Back to Journals » Journal of Inflammation Research » Volume 19
CD73 Blockade Aggravate Dextran-Sodium-Sulfate-Induced Colitis via Down Regulating Inosine
Authors Yang L, Wang X, Lin F, Liu C, Liu Y, Deng QC ![]() , Gao ZW
, Gao ZW
Received 6 November 2025
Accepted for publication 20 February 2026
Published 4 March 2026 Volume 2026:19 573140
DOI https://doi.org/10.2147/JIR.S573140
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Alberto Caminero
Lan Yang,1,* Xi Wang,1,* Fang Lin,1 Chong Liu,1 Yan Liu,1 Qing-chun Deng,2 Zhao-wei Gao1
1Department of Clinical Laboratory, Tangdu Hospital, Fourth Military Medical University, Xi’an, Shaanxi, 710038, People’s Republic of China; 2Department of Gynecology, The Second Affiliated Hospital of Hainan Medical University, Haikou, Hainan, 570102, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Zhao-wei Gao; Qing-chun Deng, Email [email protected]; [email protected]
Background: CD73, a pivotal enzyme in the purinergic signaling cascade, modulates the concentrations of adenosine and inosine. These metabolites are involved in immune responses and inflammatory processes. This study aims to investigate the function of CD73 in the pathogenesis of inflammatory bowel disease (IBD) and explore the potential mechanism.
Methods: Dextran-Sodium-Sulfate (DSS)-induced colitis mice models were established by orally administering 3% DSS. CD73 was blocked by intraperitoneal injection of Adenosine 5’- (α, β-methylene) diphosphate (APCP). Inosine was supplemented by intraperitoneal injection. Hematoxylin-eosin (H&E), PAS and Alcian blue staining were used to evaluate the inflammation infiltration and colon damage. Serum IL-6 levels were detected by ELISA assay. High-performance liquid chromatography-mass spectrometry (LC-MS) was used to investigate the level of inosine.
Results: Blockade of CD73 by APCP aggravated disease severity in DSS-induced colitis mice models, characterized by increased weight loss, colon shortening and pathological damage, increased disease activity and IL-6 production. Blocking CD73 impairs intestinal barrier function and integrity by reducing the expression of tight junction proteins (claudin-1, occludin and ZO-1), both in colon tissues and intestinal epithelial cell-MODE-K. In addition, APCP increased oxidative stress in colon tissue and MODE-K (increased MDA level, decreased SOD and GSH activities). Moreover, blocking CD73 reduced inosine levels in vivo and in vitro. We found that inosine treatment significantly ameliorated DSS-induced colitis in mice, as demonstrated by decreased weight loss, less colon shortening and histological injury, reduced disease activity and IL-6 production. Notably, the effects of inosine on MODE-K cells were opposite to those of APCP, including the effects on the expression of oxidative stress molecules and tight junction proteins.
Conclusion: This study indicates that CD73 exerts a protective effect in the progress of DSS-induced colitis. Inosine supplementation might be a potential therapeutic strategy for colitis.
Keywords: CD73, inosine, adenosine, inflammatory bowel disease, oxidative stress
Introduction
Inflammatory bowel disease (IBD) is a chronic intestinal inflammation with systemic manifestations, including ulcerative colitis (UC) and Crohn’s disease (CD).1 Recurrent diarrhea, rectal bleeding, abdominal pain and bowel obstruction are the main clinical manifestation.2,3 Periodic of symptomatic relapse and remission resulting in epithelial barrier disruption, ulceration formation and causing damage of epithelial structure.4 IBD has emerged as a public health challenge worldwide with rapidly increasing incidence.5,6 The pathogenesis of IBD were complex and remains unclear. Environmental factors, genetics, immune dysregulation, mucous barrier and epithelial barrier defects are believed to be association with IBD progress.7
CD73 is a key molecule in the purine signaling pathway, influencing the levels of adenosine and inosine, which play important roles in inflammation regulation. Wang R’s study showed that inhibition of CD73 alleviated intestinal inflammation by regulating macrophage differentiation in ulcerative colitis and significantly alleviated DSS-induced colitis in mice,8 which indicated the facilitating role of CD73 in colitis progression. However, there were other study showed the protective role of CD73 in the progression of colitis. Bynoe et al reported that CD73 deficient mice showed high susceptibility to DSS-induced colitis, along with significant weight loss, slower weight recovery, an increased intesnital permeability.9 Thus, the function of CD73 in colitis pathogenesis remains to be fully elucidated. Elevated oxidative stress levels and reduced antioxidant levels contribute to the pathogenesis of IBD.10 In addition, intestinal barrier dysfunction is a key factor in the initiation and progression of IBD. Tight junction proteins expressed by intestinal epithelial cells play a crucial role in maintaining intestinal barrier integrity.11,12 However, the effects of CD73 on oxidative stress and tight junction proteins has not been clearly defined.
In this study, to investigate the function, potential mechanism and therapeutic potency of CD73 in IBD, we systematically evaluated the effects of CD73 blockade on colitis severity, oxidative stress and tight junction proteins expression in DSS-induced colitis mice models. Given that CD73 regulates inosine levels, we also investigated the effect of inosine administration on DSS-induced colitis progression. In addition, we investigated the influences of CD73 inhibition and inosine treatment on intestinal epithelial cells. These findings will provide new insights into the role of CD73 and inosine in IBD development.
Materials and Methods
Materials
Dextran-Sodium-Sulfate (DSS) was purchased from GLPBIO (GC19829, USA). CD73 inhibitor-APCP was purchased from sigma (M3763, USA) and inosine was purchased from GLPBIO (GC17902, USA). ELISA kit for serum IL-6 detection was purchased from Novus (VAL604G, USA). Lipid Peroxidation MDA Assay Kit was purchased from Beyotime (S0131S, China). Total Superoxide Dismutase Assay Kit was purchased from Beyotime (S0101S, China). Total Glutathione Peroxidase Assay Kit was purchased from Beyotime (S0059S, China).
DSS-Induced Colitis Mice Models
DSS-induced colitis mice models were used in this study, which was a classical colitis animal model. Male C57BL/6J mice (6–8weeks) were purchased from experimental animal center, Fourth Military Medical University. All mice were housed in a standard environment (56–65% humidity, 24 ± 2 °C, 12 h light/dark cycle). The mice were randomly divided to four groups: control group (n = 8), DSS group (n = 8), APCP group (4mg/kg, n=8), and inosine group (300mg/kg, n = 8). Control group were provided with normal distilled water. DSS group were provided with distilled water containing 3% DSS (w/v) from day 1 to day 7. APCP group: APCP (4mg/kg) was administered via intraperitoneal (i.p.) injected into DSS mice once a day from day 3 to day 7. Inosine group: inosine (300 mg/kg i.p.) were injected into DSS mice once a day from day 3 to day 7. On day 8, mice were sacrificed with an overdose of isoflurane (RWD, R510-22-10), and then, blood and colon tissues were isolated according to the American Veterinary Medical Association. Colon length, body weight and the disease activity index (DAI) of mice were measured.
