adjudication to reach a consensus. The search strategy was implemented across multiple databases in accordance with PRISMA guidelines (PRISMA 2020 citation). The detailed literature screening and inclusion process is illustrated in Figure 1.
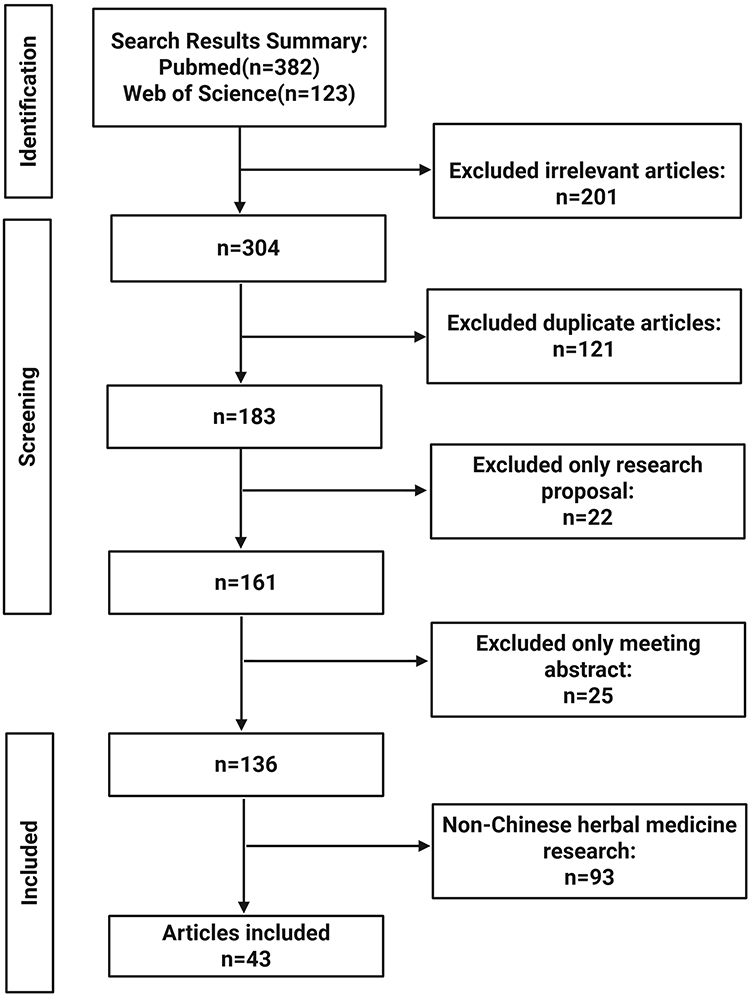 |
Figure 1 The detailed process of literature screening. Conduct a literature search using search strategy in the PubMed and Web of Science, remove duplicates and irrelevant literature to the research topic, and conduct an in-depth study of the included literature. |
Mechanism of Hyperglycemia-Induced Damage to Vascular Cells Accelerating Atherosclerosis
Mechanism of Hyperglycemia-Induced Endothelial Cell Damage
Endothelial cells (ECs) are a monolayer lining the inner surface of blood vessels. They play a central role in maintaining cardiovascular homeostasis by forming a selective barrier, regulating vascular tone, and inhibiting coagulation and inflammation.15–17 Studies have demonstrated that hyperglycemia is an independent risk factor for atherosclerosis, with its pathogenic effects originating from the functional disturbances of vascular endothelial cells.18 In the presence of persistent elevated glucose levels, endothelial cells exhibit a series of pathological alterations, including the upregulation of pro-inflammatory adhesion molecules, the uncoupling of endothelial nitric oxide synthase (eNOS), resulting in diminished bioavailability of nitric oxide (NO), augmented oxidative stress, as well as cell death and senescence.19–22 The combined consequences of these changes are disruption of endothelial barrier integrity and a significant increase in permeability, creating the initial conditions for the deposition of low-density lipoprotein (LDL) beneath the endothelium and the infiltration of macrophages. Consequently, hyperglycemia-induced endothelial dysfunction is recognized as the “initiator” of the atherosclerotic process, signifying the pivotal initial connection between metabolic disorders and vascular pathology. The mechanism of hyperglycemia-induced endothelial cell injury is illustrated in Figure 2.
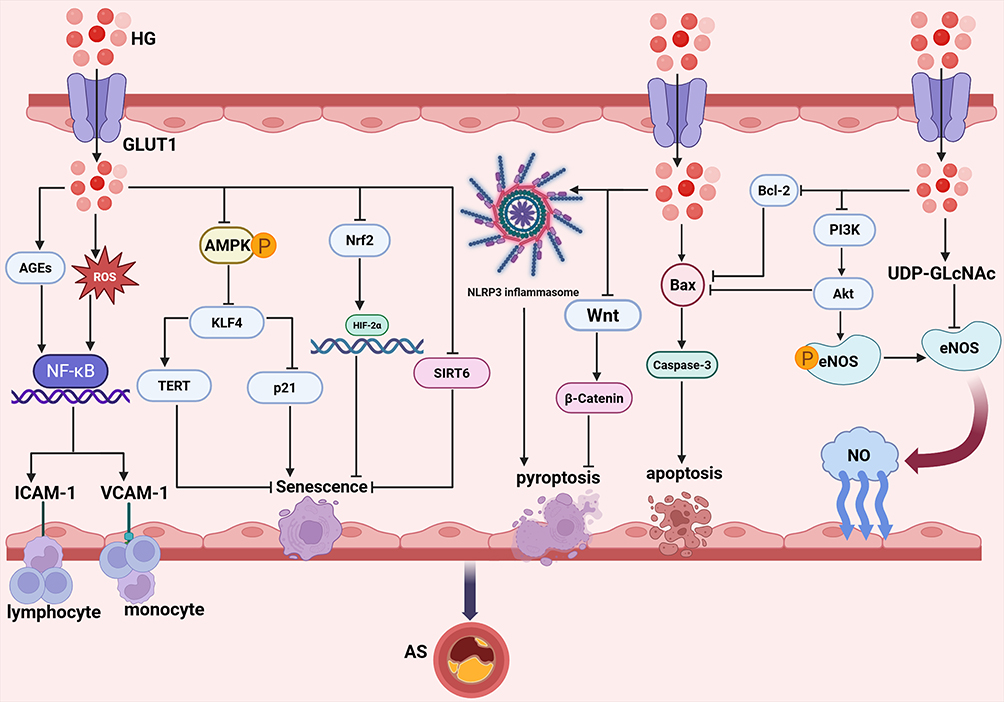 |
Figure 2 Molecular mechanisms of hyperglycemia-driven endothelial dysfunction promoting atherosclerosis. Hyperglycemia (HG) enters endothelial cells via GLUT1 and activates multiple pathological signaling pathways. On the one hand, HG promotes the generation of ROS and AGEs, activating the NF-κB signaling pathway and upregulating the expression of ICAM-1 and VCAM-1, thereby facilitating the adhesion of monocytes and lymphocytes. Additionally, HG activates the hexosamine pathway to produce UDP-GlcNAc, which inhibits eNOS activity and significantly reduces NO production. Moreover, HG suppresses the PI3K/Akt/eNOS pathway, decreasing eNOS phosphorylation levels and reducing both eNOS activity and NO generation. Moreover, HG can directly upregulate Bax expression and downregulate Bcl-2 protein, thereby promoting apoptosis. Simultaneously, HG can inhibit the Wnt/β-catenin pathway, thereby inducing pyroptosis. In addition, HG can also activate the NLRP3 inflammasome to induce pyroptosis. On the other hand, HG inhibits AMPK phosphorylation, thereby relieving the suppression of KLF4. The activation of KLF4 can relieve the inhibition of p21 and suppress TERT activity, ultimately inducing cellular senescence. Furthermore, HG can disrupt the interaction between Nrf2 and HIF-2α, thereby inducing endothelial senescence. Meanwhile, HG can also directly downregulate SIRT6 expression, promoting cellular senescence. Under the combined effects of these multiple mechanisms of high glucose on endothelial cells, vascular endothelial integrity is compromised, accelerating the formation and progression of diabetes-associated atherosclerosis. |
eNOS Activity Decline
eNOS is a calcium-dependent enzyme predominantly located in vascular endothelial cells. Under normal physiological conditions, eNOS catalyzes the conversion of L-arginine to NO, which subsequently exerts multiple physiological functions, including regulating vascular tone and maintaining vascular dilation.23 In addition to these effects, NO has been shown to inhibit platelet aggregation, suppress inflammatory responses, and promote angiogenesis.24,25 However, under certain pathological conditions, eNOS may alter its activity and leading to a significant reduction in NO production. Research has demonstrated that under conditions of hyperglycemia, the hexosamine pathway is overactivated, which inhibits the activity of eNOS, thereby reducing the production of NO.26 Furthermore, the hyperglycemia environment has been shown to reduce the phosphorylation levels of protein kinase B (AKT) and phosphatidylinositol 3-kinase (PI3K), thereby inhibiting the activity of the PI3K/AKT signaling pathway. This phenomenon also results in a decrease in eNOS phosphorylation, which, in turn, leads to the suppression of eNOS activity and, consequently, a further reduction in NO production.27
The reduction in NO production can trigger a cascade of adverse consequences. On the one hand, the anti-inflammatory capacity of endothelial cells is diminished, resulting in the infiltration of inflammatory cells into the vascular wall. Conversely, a reduction in NO production has been shown to impair endothelial cell repair, resulting in a decrease in their number and, consequently, a compromise in the integrity of the vascular endothelium and a weakening of the vascular barrier.28,29 The synergistic effect of these factors can lead to a heightened rate of atherosclerosis progression.
Upregulation of Adhesion Molecule Expression
Chronic inflammation is a hallmark of hyperglycemia. Hyperglycemia has been demonstrated to induce the production of substantial quantities of reactive oxygen species (ROS) and advanced glycation end products (AGEs) within the body. These phenomena have been observed to activate the NF-κB pathway.30,31 Upon activation, NF-κB initiates the transcription of a series of pro-inflammatory genes, including those encoding intercellular adhesion molecule-1 (ICAM-1) and vascular cell adhesion molecule-1 (VCAM-1). ICAM-1 and VCAM-1 are both expressed at high levels on the surface of endothelial cells. These adhesion molecules function as “hooks” on the rails, enabling circulating monocytes and lymphocytes to firmly adhere to the typically smooth vascular endothelium by binding to ICAM-1 and VCAM-1.32 Subsequent to adhesion, leukocytes undergo further activation, resulting in the release of additional inflammatory cytokines. These cytokines have been shown to recruit additional immune cells to the site, thereby exacerbating the local inflammatory response and promoting plaque progression.33
Endothelial Cell Death and Senescence
Cell death is a significant phenomenon in life processes, playing a pivotal role in the development and maintenance of physiological homeostasis and health in organisms. The phenomenon of cell death can be broadly categorized into two distinct classifications: programmed cell death and non-programmed cell death. Programmed cell death, otherwise known as apoptosis and pyroptosis, contrasts with non-programmed cell death, which is primarily characterized by necrosis.34 During the progression of atherosclerotic lesions, a hyperglycemic microenvironment induces aberrant endothelial cell death, which impairs endothelial integrity. Consequently, a substantial amount of LDL in the bloodstream infiltrates the subendothelial space through these defective sites, undergoes oxidation to form oxidized low-density lipoprotein (ox-LDL), and accumulates as lipid streaks—the pathological core of atherosclerosis-thereby accelerating disease progression.35 Research has demonstrated that hyperglycemia treatment significantly inhibits the activity of the PI3K/Akt signaling pathway in human umbilical vein endothelial cells (HUVECs). Concurrently, the study found that hyperglycemia treatment upregulates the expression of caspase 3 and Bcl-2-associated X protein (Bax) proteins and downregulates the level of the B cell lymphoma-2 (Bcl-2) protein. The result of these changes is the promotion of apoptosis in HUVECs.27 Additionally, hyperglycemia has been reported to further increase apoptosis in HUVECs by inhibiting the Wnt/β-catenin pathway.36 Moreover, hyperglycemia has also been found to activate the NOD-like receptor family pyrin domain-containing 3 (NLRP3) inflammasome, thereby inducing pyroptosis in endothelial cells.37
Cellular senescence is an inevitable natural process in organisms. It serves as a self-protective mechanism of cells in response to external stimuli and is also closely associated with organismal aging and the development of various chronic diseases.19
It has been reported that high glucose induced cellular senescence exerts a direct pro-atherosclerotic effect.38 Endothelial cell senescence is characterized by permanent arrest of cell growth and proliferation. Pathological studies have demonstrated that senescent endothelial cells accumulate in the subendothelial space at the onset of AS, driving disease pathogenesis by upregulating the expression of key pro-atherosclerotic inflammatory cytokines and chemokines, thereby acquiring a phenotype termed the senescence-associated secretory phenotype.39,40 Furthermore, due to impaired cell proliferation, permeability, and motility, endothelial cell senescence compromises endothelial barrier function, which may lead to endothelial erosion and intraplaque hemorrhage.41,42 Research has demonstrated that a hyperglycemia environment has the capacity to impede AMP-activated kinase (AMPK) phosphorylation, consequently diminishing its inhibitory effect on Krüppel-like factor 4 (KLF4) and resulting in augmented KLF4 expression. As a transcription factor, KLF4 has been observed to exert dual regulatory functions, namely the activation of the pro-senescence gene p21 and the repression of telomerase reverse transcriptase (TERT). This phenomenon ultimately leads to the senescence of HUVECs and the expansion of the area occupied by atherosclerotic plaques.43 Additionally, it has been reported that hyperglycemia can downregulate Sirtuin type-6 (SIRT6) expression, thereby inducing endothelial cell senescence.44 Furthermore, the available evidence suggests that elevated glucose levels can interfere with the interaction between nuclear factor erythroid-2 related factor 2 (Nrf2) and hypoxia-inducible factor-2α (HIF-2α), consequently weakening the inhibition of HIF-2α’s pro-oxidative effects. Consequently, this results in the activation of senescence pathways, which in turn triggers endothelial cell senescence and promotes the development and progression of diabetic vascular complications.45
Mechanism of Hyperglycemia-Induced Damage to Macrophages
Foam cells represent the hallmark pathological feature of atherosclerosis. These macrophage-derived cells, characterized by abundant cytoplasmic lipid droplets, constitute the lipid core of atherosclerotic plaques. Previous studies have established that the excessive uptake of LDL by macrophages via scavenger receptors is the central mechanism underlying foam cell formation.46 Emerging evidence indicates that hyperglycemia can further promote lipid accumulation and foam cell transformation in macrophages on this basis, thereby accelerating the progression of atherosclerotic lesions.47 Elucidating the mechanisms by which hyperglycemia drives macrophage foam cell formation will provide novel targets for atherosclerosis intervention strategies targeting the metabolism-immunity interface. The mechanism of hyperglycemia-induced Macrophages injury is illustrated in Figure 3.
