Back to Journals » OncoTargets and Therapy » Volume 16
CCDC103 as a Prognostic Biomarker Correlated with Tumor Progression and Immune Infiltration in Glioma
Authors Xu Z, Xu H, Chen X, Huang X, Tian J, Zhao J, Liu B, Shi F, Wu J, Pu J
Received 27 July 2023
Accepted for publication 3 October 2023
Published 18 October 2023 Volume 2023:16 Pages 819—837
DOI https://doi.org/10.2147/OTT.S429958
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Dr John Maher
Zhixing Xu,1,2,* Haitao Xu,1,* Xi Chen,1,* Xiaobing Huang,1 Jintao Tian,1 Jinxi Zhao,1 Bohu Liu,1 Fengcai Shi,2 Jin Wu,2 Jun Pu1
1Department of Neurosurgery, The Second Affiliated Hospital of Kunming Medical University, Kunming, 650223, People’s Republic of China; 2Department of Neurosurgery, The Pu’er People’s Hospital, Pu’er, 665000, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Jin Wu; Jun Pu, Email [email protected]; [email protected]
Background: The Coiled-coil domain-containing proteins (CCDCs) are expressed in many cancers, but the role of Coiled-coil domain-containing protein 103 (CCDC103) in cancers remains unclear. Further investigations are necessary to ascertain its diagnostic significance and understand its biological function in cancers. This study aims to elucidate the biological functionalities of CCDC103 in glioma and evaluate the correlation between CCDC103 expression with glioma progression.
Methods: Clinical data on glioma patients were acquired from The Cancer Genome Atlas (TCGA), the Chinese Glioma Genome Atlas (CGGA), and the Gene Expression Omnibus (GEO). The evaluation encompassed the examination of correlations between CCDC103 expression, pathological characteristics, and clinical outcomes. Furthermore, the analysis included the assessment of the correlations between CCDC103 expression and immune cell infiltration as well as glioma progression.
Results: Gliomas have higher levels of CCDC103 expression than the para-carcinoma tissues. Poorer prognosis, unfavorable histological characteristics, the absence of IDH gene mutations, and the absence of chromosome 1p and 19q deletions were all associated with higher expression of CCDC103 in gliomas. In addition to patient age, tumor grade, the absence of IDH mutations, and the absence of chromosome 1p and 19q deletions, univariate and multivariate Cox analyses showed that CCDC103 expression was independently prognostic of overall survival, disease-free survival, and progression-free survival in patients with glioma. Furthermore, tumor infiltration of B cells, neutrophils, macrophages, and dendritic cells were all linked with elevated expression of CCDC103. High CCDC103 expression was linked to immune response-related signaling pathways and cell proliferation, according to gene set enrichment analysis (GSEA). Notably, the knockdown of CCDC103 in glioma cell lines resulted in a significant reduction in cell proliferation and migration.
Conclusion: The correlation between CCDC103 expression and both glioma progression and immune cell infiltration implies that CCDC103 expression holds promise as a valuable prognostic biomarker for glioma.
Keywords: glioma, CCDC103, prognosis, biomarker, immune infiltration
Introduction
Gliomas originate from brain glial cells. It is the most common primary intracranial tumor, accounting for about 81% of malignant tumors in the central nervous system.1 Temozolomide combined with radiotherapy and chemotherapy is the standard treatment after surgery. However, the median survival time of patients is still not more than 2 years.2 The discovery of functional lymphatic vessels within the dural sinuses unveils fresh evidence supporting active immune surveillance in the central nervous system (CNS). Novel therapeutic avenues for glioma treatment encompass immune checkpoint inhibitors (ICIs), viral therapy, CAR-T therapy, and vaccine treatment.3 However, the present immunotherapy options for glioma exhibit inadequate therapeutic efficacy due to the intricate immunosuppressive environment and systemic immunosuppression prevalent in gliomas. Hence, the quest for novel immune-related indicators and therapeutic targets remains pivotal in fostering advancements in glioma treatment.
Recent research has demonstrated that tumor cells may adapt their actin cytoskeleton in order to respond to a wide range of complicated mechanical, biochemical, and microenvironmental cues.4 For instance, cells can influence cell proliferation, control changes in cell shape and the structure of the actin skeleton through integrins, and attach to certain extracellular matrix (ECM) proteins.5 Similar to this, the development of F-actin stress fibers and actin cytoskeletal polymers, which are better able to respond to changes in the extracellular matrix, are crucial for the advancement of the cell cycle.6,7 Actin/globin and microtubules are both involved in the nuclear migration of glioma cells, according to studies.8 The formation of dynamin-cortactin complexes and the phosphorylation of cortactin by cyclin-dependent kinase 5 (CDK5), among others, control actin bundles, enabling glioma-derived cells to form filamentous uropods and flaky caudate, which further affect tumor cell invasion, migration, and metastasis.9 Similar to how actin serves as the essential link in the cytoskeleton of microtubules, which are necessary for long-distance glioma cell communication, cytoplasmic dynein inhibition lowers the activity of glioma cells and prevents their proliferation.10,11
The proteins containing coiled-coil domains (CCDC) are proteins with unique α-helical coils that mediate a variety of biological functions, including intracellular material transport, adjustment and control of cell polarity, composition of cytoskeleton, molecular identification, and signal transduction pathway.12 At present, a variety of CCDC proteins have been confirmed to be related to the occurrence and development of papillary thyroid carcinoma,13 gastric cancer,14 breast tumor,15 bladder cancer16 and esophageal squamous cell carcinoma,17 which offer an academic foundation for targeted therapies of tumors. Unlike other proteins containing coiled-coil domains. CCDC103 (Coiled-Coil Domain Containing 103) is a genetic locus responsible for encoding a protein with a predicted molecular weight of 29 kDa. This protein exhibits notable features, including N- and C-terminal coiled-coil regions, as well as an internal α-helical RPAP3_C structural domain. Notably, the RPAP3_C domain is also found in the RNA polymerase II-associated protein3 and SPAG1, a recognized dynein assembly factor.18 The biophysical properties of CCDC103 are indeed intriguing, as it demonstrates the capacity to form dimers and higher-order structures in the presence of detergent. Moreover, it exhibits remarkable stability against heating-induced denaturation and the ability to reassemble such structures after exposure to thermal stress.19 Dysregulation of its expression can result in primary ciliary dyskinesia, which leads to the development of asthenozoospermia and infertility.20 However, there are no report of CCDC103 in solid tumors.