This animal study protocol was reviewed and approved by Fourth Military Medical University’s Institutional Animal Care and Use Committee (No.20250036). All the experiments were performed in accordance with ARRIVE guidelines and guidelines for the ethical review of laboratory animal welfare (GB/T35892-2018) issued by China.
Assessment of Disease Activity Index (DAI)
DAI score was used to evaluate the severity of colitis, including weight loss, diarrhea, and hematochezia. The DAI scores were monitored and recorded according to the previous study.13,14 Briefly, weight loss was scored as follows: score 0: none; score 1: 1–5%; score 2: 5–10%; score 3: 10–20%; score 4: >20%. Diarrhea was scored as follows: score 0: normal; score 2: loose stools; score 4: watery diarrhea. Hematochezia was scored as follows: score 0: normal; score 2: slight bleeding; score 4: gross bleeding. DAI score is the sum of the scores of weight loss, diarrhea, and hematochezia.
Histopathology Analysis
The colons were collected followed by washing with PBS and then fixed with 4% paraformaldehyde at room temperature overnight. Then, the paraffin-embedded sections of colon tissues were prepared (5μm thick). The H&E staining was performed to evaluate the inflammatory infiltration. Alcian blue and PAS staining were used to assess the number and structure integrity of goblet cell. Panoramic and Case Viewer 2.4 software (3DHISTECH, Hungary) were used for image acquisition and analysis. Subjective assessment was used in this study.
Immunohistochemical (IHC) Analysis
IHC analysis for paraffin-embedded sections of colon tissues were performed to detect the expression of CD73, F4/80, occludin, claudin-1 and ZO-1. Briefly, anti-occludin antibody (1:750 dilution; Servicebio, GB111401), anti-claudin-1 antibody (1:1000 dilution; Servicebio, GB15032), anti-ZO-1 antibody (1:1000 dilution; Servicebio, GB115686), anti-F4/80 antibody (1:1000 dilution; Servicebio, GB113373) and anti-CD73 antibody (1:200 dilution; abcam, ab175396) were used as the primary antibodies. The primary antibodies were incubated at 4°C for 12h. HRP-conjugated anti-rabbit secondary antibody (1:200 dilution; Servicebio, GB23303) was used as second antibody and incubated at room temperature for 1h. Panoramic and Case Viewer 2.4 software (3DHISTECH, Hungary) were used for image acquisition and analysis.
Serum Interleukin-6 (IL-6) Detection
The blood of mice was collected. After centrifugation (5000rpm, 4°C, 10min), the serum was collected. The serum IL-6 levels were detected using ELISA assay (VAL604G, Novus, USA) according to the instruction. First, 100µL serum (diluted 3 times with calibrator dilution) was added into well and incubated for 2h at room temperature; Second, 100µL detection antibody was added and incubated for 2h at room temperature; Third, 100µL streptavidin-HRP was added and incubated for 30min at room temperature; Fourth, 100µL 3,3′,5,5′-Tetramethylbenzidine substrate was added and incubate for 20min at room temperature; Finally, 50µL stop solution was added and OD450 was measured immediately.
Measurement the Activity of Antioxidant Enzyme
For MDA (S1031S, Beyotime, China) and GSH (S0059S, Beyotime, China) detection, the intestinal tissues were lysed by RIPA. The lysate was collected and used to MDA/GSH detection. For SOD detection, the intestinal tissues were lysed by SOD sample preparation solution from Total Superoxide Dismutase Assay Kit (S0101S, Beyotime, China). The supernate was separated for test. The levels of MDA, GSH and SOD were detected according to the operating instructions of kit.
Quantitative Real-Time PCR Analysis (qRT-PCR)
Total RNA from cells and colon histiocytic suspension were extracted by using Trizol reagent (TAKARA, Japan, Cat: 9109). The cDNA was generated by using Prime Script™ RT Master Mix (ACCUBATE BIOLOGY, China, AG11728). qRT-PCR reaction system was prepared using BlasTaqTM 2X qPCR Master Mix (abm, Canada, Cat: G891). The reaction was performed on Qiagen Amplifer (Germany, Rotor-gene QMDX 5). All operations was according to the manufacturer’s instructions. The mRNA expression levels in different groups were calculated using GAPDH as an internal standard. The primers sequences are listed in Table 1.
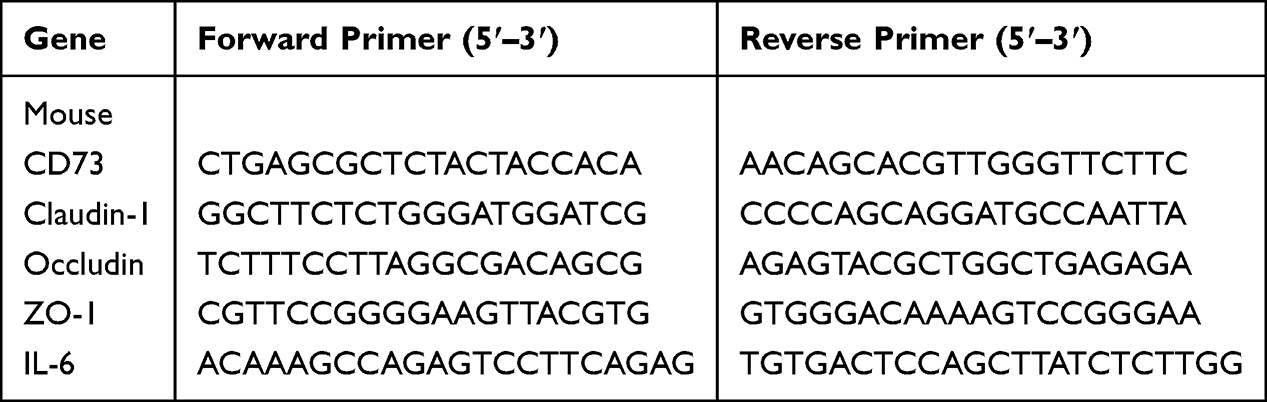 |
Table 1 Primer Sequence |
Cell Culture
The mouse intestinal epithelial cell line-MODE-K was cultured in Dulbecco’s modified Eagle’s medium (DMEM, Gibco, Cat: 12800-017) plus 10% fetal bovine serum (FBS, ExCell Bio, Cat: FSS500) at 37°C with 5% CO2 in a humidified incubator. The cells were treated with APCP (100µM) and inosine (3mM) for 48h and then incubated with LPS for 6h to induce inflammation.
Adenosine and Inosine Detection
High performance liquid chromatography mass spectrometry (LC-MS; Shimadzu, Japan) was employed to measure the levels of serum adenosine and inosine. Electrospray ionization was employed for detection in the positive ion mode. The multiple reaction monitoring mode (MRM) was utilized as the scanning mode. Transitions were monitored: for adenosine, from the m/z 268 precursor ion to the m/z 136 product ion (268 > 136); for inosine, 268.9>137.1; and for lamivudine (the internal standard) (Cat: 156082, TOPSCIENCE, Shanghai, China), 230 > 112. The analytical data were processed using the labsolution 5.81 (Shimadzu, Japan).