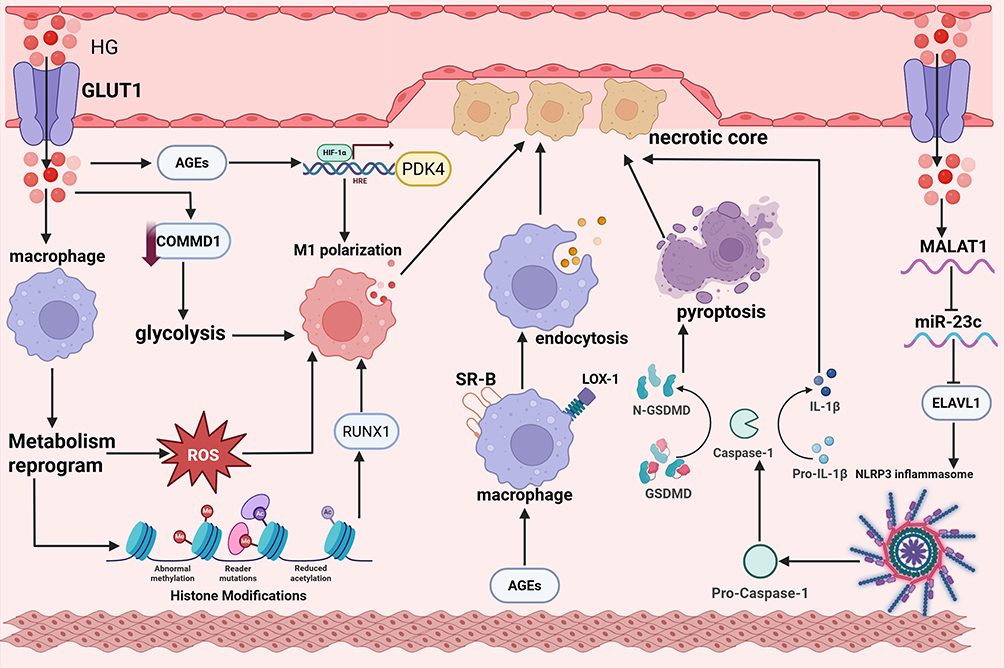 |
Figure 3 Molecular mechanisms of hyperglycemia-driven macrophage dysfunction promoting atherosclerosis. Upon entering the vascular wall via GLUT1, HG induces macrophage metabolic reprogramming: On one hand, HG promotes ROS generation, inducing macrophage M1 polarization; on the other hand, through histone modification-mediated epigenetic reprogramming, HG activates the RUNX1 transcription factor, driving macrophage polarization toward the pro-inflammatory M1 phenotype. Additionally, HG induces COMMD1 deficiency, enhances the glycolytic pathway, and thereby promotes macrophage M1 polarization. Meanwhile, HG also promotes AGE formation; AGEs further enhance M1 polarization via the HIF-1α/PDK4 pathway, and can bind to macrophage surface receptors such as SR-B and LOX-1, enhancing macrophage endocytosis and promoting lipid uptake. Simultaneously, HG can also upregulate MALAT1 expression, inhibit miR-23c expression, relieve the suppression of ELAVL1, thereby promoting NLRP3 inflammasome activation, activating Caspase-1-dependent GSDMD cleavage, inducing pyroptosis, and the release of inflammatory factors such as IL-1β. Ultimately, these pathological processes promote the development and progression of necrotic cores in diabetes-associated atherosclerosis. |
Metabolic Reprogramming
Macrophage polarization is defined as the process by which macrophages acquire distinct functional phenotypes and stably express corresponding marker molecules in response to signals from the microenvironment.48 The categorisation of this condition is primarily into M1 and M2 subtypes. M1-type macrophages have the capacity to secrete substantial quantities of proinflammatory cytokines, thereby exacerbating inflammatory responses. Furthermore, M1 macrophages have also been shown to produce matrix metalloproteinases. These enzymes have the capacity to degrade the fibrous cap of atherosclerotic plaques, thereby impairing plaque stability and increasing the risk of plaque rupture.49 Upon uptake of ox-LDL, M1-type macrophages undergo a transformation into foam cells. These foam cells accumulate within the arterial wall, forming the core of atherosclerotic plaques. The accumulation of foam cells has been demonstrated to augment plaque volume, thereby amplifying the inflammatory response.50 Conversely, M2-type macrophages have been shown to exhibit anti-inflammatory properties, thereby facilitating tissue repair and angiogenesis.51 Furthermore, M2-type macrophages exhibit robust phagocytic activity, enabling them to clear apoptotic cells and cellular debris, which in turn contributes to atherosclerotic plaque stabilisation.52
In recent years, the field of immunometabolism has undergone rapid development, leading to a significant increase in research activity. This has included research into metabolic reprogramming in the context of cardiovascular diseases. Glucose metabolic reprogramming, in particular, has attracted considerable attention due to its role as a regulatory node. Studies have confirmed that the atherosclerotic plaque microenvironment is characterized by hyperglycemia, hypoxia, and lipid enrichment. Macrophages infiltrating this microenvironment undergo an irreversible metabolic pattern switch—specifically, shifting from mitochondrial-dependent oxidative phosphorylation to aerobic glycolysis.53,54 This metabolic reprogramming event has been confirmed to be a crucial driver regulating the polarization of macrophages toward a proinflammatory phenotype, thereby accelerating the formation and unstable progression of atherosclerosis plaques.55
Previous studies have demonstrated that hyperglycemia can significantly enhance cellular glucose uptake capacity by upregulating the expression level of glucose transporter 1 (GLUT1) on the macrophage surface, ultimately inducing glucose metabolic reprogramming in macrophages.56,57 This process is accompanied by increased ROS production, which in turn drives the activation of macrophages toward a proinflammatory phenotype. In addition, deficiency of copper metabolism domain-containing protein 1 (COMMD1) is also a critical predisposing factor for diabetes-associated atherosclerosis. The underlying mechanism involves the direct promotion of glycolysis pathway activation in macrophages, which in turn enhances the pro-inflammatory activation of macrophages and foam cell formation. Notably, 2-deoxy-D-glucose (2-DG), a glycolysis-specific inhibitor, can effectively reverse the aforementioned pathological effects, further corroborating the central role of glucose metabolic reprogramming in this pathological process.58 A study by Edgar59 et al further expands this understanding, demonstrating that hyperglycemia can not only induce a shift in aerobic glycolysis-mediated metabolic reprogramming in mature macrophages but also in their precursor hematopoietic stem cells. This metabolic switch triggers histone modification-mediated epigenetic reprogramming, which activates the transcription factor Runt-related transcription factor 1 (RUNX1) to regulate the sustained expression of downstream pro-inflammatory genes, ultimately leading to macrophages maintaining a pro-atherosclerotic phenotype. Furthermore, when the research team transplanted bone marrow from diabetic mice into atherosclerosis-susceptible mice with normoglycemia, they found that this transplantation exacerbated the severity of atherosclerotic lesions at the aortic root in recipient mice. Additionally, they validated the conservation of the aforementioned metabolic and epigenetic regulatory patterns in human peripheral blood mononuclear cells and macrophages derived from atherosclerotic plaques. This finding suggests that hyperglycemia can induce trained immunity in macrophages and their progenitor hematopoietic stem cells via a glycolysis-dependent mechanism. This provides a crucial mechanistic explanation for the observation that clinical hypoglycemic therapy is challenging to effectively reduce the risk of macrovascular complications in diabetic patients.
Accumulation of Advanced Glycation End Products
AGEs are heterogeneous, irreversible adducts generated by the non-enzymatic glycation and oxidation of proteins, lipids, and nucleic acids.60 In diabetic patients, chronic hyperglycemia promotes the formation and accumulation of AGEs, thereby facilitating the development and progression of diabetic vascular complications.61 Studies have demonstrated that AGEs enhance M1 polarization of macrophages via hypoxia-inducible factor-1α (HIF-1α)/Pyruvate dehydrogenase kinase 4 (PDK4) pathway, thereby accelerating the progression of diabetes-associated atherosclerosis. Notably, this inductive effect exhibits concentration- and time-dependent characteristics.62 Furthermore, AGEs can enhance the endocytosis of macrophages, thereby increasing lipid uptake. This effect is attributed to the interaction of AGEs with macrophage surface receptors (eg, scavenger receptors class B (SR-B) and lectin-like oxidized low-density lipoprotein receptor 1 [LOX-1]), which upregulates the expression of these receptors.63
Macrophage Death
Accumulating research evidence indicates that the aberrant death of macrophages contributes to the progression of atherosclerosis. This is primarily attributed to the massive release of pro-inflammatory cytokines from dying macrophages, which exacerbates the instability of atherosclerotic plaques.64,65 It has been reported that high glucose can regulate macrophage death. Hyperglycemia upregulates the expression of the long non-coding Ribonucleic Acid (RNA) metastasis-associated lung adenocarcinoma transcript 1 (MALAT1). MALAT1 specifically binds to and inhibits the function of miR-23c, which normally targets and suppresses the expression of ELAV like RNA binding protein 1 (ELAVL1) by direct binding. The sequestration of miR-23c by MALAT1 impairs its inhibitory activity, thereby relieving the repression of ELAVL1 and leading to a marked elevation in ELAVL1 protein levels. As a positive regulator of the NLRP3 inflammasome, the upregulated ELAVL1 directly activates the NLRP3 inflammasome complex. Once activated, the inflammasome cleaves pro-caspase-1 into its biologically active form, caspase-1. On the one hand, active caspase-1 induces macrophage pyroptosis; on the other hand, it promotes the maturation and secretion of the pro-inflammatory cytokine Interleukin-1β (IL-1β). These processes further amplify the inflammatory response and accelerate the progression of diabetes-associated atherosclerosis.66 The persistent inflammatory microenvironment and cell pyroptosis accelerate the formation of atherosclerotic plaques, thereby enhancing the progression of diabetes-related atherosclerosis.67
Mechanism of Hyperglycemia-Induced Damage to Vascular Smooth Muscle Cells
Vascular smooth muscle cells (VSMCs) play a pivotal role in determining vascular caliber, regulating blood pressure, and mediating local tissue perfusion.68 Dysfunction of VSMCs constitutes a fundamental mechanism underlying the pathogenesis of vascular diseases. Moreover, hyperglycemia can induce VSMCs dysfunction, thereby contributing to the development of vascular atherosclerosis. The mechanism of hyperglycemia-induced VSMCs injury is illustrated in Figure 4.
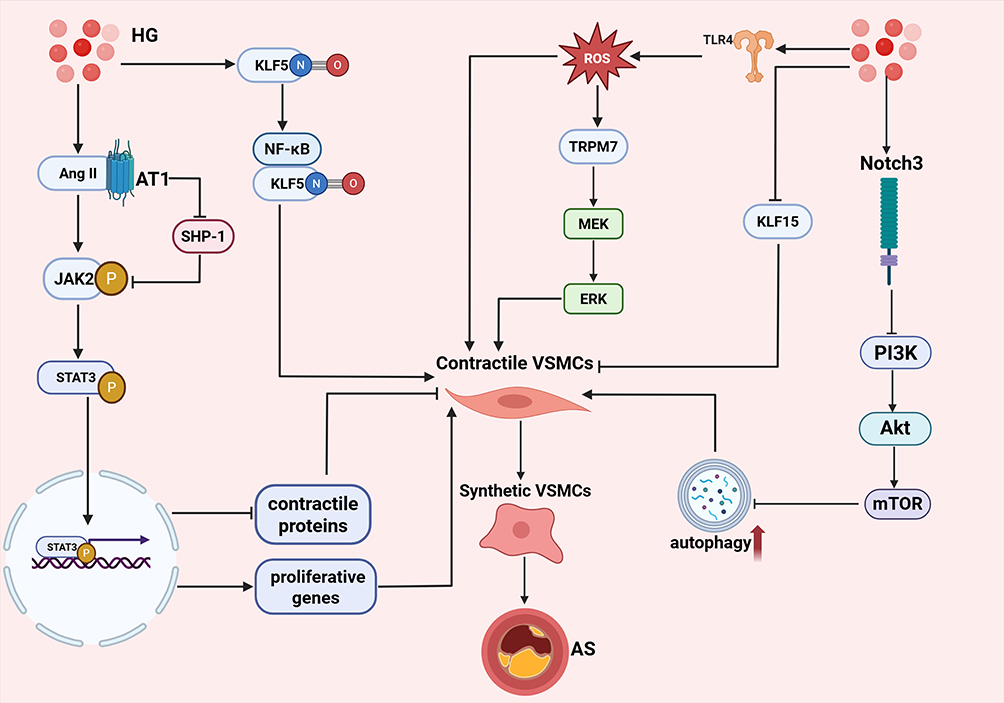 |
Figure 4 Molecular mechanisms of hyperglycemia-driven VSMCs dysfunction promoting atherosclerosis. On one hand, HG promotes Ang II binding to the AT1 receptor, activating the JAK2/STAT3 signaling pathway; phosphorylated STAT3 translocates into the nucleus to suppress contractile protein expression while upregulating proliferation genes, driving VSMCs phenotypic switching. On the other hand, HG induces tyrosine nitration of KLF5, promoting its interaction with NF-κB and increasing pro-inflammatory cytokine expression; meanwhile, HG downregulates KLF15 expression, relieving its negative regulation of phenotypic switching. Additionally, HG activates the TLR4 signaling pathway to promote excessive ROS generation; ROS upregulates TRPM7 expression and activates the MEK/ERK signaling pathway to facilitate VSMCs phenotypic switching, while accumulated ROS can also directly promote phenotypic transformation. Furthermore, HG upregulates Notch3 expression, inhibiting the PI3K/Akt/mTOR pathway and causing excessive autophagy activation, which further promotes VSMCs switching toward the synthetic phenotype. Ultimately, phenotypically transformed VSMCs exhibit enhanced proliferation, migration, and secretion capabilities, accelerating diabetes-associated atherosclerotic plaque progression. |
Migration, proliferation, and inflammation represent the major hallmarks of phenotypic changes in VSMCs. Under physiological conditions, VSMCs exist in a contractile phenotype, whose primary function is to maintain vascular tone and hemodynamic homeostasis.69 However, under hyperglycemic conditions, VSMCs rapidly initiate a dedifferentiation program and transdifferentiate into a synthetic/inflammatory phenotype. This phenotypic shift is characterized by a marked downregulation of contractile proteins, coupled with enhanced synthetic, secretory, and migratory capacities. Consequently, VSMCs become the major cellular source of the fibrous cap and neointima in atherosclerosis plaques.70,71
Activation of Janus kinase (JAK)/signal transducer and activator of transcription (STAT) signaling pathway represents one of the critical mechanisms underlying hyperglycemia-induced phenotypic switching of VSMCs. Under hyperglycemic conditions, angiotensin II (AngII) exerts a direct interaction with Janus kinase 2 (JAK2) via the angiotensin type 1 receptor (AT1), which promotes the tyrosine phosphorylation of JAK2 and inhibits the activity of Src homology region 2 domain-containing phosphatase-1 (SHP-1)-its negative regulator—thereby prolonging the phosphorylated state of JAK2.72,73 Subsequently, signal transducer and activator of transcription 3 (STAT3) is phosphorylated and translocates into the nucleus, where it acts as a transcription factor to regulate the expression of downstream target genes. Specifically, phosphorylated STAT3 suppresses myocardin-mediated expression of contractile proteins, while upregulating the expression of proliferative genes.74 These regulatory effects collectively induce the phenotypic transition of VSMCs from a contractile to a synthetic phenotype, and further enhance VSMCs proliferation and migration.
Krüppel-like factors (KLFs) also play a crucial role in hyperglycemia-induced phenotypic switching of VSMCs. VSMCs isolated from diabetic patients and murine models treated with hyperglycemia exhibit enhanced Krüppel-like factor 5 (KLF5) expression.75 Specifically, nitration modification occurs at the tyrosine residues of KLF5, which facilitates its interaction with NF-κB and thereby increases the expression of pro-inflammatory cytokines. Furthermore, studies76 have demonstrated that the aortas of mice treated with hyperglycemia exhibit decreased KLF15 mRNA expression, elevated levels of pro-inflammatory cytokines, and enhanced cellular proliferation. Notably, overexpression of KLF15 can reverse these aforementioned effects, suggesting that KLF15 may serve as a negative regulator of VSMCs phenotypic switching under hyperglycemic conditions.