To comprehend the potential role of CCDC103 in glioma, this study conducted a comprehensive investigation into the diagnostic and prognostic significance of CCDC103 in glioma through the utilization of data mining techniques on datasets from the Chinese Glioma Genome Atlas (CGGA), The Cancer Genome Atlas (TCGA), and the Gene Expression Omnibus (GEO) database. Subsequently, gene ontology (GO) analysis and gene set enrichment analysis (GSEA) were performed to determine the possible biological functions and pathways of CCDC103 in glioma. The relationship between CCDC103 expression and the infiltration of immune cells in glioma was analyzed from different algorithms, including the “ssGSEA” algorithm, the “ESTIMATE” algorithm, and the “QUANTISEQ” algorithm. In addition, the role of CCDC103 in glioma expression was analyzed experimentally by immunohistochemistry (IHC), qRT-PCR, growth curve, transwell, and wound healing assays. These findings suggest that CCDC103 possesses the capability to modulate the infiltration of immune cells within gliomas. Furthermore, the expression level of CCDC103 holds promise as a potential prognostic biomarker for patients afflicted with these tumors.
Materials and Methods
Data Processing and Differential Expression Analysis
Tumor RNA-seq data (TCGA) (https://portal.gdc.cancer.gov/)21 from the Cancer Genome Atlas (TCGA) database could be acquired through the Genomic Data Commons (GDC) data portal website. Each tumor was linked with a normal tissue sample that included mRNA expression data. For statistical analysis, R software v4.0.3 was used. Two sets of data were detected using the rank sum test, and a P value of 0.05 was considered statistically significant. We downloaded the standardized pan-cancer dataset from the UCSC(https://xenabrowser.net/) database: TCGA TARGET GTEx (PANCAN,N=19131,G=60499), further we extracted ENSG00000167131(CCDC103) gene expression data in each sample, and further we screened the sample sources as follows: Solid Tissue Normal, Primary Solid Tumor, Primary Tumor, Normal Tissue, Primary Blood Derived Cancer - Bone Marrow, Primary Blood Derived cancer-peripheral Blood samples, in addition, samples whose expression level is 0 were filtered, and log2(x+0.001) was further transformed for each expression value. Finally, Cancer species with less than 3 samples in a single cancer species were excluded. Finally, the expression data of 34 cancer species were obtained. TCGA-GBMLGG (lower grade glioma and glioblastoma) cohort data and corresponding clinical information of 706 glioma patients were downloaded from the TCGA website. CGGA-GBMLGG cohort data and corresponding clinical information of 325 glioma patients were downloaded from The CGGA (Chinese Glioma Genome Atlas) (www.cgga.org.cn) website. Data analysis was performed with the R (version 3.6.3) and ggplot2 [3.3.3] packages. The expression data were normalized to transcripts per kilobase million (TPM) values before further analysis. Two sets of data were detected using the rank sum test, and significant analysis was considered when the false discovery rate (FDR) was less than 0.01 and the p-value was less than 0.05.
The GSE4290 was downloaded from the GEO database22 (https://www.ncbi.nlm.nih.gov/gds), and it was used to analyze the differential expression of genes.
Correlation Between CCDC103 Expression and Clinicopathological Features of Glioma Patients
We analyzed the correlation between CCDC103 expression and clinicopathological features of glioma patients by using TCGA and CGGA (Chinese Glioma Genome Atlas) (www.cgga.org.cn)23 database. Clinicopathological features included tumor tissue types, tumor grade, isocitrate dehydrogenase (IDH) status, IDH codel subtype, chromosomes 1p/19q codeletion status, MGMT promoter status, transcriptome subtype, gender, chemo status, etc.
Survival Prognosis Analysis
We acquired the CCDC103 expression data and clinically relevant information of glioma from the TCGA and CGGA databases. We analyzed the effect of CCDC103 on the prognosis of glioma patients. At the same time, we also analyzed whether the impact of CCDC103 on the prognosis of glioma patients was correlated with clinicopathological features.
GO and KEGG Pathway Enrichment Analysis
The Kyoto Encyclopedia of Genes and Genomes (KEGG),24 Gene Ontology enrichment pathways25 and GSEA enrichment analysis26 were used to explore the biological processes and molecular functions of CCDC103 in glioma. To determine the underlying mechanism of CCDC103, we first performed Pearson correlation analysis (|r| > 0.4 and p < 0.05) to analyze the relevant genes in the data set. Significant analysis was considered when the false discovery rate (FDR) was less than 0.01 and the p-value was less than 0.05.
Relationship Between CCDC103 and Tumor Immune Environment in Glioma
In order to roundly explore whether the expression of CCDC103 was related to the immune cell infiltration in glioma, the data of immune infiltrating cells were analyzed from the different algorithms, included “ssGSEA” algorithm,27 “ESTIMATE” algorithm28 and “QUANTISEQ” algorithm.29 In addition, we also analyzed the correlation between CCDC103 and immune checkpoint molecules through the database. In the end, we used the TIDE (http://tide.dfci.harvard.edu/)30 to predict the correlation between CCDC103 expression and immunotherapy sensitivity.
Cell Culture
The glioma cell lines A172, T98G, U251, and SF295, along with Normal Human Astrocytes(NHA), were procured from the Cell Bank of the Kunming Institute of Zoology and cultured in DMEM (Corning) containing with 10% fetal bovine serum (FBS) and 1% penicillin/streptomycin at 37°C with 5% CO2. All cells were passaged three times, and the medium was changed every two days.
Cell Transfection
The CCDC103 siRNA plasmid and control siRNA were synthesized by GenePharma (Shanghai, China). U251 and SF295 cells were treated with Lipofectamine 3000 reagent (Invitrogen, Carlsbad, CA, United States) on 6-well plate. The CCDC103 siRNA sequence were: GCAGGACCAUAAUAAGAAACC. Scrambled siRNA was used as negative control.