Statistical Analysis
All quantitative results were presented as mean ± standard deviations (SD). All data analyses were performed by using GraphPad prism 8.0. T-test was used to analyze the differences between two independent samples. One-way analysis of variance (ANOVA) test was used for the comparison of multiple groups. P<0.05 was considered to be statistically significant.
Results
CD73 Blockade Aggravated Disease Progression in DSS-Induced Colitis Mice Models
The experimental flowchart for mice experiment was presented in Figure 1A. Firstly, we detected the expression of CD73 in colon tissues from DSS-induced colitis mice by qRT-PCR and immunohistochemistry. We observed that the CD73 expression was significant decreased in colitis mice compared with control mice (Figure 1B and C).
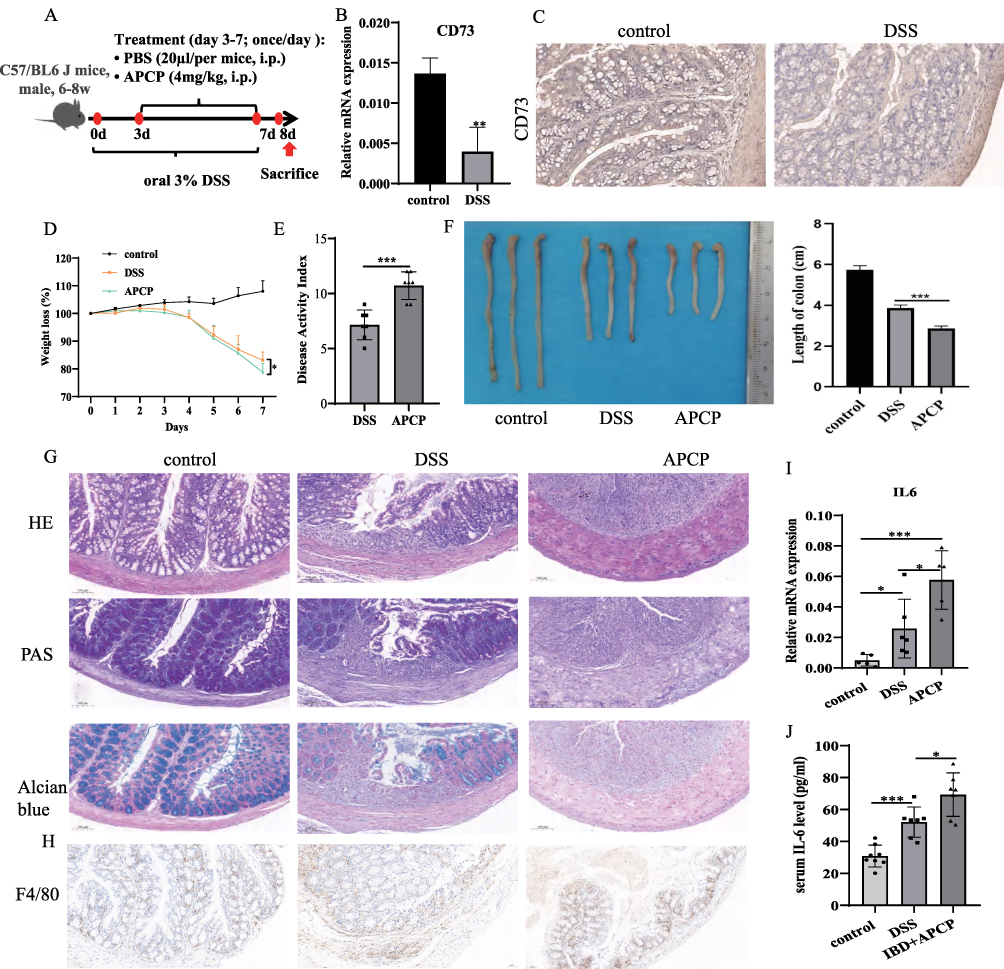 |
Figure 1 CD73 blockade aggravated disease progression in DSS-induced colitis mice. (A) low chart of animal experiments. (B and C) The expression levels of CD73 in colon tissues from control and DSS treated mice were analyzed by qRT-PCR and immunohistochemical (IHC) staining analysis. Scale bar, 50μm; magnification, 20x. (D–F) Body weight change, DAI and colon length in mice models. (G) The inflammation infiltration and colon damage of control, DSS and APCP treated group were detected by HE, PAS and Alcian blue staining. Scale bar, 100μm; magnification, 10x. (H) The infiltration of macrophage was measured by F4/80 staining. Scale bar, 50μm; magnification, 20x. (I and J) The expression levels of IL-6 in colon tissues from control, DSS and APCP treated mice were analyzed by qRT-PCR. *P < 0.05, ***P < 0.0005. |
To further investigate the role of CD73 in colitis, the effects of CD73 inhibition with APCP were examined in DSS-induced colitis mice. After 1 week intervention, mice in DSS group showed significant weight loss, which were further exacerbated by APCP treatment (Figure 1D). The DAI scores were higher in APCP treatment group t compared to the DSS group (Figure 1E). APCP treatment also led to more severe colon shortening (Figure 1F). Hematoxylin-eosin (H&E), PAS and Alcian blue staining were used to evaluate the inflammation infiltration and colon damage.
We observed that DSS induced the necrosis and exfoliation of mucosal epithelium, goblets cells depletion, tissue disorganization, and immune cells infiltration. APCP treatment could exacerbate the severity of tissue damage and inflammation cell infiltration (Figure 1G). IHC staining for F4/80 showed markedly increased macrophage infiltration following APCP treatment (Figure 1H). Notably, IL-6 levels were significant higher in APCP treated DSS-induced colitis mice compared to control and DSS induced mice (Control: 30.88 ± 6.841, DSS group: 52.13 ± 9.438, APCP group: 69.33 ±13.61pg/mL, Figure 1I and J).
APCP Inhibited the Colonic Antioxidant Abilities and Aggravated the Loss of Tight Junction Proteins in Colitis Mice
To assess the effect of APCP intervention on colonic oxidative stress in DSS treated mice, we measured colonic MDA levels along with SOD and GSH activities. Compared with the control group, the DSS-induced colitis mice showed elevated MDA levels, which were further increased following APCP treatment (Control: 4.199 ± 0.532, DSS group: 5.513 ± 0.825, APCP group: 9.553 ± 1.574 µM/mg, Figure 2A). Moreover, the activities of SOD (Control: 0.423 ± 0.019, DSS group: 0.320 ± 0.012, APCP group: 0.245 ± 0.033 U/mg) and GSH (Control: 15.110 ± 1.657, DSS group: 11.570 ± 1.842, APCP group: 9.365 ± 0.6951 mU/mg, Figure 2B and C) in DSS-induced colitis mice were reduced compared with control groups, which was the lowest in APCP treated mice (Figure 2B and C).