Hyperglycemia also modulates the generation of ROS. Specifically, it promotes excessive ROS production by activating Toll-like receptor 4 (TLR4) signaling in VSMCs. Conversely, inhibition of TLR4 leads to a significant reduction in ROS levels in hyperglycemia-treated VSMCs.77 Excessive ROS accumulation can drive the proliferation, migration, and apoptosis of VSMCs, thereby facilitating the completion of phenotypic switching. Accumulating evidence has demonstrated that hyperglycemia induces oxidative stress, which in turn upregulates the mRNA and protein expression of transient receptor potential melastatin 7 (TRPM7) and activates the mitogen-Activated protein kinase(MEK)/extracellular signal-regulated kinase(ERK)signaling pathway. Collectively, these events promote the phenotypic switching of aortic VSMCs from the contractile to the synthetic phenotype and enhance cellular proliferation in rats. Notably, silencing of TRPM7, inhibition of MEK1/2, or scavenging of ROS can significantly abrogate the aforementioned effects, thus confirming that the ROS-TRPM7-ERK1/2 axis serves as a critical mechanism underlying hyperglycemia-induced abnormal proliferation and phenotypic switching of VSMCs.78 Notably, autophagy, a catabolic process responsible for the clearance and recycling of damaged cells and organelles, is also involved in the phenotypic switching of VSMCs. Accumulating evidence suggests that autophagy is crucial for maintaining the contractile phenotype of VSMCs, whereas excessive autophagy promotes the phenotypic transition of VSMCs toward a synthetic phenotype. Furthermore, studies have demonstrated that hyperglycemia upregulates Notch receptor 3 (Notch3) expression, which in turn inhibits PI3K/Akt/mechanistic target of rapamycin (mTOR) signaling pathway, thereby exacerbating the excessive activation of autophagy.79
Ultimately, VSMCs that have undergone phenotypic switching exhibit robust proliferative, migratory, and secretory capacities. Studies have revealed that hyperglycemia treatment leads to an elevated rate of DNA synthesis and an increased migratory rate of VSMCs. Moreover, this treatment results in enhanced activities of matrix metalloproteinase-2 and −9, which further potentiates the migratory capacity of these cells.80,81 Furthermore, dedifferentiated VSMCs secrete substantial amounts of OPN, type I collagen, and fibronectin, which contribute to the formation of the fibrous cap. Concomitantly, these cells release pro-inflammatory cytokines including interleukin-6 (IL-6), monocyte chemoattractant protein-1 (MCP-1), and granulocyte-macrophage colony-stimulating factor, thereby recruiting monocytes/macrophages and amplifying local vascular inflammation.82 Single-cell sequencing studies have further revealed that VSMCs under hyperglycemic conditions can undergo further multidirectional phenotypic differentiation into diverse subpopulations, including osteogenic-like, chondrogenic-like, macrophage-like, and even stem cell-like subsets, thereby providing a cellular basis for plaque calcification and neovascularization.83,84
Mechanisms of Chinese Herbal Medicines in Intervening with Diabetes-Associated Atherosclerosis
The pathogenic mechanisms underlying hyperglycemia-driven atherosclerosis are highly intricate, with macrophages, endothelial cells, and VSMCs playing pivotal roles. Recent investigations have revealed that traditional chinese herbal medicines exhibit a “multi-component–multi-target–multi-pathway” pattern of synergistic regulation. Contemporary pharmacological investigations have further confirmed that traditional chinese herbal medicines and their bioactive constituents possess anti-apoptotic, antioxidant, anti-inflammatory, and hypoglycemic properties.85,86 These attributes endow traditional chinese herbal medicines with unique advantages in the prevention and treatment of hyperglycemia-associated atherosclerosis. Moreover, traditional Chinese herbal remedies have been formally incorporated into China’s clinical management pathway for hyperglycemia-associated atherosclerosis, and their efficacy has been substantiated by a growing body of evidence-based investigations.87,88 Building on these foundations, the precise delineation of molecular targets and the mechanistic underpinnings of bioactive constituents derived from traditional chinese herbal medicines is expected to furnish novel strategies for the precision management of hyperglycemia-driven atherosclerosis. Consequently, a systematic dissection of the molecular networks through which these phytochemicals modulate hyperglycemia-associated atherosclerosis will provide innovative translational avenues for subsequent drug discovery and clinical application. The table summarizes the protective effects of chinese herbal medicines decoctions and active components against hyperglycemia-associated atherosclerosis (Table 1).
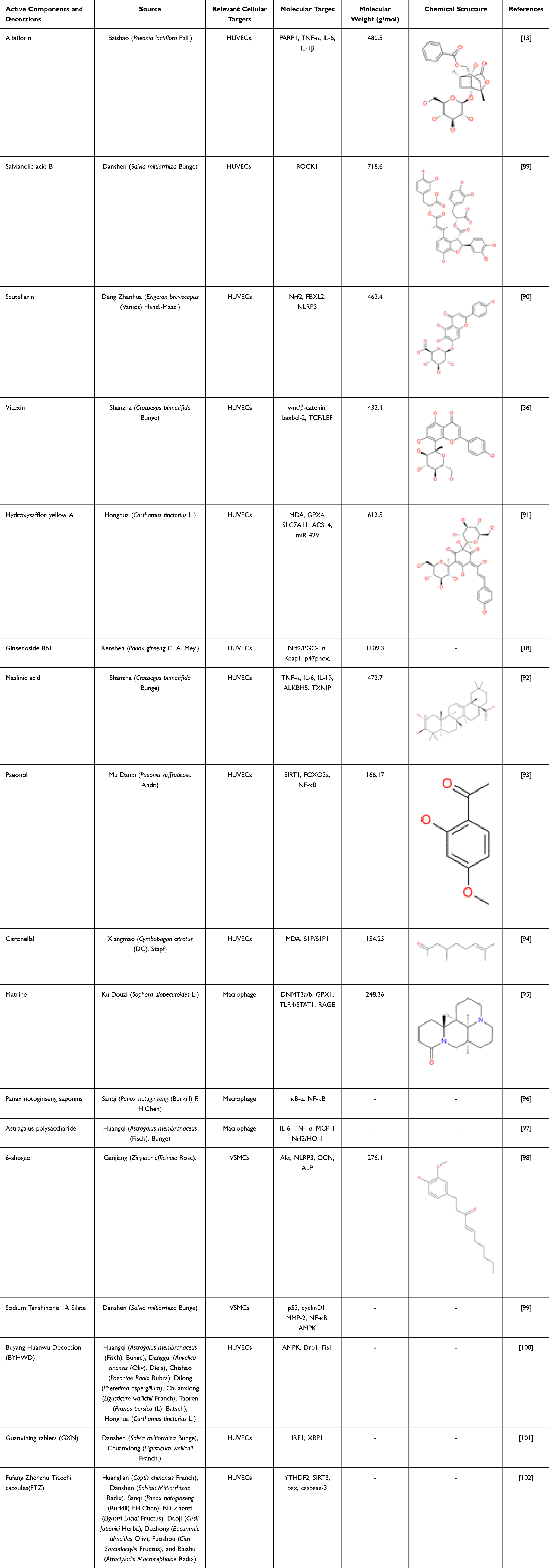 |
Table 1 Active Components and Decoctions Protective Effects on Diabetes-Associated Atherosclerosis |
Protection Mechanisms of Chinese Herbal Medicines Intervention on Endothelial Cells in Hyperglycemia Environments
Albiflorin, a monoterpene glycoside, is the principal bioactive constituent isolated from the traditional Chinese herb Baishao (Paeonia lactiflora Pall). Studies have found that Albiflorin can enhance the sensitivity of peripheral tissues to insulin and improve insulin-stimulated glucose clearance. Additionally, it can promote the translocation of glucose transporter 4 (GLUT4) to the cell membrane, accelerate glucose uptake by cells, and reduce blood glucose levels. Furthermore, Albiflorin can regulate systemic energy metabolism, thereby maintaining blood glucose homeostasis.14,103 Recent research has revealed that Albiflorin also exhibits a protective effect on blood vessels. In an in vitro model, Yang et al demonstrated that albiflorin can reverse the loss of viability in HUVECs induced by hyperglycemia. They also found that albiflorin can promote the proliferation and migration of HUVECs, as well as suppress the release of pro-inflammatory cytokines, such as tumour necrosis factor-alpha (TNF-α), IL-6 and IL-1β. Finally, they observed that albiflorin can attenuate endothelial apoptosis. Molecular-docking simulations in combination with plasmid transfection assays, performed by the same research group, identified poly (ADP-ribose) polymerase-1 (PARP1) as the critical molecular target through which albiflorin exerts its protective actions.13
Salvianolic acid B, a polyphenolic compound extracted from the traditional Chinese medicinal herb Danshen (Salvia miltiorrhiza Bunge), has attracted extensive scholarly attention because of its comprehensive cardiovascular protective effects.104 Studies have shown that salvianolic acid B lowers blood glucose levels. Salvianolic acid B upregulates the expression of GLUT4 and glycogen synthase in skeletal muscle, promoting the uptake and utilization of glucose by peripheral tissues.105 At the same time, salvianolic acid B can also inhibit hepatic gluconeogenic gene expression, reduce fasting blood glucose levels, and improve abnormal glucose tolerance. In addition, salvianolic acid B alleviates insulin resistance by inhibiting endoplasmic reticulum stress in the liver.106,107 Recent studies have found that salvianolic acid B can reduce vascular inflammation and inhibit the progression of atherosclerosis. Ko89 et al used a hyperglycemia combined with oxLDL-induced human endothelial cell injury model to confirm that salvianolic acid B dose-dependently inhibits mitochondrial reactive oxygen species generation, thereby reducing oxidative stress. Specifically blocking Rho-associated coiled-coil-containing protein kinase 1 (ROCK1) pathway inhibits endothelial cell apoptosis and mitochondrial fission abnormalities, ultimately improving endothelial cell injury, alleviating endothelial dysfunction, and ameliorating hyperglycemia-induced endothelial cell injury.
Scutellarin, a flavonoid isolated from the traditional Chinese medicinal herb Deng Zhanhua (Erigeron breviscapus (Vaniot) Hand.-Mazz), exhibits potent anti-inflammatory, antioxidant, and vasorelaxant properties.108,109 Currently, it is primarily employed in the clinical management of stroke and myocardial infarction.108,110 Studies have found that Scutellarin can activate the AMPK pathway, thereby improving insulin resistance; in addition, Scutellarin can accelerate glucose uptake and utilization by cells, thereby reducing blood glucose levels.111,112 Recent research has shown that Scutellarin can alleviate hyperglycemia-induced endothelial cell damage. Meng90 et al utilized HUVECs to establish an in vitro model of diabetes-associated atherosclerosis induced by hyperglycemia. They found that scutellarin up-regulates Nrf2. Acting as a transcription factor, Nrf2 binds to the F-box and leucine-rich repeat protein 2 (FBXL2) promoter and enhances its transcription. FBXL2, in turn, targets the inflammasome core component NLRP3 for ubiquitin-dependent degradation. This process has been demonstrated to suppress endothelial pyroptosis and inflammatory responses, thereby ultimately ameliorating hyperglycemia-induced endothelial injury. Moreover, the team employed shRNA-mediated knockdown and demonstrated that silencing Nrf2 down-regulated FBXL2 and abolished the protective effect of scutellarin. FBXL2 knockdown attenuated NLRP3 degradation and exacerbated cellular injury, whereas NLRP3 knockdown alleviated hyperglycemia-induced damage. These findings indicate that scutellarin confers protection against diabetic AS via the Nrf2-FBXL2–NLRP3 signaling axis.
Vitexin is a flavonoid compound extracted from traditional chinese medicinal herbs such as Shanzha (Crataegus pinnatifida Bunge).113 Studies have shown that Vitexin can regulate insulin signaling pathways and improve insulin resistance. Additionally, it can inhibit α-glucosidase activity, delay the digestion and absorption of carbohydrates in the intestine, and thereby reduce postprandial blood glucose spikes.114,115 In vitro experiments conducted by Zhang et al revealed that vitexin facilitates the nuclear translocation of β-catenin and its subsequent interaction with T-cell factor (TCF)/lymphoid enhancer-binding factor (LEF) transcription factors, thereby suppressing the pro-apoptotic protein Bax while enhancing the expression of the anti-apoptotic protein Bcl-2, ultimately attenuating apoptosis in HUVECs.36
Hydroxysafflor yellow A is a monomeric chalcone glycoside principally isolated from the traditional Chinese medicinal herb Honghua (Carthamus tinctorius L).116 Studies have found that Hydroxysafflor yellow A can reduce pancreatic β-cell apoptosis and maintain insulin secretion function; additionally, Hydroxysafflor yellow A can inhibit α-glucosidase activity, delay intestinal carbohydrate absorption, and lower postprandial blood glucose levels.117,118 It has been reported that hydroxysafflor yellow A exerts vascular-protective effects.119 Additionally, Xie et al demonstrated that hydroxysafflor yellow A effectively ameliorates H2O2-induced oxidative injury in human umbilical vein endothelial cells.120 Rong91 et al constructed a diabetes AS model using ApoE−/− mice and found that hydroxysafflor yellow A can improve glucose and lipid metabolism disorders, reduce the production of Malondialdehyde (MDA) and ROS, upregulate the expression of anti-ferroptosis-related proteins Glutathione Peroxidase 4 (GPX4) and Solute Carrier Family 7 Member 11 (SLC7A11), inhibit the pro-ferroptosis protein ACSL4, and downregulate the expression of miR-429. Moreover, the team also constructed a high-glucose and high-fat injury model using HUVEC, and further explored the mechanism through cell transfection technology. It was found that after transfection with miR-429 mimics (overexpressing miR-429) or SLC7A11 siRNA, the antioxidant stress and anti-ferroptosis effects of hydroxysafflor yellow A were weakened, and the protective effect on endothelial cells was significantly reduced. This indicates that hydroxysafflor yellow A exerts a protective effect on diabetes-associated atherosclerosis mice through the miR-429/SLC7A11 axis.
Ginsenoside Rb1 is a pivotal constituent of the ginsenoside family. Reports indicate that ginsenoside Rb1 can modulate central feeding mechanisms, suppress appetite, and reduce carbohydrate intake. Additionally, ginsenoside Rb1 can activate the AMPK pathway to promote glucose uptake, improve insulin resistance, and enhance insulin sensitivity, thereby achieving blood glucose regulation.121 Recent studies have shown that ginsenoside Rb1 also possesses cardiovascular and cerebrovascular protective effects.122,123 Wang18 et al established a murine model of diabetes-associated atherosclerosis in ApoE−/− mice and demonstrated that ginsenoside Rb1 markedly attenuated oxidative stress and inflammatory responses while reducing aortic atherosclerotic plaque burden. Notably, tail-vein delivery of AAV9-Tie2 vectors to silence endothelial Nrf2 or Peroxisome proliferator-activated receptor gamma coactivator 1-α(PGC-1α) completely abolished these protective actions, unequivocally establishing that the anti-atherogenic efficacy of Ginsenoside Rb1 in vivo is contingent upon activation of Nrf2/PGC-1α axis. To pinpoint the direct molecular targets of ginsenoside Rb1, Wang et al integrated molecular docking, surface plasmon resonance, and microscale thermophoresis analyses, revealing that ginsenoside Rb1 physically engages both Kelch-like ECH-associated protein 1(Keap1) and p47phox. ginsenoside Rb1–Keap1 interaction facilitated Keap1 ubiquitination and proteasomal degradation, liberating Nrf2 for nuclear translocation and subsequent heterodimerization with PGC-1α. Concurrently, ginsenoside Rb1-bound p47phox exhibited diminished phosphorylation and membrane translocation, preventing NOX2 holoenzyme assembly. This dual-target engagement coordinately ignites the Nrf2/PGC-1α transcriptional program. Collectively, Keap1 and p47phox are validated as the bona fide direct targets of Ginsenoside Rb1, and the Nrf2/PGC-1α signaling axis constitutes the central pathway underpinning its anti-diabetic atheroprotective activity.
Maslinic acid is a pentacyclic triterpenoid principally enriched in the medicinal herb Shan zha (Crataegus pinnatifida Bunge) and related botanicals.124 It has been reported that maslinic acid can inhibit glycogen phosphorylase activity, reduce hepatic glycogenolysis, and decrease hepatic glucose output; it can also inhibit α-glucosidase activity, thereby delaying carbohydrate digestion and absorption in the intestine and lowering postprandial blood glucose.125,126 Furthermore, studies have demonstrated that maslinic acid attenuates hyperglycemia-induced endothelial cell injury. Wang92 et al demonstrated that maslinic acid lowers the levels of TNF-α, IL-6, and IL-1β, attenuates ROS generation, and diminishes apoptosis in HUVECs. Mechanistically, maslinic acid facilitates the recruitment of the RNA demethylase alpha-ketoglutarate-dependent dioxygenase alkB homolog 5 (ALKBH5) to thioredoxin-interacting protein (TXNIP) mRNA, augmenting m6A demethylation of TXNIP transcripts and thereby reducing their stability. The consequent down-regulation of TXNIP—a glucose-sensing adaptor protein—suppresses hyperglycemia-induced ROS production and pro-inflammatory cytokine release, ultimately mitigating endothelial inflammation and apoptosis.