Quantitative Real-Time PCR
Total RNA from glioma was extracted by TRIzol reagent (Invitrogen, USA) and reverse transcribed by EasyScript First-Strand cDNA Synthesis SuperMix Kit (TransGen Biotech, China). Fast Start Universal SYBR Green Master was used for RT-PCR (Roche, Cat. 04194194001). The following primers were used: CCDC103-F: GAATGCTGCCAAGTTACGGG, CCDC103-R: TGATGCAAGGACAATACCCCT; 18sRNA-F: AACCCGTTGAACCCCATT, 18sRNA-R: CCATCCAATCGGTAGTAGCG. Expression was quantified using the 2−ΔΔCt method.30
Western Blotting
Protein samples were separated using 10% sodium lauryl sulfate–polyacrylamide gel electrophoresis (SDS–PAGE) and then transferred to a polyvinylidene fluoride (PVDF) membrane before blocking at room temperature for 1 h in 5% skim milk. Next, the membrane was incubated with the primary antibody (CCDC103,CUSABIO, CSB-PAS12877LA01HU, China,1:1000;β-actin, Proteintech, Cat No. 81115-1-RR, 1:5000, China) at 4 °C overnight and then incubated with the secondary antibody (Proteintech, Cat. No. SA00001− 2;1:5000, China) at room temperature.
Immunohistochemistry
Antigen retrieval was performed using an antigen retrieval kit (Transgenitor bio). The slides were dewaxed and treated with 3% hydrogen peroxide and BSA (5%, Bioss). The diluted primary antibody was added to the slides and incubated overnight. The primary antibody was anti-CCDC103 (CUSABIO, CSB-PAS12877LA01HU, China), and the secondary antibody was anti-Rabbit IgG (K5007, Agilent Technologies, USA). DAB kit (K5007, Agilent Technologies, USA) was used for staining and hematoxylin was used for restaining. Slides are viewed under a laboratory microscope (Nikon, Japan).
GrowthCurve
1×104 cells were planted on 6-well plates. After 24 hours, tumor cells were digested with trypsin and 20ul cells were absorbed and placed on slides for counting, once a day for 6 consecutive days.
Colony Formation Experiment
The transfected cells were grown in 6-well plates with the medium changed every two days. After 14 days, the cells were fixed with a fixative solution, stained with crystal violet, and photographed for statistics.
Transwell Assay
The migration ability of tumor cells was detected by transwell cell culture system. 1×105 cells were suspended in 1mL serum-free DMEM medium, and 100ul cell suspension was placed in transwell chamber. After one day, the cells were stained with crystal violet and counted.
Wound healing Assay
The 1×105 cells were planted in the six-well plate. After 24 hours, the 6-well plates were scratched with the head of a pipetting gun. At this time, the width of the scratches was recorded under a microscope. Thirty-six hours later, the six-well plate was again placed under a microscope and photographed to record the width of the scratches.
Statistical Analysis
The expression of CCDC103 in tumors was analyzed by using the CGGA and TCGA databases. Log rank test was used for survival analysis. The relationships between genes were assessed by P value and Spearman correlation modulus. Wilcoxon rank-sum test was used for continuous variables and chi-square test was used for categorical variables to assess the correlation between CCDC103 and pathological features. The expression of CCDC103 in normal glial cells and tumor cells was detected by ANOVA (Analysis of Variance). All cytological experiments were repeated three times. P-value less than 0.05 was defined statistically significant.
Results
Expression Analysis of CCDC103 in Normal Tissues and Tumor Tissues in Human
We evaluated the differential expression of CCDC103 between human normal tissues and tumor tissues through TCGA database. Our results indicated that CCDC103 was upregulated in Glioblastoma(GBM), Low grade glioma (LGG), Breast invasive carcinoma (BRCA), Lung adenocarcinoma(LUAD), Esophageal carcinoma., (ESCA), Kidney renal papillary cell carcinoma(KIRP), Colon adenocarcinoma, (COAD), Prostate adenocarcinoma(PRAD), Stomach adenocarcinoma, (STAD), Head and Neck squamous cell carcinoma(HNSC), Kidney renal clear cell carcinoma(KIRC), Liver hepatocellular carcinoma(LIHC), Skin Cutaneous Melanoma(SKCM), Bladder Urothelial Carcinoma(BLCA), Thyroid carcinoma(THCA), Ovarian serous cystadenocarcinoma(OV), Pancreatic adenocarcinoma(PAAD), Acute Myeloid Leukemia(LAML), Adrenocortical carcinoma(ACC) and Gallbladder cancer(CHOL), but downregulated in Testicular Germ Cell Tumors(TGCT) and Kidney Chromophobe(KICH) (Figure 1A and Supplementary Figure 1). Subsequently, we also demonstrated the high expression of CCDC103 in glioma by using TCGA (Figure 1B) and GEO (Figure 1C) databases. In addition, through TCGA and CGGA database analysis, we found that CCDC103 expression was significantly different in different glioma tissue types (Figure 1D), and the expression level of CCDC103 was positively correlated with the grade of glioma (Figure 1E). Receiver operating characteristic (ROC) analysis CCDC103 showed an Area Under Curve (AUC) value of 0.905 in low grade glioma patients and an AUC value of 0.939 in GBM patients (Figure 1F). These results confirmed that CCDC103 has the potential to be used as a diagnostic indicator for glioma.