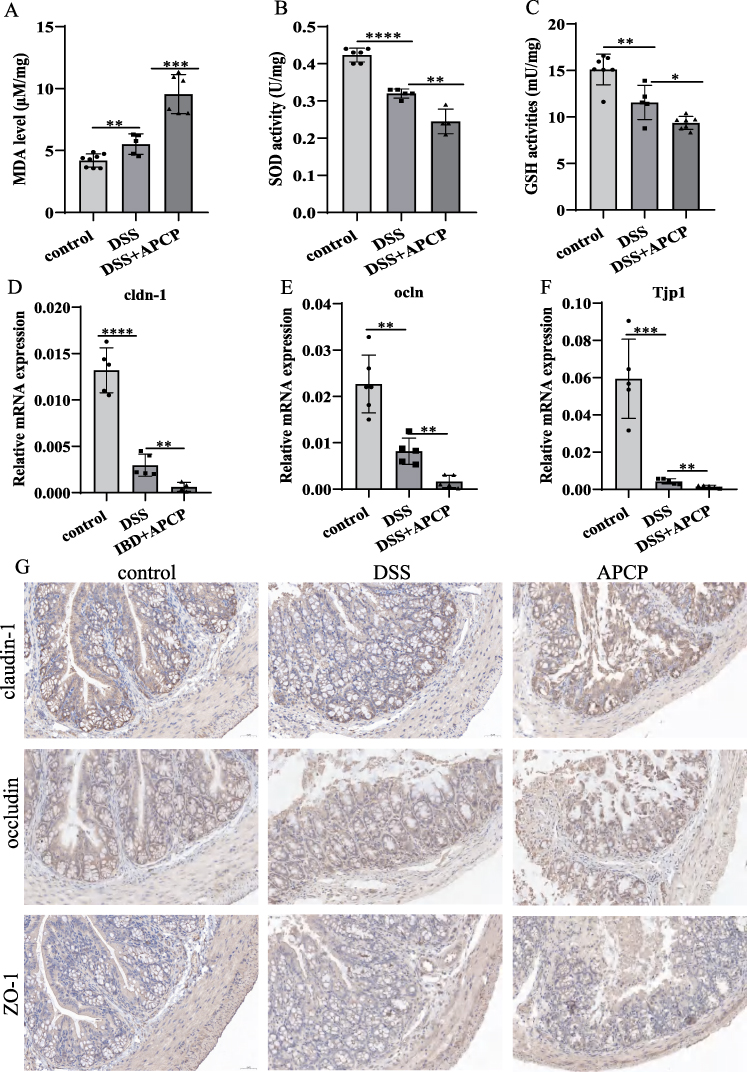 |
Figure 2 APCP inhibited the Colonic Antioxidant Abilities and aggravated the loss of tight junction proteins in colitis mice. (A–C): The effect of APCP intervention on the colonic oxidative stress in colon tissues. (D–G): The expression levels of tight junction proteins in colon tissues from control, DSS and APCP treated mice were analyzed by qRT-PCR and immunohistochemical (IHC) staining analysis. Scale bar, 50μm; magnification, 20x. *P < 0.05, **P < 0.01, ***P < 0.0005, ****P < 0.0001. |
To assess the integrity of intestinal epithelial structure and barrier function, we investigated the expression of tight junction proteins in colon tissues. We observed that DSS treatment could reduce the expression levels of claudin-1, occludin and ZO-1, which were more decreased in APCP treated mice (Figure 2D–F). Moreover, IHC analysis further confirmed the decreased protein expression of claudin-1, occludin and ZO-1 in APCP treated mice (Figure 2G). These finding suggested that the CD73 inhibition exacerbates disruption of intestinal epithelial structure integrity and barrier function.
Inosine Alleviated Disease Activity in DSS Induced Mice
The experimental flowchart for mice experiment was shown in Figure 3A. CD73 catalyzes the production of adenosine, which could be further degraded into inosine. Accumulating evidences indicates that both adenosine and inosine play significant roles in immunoregulation.15,16
 |
Figure 3 Inosine alleviated disease activity in DSS induced mice. (A) Flow chart of animal experiments. (B and C) Serum adenosine and inosine levels in control, DSS and APCP treated mice. (D–F) Body weight change, DAI and colon length in mice models. (G) The inflammation infiltration of control, DSS and inosine treated group were detected by HE staining. Scale bar, 100μm; magnification, 10x. (H) The infiltration of macrophage was measured by F4/80 staining. Scale bar, 50μm; magnification, 20x. (I and J) The expression levels of IL-6 in colon tissues from control, DSS and inosine treated mice were analyzed by qRT-PCR. *P < 0.05, **P < 0.01, ***P < 0.0005. |
LC-MS revealed decreased serum levels of adenosine (Control: 543.9 ± 101, DSS group: 323.3 ± 122, APCP group: 204.4 ± 111.4 pg/mL) and inosine (Control: 12.440 ± 3.879, DSS group: 9.074 ± 3.353, APCP group: 5.987 ± 2.465 pg/mL) in DSS-induced colitis mice, which were more reduced in APCP treated DSS-induced colitis mice (Figure 3B and C). In this study, we further investigated the effect of inosine on colitis mice. Compared with DSS induced mice, we observed that inosine i.p. significantly alleviated the extent of weight loss, reduced the DAI scores, and ameliorated colon shortening (Figure 3D–F). H&E staining showed that inosine i.p. could ameliorate the severity of colon damage and immune cell infiltration (Figure 3G). Moreover, we observed a significant decrease in macrophages infiltration in inosine treated DSS-induced colitis mice (Figure 3H). Additionally, IL-6 levels in serum and colon tissues were significantly increased by DSS treatment, while were reduced by inosine treatment (Control: 30.880 ± 6.841, DSS group: 52.13 ± 9.438, Inosine group: 36.070 ± 7.039 pg/mL, Figure 3I and J).
Inosine Promoted the Colonic Antioxidant Abilities and Prevented the Loss of Tight Junction Proteins in Colitis
The effects of inosine i.p. on oxidative stress and tight junction proteins in colon tissues were investigated. Compared with the control group, the DSS-induced colitis mice exhibited elevated MDA levels, which were significantly reduced by inosine treatment (Control: 4.199 ± 0.532, DSS group: 5.513 ± 0.825, Inosine group: 4.473 ± 0.464 µM/mg, Figure 4A). Additionally, SOD and GSH activities were significantly decreased in the DSS group, whereas inosine i.p. significantly elevated the activities of SOD (Control: 0.423 ± 0.019, DSS group: 0.320 ± 0.012, Inosine group: 0.383 ± 0.021 U/mg) and GSH (Control: 15.110 ± 1.657, DSS group: 11.570 ± 1.842, Inosine group: 13.690 ± 0.552 mU/mg, Figure 4B and C) in colon tissues. Furthermore, inosine treatment significantly elevated the expression of claudin-1, occludin and ZO-1 (Figure 4D–G).