Paeonol is a phenolic acid compound derived from chinese herbal medicines such as Mu Danpi (Paeonia suffruticosa Andr).127 Previous studies have found that Paeonol possesses anti-inflammatory and antioxidant properties, capable of mitigating oxidative and inflammatory damage to endothelial cells.128,129 Research has demonstrated that Paeonol can enhance insulin sensitivity, promote glycogen synthesis, and inhibit gluconeogenesis, thereby achieving blood glucose-lowering effects.130,131 Furthermore, Paeonol exhibits anti-inflammatory and antioxidant properties, capable of alleviating oxidative and inflammatory damage in endothelial cells. Tang93 et al further demonstrated that paeonol mitigates hyperglycemia plus palmitic acid-induced HUVEC injury by up-regulating Sirtuin 1 (SIRT1) expression. Activated SIRT1 deacetylates Forkhead box protein O3 a (FOXO3a), thereby repressing its pro-apoptotic and pro-oxidative activities, while concurrently suppressing NF-κB signaling to curtail transcription and release of inflammatory mediators. These complementary actions collectively ameliorate endothelial apoptosis, oxidative stress, and inflammatory damage.
Citronellal, a monoterpenoid aldehyde, is principally isolated from the essential oils of Xiangmao (Cymbopogon citratus (DC). Stapf).132 It has been reported that citronellal can inhibit α-glucosidase and α-amylase activities in the intestine, delay the digestion and decomposition of carbohydrates and the absorption rate of glucose, and reduce postprandial blood glucose peaks. Additionally, it can regulate insulin signaling pathways and enhance insulin sensitivity.133 Previous investigations have demonstrated that citronellal attenuates atherosclerotic plaque formation and arterial stenosis by suppressing oxidative stress and promoting endothelial cell migration.134 Qiu94 et al employed a diabetic rat model induced by a high-fat diet plus streptozotocin(STZ) and cultured HUVECs under hyperglycemia to recapitulate the vascular injury elicited by hyperglycemia. It was observed that citronellal significantly reduced vascular lipid deposition, restored endothelial alignment, and enhanced endothelium-dependent vasodilation. Concurrently, citronellal lowered malondialdehyde and ROS production, thereby conferring protection against diabetes-associated endothelial damage. The beneficial effects of citronellal appear to be mediated by the activation of the sphingosine-1-phosphate (S1P)/Sphingosine 1-phosphate (S1P1) axis, which is induced by citronellal. This activation promotes the phosphorylation-dependent dimerisation of endothelial nitric-oxide synthase and augments nitric-oxide generation.
Buyang Huanwu Decoction (BYHWD), a classical formulation in traditional Chinese medicine, is renowned for its therapeutic efficacy against cardio-cerebrovascular disorders, including ischemic stroke and atherosclerosis.135,136 It is composed of 7 chinese medicinal herbs: Huangqi (Astragalus membranaceus (Fisch). Bunge), Danggui (Angelica sinensis (Oliv). Diels), Chishao (Paeoniae Radix Rubra), Dilong (Pheretima aspergillum), Chuanxiong (Ligusticum wallichii Franch), Taoren(Prunus persica (L). Batsch), and Honghua (Carthamus tinctorius L).137 Research has found that BYHWD protects pancreatic β-cell function and improves insulin secretion capacity; moreover, BYHWD can also promote glucose uptake in skeletal muscle and adipose tissue, ultimately achieving blood glucose regulation.138 Recent studies have shown that BYHWD also has protective effects against hyperglycemia-induced vascular cell injury. Tong100 et al utilised STZ-induced ApoE−/− mice to establish a diabetes-associated atherosclerosis model and ascertained that BYHWD activates AMPK pathway, down-regulates the expression of Dynamin-related protein 1 (Drp1) and Mitochondrial fission 1 (Fis1) proteins, suppresses aberrant mitochondrial fission, and reduces ROS generation, thereby improving endothelial function and inhibiting hyperglycemia-accelerated atherosclerosis. In order to provide further validation of the key molecular component of this pathway, the team silenced the AMPK gene in HUVECs via siRNA transfection. The results demonstrated that following AMPK knockdown, the protective effect of BYHWD against hyperglycemia-induced endothelial injury was significantly diminished, thereby confirming that AMPK is a critical mediator of BYHWD’s protective action. Furthermore, molecular docking with AutoDock Vina revealed that BYHWD’s core active constituents – ferulic acid and calycosin-7-glucoside – directly bind to the GTPase domain of Drp1. Subsequent GTPase activity assays further verified that both compounds effectively inhibit Drp1 GTPase activity, cooperatively blocking the mitochondrial fission signal. These findings not only clarify the therapeutic targets and active ingredients of BYHWD, but also exemplify the multi-component, multi-target synergistic advantage of traditional Chinese medicine.
Guanxining tablets (GXN) are composed of two well-known chinese herbal medicines—Danshen (Salvia miltiorrhiza Bunge) and Chuanxiong (Ligusticum wallichii Franch).—that are renowned for promoting blood circulation.139 Studies have shown that GXN can decrease levels of Endothelin-1, von Willebrand factor, SOD, and TNF-α, thereby alleviating vascular inflammation and oxidative stress and protecting vascular endothelium.140 Furthermore, Li141 et al reported that GXN effectively modulates glucose and lipid metabolism in Zucker diabetic fatty rats, lowering fasting and postprandial blood glucose, glycated hemoglobin, and total cholesterol levels. Wang101 et al further demonstrated in vitro that GXN modulates the interplay between endoplasmic-reticulum stress and autophagy: it up-regulates Endoplasmic reticulum stress proteins to potentiate the unfolded-protein response, while simultaneously promoting protective autophagy and suppressing endothelial-cell apoptosis, thereby attenuating hyperglycemia-induced injury to vascular endothelium.
Fufang Zhenzhu Tiaozhi capsules(FTZ) is composed of eight chinese medicinal herbs: Huanglian (Coptis chinensis Franch), Danshen (Salviae Miltiorrhizae Radix), Sanqi (Panax notoginseng (Burkill) F.H.Chen), Nü Zhenzi (Ligustri Lucidi Fructus), Daji (Cirsii Japonici Herba), Duzhong (Eucommia ulmoides Oliv), Fuoshou (Citri Sarcodactylis Fructus), and Baizhu (Atractylodis Macrocephalae Radix).142 Studies have shown that FTZ can improve glucose and lipid metabolism, lower fasting blood glucose, ameliorate oxidative stress, and protect vascular endothelium.143 Zhang102 et al first conducted a clinical trial, finding that FTZ lowers fasting plasma glucose, total cholesterol, and triglycerides, and reduces carotid intima-media thickness and plaque area in diabetic patients with atherosclerosis. They next generated a diabetes-associated atherosclerosis model by inducing streptozotocin in ApoE−/− mice and showed that FTZ ameliorates glucose- and lipid-metabolic disorders, decreasing aortic lipid deposition and plaque area. To clarify the molecular mechanism, the same team established an in vitro model by stimulating HUVECs with hyperglycemia plus ox-LDL and demonstrated that FTZ suppresses YTH N6-methyladenosine RNA binding protein 2 (YTHDF2)-mediated m6A modification of SIRT3 mRNA, diminishes sirtuin 3 (SIRT3) mRNA degradation, up-regulates SIRT3 expression, lowers the levels of the pro-apoptotic proteins Bax and cleaved caspase-3, inhibits endothelial cell apoptosis, and alleviates atherosclerosis.
Protection Mechanisms of Chinese Herbal Medicines Intervention on Macrophages in Hyperglycemia Environments
Matrine is an alkaloid extracted from the Chinese herb Ku Douzi (Sophora alopecuroides L).144 It has been shown to promote the secretion of glucagon-like peptide-1 from intestinal L cells, thereby improving insulin resistance and glucose metabolism. Additionally, it enhances glucose uptake in skeletal muscle and adipose tissue, ameliorating disorders of glucose metabolism.145 Evidence indicates that matrine exerts cardiovascular protective effects, primarily attributed to its antioxidant, anti-apoptotic, and anti-atherosclerotic properties.146,147 Cui95 et al stimulated primary mouse macrophages with AGEs to establish a model that mimics vascular injury caused by AGEs accumulation under diabetic conditions. They found that matrine down-regulates the expression of Deoxyribonucleic Acid methyltransferases 3a/b (DNMT3a/b), thereby reducing promoter methylation of glutathione peroxidase 1 (GPX1) and restoring GPX1 expression. As a key antioxidant molecule, GPX1 suppresses oxidative stress mediated by AGEs binding to their receptor, consequently inhibiting aberrant activation of TLR4/signal transducer and activator of transcription 1 (STAT1) signaling pathway and ultimately blocking macrophage polarization toward the pro-inflammatory M1 phenotype. To further elucidate the molecular mechanism, the team employed plasmid-mediated gene overexpression. The results showed that RAGE overexpression markedly attenuated matrine’s inhibitory effects on oxidative stress, TLR4/STAT1 signaling activation, and M1 polarization, whereas TLR4 overexpression only weakened matrine’s suppression of TLR4/STAT1 signaling and M1 polarization without appreciably affecting oxidative stress. These findings establish RAGE as the critical upstream target for matrine’s antioxidant action and TLR4 as the core molecule governing the downstream signaling cascade, ultimately confirming that matrine’s protection against diabetes-associated vascular injury depends on precise modulation of the RAGE/TLR4/STAT1 pathway.
Panax notoginseng saponins are the bioactive constituents extracted from Santi(Panax notoginseng (Burkill) F.H.Chen). According to reports, Panax notoginseng saponins can regulate insulin secretion, improve pancreatic β-cell function, and consequently ameliorate insulin resistance.148,149 Additionally, Panax notoginseng saponins possess anti-inflammatory and anti-atherosclerotic properties.150 Zhao96 et al found that Panax notoginseng saponins dose-dependently regulate macrophage polarization toward the anti-inflammatory M2 phenotype while suppressing the pro-inflammatory M1 phenotype. The underlying mechanism is primarily associated with the inhibition of IκB-α phosphorylation and the activation of NF-κB p50 and p65 subunits by Panax notoginseng saponins, thereby blocking NF-κB signaling and improving diabetes-associated atherosclerosis.
Astragalus polysaccharide is the principal bioactive component extracted from the root of the traditional Chinese herb Huangqi (Astragalus membranaceus (Fisch). Bunge).151 Modern pharmacological studies have found that astragalus polysaccharide can exert hypoglycemic effects by regulating intestinal flora structure and improving intestinal mucosal barrier function. In addition, astragalus polysaccharide can also improve insulin resistance, increase insulin sensitivity, and ameliorate glucose metabolism disorders.152,153 Recent studies have also shown that astragalus polysaccharide possesses cardiovascular-protective and anti-inflammatory properties. Sha97 et al reported that Astragalus polysaccharide reduced the thickness of the thickened thoracic aortic wall in diabetic rats, whilst concomitantly reducing the expression of inflammatory mediators, including IL-6, TNF-α, and MCP-1, in thoracic aortic tissue. Furthermore, astragalus polysaccharide has been demonstrated to skew macrophage polarization towards the anti-inflammatory M2 phenotype whilst suppressing the pro-inflammatory M1 phenotype. In order to elucidate the underlying molecular mechanism, the group performed in vitro experiments in which macrophages were stimulated with lipopolysaccharide plus hyperglycemia. It was found that astragalus polysaccharide up-regulated Nrf2 and heme oxygenase-1 (HO-1) expression within the Nrf2/HO-1 signalling axis, thereby exerting the aforementioned protective effects. When Nrf2 was silenced, the protective actions of astragalus polysaccharide were largely abolished, indicating that Nrf2 is a critical target through which astragalus polysaccharide modulates macrophage polarization and ameliorates diabetes-associated vascular inflammation and injury.
Protection Mechanisms of Chinese Herbal Medicines Intervention on Vascular Smooth Muscle Cells in Hyperglycemia Environments
6-Shogaol, the principal bioactive constituent of the herbal medicine Ganjiang (Zingiber officinale Rosc), exerts anti-inflammatory and antioxidant effects.154,155 In addition, Studies have also found that 6-shogaol can improve insulin sensitivity in peripheral tissues, protect pancreatic β-cell morphology, reduce apoptosis, maintain islet function, and lower blood glucose levels.156,157 Recent research has demonstrated that 6-shogaol can delay the progression of diabetes-associated atherosclerosis. Chen98 et al demonstrated that 6-shogaol mitigates hyperglycemia-induced injury in Human artery smooth muscle cells. Mechanistically, 6-shogaol blocks Akt phosphorylation, attenuates ROS generation, and consequently suppresses NLRP3 inflammasome activation. This cascade down-regulates osteogenic proteins—including osteopontin (OPN), osteocalcin (OCN), and alkaline phosphatase (ALP)—markedly inhibiting hyperglycemia-triggered vascular calcification and thereby protecting arterial smooth muscle cells.
Sodium Tanshinone IIA Silate is a water-soluble derivative of Tanshinone IIA. Previous studies have shown that Tanshinone IIA lowers blood glucose and improves glucose tolerance; it also suppresses vascular smooth muscle cell proliferation.158 Wu99 et al reported that, under hyperglycemia conditions, Sodium Tanshinone IIA Silate arrests VSMCs in G0/G1, markedly shrinking S and G2/M fractions. This is achieved by up-regulating p53 and the CDK inhibitor p21 while down-regulating cyclin D1 expression and promoter activity in an AMPK-dependent manner. Transwell assays further showed that Sodium Tanshinone IIA Silate dose-dependently blunts hyperglycemia-stimulated VSMCs migration, suppresses MMP-2, and blocks both NF-κB nuclear translocation and cytosolic NF-κB activation. Crucially, siRNA knockdown of AMPK abolished Sodium Tanshinone IIA Silate-mediated inhibition of proliferation and migration, confirming that the vascular protection conferred by Sodium Tanshinone IIA Silate requires AMPK activation.
Conclusions
With the continued rise in global diabetes prevalence, the incidence of diabetes-related vascular complications is also showing a significant upward trend.159 Current clinical management primarily relies on glucose-lowering medications, dietary interventions, and exercise regimens to control blood glucose levels.160,161 However, these conventional interventions have limited efficacy in delaying or reversing the progression of diabetic vasculopathy and fail to effectively curb the onset and advancement of vascular complications such as atherosclerosis.162 Therefore, there is an urgent need to develop novel therapeutic strategies that can directly target the mechanisms of diabetic vascular injury. Hence, there is an urgent need to develop innovative therapeutic strategies capable of directly intervening in the pathogenic mechanisms underlying diabetic vascular injury.
Atherosclerosis, the most prevalent pathological basis for a variety of cardiovascular and cerebrovascular diseases, involves the functional disruption of multiple cell types. Vascular endothelial cells, vascular smooth-muscle cells, and macrophages play pivotal roles throughout the progression of atherosclerosis.163 Recent studies have shown that hyperglycemia can act on these three types of cells through various molecular mechanisms, inducing their functional abnormalities and thereby promoting the initiation and progression of atherosclerosis.164 Therefore, elucidating the specific mechanisms by which hyperglycemia affects vascular endothelial cells, smooth muscle cells, and macrophages not only helps to uncover the pathogenesis of diabetic vascular complications but also provides a theoretical basis for developing novel intervention strategies that target the functions of vascular cells. Therefore, elucidating the specific molecular mechanisms by which hyperglycemia disrupts the functions of vascular endothelial cells, smooth muscle cells, and macrophages not only deepens our understanding of the pathogenesis of diabetic vascular complications, but also provides a theoretical foundation for developing novel interventions that target vascular-cell function. This review systematically summarizes the molecular pathways through which hyperglycemia induces vascular-cell dysfunction, examining—through the three perspectives of endothelial cells, vascular smooth muscle cells, and macrophages—the mechanisms by which hyperglycemia acts on vascular cells, with the aim of offering a comprehensive theoretical framework for understanding how hyperglycemia promotes atherosclerosis.