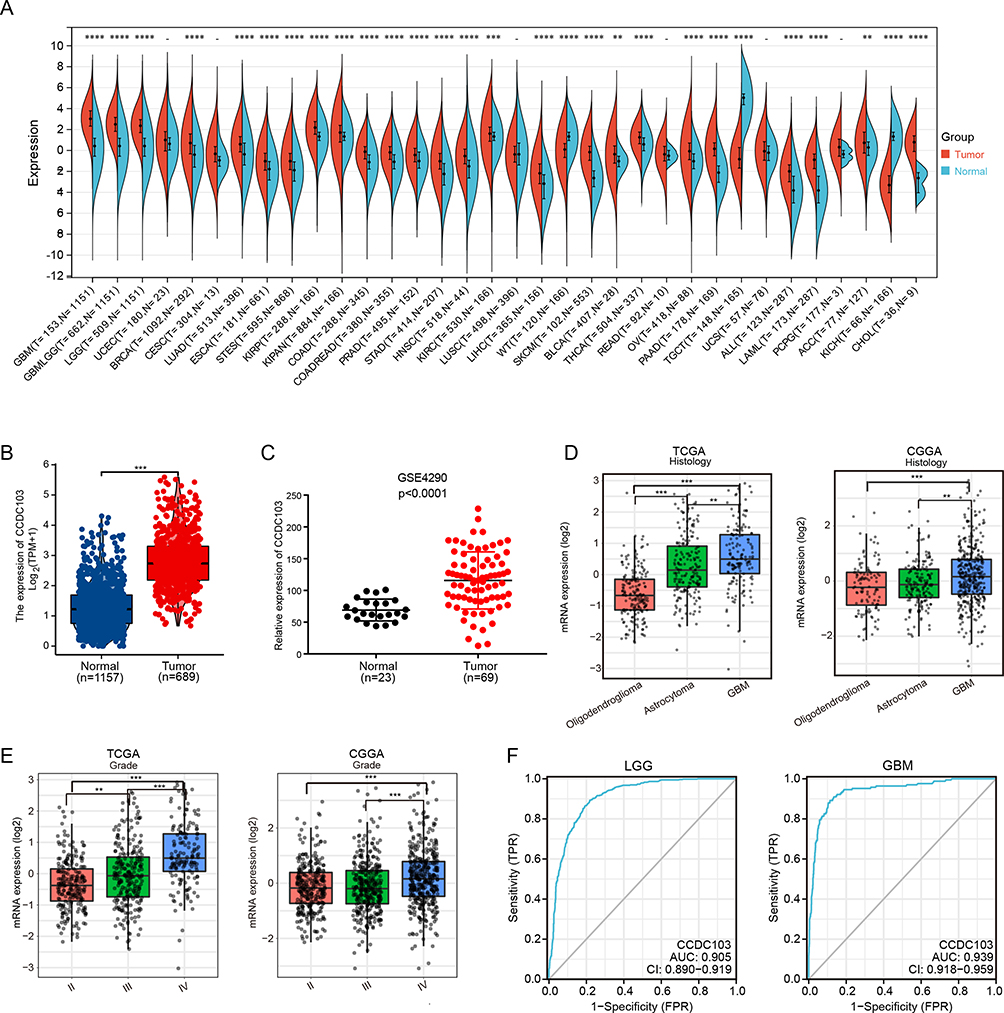 |
Figure 1 The expression of CCDC103 in human cancer. (A)The expression of CCDC103 in pan-cancer examined using the TCGA database. (B–C) The expression of CCDC103 in glioma examined by using TCGA and GEO databases. (D and E) The expression of CCDC103 in various glioma tissues types and tumor-grade based on the TCGA and CGGA databases. (F) ROC curve demonstrated the sensitivity and specificity of CCDC103 in the diagnosis of glioma. **P<0.01, ***P<0.001, ****P<0.0001. |
Correlation Between CCDC103 Expression and Clinicopathological Features of Glioma Patients
TCGA and CGGA databases were used to evaluate the relationships between CCDC103 and clinicopathological features of glioma patients. The results showed that CCDC103 expression was related with isocitrate dehydrogenase (IDH) mutation status, IDH codel subtype, chromosomes 1p/19q co-deletion status, MGMT promoter status, pan-glioma RNA expression cluster, pan-glioma DNA methylation cluster, transcriptome subtype, chemotherapy status and gender in glioma (Figure 2A–C and Table 1). These results demonstrated that CCDC103 expression was significantly associated with the development and progression of glioma.
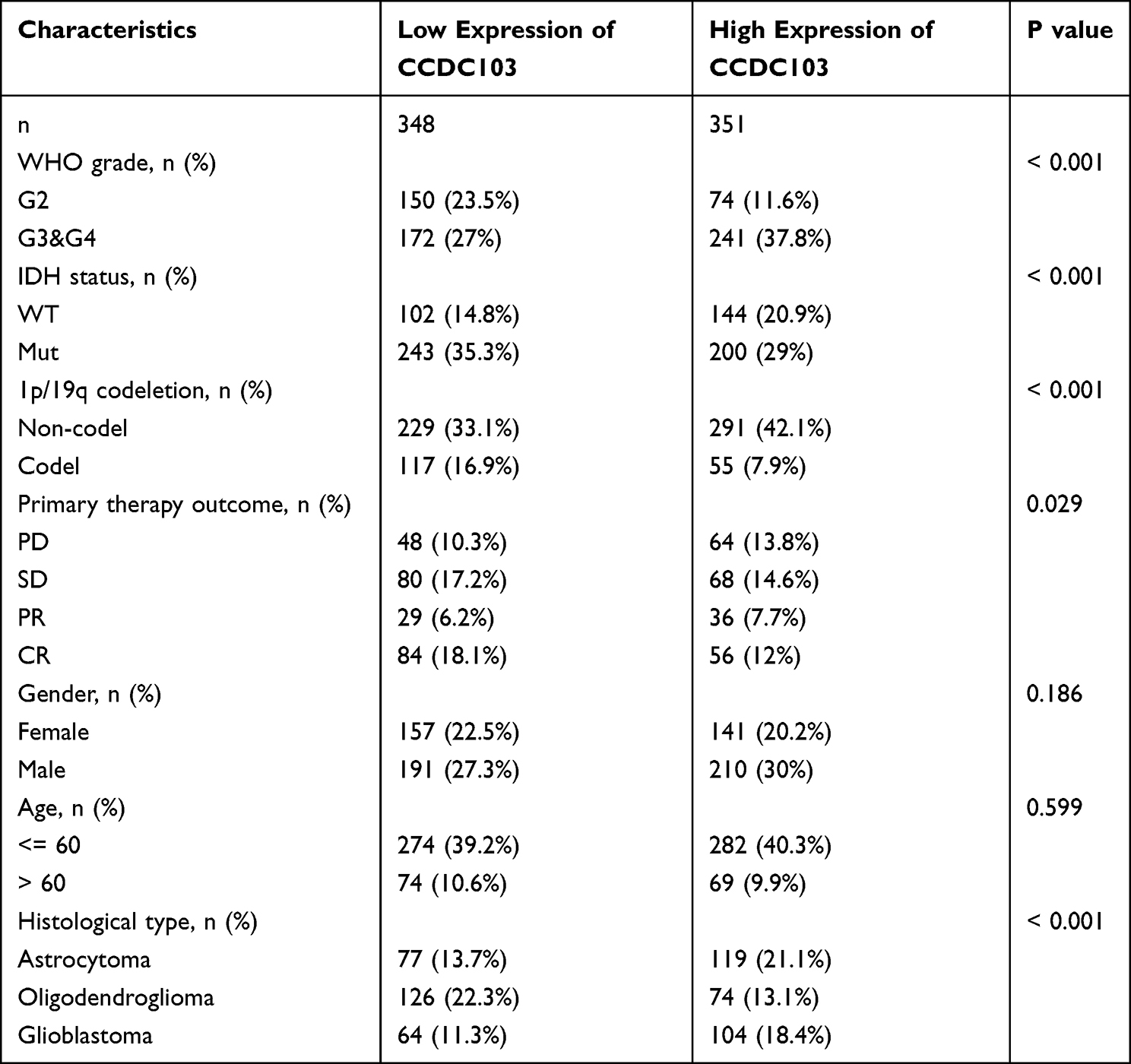 |
Table 1 The Correlation Between CCDC103 Expression and Clinical Features in Glioma |
 |
Figure 2 Association Between CCDC103 Expression and Clinicopathologic Characteristics of glioma patients. (A and B) Correlation between CCDC103 expression and clinicopathological characteristics in glioma by TCGA database. (C) Correlation between CCDC103 expression and clinicopathological characteristics in glioma by CGGA database.*P<0.05, **P<0.01, ***P<0.001. |
Prognostic Value of CCDC103 in Glioma