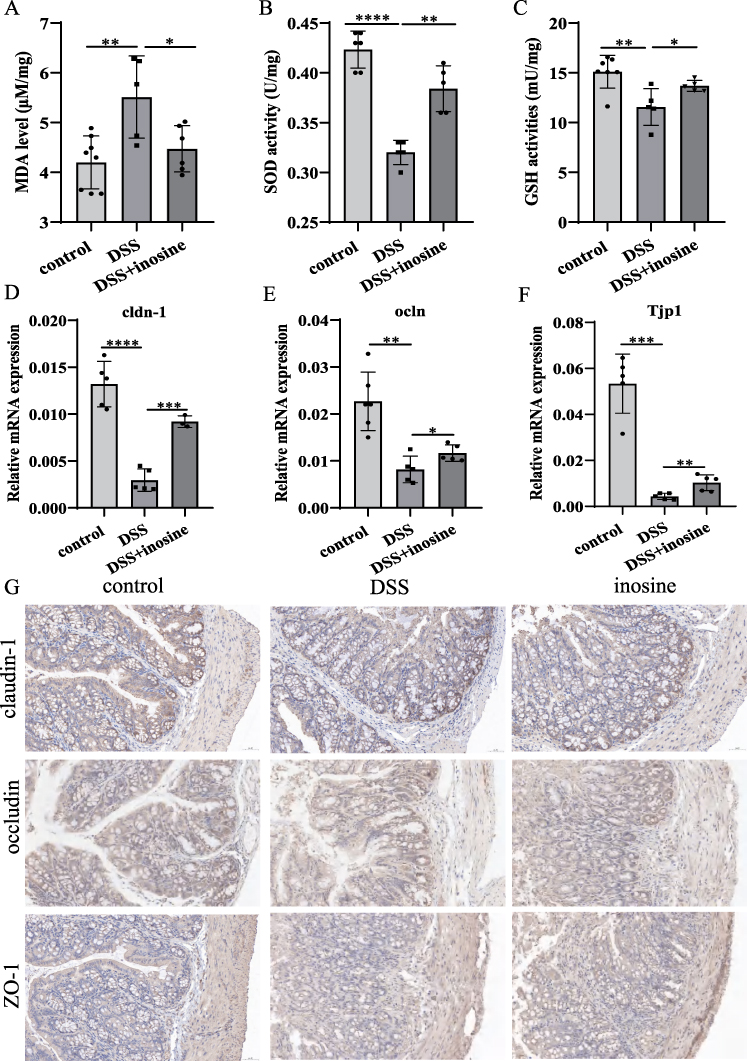 |
Figure 4 Inosine promoted the Colonic Antioxidant Abilities and prevented the loss of tight junction proteins in colitis. (A–C) The effect of inosine intervention on the colonic oxidative stress in colon tissues. (D–G) The expression levels of tight junction proteins in colon tissues from control, DSS and inosine treated mice were analyzed by qRT-PCR and immunohistochemical (IHC) staining analysis. Scale bar, 50μm; magnification, 20x. *P < 0.05, **P < 0.01, ***P < 0.0005, ****P < 0.0001. |
The Opposite Effects of APCP and Inosine on Intestinal Epithelial Cell
In MODE-K cells, we observed that APCP treatment reduced inosine production (Control: 86.920 ± 3.839, APCP group: 59.5 ± 2.465 ng/mL, Figure 5A). LPS stimulation significantly increased IL-6 expression, which were further enhanced by APCP treatment. In contrast, inosine suppressed IL-6 expression (Figure 5B and C). Antioxidant capacity assessment revealed that APCP treatment markedly increased MDA levels, whereas inosine treatment attenuated this effect (Control: 64.220 ± 3.033, DSS group: 69.740 ± 1.162, Inosine group: 51.010 ± 1.389 µM/mg, Figure 5D). Moreover, after APCP treatment, SOD and GSH activities were markedly reduced. However, inosine treatment significantly enhanced the activities of SOD (Control: 0.420 ± 0.044, DSS group: 0.296 ± 0.041, Inosine group: 0.565 ± 0.055 U/mg) and GSH (Control: 303.2 ± 12.2, DSS group: 209.8 ± 7.659, Inosine group: 374.9 ± 14.980 U/mg, Figure 5E and F). The expression levels of claudin-1, occludin, and ZO-1 were significantly reduced by APCP treatment, whereas inosine treatment could restore their expression. (Figure 5G–I). Taken together, the impacts of APCP and inosine on MODE-K were completely opposite, consistent with the results from mice experiments.
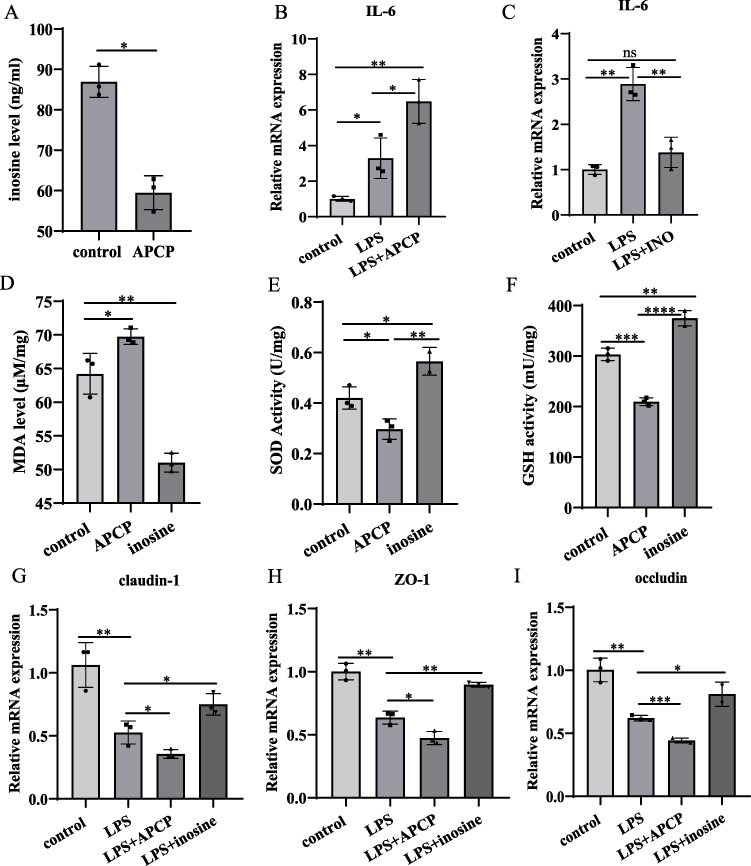 |
Figure 5 The opposite effects of APCP and inosine on intestinal epithelial cell. (A): Serum inosine levels in APCP treated MODE-K cells. (B and C) The expression levels of IL-6 in control, LPS, APCP and inosine treated MODE-K cells were analyzed by qRT-PCR. (D–F): The effect of APCP and inosine intervention on the oxidative stress in MODE-K cells. (G–I): The expression levels of tight junction proteins in control, LPS APCP and inosine treated MODE-K cells were analyzed by qRT-PCR. *P < 0.05, **P < 0.01, ***P < 0.0005, ****P < 0.0001. |
Discussion
Inflammatory bowel disease (IBD) is characterized by intestinal inflammation and is associated with structural disruption of the intestinal epithelium.17 Immune disorder and inflammation are implicated in the whole process of IBD. CD73 plays a critical role in immune regulation. In this study, we first demonstrated that blocking CD73 could exacerbate the disease activity in DSS-induced colitis mice, as indicated by increased DAI scores, reduced colon length, more severe intestinal pathological, and elevated IL-6 levels.