In China, chinese herbal medicines have demonstrated promising clinical prospects as an important therapeutic approach for diabetic atherosclerosis. Studies have shown that chinese herbal medicines can slow the progression of diabetes-associated atherosclerosis.165 Clinical retrospective analyses have also demonstrated that, when combined with conventional Western therapies, chinese herbal medicines can further reduce carotid intima–media thickness and coronary artery calcification scores, with a favorable safety profile.166 Moreover, relevant randomized controlled trials are currently underway and are expected to furnish robust evidence supporting the use of traditional Chinese herbal medicine in the treatment of diabetes-associated atherosclerosis.87,88 In recent years, research on the metabolically active components of chinese herbal medicines has been gradually deepening. Therefore, this article systematically reviews the mechanisms by which chinese herbal medicines and their active metabolic components improve diabetic atherosclerosis by regulating the functions of vascular endothelial cells, vascular smooth muscle cells, and macrophages. Furthermore, it summarizes the sources of acquisition for the relevant active components of chinese herbal medicines (Table 2) and the Latin names and main efficacy of the major chinese herbal medicines (Table 3) to demonstrate the reliability and rationality of the metabolically active components of chinese herbal medicines. The aim is to provide new insights for diabetes-associated atherosclerosis.
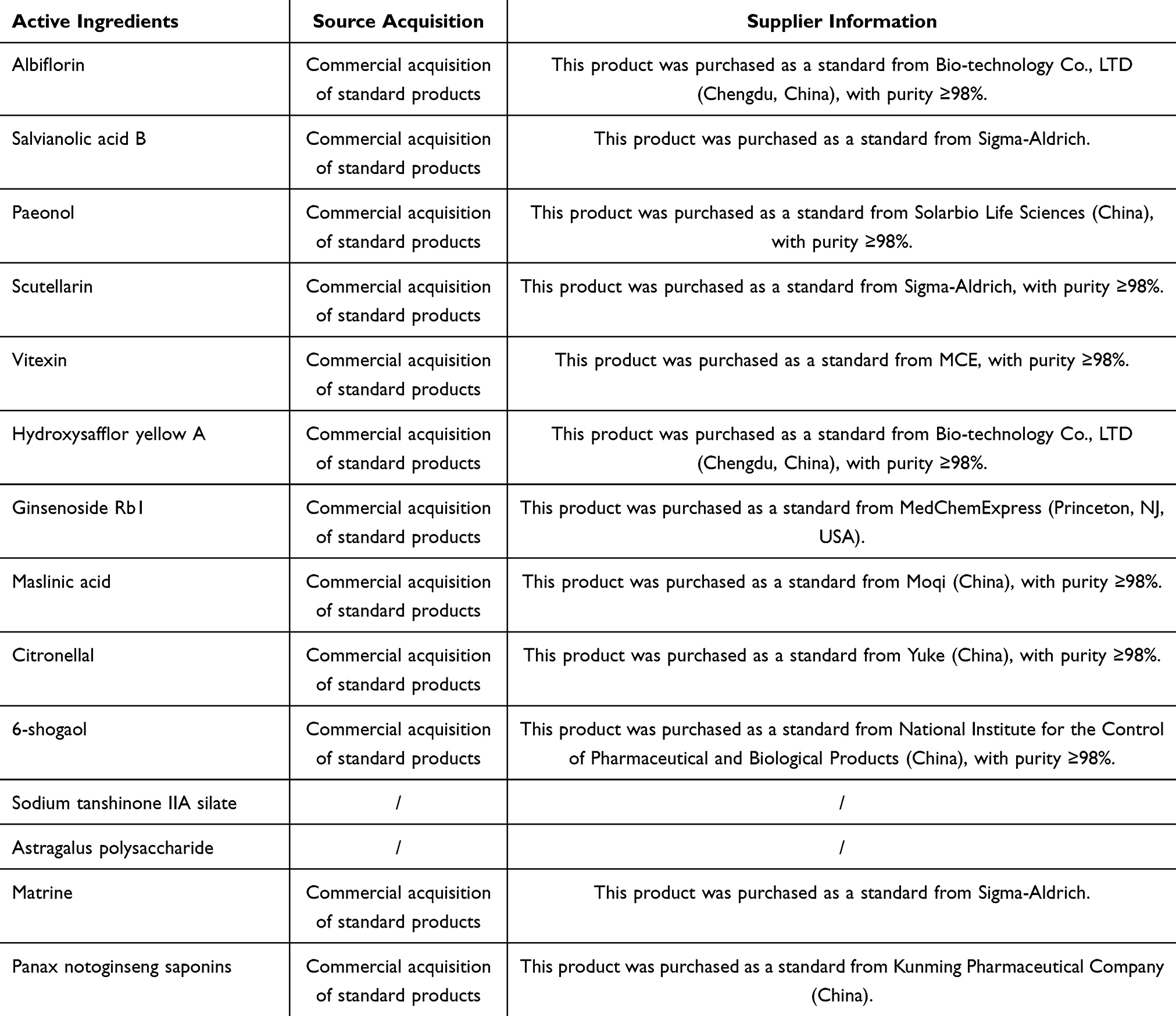 |
Table 2 Sources of Active Components from Chinese Herbal Medicines |
 |
Table 3 Sources, supplies adn purity of Active Components from Chinese Herbal Medicines |
Despite the growing body of research on Chinese herbal medicines for diabetes-associated atherosclerosis, several critical challenges continue to impede their translation into clinical practice. Firstly, limitations inherent to current animal and cell models. The most widely used diabetes-associated atherosclerosis model—the ApoE−/− mouse—exhibits a lipoprotein profile dominated by VLDL and abnormally elevated HDL, markedly different from the LDL-driven pathology seen in humans. In vitro studies, meanwhile, typically rely on static single-cell cultures that lack crucial micro-environmental cues present in vivo—such as hemodynamic shear stress and inter-cellular crosstalk—resulting in systems that poorly recapitulate physiological or pathological conditions. These disparities can skew assessments of both the efficacy and the molecular targets of herbal constituents, and may overlook bioactive mechanisms that emerge only through multi-cellular cooperation, ultimately compromising the accuracy of downstream translational investigations. Secondly, inadequate safety evaluation. The chemical complexity of herbal medicines means that current studies rarely provide systematic data on hepatotoxicity, nephrotoxicity, chronic toxicity, or reproductive toxicity; moreover, information on herb–drug interactions with commonly prescribed hypoglycaemic or anti-atherosclerotic agents is scarce. The absence of a standardized safety-assessment framework makes it impossible to quantify the clinical risks associated with either whole herbs or their bioactive metabolites. Finally, insufficient clinical evidence. Most existing work centres on single purified constituents tested in cellular or animal models, whereas well-designed clinical trials lag far behind and are methodologically weak. Studies rarely stratify participants by diabetes subtype or stage of atherosclerotic lesion, further limiting the external validity and therapeutic guidance that can be derived from the data.
Future research should therefore focus on three priorities. First, model refinement: adopt systems that more faithfully replicate human LDL-driven atherosclerotic metabolism to enhance both scientific rigor and translational relevance. Second, establish a comprehensive pharmacological safety platform specifically tailored to herbal medicines, systematically evaluating hepatorenal toxicity, chronic toxicology, and herb–drug interactions. Third, launch large-scale, multicentre, long-term clinical trials that generate high-level evidence. Such efforts will not only foster wider acceptance of herbal therapies among patients with diabetes-associated atherosclerosis, but also provide safer, more effective therapeutic options grounded in robust evidence-based medicine.
Abbreviations
ECs, Endothelial cells; eNOS, endothelial nitric oxide synthase; NO, nitric oxide; LDL, low-density lipoprotein; AKT, protein kinase B; PI3K, phosphatidylinositol 3-kinase; ROS, reactive oxygen species; AGEs, advanced glycation end products; NF-κB, Nuclear factor-κB; ICAM-1, intercellular adhesion molecule-1; VCAM-1, vascular cell adhesion molecule-1; ox-LDL, oxidized low-density lipoprotein; HUVECs, human umbilical vein endothelial cells; Bcl-2, B cell lymphoma-2; Bax, Bcl-2-associated X protein; NLRP3, NOD-like receptor family pyrin domain-containing 3; AMPK, AMP-activated kinase; KLF4, Krüppel-like factor 4; TERT, telomerase reverse transcriptase; SIRT6, Sirtuin type-6; Nrf2, nuclear factor erythroid-2 related factor 2; HIF-2α, hypoxia-inducible factor-2α; GLUT1, glucose transporter 1; COMMD1, copper metabolism domain-containing protein 1; 2-DG, 2-deoxy-D-glucose; RUNX1, Runt-related transcription factor 1; HIF-1α, hypoxia-inducible factor-1α; PDK4, Pyruvate dehydrogenase kinase 4; SR-B, scavenger receptors class B; LOX-1, low-density lipoprotein receptor 1; RNA, Ribonucleic Acid; MALAT1, metastasis-associated lung adenocarcinoma transcript 1; ELAVL1, ELAV like RNA binding protein 1; IL-1β, Interleukin-1β, VSMCs, Vascular smooth muscle cells; JAK, Janus kinase; STAT, signal transducer and activator of transcription; JAK2, Janus kinase 2; AngII, angiotensin II; AT1, angiotensin type 1 receptor; SHP-1, Src homology region 2 domain-containing phosphatase-1; KLF5, Krüppel-like factor 5; TLR4, Toll-like receptor 4; TRPM7, transient receptor potential melastatin 7; MEK, mitogen-Activated protein kinase; ERK, extracellular signal-regulated kinase; Notch3, Notch receptor 3; mTOR, mechanistic target of rapamycin; MCP-1, monocyte chemoattractant protein-1; TNF-α, tumour necrosis factor-alpha; PARP1, poly(ADP-ribose) polymerase-1; ROCK1, Rho-associated coiled-coil-containing protein kinase 1; FBXL2, F-box and leucine-rich repeat protein 2; TCF, T-cell factor; LEF, lymphoid enhancer-binding factor; GPX4, Glutathione Peroxidase 4; SLC7A11, Solute Carrier Family 7 Member 11; PGC-1α, proliferator-activated receptor gamma coactivator 1-α; Keap1, Kelch-like ECH-associated protein 1; ALKBH5, alpha-ketoglutarate-dependent dioxygenase alkB homolog 5; TXNIP, thioredoxin-interacting protein; SIRT1, Sirtuin 1; FOXO3a, Forkhead box protein O3 a; STZ, streptozotocin; S1P, sphingosine-1-phosphate; S1P1, Sphingosine 1-phosphate; Drp1, Dynamin-related protein 1; Fis1, fission 1; YTHDF2, YTH N6-methyladenosine RNA binding protein 2; SIRT3, sirtuin 3; GPX1, glutathione peroxidase 1; STAT1, signal transducer and activator of transcription 1; HO-1, heme oxygenase-1; OPN, osteopontin; OCN, osteocalcin; ALP; alkaline phosphatase; AS, atherosclerosis; UDP-GlcNAc, Uridine Diphosphate N-acetylglucosamine; GSDMD, Gasdermin D.
Additional Notes
The plant names used in this article have been verified by Chinese Medicinal Materials Image Database of HONG KONG BAPTIST UNIVERSITY (https://sys01.lib.hkbu.edu.hk/cmed/mmid/) and The Plant List (https://www.theplantlist.org/). Verification date: 15 January, 2026.
Acknowledgments
The molecular structures and molar masses of the active components of traditional Chinese herbal medicine are obtained with the help of PubChem (https://pubchem.ncbi.nlm.nih.gov/). Figures 1–4 are created with the assistance of BioRender(https://www.biorender.com).
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
No funding was received for this study.
Disclosure
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
1. Hossain MJ, Al-Mamun M, Islam MR. Diabetes mellitus, the fastest growing global public health concern: early detection should be focused. Health Sci Rep. 2024;7(3):e2004. doi:10.1002/hsr2.2004
2. DC GBD. Global, regional, and national burden of diabetes from 1990 to 2021, with projections of prevalence to 2050: a systematic analysis for the global burden of disease study 2021. Lancet. 2023;402(10397):203–28. doi:10.1016/S0140-6736(23)01301-6
3. Mosenzon O, Alguwaihes A, JLA L, et al. CAPTURE: a multinational, cross-sectional study of cardiovascular disease prevalence in adults with type 2 diabetes across 13 countries. Cardiovasc Diabetol. 2021;20(1):154. doi:10.1186/s12933-021-01344-0
4. Rudijanto A. The role of vascular smooth muscle cells on the pathogenesis of atherosclerosis. Acta Med Indones. 2007;39(2):86–93.
5. Ambreen S, Fatima S, Elwakiel A, et al. Hypercoagulability impairs plaque stability in diabetes-induced atherosclerosis. Nutrients. 2022;14(10):1991. doi:10.3390/nu14101991
6. Poznyak A, AV G, Poggio P, et al. The diabetes mellitus-atherosclerosis connection: the role of lipid and glucose metabolism and chronic inflammation. Int J Mol Sci. 2020;21(5):1835. doi:10.3390/ijms21051835
7. Chen X, Xie N, Feng L, et al. Oxidative stress in diabetes mellitus and its complications: from pathophysiology to therapeutic strategies. Chinese Med J. 2025;138(1):15–27.