Through TCGA analysis, it was discovered that CCDC103 is associated with the prognosis of various types of tumors. Specifically, high expression of CCDC103 was found to be correlated with poor prognosis in glioma patients. This finding was further validated using data from the CGGA database, which indicated that CCDC103 plays a negative role in the prognosis of glioma patients (Figure 3A and Supplementary Figure 1). The Kaplan–Meier curves were plotted in subgroups of data from TCGA database, including tumor grade, IDH mutation status, 1p/19q codeletion, patient gender, patient age, histological type, and primary therapy outcome. Subgroup analysis indicated that CCDC103 overexpression was significantly affected the overall survival in glioma cases of grade G2 (P=0.037), G3&G4 (p<0.001), IDH status WT (P<0.001), 1p/19q non-codel (P<0.001), male (P<0.001), female (p=0.003), astrocytoma (P=0.006), and primary therapy outcome SD (P=0.007), respectively (Figure 3B–D). CGGA database was used to confirm the above results from CGGA database (Figure 3E and F).
 |
Figure 3 Analysis of the prognostic value of CCDC103 in different glioma subgroups. Analysis of the prognostic value of CCDC103 in different subgroups, including(A) OS (B) WHO grade (C–D) IDH mutation status,1p/19q, therapy outcome, age, histology type and gender.(E–F) in CGGA result.*P<0.05, **P<0.01, ***P<0.001. |
Logistics regression analysis was used to prove which factors were risk factors for glioma patients. We found that glioma tumor grade, IDH mutation status, 1p/19q co-deletion status, patient age and glioma tissue type are risk factors for glioma (Table 2). Meanwhile, Nomogram diagram and Calibration curve were used to predict the effects of various variables on the prognosis of glioma patients. We found that 1-,3-, and 5-year Overall Survival and Disease Free Survival were associated with CCDC103 expression, IDH mutation status, 1p/19q co-deletion status, primary therapy outcome and patient age in patients with glioma (Figure 4A–D). Univariate and multivariate analyses showed that CCDC103 was an independent predictor of prognosis in patients with glioma (Figure 4E, F and Table 3). These results suggested that high expression of CCDC103 in gliomas was associated with poor prognosis.