Similarly, Zhu’s study indicated that Moxibustion could relieve gut inflammation in DSS-induced mice by enhancing the expression of CD39 and CD73.18 Notably, Shao and colleague’s study demonstrated that knocking out CD73 diminishes the therapeutic efficacy of mesenchymal-like endometrial regenerative cells in treating colitis.19 However, in contrast to our study, Wang et al’s study showed that CD73 blockade markedly alleviated DSS-induced colitis in mice, as characterized by reduced weight loss, diarrhea and bloody stool.8 Moreover, Liu et al’s study showed that CD73 blockade suppressed the severity of colitis-associated tumorigenesis with attenuated weight loss, longer colons, lower tumor number and smaller tumor size compared to the model group.20
Secondly, we found that CD73 blockade significantly reduced adenosine and inosine levels in vivo. Thus, we assumed that adenosine and inosine might be involved in IBD progress. Inosine injection into DSS-mice was used to evaluate the effect of inosine on IBD. As expected, inosine significantly alleviated DSS-induced colitis, as characterized by lower DAI score, longer bowel length, milder intestinal pathology and lower inflammatory cytokines levels. Supportingly, Zhu et al’s study demonstrated inosine treatment could improve gut ecosystem and intestinal barrier function.21 Guo et al’s study showed gut microbiota-derived inosine supplementation attenuates colitis through NF-κB and Nrf2 signaling.22 Notably, although different studies had disagreements about the role of CD73 in colitis, which could be due to the functional diversity of CD73 itself; however, the alleviated effects of inosine on IBD were consistent among different studies. To date, several therapeutic approaches, including anti-TNF-α, corticosteroids, amino salicylates, and antibiotics, have been employed to alleviate IBD. However, long-term use of these medications may give rise to a series of side effects, such as high blood pressure, headaches, diabetes, liver damage, and nausea.22,23 Inosine is commonly employed as a hepatoprotective and anti-inflammatory agent. Our study indicated that inosine could be a potential therapeutic option for colitis, which needed to be verified by clinical trial in IBD patients. Notably, considering that the lack of a sustained delivery system for adenosine, which led to a high lethality rate in our preliminary experiment, we did not examine the effect of adenosine in DSS-induced mice.
Thirdly, we explored the impact of CD73 blockade and inosine on oxidative stress, both in vivo and in vitro. Oxidative stress is a crucial pathophysiological factor during IBD development. An overabundance of oxidative stress can undermine the integrity of intestinal barrier.24 MDA can influence cell mitosis by damaging the DNA structure,25 which has been regarded as an indicator of oxidative stress and involved in IBD progress.26 Sahoo’s research revealed that the colonic MDA level was increased in IBD.10 In this study, we found that CD73 blockade led to an increase in MDA. The antioxidant enzymes have the ability to reduce oxidative stress, thereby protecting intestinal cells from oxidative damage.22 Superoxide dismutase (SOD) could catalyze the conversion of superoxide radicals into oxygen and hydrogen peroxide, which could reduce oxidative damage.27 GSH can protect cells from free radicals and pro-oxidant damage.10 Zhang et al’s research found the SOD activity was significantly decreased in DSS-induced colitis mice.28 Grabias’s study discovered that enhancing GSH activity can decrease oxidative stress in the colonic mucosa.29 We observed CD73 blockade significantly reduced SOD and GSH activities. Antioxidants have been proposed as a potential alternative therapy of anti-inflammatory/immunomodulatory drugs for patients with uncomplicated gastrointestinal disorders.30 In this study, we discovered that inosine could alleviate oxidative stress, through down-regulating MDA and up-regulating SOD and GSH activities.
Fourthly, intact intestinal and mucosal barriers are critical in maintaining colon homeostasis.31 Tight junction proteins are important for intestinal tract barrier.12,32 Decreased tight junction proteins lead to a reduction in the firmness of the intestinal barrier and contribute to colitis development.11 In this study, we explore the impact of CD73 blockade and inosine on claudin-1, occludin, and ZO-1. We observed that APCP treatment decreased claudin-1, occludin, and ZO-1, in both colon tissues and intestinal epithelial cells. Claudin-1 is important for maintaining the integrity of epithelial tight junctions and mainly regulates intestinal permeability.33 Decreased Claudin-1 lead to the hyperpermeability of intestinal epithelial cells.34 Severe intestinal leakage was observed in mice with intestinal epithelial-specific knockout of the occludin,35,36 while the overexpression of Ocln promotes the epithelial barrier function in vitro.37 Moreover, ZO-1 regulates epithelial proliferation and mucosal repair. ZO-1 deficiency disturbed mitotic spindle orientation induced abortive proliferation in intestinal epithelial cells.12 In this study, we observed that inosine can boost the expression of tight junction proteins. Thus, these effects of inosine regulate oxidative stress and tight junction proteins could explain the alleviative effects of inosine on IBD disease. Notably, there were other biomarkers for assessing the structural integrity of the intestinal epithelium, including desmosomes, adherens junctions and cytoskeleton.38 Except for these, Hempstock’s et al also evaluated the paracellular permeability of ions by using Ussing chamber technique.39 The effect of inosine on structural integrity can be tested by using the above method in further study.
In summary, this study explored the function and mechanism of CD73 in IBD, thereby offering a novel theoretical foundation and potential therapeutic strategy for IBD. Nevertheless, this study has certain limitations. Firstly, the intervention of inosine’s effects on IBD disease was performed only in mice model. The potential therapeutic action of inosine on IBD patients needed to be verified by clinical trial in further study. Secondly, gut microbiota plays important roles during IBD progress. Gut bacteria constitute a vital niche within the gastrointestinal tract and play an essential role in maintaining intestinal microecological balance. Compared with healthy individuals, IBD patients exhibit reduced bacterial diversity, diminished levels of anti-inflammatory bacteria, and an expansion of pro-inflammatory bacterial species.40 However, the influences of CD73 blockade and inosine were not investigated in this study, which needed to be explored in further study.
Conclusion
In conclusion, our studies revealed that CD73 played a significant role in the progress of DSS-induced colitis via regulating the level of inosine. Inosine could be a potential drug for IBD therapy, which need further verification by clinical trial. Moreover, the alleviative efficacy of inosine on DSS-induced colitis associated with decreased oxidative stress levels, elevated antioxidants levels and tight junction.
Data Sharing Statement
All data are available through the corresponding author.