8. An Y, Xu B, Wan S, et al. The role of oxidative stress in diabetes mellitus-induced vascular endothelial dysfunction. Cardiovasc Diabetol. 2023;22(1):237. doi:10.1186/s12933-023-01965-7
9. Mao L, Yin R, Yang L, et al. Role of advanced glycation end products on vascular smooth muscle cells under diabetic atherosclerosis. Front Endocrinol. 2022;13:983723. doi:10.3389/fendo.2022.983723
10. Maruhashi T, Higashi Y. Pathophysiological association between diabetes mellitus and endothelial dysfunction. Antioxidants. 2021;10(8):1306. doi:10.3390/antiox10081306
11. Nijat D, Zhao Q, Abdurixit G, et al. An up-to-date review of traditional Chinese medicine in the treatment of atherosclerosis: components, mechanisms, and therapeutic potentials. Phytother Res. 2025;39(8):3709–3735. doi:10.1002/ptr.70037
12. Zhang Q, Hu S, Jin Z, et al. Mechanism of traditional Chinese medicine in elderly diabetes mellitus and a systematic review of its clinical application. Front Pharmacol. 2024;15:1339148. doi:10.3389/fphar.2024.1339148
13. Yang R, Yang Y. Albiflorin attenuates high glucose-induced endothelial apoptosis via suppressing PARP1/NF-κB signaling pathway. Inflamm Res. 2023;72(1):159–169. doi:10.1007/s00011-022-01666-z
14. Ma X, Song M, Yan Y, et al. Albiflorin alleviates cognitive dysfunction in STZ-induced rats. Aging. 2021;13(14):18287–18297. doi:10.18632/aging.203274
15. Pilard M, EL O, Gourdou-Latyszenok V, et al. Endothelial cell phenotype, a major determinant of venous thrombo-inflammation. Front Cardiovasc Med. 2022;9:864735. doi:10.3389/fcvm.2022.864735
16. Gheibi S, Jeddi S, Kashfi K, et al. Regulation of vascular tone homeostasis by NO and H(2)S: implications in hypertension. Biochem Pharmacol. 2018;149:42–59. doi:10.1016/j.bcp.2018.01.017
17. van Hinsbergh VWM. Endothelium--role in regulation of coagulation and inflammation. Semin Immunopathol. 2012;34(1):93–106. doi:10.1007/s00281-011-0285-5
18. Wang Z, Niu K, Wu Y, et al. A dual Keap1 and p47(phox) inhibitor Ginsenoside Rb1 ameliorates high glucose/ox-LDL-induced endothelial cell injury and atherosclerosis. Cell Death Dis. 2022;13(9):824. doi:10.1038/s41419-022-05274-x
19. Guo J, Huang X, Dou L, et al. Aging and aging-related diseases: from molecular mechanisms to interventions and treatments. Signal Transduct Target Ther. 2022;7(1):391. doi:10.1038/s41392-022-01251-0
20. ML F, AT T, DP M, et al. The impact of glucose exposure on bioenergetics and function in a cultured endothelial cell model and the implications for cardiovascular health in diabetes. Sci Rep. 2020;10(1):19547. doi:10.1038/s41598-020-76505-4
21. CMO V, PH V-D, Dos APMF, et al. Cellular death, reactive oxygen species (ROS) and diabetic complications. Cell Death Dis. 2018;9(2):119. doi:10.1038/s41419-017-0135-z
22. Kolluru GK, Bir SC, Kevil CG. Endothelial dysfunction and diabetes: effects on angiogenesis, vascular remodeling, and wound healing. Int J Vasc Med. 2012;2012:918267. doi:10.1155/2012/918267
23. Scarpellino G, Brunetti V, Berra-Romani R, et al. The unexpected role of the endothelial nitric oxide synthase at the neurovascular unit: beyond the regulation of cerebral blood flow. Int J Mol Sci. 2024;25(16). doi:10.3390/ijms25169071
24. Kim ME, Lee JS. Advances in the regulation of inflammatory mediators in nitric oxide synthase: implications for disease modulation and therapeutic approaches. Int J Mol Sci. 2025;26(3). doi:10.3390/ijms26031204
25. GR W, Zhu Y, PV H, et al. Mechanism of platelet inhibition by nitric oxide: in vivo phosphorylation of thromboxane receptor by cyclic GMP-dependent protein kinase. Proc Natl Acad Sci U S A. 1998;95(9):4888–4893. doi:10.1073/pnas.95.9.4888
26. Stefano GB, Challenger S, Kream RM. Hyperglycemia-associated alterations in cellular signaling and dysregulated mitochondrial bioenergetics in human metabolic disorders. Eur J Nutr. 2016;55(8):2339–2345. doi:10.1007/s00394-016-1212-2
27. Lin F, Yang Y, Wei S, et al. Hydrogen sulfide protects against high glucose-induced human umbilical vein endothelial cell injury through activating PI3K/Akt/eNOS pathway. Drug Des Devel Ther. 2020;14:621–633. doi:10.2147/DDDT.S242521
28. Penna C, Pagliaro P. Endothelial dysfunction: redox imbalance, nlrp3 inflammasome, and inflammatory responses in cardiovascular diseases. Antioxidants. 2025;14(3). doi:10.3390/antiox14030256
29. Wang X, He B. Endothelial dysfunction: molecular mechanisms and clinical implications. MedComm. 2024;5(8):e651. doi:10.1002/mco2.651
30. Singh S, Siva BV, Ravichandiran V. Advanced glycation end products: key player of the pathogenesis of atherosclerosis. Glycoconjugate J. 2022;39(4):547–563. doi:10.1007/s10719-022-10063-x
31. Batchuluun B, Inoguchi T, Sonoda N, et al. Metformin and liraglutide ameliorate high glucose-induced oxidative stress via inhibition of PKC-NAD(P)H oxidase pathway in human aortic endothelial cells. Atherosclerosis. 2014;232(1):156–164. doi:10.1016/j.atherosclerosis.2013.10.025
32. Bei Y, Zhang S, Song Y, et al. EPSTI1 promotes monocyte adhesion to endothelial cells in vitro via upregulating VCAM-1 and ICAM-1 expression. Acta Pharmacol Sin. 2023;44(1):71–80. doi:10.1038/s41401-022-00923-5
33. AV P, VA O, VN S, et al. Overview of atherosclerotic plaque: from formation to complication. Online J Biol Sci. 2024;24(3). doi:10.3844/ojbsci.2024.382.394
34. Hajibabaie F, Abedpoor N, Mohamadynejad P. Types of cell death from a molecular perspective. Biology. 2023;12(11). doi:10.3390/biology12111426
35. Wei C, Shi Y, Zhang H, et al. The role of endothelial cells in the formation and progression of atherosclerosis. 2023.
36. Zhang S, Jin S, Zhang S, et al. Vitexin protects against high glucose-induced endothelial cell apoptosis and oxidative stress via Wnt/beta-catenin and Nrf2 signalling pathway. Arch Physiol Biochem. 2024;130(3):275–284. doi:10.1080/13813455.2022.2028845
37. Li J, Xu J, Wang C, et al. Knockdown of long non-coding RNA-MALAT1 ameliorates diabetic lower limb atherosclerotic disease through MiR-17-5p-Mediated endothelial cell pyroptosis. Biochem Genet. 2025. doi:10.1007/s10528-025-11236-7
38. SD S, Fiedler J, Bauersachs J, et al. Senescence-induced inflammation: an important player and key therapeutic target in atherosclerosis. Eur Heart J. 2020;41(31):2983–2996. doi:10.1093/eurheartj/ehz919
39. Childs BG, Li H, van Deursen JM. Senescent cells: a therapeutic target for cardiovascular disease. J Clin Invest. 2018;128(4):1217–1228. doi:10.1172/JCI95146
40. BG C, DJ B, Wijshake T, et al. Senescent intimal foam cells are deleterious at all stages of atherosclerosis. Science. 2016;354(6311):472–477. doi:10.1126/science.aaf6659
41. DG S, EC B, JAF D, et al. Vasa vasorum angiogenesis: key player in the initiation and progression of atherosclerosis and potential target for the treatment of cardiovascular disease. Front Immunol. 2018;9:706. doi:10.3389/fimmu.2018.00706
42. Boyle EC, Sedding DG, Haverich A. Targeting vasa vasorum dysfunction to prevent atherosclerosis. Vasc Pharmacol. 2017;96-98:5–10. doi:10.1016/j.vph.2017.08.003
43. Wang G, Han B, Zhang R, et al. C1q/TNF-related protein 9 attenuates atherosclerosis by inhibiting hyperglycemia-induced endothelial cell senescence through the AMPKalpha/KLF4 signaling pathway. Front Pharmacol. 2021;12:758792. doi:10.3389/fphar.2021.758792
44. Li M, Liu Z, Cao X, et al. [Gly14]-Humanin ameliorates high glucose-induced endothelial senescence via SIRT6. Sci Rep. 2024;14(1):30924. doi:10.1038/s41598-024-81878-x
45. Shen J, Lai Y, Lu Y, et al. NRF2-HIF2alpha signaling attenuates endothelial cell senescence and maintains intercellular junctions in diabetes. Int J Biol Sci. 2024;20(10):4055–4073. doi:10.7150/ijbs.96719
46. Denimal D. Carbamylated lipoproteins in diabetes. World J Diab. 2023;14(3):159–169. doi:10.4239/wjd.v14.i3.159
47. Cervantes J, Kanter JE. Monocyte and macrophage foam cells in diabetes-accelerated atherosclerosis. Front Cardiovasc Med. 2023;10:1213177. doi:10.3389/fcvm.2023.1213177
48. Strizova Z, Benesova I, Bartolini R, et al. M1/M2 macrophages and their overlaps - myth or reality? Clin Sci. 2023;137(15):1067–1093. doi:10.1042/CS20220531
49. Liu J, Meng H, Chen L, et al. OCT-based vulnerable plaque features and MACE prediction in premature coronary artery disease. Front Cardiovasc Med. 2025;12:1621824. doi:10.3389/fcvm.2025.1621824
50. Saki N, Haybar H, Maniati M, et al. Modification macrophage to foam cells in atherosclerosis disease: some factors stimulate or inhibit this process. J Diab Metab Disord. 2024;23(2):1687–1697. doi:10.1007/s40200-024-01482-8
51. Bi Y, Chen J, Hu F, et al. M2 macrophages as a potential target for antiatherosclerosis treatment. Neural Plast. 2019;2019:6724903. doi:10.1155/2019/6724903
52. Chen R, Zhang H, Tang B, et al. Macrophages in cardiovascular diseases: molecular mechanisms and therapeutic targets. Signal Transduction Targeted Ther. 2024;9(1):130. doi:10.1038/s41392-024-01840-1
53. Verweij SL, Duivenvoorden R, Stiekema LCA, et al. CCR2 expression on circulating monocytes is associated with arterial wall inflammation assessed by 18F-FDG PET/CT in patients at risk for cardiovascular disease. Cardiovasc Res. 2018;114(3):468–475. doi:10.1093/cvr/cvx224
54. Tomas L, Edsfeldt A, Mollet IG, et al. Altered metabolism distinguishes high-risk from stable carotid atherosclerotic plaques. Eur Heart J. 2018;39(24):2301–2310. doi:10.1093/eurheartj/ehy124
55. Liu Y, Guo F, Qiu W, et al. Metabolic reprogramming and cell interaction in atherosclerosis: from molecular mechanisms to therapeutic strategies. J Cardiovasc Dev Dis. 2025;12(10):384. doi:10.3390/jcdd12100384
56. Cornwell A, Ziółkowski H, Badiei A. Glucose transporter Glut1-dependent metabolic reprogramming regulates lipopolysaccharide-induced inflammation in RAW264.7 macrophages. Biomolecules. 2023;13(5):770. doi:10.3390/biom13050770
57. B GDS, Goedeke L. Macrophage immunometabolism in diabetes-associated atherosclerosis. Immunometabolism. 2023;5(4):e32. doi:10.1097/IN9.0000000000000032
58. Zhang L, Li L, Li Y, et al. Disruption of COMMD1 accelerates diabetic atherosclerosis by promoting glycolysis. Diabetes Vasc Dis Res. 2023;20(1):1497001697. doi:10.1177/14791641231159009
59. Edgar L, Akbar N, Braithwaite AT, et al. Hyperglycemia induces trained immunity in macrophages and their precursors and promotes atherosclerosis. Circulation. 2021;144(12):961–982. doi:10.1161/CIRCULATIONAHA.120.046464
60. Gill V, Kumar V, Singh K, et al. Advanced glycation end products (AGEs) may be a striking link between modern diet and health. Biomolecules. 2019;9(12):888. doi:10.3390/biom9120888
61. Khalid M, Petroianu G, Adem A. Advanced glycation end products and diabetes mellitus: mechanisms and perspectives. Biomolecules. 2022;12(4):542. doi:10.3390/biom12040542
62. Han X, Ma W, Zhu Y, et al. Advanced glycation end products enhance macrophage polarization to the M1 phenotype via the HIF-1α/PDK4 pathway. Mol Cell Endocrinol. 2020;514:110878. doi:10.1016/j.mce.2020.110878
63. Hamasaki S, Kobori T, Yamazaki Y, et al. Effects of scavenger receptors-1 class A stimulation on macrophage morphology and highly modified advanced glycation end product-protein phagocytosis. Sci Rep. 2018;8(1):5901. doi:10.1038/s41598-018-24325-y
64. Poznyak AV, Nikiforov NG, Starodubova AV, et al. Macrophages and foam cells: brief overview of their role, linkage, and targeting potential in atherosclerosis. Biomedicines. 2021;9(9):1221. doi:10.3390/biomedicines9091221
65. Tall AR, Westerterp M. Inflammasomes, neutrophil extracellular traps, and cholesterol. J Lipid Res. 2019;60(4):721–727. doi:10.1194/jlr.S091280
66. Han Y, Qiu H, Pei X, et al. Low-dose sinapic acid abates the pyroptosis of macrophages by downregulation of lncRNA-MALAT1 in rats with diabetic atherosclerosis. J Cardiovasc Pharmacol. 2018;71(2):104–112. doi:10.1097/FJC.0000000000000550
67. Qiu Y, Li L, Guo X, et al. Exogenous spermine inhibits high glucose/oxidized LDL-induced oxidative stress and macrophage pyroptosis by activating the Nrf2 pathway. Exp Ther Med. 2022;23(4):310. doi:10.3892/etm.2022.11239
68. Suzuki Y. Ca(2+) microdomains in vascular smooth muscle cells: roles in vascular tone regulation and hypertension. J Pharmacol Sci. 2025;158(1):59–67. doi:10.1016/j.jphs.2025.03.008
69. Gomez D, Owens GK. Smooth muscle cell phenotypic switching in atherosclerosis. Cardiovasc Res. 2012;95(2):156–164. doi:10.1093/cvr/cvs115
70. Pham TH, Trang NM, Kim E, et al. Citropten inhibits vascular smooth muscle cell proliferation and migration via the TRPV1 receptor. ACS Omega. 2024;9(27):29829–29839. doi:10.1021/acsomega.4c03539
71. Bao Z, Li L, Geng Y, et al. Advanced glycation end products induce vascular smooth muscle cell-derived foam cell formation and transdifferentiate to a macrophage-like state. Mediators Inflamm. 2020;2020:6850187. doi:10.1155/2020/6850187
72. Ketsawatsomkron P, Stepp DW, Fulton DJ, et al. Molecular mechanism of angiotensin II-induced insulin resistance in aortic vascular smooth muscle cells: roles of protein tyrosine phosphatase-1B. Vasc Pharmacol. 2010;53(3–4):160–168. doi:10.1016/j.vph.2010.06.001
73. Shaw S, Wang X, Redd H, et al. High glucose augments the angiotensin II-induced activation of JAK2 in vascular smooth muscle cells via the polyol pathway. J Biol Chem. 2003;278(33):30634–30641. doi:10.1074/jbc.M305008200
74. Liao X, Wang N, Zhao D, et al. STAT3 protein regulates vascular smooth muscle cell phenotypic switch by interaction with myocardin. J Biol Chem. 2015;290(32):19641–19652. doi:10.1074/jbc.M114.630111
75. Zhang M, Zheng B, Tong F, et al. iNOS-derived peroxynitrite mediates high glucose-induced inflammatory gene expression in vascular smooth muscle cells through promoting KLF5 expression and nitration. Biochim Biophys Acta Mol Basis Dis. 2017;1863(11):2821–2834. doi:10.1016/j.bbadis.2017.07.004
76. Yang B, Gao X, Sun Y, et al. Dihydroartemisinin alleviates high glucose-induced vascular smooth muscle cells proliferation and inflammation by depressing the miR-376b-3p/KLF15 pathway. Biochem Biophys Res Commun. 2020;530(3):574–580. doi:10.1016/j.bbrc.2020.07.095