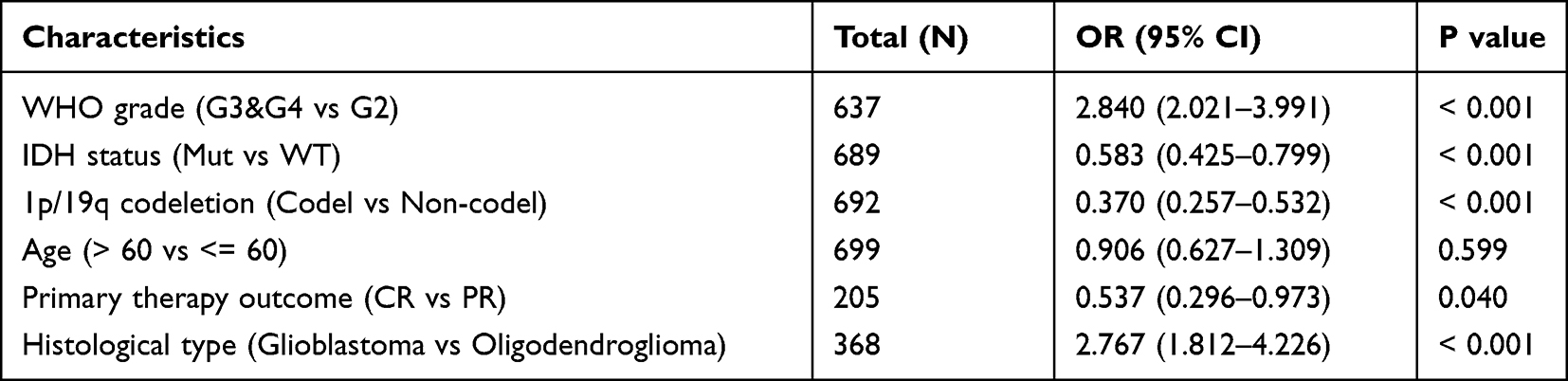 |
Table 2 Logistic Analysis of the Correlation Between CCDC103 Expression and Clinical Features in Glioma |
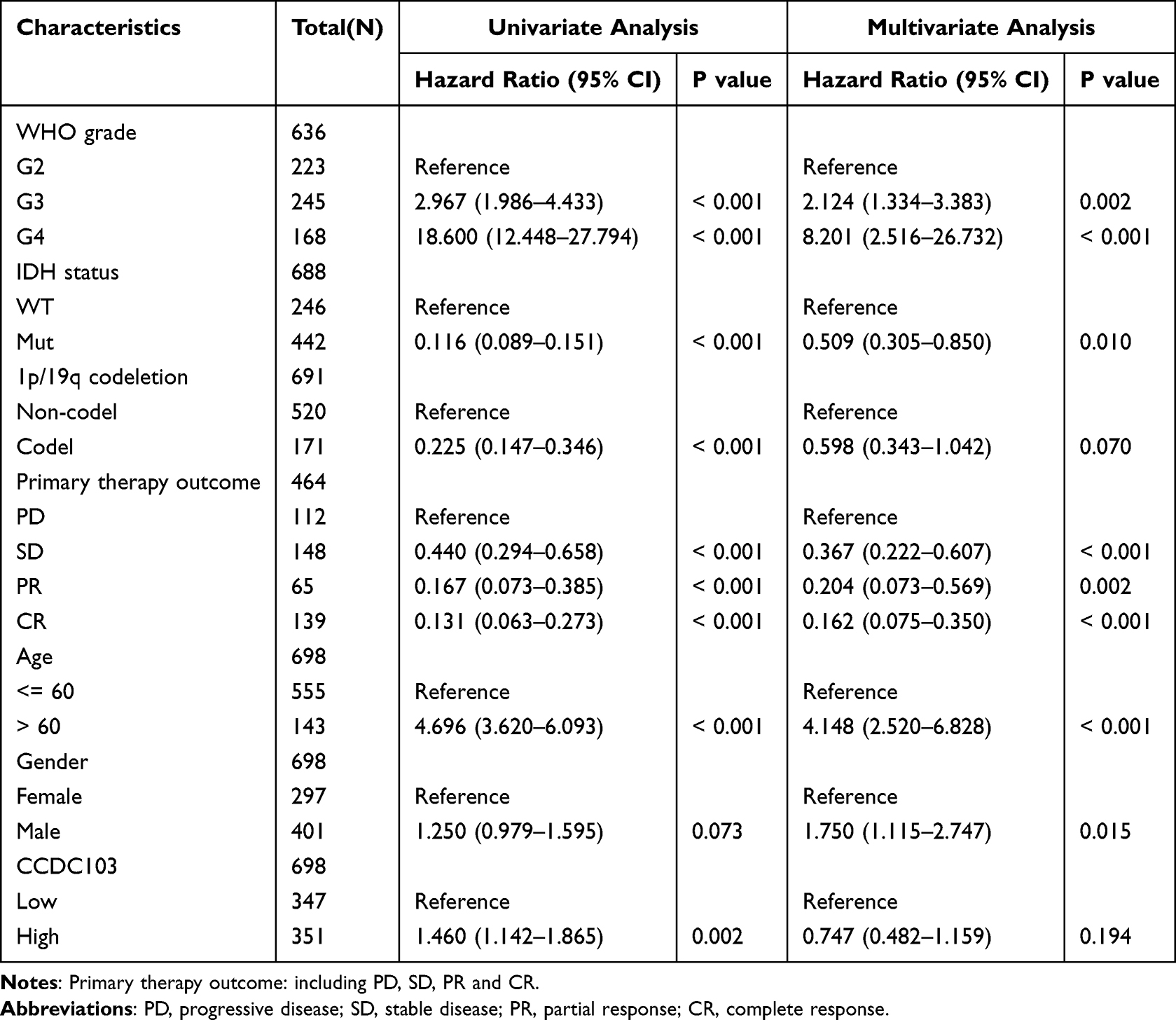 |
Table 3 Univariate and Multivariate Analysis of the Prognostic Value of CCDC103 in Glioma |
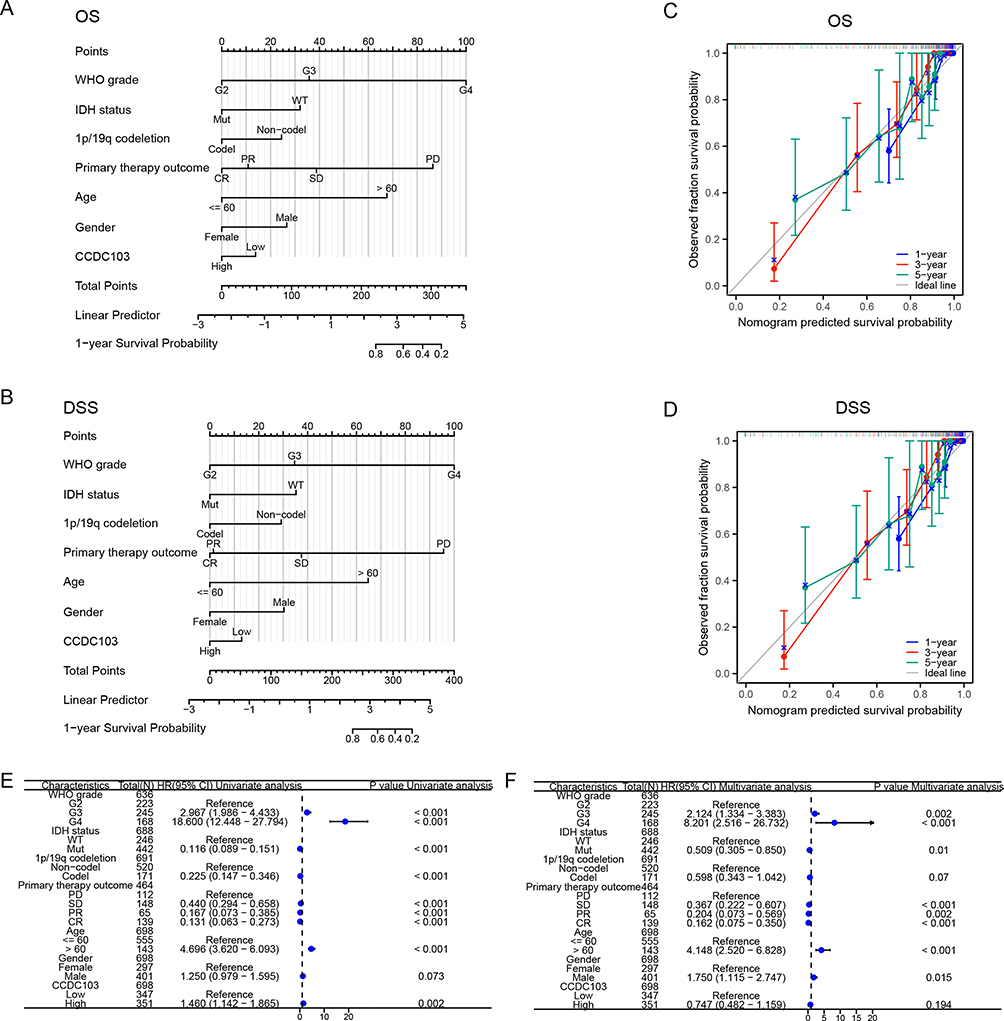 |
Figure 4 Construction a nomogram to evaluate the prognosis of CCDC103 in glioma. (A and B) Construction of a nomogram to evaluate the survival in patients with glioma. (C and D) The calibration curve used to analyze the TCGA-glioma cohort for OS and DSS. (E and F) Univariate and multivariate regression survival model of prognostic covariates in patients with glioma. |
Enrichment Analysis of CCDC103 in Glioma
To investigate the molecular mechanisms of CCDC103 in glioma, functional enrichment analyses were executed. The enriched Gene Ontology(GO) items in Biological Process(BP) included humoral immune response, lymphocyte mediated immunity, B cell mediated immunity and immunoglobulin mediated immune response (Figure 5A). The enriched GO items in Cellular Component(CC) included synaptic membrane, immunoglobulin complex, presynapse, extemal side of plasma membrane, transporter complex and postsynaptic membrane (Figure 5B). The enriched GO items in Molecular Function(MF) included passive transmembrane transporter activity, channel activity, substrate-specific channel activity, ion channel activity, metal ion transmembrane transporter activity, gated channel activity and antigen binding (Figure 5C). KEGG pathway analysis indicated enrichment in MAPK signaling pathway, Ras signaling pathway, cAMP signaling pathway, Calcium signaling pathway, GABAergic synapse and synaptic vesicle cycle (Figure 5D). These results showed that CCDC103 was highly correlated with immune activation.