Ethics Statement
This animal study protocol was reviewed and approved by Fourth Military Medical University’s Institutional Animal Care and Use Committee (No.20250036). All the experiments were performed and analysed in accordance with ARRIVE guidelines and guidelines for the ethical review of laboratory animal welfare (GB/T35892-2018) issued by China.
Author Contributions
All authors made a significant contribution to the work reported. Lan Yang: Conception and written the article; Xi Wang: Acquisition of data and written the article; Fang Lin: Execution and draft the article; Chong Liu: Analysis, interpretation and draft the article; Yan Liu: acquisition of data and draft the article; Qing-chun Deng: Analysis, interpretation, written the article and substantially revised the article; Zhao-wei Gao: Study design and substantially revised the article. All of the authors agreed on the Journal of inflammation research to which the article has been submitted. All the authors reviewed and agreed on all versions of the article before submission, during revision, the final version accepted for publication, and any significant changes introduced at the proofing stage. All of the authors agree to take responsibility and be accountable for the contents of the article.
Funding
This research was funded by the Talent Fund of Tangdu Hospital (No. 2021SHRC004).
Disclosure
The authors declare that they have no conflict of interest.
References
1. Faye AS, Colombel J-FJ-F. Aging and IBD: a New Challenge for Clinicians and Researchers. Inflamm Bowel Dis. 2022;28(1):126–13. doi:10.1093/ibd/izab039
2. Zeng B, Huang Y, Chen S, et al. He X. Dextran sodium sulfate potentiates NLRP3 inflammasome activation by modulating the KCa3.1 potassium channel in a mouse model of colitis. Cell Mol Immunol. 2022;19(8):925–943. doi:10.1038/s41423-022-00891-0
3. Ng SC, Shi HY, Hamidi N, et al. Worldwide incidence and prevalence of inflammatory bowel disease in the 21st century: a systematic review of population-based studies. Lancet. 2017;390(10114):2769–2778. doi:10.1016/S0140-6736(17)32448-0
4. Yang L, Wu G, Wu Q, Peng L, Yuan L. METTL3 overexpression aggravates LPS-induced cellular inflammation in mouse intestinal epithelial cells and DSS-induced IBD in mice. Cell Death Discov. 2022;8(1):62. doi:10.1038/s41420-022-00849-1
5. Kaplan GG. The global burden of IBD: from 2015 to 2025. Nat Rev Gastroenterol Hepatol. 2015;12(12):720–727. doi:10.1038/nrgastro.2015.150
6. Kaulmann A, Bohn T. Bioactivity of Polyphenols: preventive and Adjuvant Strategies toward Reducing Inflammatory Bowel Diseases-Promises, Perspectives, and Pitfalls. Oxid Med Cell Longev. 2016;2016:9346470. doi:10.1155/2016/9346470
7. Wu H, Chen QY, Wang WZ, et al. Compound sophorae decoction enhances intestinal barrier function of dextran sodium sulfate induced colitis via regulating notch signaling pathway in mice. Biomed Pharmacother. 2021;133:110937. doi:10.1016/j.biopha.2020.110937
8. Wang R, Wang Y, Wu C, et al. CD73 blockade alleviates intestinal inflammatory responses by regulating macrophage differentiation in ulcerative colitis. Exp Ther Med. 2023;25(6):272. doi:10.3892/etm.2023.11972
9. Bynoe MS, Waickman AT, Mahamed DA, Mueller C, Mills JH, Czopik A. CD73 is critical for the resolution of murine colonic inflammation. J Biomed Biotechnol. 2012;2012:260983. doi:10.1155/2012/260983
10. Sahoo DK, Heilmann RM, Paital B, et al. Oxidative stress, hormones, and effects of natural antioxidants on intestinal inflammation in inflammatory bowel disease. Front Endocrinol (Lausanne). 2023;14:1217165. doi:10.3389/fendo.2023.1217165
11. Cavalu S, Sharaf H, Saber S, et al. Ambroxol, a mucolytic agent, boosts HO-1, suppresses NF-κB, and decreases the susceptibility of the inflamed rat colon to apoptosis: a new treatment option for treating ulcerative colitis. FASEB J. 2022;36(9):e22496. doi:10.1096/fj.202200749R
12. Kuo WT, Zuo L, Odenwald MA, et al. The Tight Junction Protein ZO-1 Is Dispensable for Barrier Function but Critical for Effective Mucosal Repair. Gastroenterology. 2021;161(6):1924–1939. doi:10.1053/j.gastro.2021.08.047
13. Chen X, Liu G, Yuan Y, Wu G, Wang S, Yuan L. NEK7 interacts with NLRP3 to modulate the pyroptosis in inflammatory bowel disease via NF-κB signaling. Cell Death Dis. 2019;10(12):906. doi:10.1038/s41419-019-2157-1
14. Wu G, Zhang D, Yang L, Wu Q, Yuan L. MicroRNA-200c-5p targets NIMA Related Kinase 7 (NEK7) to inhibit NOD-like receptor 3 (NLRP3) inflammasome activation, MODE-K cell pyroptosis, and inflammatory bowel disease in mice. Mol Immunol. 2022;146:57–68. doi:10.1016/j.molimm.2022.03.121
15. Vuerich M, Nguyen DH, Ferrari D, Longhi MS. Adenosine-mediated immune responses in inflammatory bowel disease. Front Cell Dev Biol. 2024;12:1429736. doi:10.3389/fcell.2024.1429736
16. Klysz DD, Fowler C, Malipatlolla M, et al. Inosine Induces Stemness Features in CAR T cells and Enhances Potency. bioRxiv. 2023;4:537859. doi:10.1101/2023.04.21.537859. Update in: Cancer Cell. 2024 Feb 12;42(2):266-282.e8. doi: 10.1016/j.ccell.2024.01.002
17. Li MX, Li MY, Lei JX, et al. Huangqin decoction ameliorates DSS-induced ulcerative colitis: role of gut microbiota and amino acid metabolism, mTOR pathway and intestinal epithelial barrier. Phytomedicine. 2022;100:154052. doi:10.1016/j.phymed.2022.154052
18. Zhu Y, Zhuang Z, Wu Q, et al. CD39/CD73/A2a Adenosine Metabolic Pathway: targets for Moxibustion in Treating DSS-Induced Ulcerative Colitis. Am J Chin Med. 2021;49(3):661–676. doi:10.1142/S0192415X21500300
19. Shao B, Ren SH, Wang ZB, et al. CD73 mediated host purinergic metabolism in intestine contributes to the therapeutic efficacy of a novel mesenchymal-like endometrial regenerative cells against experimental colitis. Front Immunol. 2023;14:1155090. doi:10.3389/fimmu.2023.1155090