77. Carrillo-Sepulveda MA, Spitler K, Pandey D, et al. Inhibition of TLR4 attenuates vascular dysfunction and oxidative stress in diabetic rats. J Mol Med. 2015;93(12):1341–1354. doi:10.1007/s00109-015-1318-7
78. Yang M, Fang J, Liu Q, et al. Role of ROS-TRPM7-ERK1/2 axis in high concentration glucose-mediated proliferation and phenotype switching of rat aortic vascular smooth muscle cells. Biochem Biophys Res Commun. 2017;494(3–4):526–533. doi:10.1016/j.bbrc.2017.10.122
79. Xu Z, Xu J, Lei W, et al. RANBP1 regulates NOTCH3-mediated autophagy in high glucose-induced vascular smooth muscle cells. Front Biosci. 2025;30(2):26850. doi:10.31083/FBL26850
80. Yang J, Gourley GR, Gilbertsen A, et al. High glucose levels promote switch to synthetic vascular smooth muscle cells via Lactate/GPR81. Cells. 2024;13(3):236. doi:10.3390/cells13030236
81. Wang K, Deng X, Shen Z, et al. High glucose promotes vascular smooth muscle cell proliferation by upregulating proto-oncogene serine/threonine-protein kinase Pim-1 expression. Oncotarget. 2017;8(51):88320–88331. doi:10.18632/oncotarget.19368
82. Grootaert MOJ, Bennett MR. Vascular smooth muscle cells in atherosclerosis: time for a re-assessment. Cardiovasc Res. 2021;117(11):2326–2339. doi:10.1093/cvr/cvab046
83. Guo J, Du L. Regulation of osteogenic differentiation in vascular smooth muscle cells under high-glucose condition. Front Endocrinol. 2025;16:1589160. doi:10.3389/fendo.2025.1589160
84. Lehners M, Schmidt H, Zaldivia MTK, et al. Single-cell analysis identifies the CNP/GC-B/cGMP axis as marker and regulator of modulated VSMCs in atherosclerosis. Nat Commun. 2025;16(1):429. doi:10.1038/s41467-024-55687-9
85. Gao Y, Zhang L, Zhang F, et al. Traditional Chinese medicine and its active substances reduce vascular injury in diabetes via regulating autophagic activity. Front Pharmacol. 2024;15:1355246. doi:10.3389/fphar.2024.1355246
86. Ao L, Chen Z, Yin J, et al. Chinese herbal medicine and active ingredients for diabetic cardiomyopathy: molecular mechanisms regulating endoplasmic reticulum stress. Front Pharmacol. 2023;14:1290023. doi:10.3389/fphar.2023.1290023
87. Leng Y, Fu X, Qian L, et al. Efficacy, safety and therapeutic mechanism of Shen-Qi Xiao-Tan formula in the treatment of peripheral atherosclerosis in patients with type 2 diabetes mellitus: a randomized, double-blind, placebo-controlled trial protocol. BMC Complement Med Therap. 2022;22(1):337. doi:10.1186/s12906-022-03813-9
88. Zhu Q, Kang J, Xu G, et al. Traditional Chinese medicine Shenqi compound to improve lower extremity atherosclerosis of patients with type 2 diabetes by affecting blood glucose fluctuation: study protocol for a randomized controlled multicenter trial. Medicine. 2020;99(11):e19501. doi:10.1097/MD.0000000000019501
89. Ko YS, Jin H, Park SW, et al. Salvianolic acid B protects against oxLDL-induced endothelial dysfunction under high-glucose conditions by downregulating ROCK1-mediated mitophagy and apoptosis. Biochem Pharmacol. 2020;174:113815. doi:10.1016/j.bcp.2020.113815
90. Meng Q, Huang Y, Long X, et al. Scutellarin mitigates high glucose-induced pyroptosis in diabetic atherosclerosis: role of Nrf2-FBXL2-mediated NLRP3 degradation. Endocr J. 2025;72(5):495–507. doi:10.1507/endocrj.EJ24-0505
91. Rong J, Li C, Zhang Q, et al. Hydroxysafflor yellow A inhibits endothelial cell ferroptosis in diabetic atherosclerosis mice by regulating miR-429/SLC7A11. Pharm Biol. 2023;61(1):404–415. doi:10.1080/13880209.2023.2225543
92. Wang L, Fan Y, Li B, et al. Maslinic acid suppresses high glucose-induced inflammation by epigenetically inhibiting TXNIP expression. Current Med Sci. 2022;42(6):1213–1219. doi:10.1007/s11596-022-2657-6
93. Tang H, Li K, Zhang S, et al. Inhibitory effect of paeonol on apoptosis, oxidative stress, and inflammatory response in human umbilical vein endothelial cells induced by high glucose and palmitic acid induced through regulating SIRT1/FOXO3a/NF-κB pathway. J Interferon Cytokine Res. 2021;41(3):111–124. doi:10.1089/jir.2019.0236
94. Qiu Y, Chao C, Jiang L, et al. Citronellal alleviate macro- and micro-vascular damage in high fat diet/streptozotocin - Induced diabetic rats via a S1P/S1P1 dependent signaling pathway. Eur J Pharmacol. 2022;920:174796. doi:10.1016/j.ejphar.2022.174796
95. Cui Q, Du H, Ma Y, et al. Matrine inhibits advanced glycation end products-induced macrophage M1 polarization by reducing DNMT3a/b-mediated DNA methylation of GPX1 promoter. Eur J Pharmacol. 2022;926:175039. doi:10.1016/j.ejphar.2022.175039
96. Zhao Y, Zheng J, Yu Y, et al. Panax notoginseng saponins regulate macrophage polarization under hyperglycemic condition via NF-κB signaling pathway. Biomed Res Int. 2018;2018:9239354. doi:10.1155/2018/9239354
97. Sha W, Zhao B, Wei H, et al. Astragalus polysaccharide ameliorates vascular endothelial dysfunction by stimulating macrophage M2 polarization via potentiating Nrf2/HO-1 signaling pathway. Phytomedicine. 2023;112:154667. doi:10.1016/j.phymed.2023.154667
98. Chen T, Yen C, Lu Y, et al. The antagonism of 6-shogaol in high-glucose-activated NLRP3 inflammasome and consequent calcification of human artery smooth muscle cells. Cell Biosci. 2020;10:5. doi:10.1186/s13578-019-0372-1
99. Wu W, Yan H, Wang X, et al. Sodium tanshinone IIA silate inhibits high glucose-induced vascular smooth muscle cell proliferation and migration through activation of AMP-activated protein kinase. PLoS One. 2014;9(4):e94957. doi:10.1371/journal.pone.0094957
100. Tong W, Leng L, Wang Y, et al. Buyang huanwu decoction inhibits diabetes-accelerated atherosclerosis via reduction of AMPK-Drp1-mitochondrial fission axis. J Ethnopharmacol. 2023;312:116432. doi:10.1016/j.jep.2023.116432
101. Wang W, Wang S, Li Y, et al. Network pharmacology, molecular docking, and in vitro experimental verification of the mechanism of Guanxining in treating diabetic atherosclerosis. J Ethnopharmacol. 2024;324:117792. doi:10.1016/j.jep.2024.117792
102. Zhang Y, Wang R, Tan H, et al. Fufang Zhenzhu Tiaozhi (FTZ) capsule ameliorates diabetes-accelerated atherosclerosis via suppressing YTHDF2-mediated m(6)A modification of SIRT3 mRNA. J Ethnopharmacol. 2023;317:116766. doi:10.1016/j.jep.2023.116766
103. Zhou X, Fouda S, Zeng X, et al. Characterization of the therapeutic profile of albiflorin for the metabolic syndrome. Front Pharmacol. 2019;10:1151. doi:10.3389/fphar.2019.01151
104. Wu Y, Xu S, Tian XY. The effect of salvianolic acid on vascular protection and possible mechanisms. Oxid Med Cell Longev. 2020;2020:5472096. doi:10.1155/2020/5472096
105. Huang M, Zhou C, Zhang Y, et al. Salvianolic Acid B ameliorates hyperglycemia and dyslipidemia in db/db mice through the AMPK pathway. Cell Physiol Biochem. 2016;40(5):933–943. doi:10.1159/000453151
106. Yang X, Wang W, Huang Y, et al. Network pharmacology-based dissection of the active ingredients and protective mechanism of the salvia miltiorrhiza and panax notoginseng herb pair against insulin resistance. ACS Omega. 2021;6(27):17276–17288. doi:10.1021/acsomega.1c01209
107. Shi Y, Pan D, Yan L, et al. Salvianolic acid B improved insulin resistance through suppression of hepatic ER stress in ob/ob mice. Biochem Biophys Res Commun. 2020;526(3):733–737. doi:10.1016/j.bbrc.2020.03.124
108. Wang L, Ma Q. Clinical benefits and pharmacology of scutellarin: a comprehensive review. Pharmacol Ther. 2018;190:105–127. doi:10.1016/j.pharmthera.2018.05.006
109. Liu Y, Jing Y, Zeng C, et al. Scutellarin suppresses NLRP3 inflammasome activation in macrophages and protects mice against bacterial sepsis. Front Pharmacol. 2018;8:975. doi:10.3389/fphar.2017.00975
110. Huang H, Geng Q, Yao H, et al. Protective effect of scutellarin on myocardial infarction induced by isoprenaline in rats. Iran J Basic Med Sci. 2018;21(3):267–276. doi:10.22038/ijbms.2018.26110.6415
111. Gao L, Tang H, Zeng Q, et al. The anti-insulin resistance effect of scutellarin may be related to antioxidant stress and AMPKα activation in diabetic mice. Obes Res Clin Pract. 2020;14(4):368–374. doi:10.1016/j.orcp.2020.06.005
112. Yang L, Xiao N, Liu J, et al. Differential regulation of baicalin and scutellarin on AMPK and Akt in promoting adipose cell glucose disposal. Biochim Biophys Acta Mol Basis Dis. 2017;1863(2):598–606. doi:10.1016/j.bbadis.2016.11.024
113. Yan W, Cheng J, Xu B. Dietary flavonoids vitexin and isovitexin: new insights into their functional roles in human health and disease prevention. Int J Mol Sci. 2025;26(14):6997. doi:10.3390/ijms26146997
114. Zhang L, Shi L, Han J, et al. Protection of β-pancreatic cells from dysfunctionality of insulin using vitexin by apoptosis of INS-1 cells. Arch Physiol Biochem. 2023;129(5):1160–1167. doi:10.1080/13813455.2021.1910714
115. Choo CY, Sulong NY, Man F, et al. Vitexin and isovitexin from the Leaves of Ficus deltoidea with in-vivo α-glucosidase inhibition. J Ethnopharmacol. 2012;142(3):776–781. doi:10.1016/j.jep.2012.05.062
116. Hou J, Wang C, Zhang M, et al. Safflower yellow improves the synaptic structural plasticity by ameliorating the disorder of glutamate circulation in Aβ1-42-induced AD model rats. Neurochem Res. 2020;45(8):1870–1887. doi:10.1007/s11064-020-03051-w
117. Lee M, Li H, Zhao H, et al. Effects of hydroxysafflor Yellow A on the PI3K/AKT pathway and apoptosis of pancreatic β-Cells in type 2 diabetes mellitus rats. Diab Metab Syndrome Obes. 2020;13:1097–1107. doi:10.2147/DMSO.S246381
118. Xu Y, Lee J, Park Y, et al. Molecular dynamics simulation integrating the inhibition kinetics of hydroxysafflor yellow A on α-glucosidase. J Biomol Struct Dyn. 2018;36(4):830–840. doi:10.1080/07391102.2017.1300544
119. Liu S, Zhang Y, Wang Y, et al. Upregulation of heme oxygenase-1 expression by hydroxysafflor yellow A conferring protection from anoxia/reoxygenation-induced apoptosis in H9c2 cardiomyocytes. Int J Cardiol. 2012;160(2):95–101. doi:10.1016/j.ijcard.2011.03.033
120. Xie Y, Guo Y, Cao S, et al. Hydroxysafflor yellow a attenuates hydrogen peroxide-induced oxidative damage on human umbilical vein endothelial cells. Evidence-Based Complem Altern Med. 2020;2020(1):8214128. doi:10.1155/2020/8214128
121. Zhou P, Xie W, He S, et al. Ginsenoside Rb1 as an anti-diabetic agent and its underlying mechanism analysis. Cells. 2019;8(3):204. doi:10.3390/cells8030204
122. Lu W, Lin Y, Haider N, et al. Ginsenoside Rb1 protects human vascular smooth muscle cells against resistin-induced oxidative stress and dysfunction. Front Cardiovasc Med. 2023;10:1164547. doi:10.3389/fcvm.2023.1164547
123. Zheng X, Wang S, Zou X, et al. Ginsenoside Rb1 improves cardiac function and remodeling in heart failure. Exp Animals. 2017;66(3):217–228. doi:10.1538/expanim.16-0121
124. Chen Y, Yan D, Wu C, et al. Maslinic acid prevents IL-1β-induced inflammatory response in osteoarthritis via PI3K/AKT/NF-κB pathways. J Cell Physiol. 2021;236(3):1939–1949. doi:10.1002/jcp.29977
125. Liu X, Zang X, Yin X, et al. Semi-synthesis of C28-modified triterpene acid derivatives from maslinic acid or corosolic acid as potential α-glucosidase inhibitors. Bioorg Chem. 2020;97:103694. doi:10.1016/j.bioorg.2020.103694
126. Guan T, Qian Y, Tang X, et al. Maslinic acid, a natural inhibitor of glycogen phosphorylase, reduces cerebral ischemic injury in hyperglycemic rats by GLT-1 up-regulation. J Neurosci Res. 2011;89(11):1829–1839. doi:10.1002/jnr.22671
127. Yang C, Cheng J, Zhu Q, et al. Review of the protective mechanism of paeonol on cardiovascular disease. Drug Design Dev Ther. 2023;17:2193–2208. doi:10.2147/DDDT.S414752
128. Shi X, Xie X, Sun Y, et al. Paeonol inhibits NLRP3 mediated inflammation in rat endothelial cells by elevating hyperlipidemic rats plasma exosomal miRNA-223. Eur J Pharmacol. 2020;885:173473. doi:10.1016/j.ejphar.2020.173473
129. Liu Y, Chen J, Dai M. Paeonol protects rat vascular endothelial cells from ox-LDL-induced injury in vitro via downregulating microRNA-21 expression and TNF-α release. Acta Pharmacol Sin. 2014;35(4):483–488. doi:10.1038/aps.2013.190
130. Xu F, Xiao H, Liu R, et al. Paeonol ameliorates glucose and lipid metabolism in experimental diabetes by activating Akt. Front Pharmacol. 2019;10:261. doi:10.3389/fphar.2019.00261
131. Zhang L, Li D, Liu L. Paeonol: pharmacological effects and mechanisms of action. Int Immunopharmacol. 2019;72:413–421. doi:10.1016/j.intimp.2019.04.033
132. Quintans-Júnior LJ, Souza TT, Leite BS, et al. Phythochemical screening and anticonvulsant activity of Cymbopogon winterianus Jowitt (Poaceae) leaf essential oil in rodents. Phytomedicine. 2008;15(8):619–624. doi:10.1016/j.phymed.2007.09.018
133. Srinivasan S, Muruganathan U. Antidiabetic efficacy of citronellol, a citrus monoterpene by ameliorating the hepatic key enzymes of carbohydrate metabolism in streptozotocin-induced diabetic rats. Chem Biol Interact. 2016;250:38–46. doi:10.1016/j.cbi.2016.02.020
134. Lu J, Guo C, Ou W, et al. Citronellal prevents endothelial dysfunction and atherosclerosis in rats. J Cell Biochem. 2019;120(3):3790–3800. doi:10.1002/jcb.27660
135. Chen S, Wang Y, Liang C, et al. Buyang Huanwu Decoction ameliorates atherosclerosis by regulating TGF-β/Smad2 pathway to promote the differentiation of regulatory T cells. J Ethnopharmacol. 2021;269:113724. doi:10.1016/j.jep.2020.113724
136. She Y, Shao L, Zhang Y, et al. Neuroprotective effect of glycosides in Buyang Huanwu Decoction on pyroptosis following cerebral ischemia-reperfusion injury in rats. J Ethnopharmacol. 2019;242:112051. doi:10.1016/j.jep.2019.112051
137. Zhou H, Lin B, Yang J, et al. Analysis of the mechanism of Buyang Huanwu Decoction against cerebral ischemia-reperfusion by multi-omics. J Ethnopharmacol. 2023;305:116112. doi:10.1016/j.jep.2022.116112
138. Liu M, Zhao Q, Liu J, et al. Buyang Huanwu decoction affects gut microbiota and lipid metabolism in a ZDF rat model of co-morbid type 2 diabetes mellitus and obesity: an integrated metabolomics analysis. Front Chem. 2022;10:1036380. doi:10.3389/fchem.2022.1036380
139. Wang M, Yang Q, Ying X, et al. Network pharmacology-based approach uncovers the mechanism of guanxinning tablet for treating thrombus by MAPKs signal pathway. Front Pharmacol. 2020;11:652. doi:10.3389/fphar.2020.00652