 |
Figure 5 Function enrichment analysis of CCDC103 in glioma. (A–C) GO analyses of genes co-expressed with CCDC103 were performed by using TCGA data. (B–D) KEGG analyze of genes co-expressed with CCDC103 was performed by using TCGA database. |
Ultimately, we analyzed the signaling pathways involved in CCDC103 by GSEA. The results showed that CCDC103 was involved in JAK-STAT signaling pathway, aurora B pathway, DNA methylation, P53 downstream pathway, antigen processing cross presentation, cancer immunotherapy by PD1 blockade, primary immunodeficiency, G2/M checkpoints and CTLA4 pathway (Figure 6A–C).
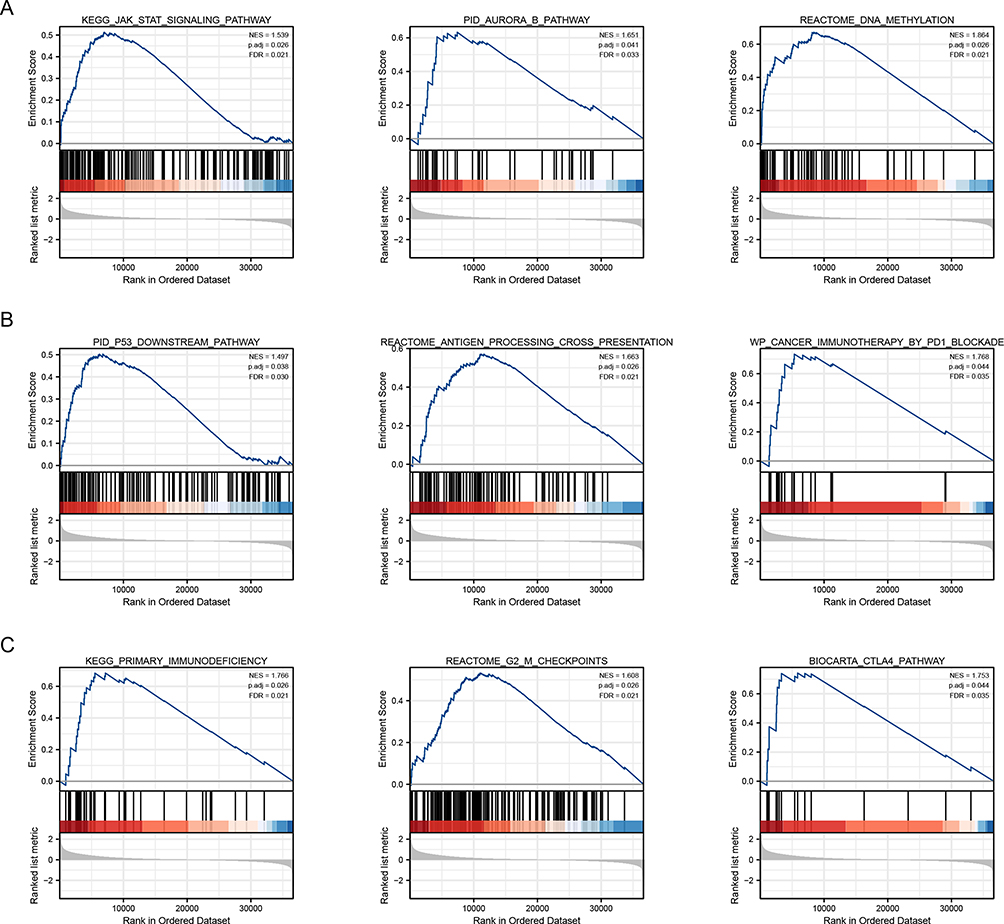 |
Figure 6 The signaling pathways involved in CCDC103 were analyzed by GSEA tool. (A–C) The involvement of genes co-expressed with CCDC103 in glioma signaling pathways as examined by GSEA. |
Relationship Between CCDC103 and Tumor Immune Environment in Glioma
Tumor-infiltrating cells influence tumor progression and patient survival.31 We investigated whether CCDC103 expression in glioma patients was related to the degree of immune infiltration. We assessed the relationship between the expression of CCDC103 and immune infiltration. Based on “ssGSEA” algorithm, we found that CCDC103 expression was correlated with immune infiltration (Figure 7A–D). In addition, based on “ESTIMATE” (Figure 7E–G) and “QuanTIseq” algorithms (Supplementary Figure 2), we again demonstrated that CCDC103 was associated with immune invasion in glioma.