20. Liu XH, Wu XR, Lan N, et al. CD73 promotes colitis-associated tumorigenesis in mice. Oncol Lett. 2020;20(2):1221–1230. doi:10.3892/ol.2020.11670
21. Zhu Y, Wang X, Zhu L, et al. Lactobacillus rhamnosus GG combined with inosine ameliorates alcohol-induced liver injury through regulation of intestinal barrier and Treg/Th1 cells. Toxicol Appl Pharmacol. 2022;439:115923. doi:10.1016/j.taap.2022.115923
22. Guo W, Tang X, Zhang Q, et al. Mitigation of Dextran-Sodium-Sulfate-Induced Colitis in Mice through Oral Administration of Microbiome-Derived Inosine and Its Underlying Mechanisms. Int J Mol Sci. 2023;24(18):13852. doi:10.3390/ijms241813852
23. Singh S, Fumery M, Sandborn WJ, Murad MH. Systematic review with network meta-analysis: first- and second-line pharmacotherapy for moderate-severe ulcerative colitis. Aliment Pharmacol Ther. 2018;47(2):162–175. doi:10.1111/apt.14422
24. Tang Z, Yang Y, Wu Z, Ji Y. Heat Stress-Induced Intestinal Barrier Impairment: current Insights into the Aspects of Oxidative Stress and Endoplasmic Reticulum Stress. J Agric Food Chem. 2023;71(14):5438–5449. doi:10.1021/acs.jafc.3c00798
25. Guo W, Cui S, Tang X, et al. Intestinal Microbiomics and Metabolomics Insights into the Hepatoprotective Effects of Lactobacillus paracasei CCFM1222 Against the Acute Liver Injury in Mice. Probiotics Antimicrob Proteins. 2023;15(5):1063–1077. doi:10.1007/s12602-022-09986-6
26. Xu L, Zeng X, Liu Y, Wu Z, Zheng X, Zhang X. Inhibitory effect of Dendrobium officinale polysaccharide on oxidative damage of glial cells in aging mice by regulating gut microbiota. Int J Biol Macromol. 2023;247:125787. doi:10.1016/j.ijbiomac.2023.125787
27. Zhou J, Li M, Chen Q, et al. Programmable probiotics modulate inflammation and gut microbiota for inflammatory bowel disease treatment after effective oral delivery. Nat Commun. 2022;13(1):3432. doi:10.1038/s41467-022-31171-0
28. Zhang X, Yuan Z, Wu J, et al. An Orally-Administered Nanotherapeutics with Carbon Monoxide Supplying for Inflammatory Bowel Disease Therapy by Scavenging Oxidative Stress and Restoring Gut Immune Homeostasis. ACS Nano. 2023;17(21):21116–21133. doi:10.1021/acsnano.3c04819
29. Dziąbowska-Grabias K, Sztanke M, Zając P, et al. Antioxidant Therapy in Inflammatory Bowel Diseases. Antioxidants. 2021;10(3):412. doi:10.3390/antiox10030412
30. Candellone A, Cerquetella M, Girolami F, Badino P, Odore R. Acute Diarrhea in Dogs: current Management and Potential Role of Dietary Polyphenols Supplementation. Antioxidants. 2020;9(8):725. doi:10.3390/antiox9080725
31. Wu J, Xu X, Duan J, et al. EFHD2 suppresses intestinal inflammation by blocking intestinal epithelial cell TNFR1 internalization and cell death. Nat Commun. 2024;15(1):1282. doi:10.1038/s41467-024-45539-x
32. Jiang Y, Song J, Xu Y, et al. Piezo1 regulates intestinal epithelial function by affecting the tight junction protein claudin-1 via the ROCK pathway. Life Sci. 2021;275:119254. doi:10.1016/j.lfs.2021.119254
33. Safari F, Sharifi M, Talebi A, Mehranfard N, Ghasemi M. Alleviation of cholestatic liver injury and intestinal permeability by lubiprostone treatment in bile duct ligated rats: role of intestinal FXR and tight junction proteins claudin-1, claudin-2, and occludin. Naunyn Schmiedebergs Arch Pharmacol. 2023;396(9):2009–2022. doi:10.1007/s00210-023-02455-z
34. Lan H, Zhang LY, He W, et al. Sinapic Acid Alleviated Inflammation-Induced Intestinal Epithelial Barrier Dysfunction in Lipopolysaccharide- (LPS-) Treated Caco-2 Cells. Mediators Inflamm. 2021;2021:5514075. doi:10.1155/2021/5514075
35. Kuo WT, Odenwald MA, Turner JR, Zuo L. Tight junction proteins occludin and ZO-1 as regulators of epithelial proliferation and survival. Ann N Y Acad Sci. 2022;1514(1):21–33. doi:10.1111/nyas.14798
36. Chanez-Paredes SD, Abtahi S, Kuo WT, Turner JR. Differentiating Between Tight Junction-Dependent and Tight Junction-Independent Intestinal Barrier Loss In Vivo. Methods Mol Biol. 2021;2367:249–271. doi:10.1007/7651_2021_389. Erratum in: Methods Mol Biol. 2021;2367:C1. doi: 10.1007/978-1-0716-1673-4_391.
37. McCarthy KM, Skare IB, Stankewich MC, et al. Occludin is a functional component of the tight junction. J Cell Sci. 1996;109(Pt 9):2287–2298. doi:10.1242/jcs.109.9.2287
38. Groschwitz KR, Hogan SP. Intestinal barrier function: molecular regulation and disease pathogenesis. J Allergy Clin Immunol. 2009;124(1):3–20. doi:10.1016/j.jaci.2009.05.038
39. Hempstock W, Ishizuka N, Hayashi H. Functional Assessment of Intestinal Tight Junction Barrier and Ion Permeability in Native Tissue by Ussing Chamber Technique. J Vis Exp. 2021;171. doi:10.3791/62468.
40. Li Q, Zhou S, Wang Y, Cong J. Changes of intestinal microbiota and microbiota-based treatments in IBD. Arch Microbiol. 2022;204(7):442. doi:10.1007/s00203-022-03069-4
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Hydrogen Regulates Ulcerative Colitis by Affecting the Intestinal Redox Environment

Li J, Huang G, Wang J, Wang S, Yu Y
Journal of Inflammation Research 2024, 17:933-945
Published Date: 12 February 2024
Altered CD73-Adenosine Signaling Linked to Infection in Patients undergoing hemodialysis

Xiang F, Zhang Z, Nie Y, Cao X, Li Y, Chen X, Gong S, Zou J, Teng J, Ding X, Shen B
Journal of Inflammation Research 2025, 18:1267-1279
Published Date: 27 January 2025
T Cell-Derived Apoptotic Extracellular Vesicles Ameliorate Bone Loss via CD39 and CD73-Mediated ATP Hydrolysis

Yang X, Zhou Y, Zhou F, Bao L, Wang Z, Li Z, Ding F, Kuang H, Liu H, Tan S, Qiu X, Jing H, Liu S, Ma D
International Journal of Nanomedicine 2025, 20:1083-1100
Published Date: 27 January 2025
Scutellaria baicalensis-Derived Extracellular Vesicles Alleviate Inflammatory Bowel Disease by Inhibiting the NF-κB/NLRP3 Pathway
Miao R, Wang S, Yin H, Zhu R, Yin Y, Liao W, Wang S, Zhang J, Li R, Xu J
International Journal of Nanomedicine 2026, 21:586382
Published Date: 27 April 2026