140. Yang Q, Xu Y, Shen L, et al. Guanxinning tablet attenuates coronary atherosclerosis via regulating the gut microbiota and their metabolites in tibetan minipigs induced by a high-fat diet. J Immunol Res. 2022;2022:7128230. doi:10.1155/2022/7128230
141. Li Y, Chen J, Tu H, et al. Protective effects of GuanXinNing tablet (GXNT) on diabetic encephalopathy in zucker diabetic obesity (ZDF) rats. BMC Complement Med Therap. 2023;23(1):385. doi:10.1186/s12906-023-04195-2
142. Song L, Zhang D, Guo C, et al. The traditional Chinese medicine formula Fufang-Zhenzhu-Tiaozhi protects myocardia from injury in diabetic minipigs with coronary heart disease. Biomed Pharmacothe. 2021;137:111343. doi:10.1016/j.biopha.2021.111343
143. Wang H, Tan H, Zhan W, et al. Molecular mechanism of Fufang Zhenzhu Tiaozhi capsule in the treatment of type 2 diabetes mellitus with nonalcoholic fatty liver disease based on network pharmacology and validation in minipigs. J Ethnopharmacol. 2021;274:114056. doi:10.1016/j.jep.2021.114056
144. Hou H, Zhang Q, Dong H, et al. Matrine improves diabetic cardiomyopathy through TGF-β-induced protein kinase RNA-like endoplasmic reticulum kinase signaling pathway. J Cell Biochem. 2019;120(8):13573–13582. doi:10.1002/jcb.28632
145. Guo S, Yan T, Shi L, et al. Matrine, as a CaSR agonist promotes intestinal GLP-1 secretion and improves insulin resistance in diabetes mellitus. Phytomedicine. 2021;84:153507. doi:10.1016/j.phymed.2021.153507
146. Zhao L, Cai H, Tang Z, et al. Matrine suppresses advanced glycation end products-induced human coronary smooth muscle cells phenotype conversion by regulating endoplasmic reticulum stress-dependent Notch signaling. Eur J Pharmacol. 2020;882:173257. doi:10.1016/j.ejphar.2020.173257
147. Ma W, Xu J, Zhang Y, et al. Matrine pre-treatment suppresses AGEs- induced HCSMCs fibrotic responses by regulating Poldip2/mTOR pathway. Eur J Pharmacol. 2019;865:172746. doi:10.1016/j.ejphar.2019.172746
148. Kitamura K, Takamura Y, Iwamoto T, et al. Dammarane-type triterpene extracts of Panax notoginseng root ameliorates hyperglycemia and insulin sensitivity by enhancing glucose uptake in skeletal muscle. Biosci Biotechnol Biochem. 2017;81(2):335–342. doi:10.1080/09168451.2016.1246173
149. Uzayisenga R, Ayeka PA, Wang Y. Anti-diabetic potential of Panax notoginseng saponins (PNS): a review. Phytother Res. 2014;28(4):510–516. doi:10.1002/ptr.5026
150. Shen Q, Li J, Zhang C, et al. Panax notoginseng saponins reduce high-risk factors for thrombosis through peroxisome proliferator-activated receptor -γ pathway. Biomed Pharmacothe. 2017;96:1163–1169. doi:10.1016/j.biopha.2017.11.106
151. Feng S, Ding H, Liu L, et al. Astragalus polysaccharide enhances the immune function of RAW264.7 macrophages via the NF-κB p65/MAPK signaling pathway. Exp Ther Med. 2021;21(1):20. doi:10.3892/etm.2020.9452
152. Liu S, Wang L, Zhang Z, et al. The potential of astragalus polysaccharide for treating diabetes and its action mechanism. Front Pharmacol. 2024;15:1339406. doi:10.3389/fphar.2024.1339406
153. Yang Z, Wang Y, Chen S. Astragalus polysaccharide alleviates type 2 diabetic rats by reversing the glucose transporters and sweet taste receptors/GLP-1/GLP-1 receptor signaling pathways in the intestine-pancreatic axis. J Funct Foods. 2021;76:104310. doi:10.1016/j.jff.2020.104310
154. Wang YK, Hong YJ, Yao YH, et al. 6-shogaol protects against oxidized LDL-induced endothelial injruries by inhibiting oxidized LDL-Evoked LOX-1 signaling. Evidence-Based Complem Altern Med. 2013;2013:503521. doi:10.1155/2013/503521
155. Kou X, Wang X, Ji R, et al. Occurrence, biological activity and metabolism of 6-shogaol. Food Funct. 2018;9(3):1310–1327. doi:10.1039/c7fo01354j
156. Yi J, Ryoo Z, Ha J, et al. Beneficial effects of 6-shogaol on hyperglycemia, islet morphology and apoptosis in some tissues of streptozotocin-induced diabetic mice. Diabetol Metab Syndr. 2019;11:15. doi:10.1186/s13098-019-0407-0
157. Wei C, Tsai Y, Korinek M, et al. 6-Paradol and 6-Shogaol, the pungent compounds of ginger, promote glucose utilization in adipocytes and myotubes, and 6-Paradol reduces blood glucose in high-fat diet-fed mice. Int J Mol Sci. 2017;18(1):168. doi:10.3390/ijms18010168
158. Hwang S, Yang JH, Jeong Y, et al. Tanshinone IIA improves endoplasmic reticulum stress-induced insulin resistance through AMP-activated protein kinase. Biochem Biophys Res Commun. 2013;430(4):1246–1252. doi:10.1016/j.bbrc.2012.12.066
159. Genitsaridi I, Salpea P, Salim A, et al. 11th edition of the IDF Diabetes Atlas: global, regional, and national diabetes prevalence estimates for 2024 and projections for 2050. Lancet Diab Endocrinol. 2026;14(2):149–156. doi:10.1016/S2213-8587(25)00299-2
160. Wong ND, Sattar N. Cardiovascular risk in diabetes mellitus: epidemiology, assessment and prevention. Nat Rev Cardiol. 2023;20(10):685–695. doi:10.1038/s41569-023-00877-z
161. Lu Y, Wang W, Liu J, et al. Vascular complications of diabetes: a narrative review. Medicine. 2023;102(40):e35285. doi:10.1097/MD.0000000000035285
162. Li Y, Liu Y, Liu S, et al. Diabetic vascular diseases: molecular mechanisms and therapeutic strategies. Signal Transduction Targeted Ther. 2023;8(1):152. doi:10.1038/s41392-023-01400-z
163. Gong S, Li Y, Yan K, et al. The crosstalk between endothelial cells, smooth muscle cells, and macrophages in atherosclerosis. Int J Mol Sci. 2025;26(4):1457. doi:10.3390/ijms26041457
164. Khan AW, Jandeleit-Dahm KAM. Atherosclerosis in diabetes mellitus: novel mechanisms and mechanism-based therapeutic approaches. Nat Rev Cardiol. 2025;22(7):482–496. doi:10.1038/s41569-024-01115-w
165. Chu S, Wang H, Liu D, et al. Heqi San alleviates diabetic atherosclerosis and alters serum metabolic profiles in rats. BMC Cardiovasc Disorders. 2025;25(1):610. doi:10.1186/s12872-025-05012-z
166. Shi G, Yu D, Wu J, et al. Adjuvant qi-invigorating blood-activating tongmai decoction promotes effect of rosuvastatin on senile diabetes mellitus complicated with atherosclerosis. Am J Transl Res. 2023;15(5):3433–3441.
167. Parker S, May B, Zhang C, et al. A pharmacological review of bioactive constituents of paeonia lactiflora pallas and paeonia veitchii lynch. Phytother Res. 2016;30(9):1445–1473. doi:10.1002/ptr.5653
168. Yang L, Huang X, Wang Z, et al. Research progress on the pharmacological properties of active ingredients from Salvia miltiorrhiza: a review. Phytomedicine. 2025;148:157272. doi:10.1016/j.phymed.2025.157272
169. Wei B, Sun C, Wan H, et al. Bioactive components and molecular mechanisms of Salvia miltiorrhiza Bunge in promoting blood circulation to remove blood stasis. J Ethnopharmacol. 2023;317:116697. doi:10.1016/j.jep.2023.116697
170. Fan H, Lin P, Kang Q, et al. Metabolism and Pharmacological Mechanisms of Active Ingredients in Erigeron breviscapus. Current Drug Metab. 2021;22(1):24–39. doi:10.2174/1389200221666201217093255
171. Gao J, Chen G, He H, et al. Therapeutic effects of breviscapine in cardiovascular diseases: a review. Front Pharmacol. 2017;8:289. doi:10.3389/fphar.2017.00289
172. Orhan IE. Phytochemical and pharmacological activity profile of Crataegus oxyacantha L. (Hawthorn) - A Cardiotonic herb. Curr Med Chem. 2018;25(37):4854–4865. doi:10.2174/0929867323666160919095519
173. Wang J, Xiong X, Feng B. Effect of crataegus usage in cardiovascular disease prevention: an evidence-based approach. Evidence-Based Complem Altern Med. 2013;2013:149363. doi:10.1155/2013/149363
174. Bai H, Yang J, Wang R. Carthamus tinctorius L.: a comprehensive review of its ethnomedicine, phytochemistry, pharmacology, and clinical applications. Front Pharmacol. 2025;16:1609299. doi:10.3389/fphar.2025.1609299
175. Li J, Xu D, Cui D, et al. Exploring the structure-activity relationship of Safflower polysaccharides: from the structural characteristics to biological function and therapeutic applications. J Ethnopharmacol. 2025;339:119131. doi:10.1016/j.jep.2024.119131
176. Wang Z, He C, Peng Y, et al. Origins, phytochemistry, pharmacology, analytical methods and safety of Cortex Moutan (Paeonia suffruticosa Andrew): a systematic review. Molecules. 2017;22(6):946. doi:10.3390/molecules22060946
177. JJ T, Yi Q, Wang X, et al. Corrigendum: review of phytomedicine, phytochemistry, ethnopharmacology, toxicology, and pharmacological activities of Cymbopogon genus. Front Pharmacol. 2022;13:1109233. doi:10.3389/fphar.2022.1109233
178. Ekpenyong CE, Akpan E, Nyoh A. Ethnopharmacology, phytochemistry, and biological activities of Cymbopogon citratus (DC.) Stapf extracts. Chin J Nat Med. 2015;13(5):321–337. doi:10.1016/S1875-5364(15)30023-6
179. Fu J, Wang Z, Huang L, et al. Review of the botanical characteristics, phytochemistry, and pharmacology of Astragalus membranaceus (Huangqi). Phytother Res. 2014;28(9):1275–1283. doi:10.1002/ptr.5188
180. Shen J, Qin H, Li K, et al. The angelica Polysaccharide: a review of phytochemistry, pharmacology and beneficial effects on systemic diseases. Int Immunopharmacol. 2024;133:112025. doi:10.1016/j.intimp.2024.112025
181. Wei W, Zeng R, Gu C, et al. Angelica sinensis in China-A review of botanical profile, ethnopharmacology, phytochemistry and chemical analysis. J Ethnopharmacol. 2016;190:116–141. doi:10.1016/j.jep.2016.05.023
182. Luo F, Zeng F, Ren X, et al. Pheretima: a system review of zoology, traditional applications, processing technology, chemical composition, pharmacologic activities, quality control and toxicity. J Ethnopharmacol. 2025;351:120093. doi:10.1016/j.jep.2025.120093
183. Zhu Z, Deng X, Xie W, et al. Pharmacological effects of bioactive agents in earthworm extract: a comprehensive review. Anim Models Exp Med. 2024;7(5):653–672. doi:10.1002/ame2.12465
184. Wang Y, Wu L, Wang H, et al. Ligusticum chuanxiong: a chemical, pharmacological and clinical review. Front Pharmacol. 2025;16:1523176. doi:10.3389/fphar.2025.1523176
185. Liu Y, Wu H, Zhang Z, et al. Traditional use, phytochemistry, pharmacology, toxicology and clinical applications of persicae semen: a review. Chin J Integr Med. 2024;30(12):1137–1147. doi:10.1007/s11655-024-3815-4
186. Wang J, Wang L, Lou G, et al. Coptidis Rhizoma: a comprehensive review of its traditional uses, botany, phytochemistry, pharmacology and toxicology. Pharm Biol. 2019;57(1):193–225. doi:10.1080/13880209.2019.1577466
187. Kim S, Hur S, Yang D. The application of Panax notoginseng with a focus on its anti-inflammatory effect: a narrative review. Ann Translat Med. 2025;13(6):80. doi:10.21037/atm-25-69
188. Gao L, Li C, Wang Z, et al. Ligustri Lucidi Fructus as a traditional Chinese medicine: a review of its phytochemistry and pharmacology. Nat Product Res. 2015;29(6):493–510. doi:10.1080/14786419.2014.954114
189. Wang J, Tao H, Wang Z, et al. Application of network pharmacology, molecular docking, and in vitro experimental evaluation to decipher the anti-inflammatory mechanisms of Cirsium japonicum. Appl Sci. 2024;14:9687.
190. Aggarwal G, Kaur G, Bhardwaj G, et al. Traditional uses, phytochemical composition, pharmacological properties, and the biodiscovery potential of the Genus Cirsium. Chemistry. 2022;4:1161–1192.
191. Huang L, Lyu Q, Zheng W, et al. Traditional application and modern pharmacological research of Eucommia ulmoides Oliv. ChinMed. 2021;16(1):73. doi:10.1186/s13020-021-00482-7
192. Xing Y, He D, Wang Y, et al. Chemical constituents, biological functions and pharmacological effects for comprehensive utilization of Eucommia ulmoides Oliver. Food Sci Human Wellness. 2019;8(2):177–188.
193. Sun Y, Zheng J, Zhang T, et al. Review of polysaccharides from Citrus medica L. var. sarcodactylis. (Fingered citron): their extraction, purification, structural characteristics, bioactivity and potential applications. Int J Biol Macromol. 2024;282(Pt 1):136640. doi:10.1016/j.ijbiomac.2024.136640
194. Benedetto N, Carlucci V, Faraone I, et al. An insight into Citrus medica Linn.: a systematic review on phytochemical profile and biological activities. Plants. 2023;12(12):2267. doi:10.3390/plants12122267
195. Zhu B, Zhang Q, Hua J, et al. The traditional uses, phytochemistry, and pharmacology of Atractylodes macrocephala Koidz.: a review. J Ethnopharmacol. 2018;226:143–167. doi:10.1016/j.jep.2018.08.023
196. He X, Fang J, Huang L, et al. Sophora flavescens Ait.: traditional usage, phytochemistry and pharmacology of an important traditional Chinese medicine. J Ethnopharmacol. 2015;172:10–29. doi:10.1016/j.jep.2015.06.010
197. Shaukat MN, Nazir A, Fallico B. Ginger bioactives: a comprehensive review of health benefits and potential food applications. Antioxidants. 2023;12(11):2015. doi:10.3390/antiox12112015
198. Mukjerjee S, Karati D. A mechanistic view on phytochemistry, pharmacognostic properties, and pharmacological activities of phytocompounds present in Zingiber officinale: a comprehensive review. Pharmacolog Res Mod Chin Med. 2022;5:100173.
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
The Future Landscape of Endothelial Cells Research in Psoriasis: Bibliometric Analysis and Literature Review
Long SY, Shang L, Shi H, Zhao S, Cao J, He Y
Clinical, Cosmetic and Investigational Dermatology 2023, 16:3107-3120
Published Date: 30 October 2023
BAP31 Promotes Adhesion Between Endothelial Cells and Macrophages Through the NF-κB Signaling Pathway in Sepsis
He J, Jing D, Zhao S, Duan M
Journal of Inflammation Research 2024, 17:1267-1279
Published Date: 26 February 2024
Crosstalk Between H-Type Vascular Endothelial Cells and Macrophages: A Potential Regulator of Bone Homeostasis

Fan J, Xie Y, Liu D, Cui R, Zhang W, Shen M, Cao L
Journal of Inflammation Research 2025, 18:2743-2765
Published Date: 25 February 2025
Co-Targeting Biomimetic Nanoparticles Alleviate Atherosclerosis by Inhibiting the Vicious Circle Between Inflammation and Lipids
Wu C, Li Y, Liu Y, Wang X, Yuan P, Xu M, Deng Y, Zhang Z, Li C, Zhou X
International Journal of Nanomedicine 2025, 20:15705-15721
Published Date: 25 December 2025
Protective Effect of Chinese Herbal Medicine on Cerebral Ischemia-Reperfusion Injury by Regulating Ferroptosis
Luo S, Zhu J, Yang L, Liu J, Deng Y
Drug Design, Development and Therapy 2026, 20:563534
Published Date: 31 January 2026