 |
Figure 7 The effects of CCDC103 expression on immune cell infiltration in glioma. (A–D) Correlation between CCDC103 and immune cell infiltration in glioma based on “ssGSEA” algorithm. (E–G) Correlation between CCDC103 and immune cell infiltration in glioma based on “ESTIMATE” algorithm. *P<0.05, **P<0.01, ***P<0.001. |
Immune checkpoint molecules regulated immune responses to prevent inappropriate immune activation and allow self-tolerance.32 Then, we analyzed the correlation between CCDC103 and immune checkpoint molecules in glioma using TCGA database. The results showed that CCDC103 was positively correlated with several immune checkpoint molecules, such as CD274, CTLA4, HAVCR2, PDCD1, PDCD1LG2 and SIGLEC15 (Figure 8A–C and Supplementary Figure 3). Importantly, there was a significant difference in the strength of the immune response in the high and low CCDC103 expression groups, and the weak immune response in the high expression group, indicating that the high expression group had a higher possibility of immune escape, while the low expression group was more beneficial to anti-PD1-CTLA4 (Figure 8D).
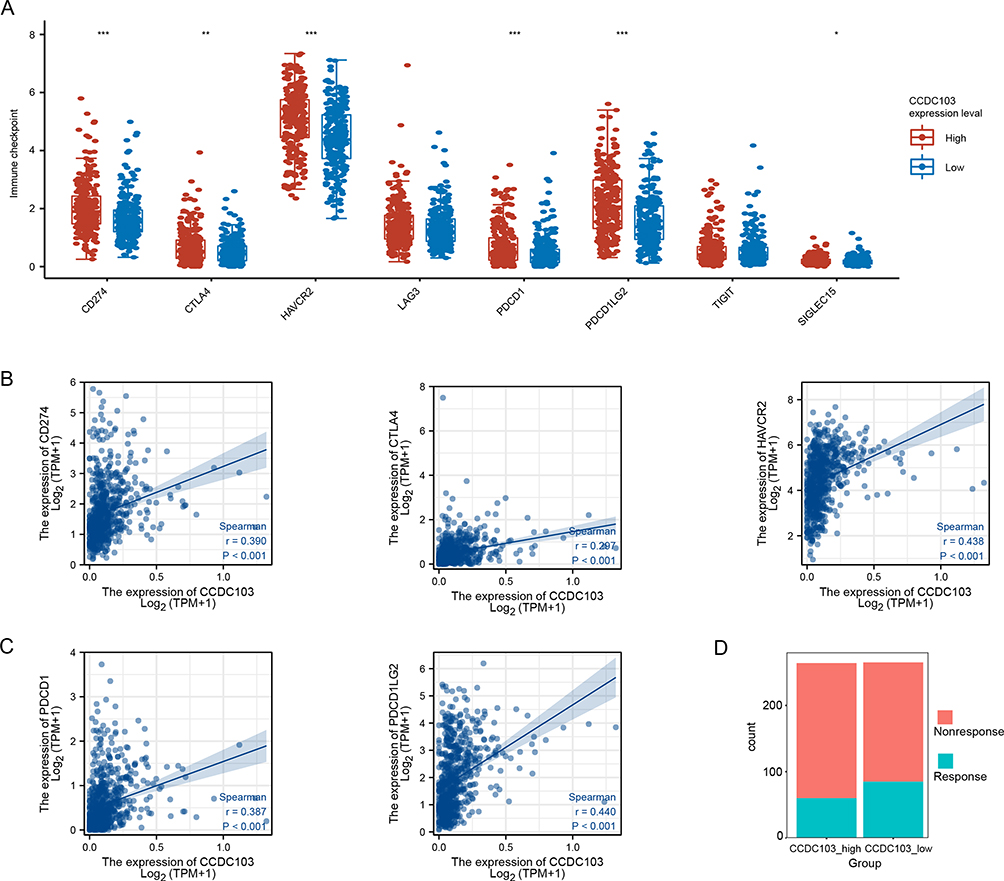 |
Figure 8 Correlation between CCDC103 expression and immune checkpoint molecules. (A–C) The correlation between CCDC103 expression and immune checkpoint molecules. (D) The correlation between CCDC103 expression and immunotherapy sensitivity. *P<0.05, **P<0.01, ***P<0.001. |
CCDC103 Knockdown Inhibits Glioma Cell Growth and Migration
By using qRT-PCR assay and Western blotting, we observed that CCDC103 demonstrated a high level of expression in glioma cell lines, particularly in U251 and SF295 cells, while normal human astrocytes exhibited no expression (Figure 9A and B). Additionally, immunohistochemistry analysis revealed that CCDC103 expression level was considerably higher in glioma tissues compared to paracancer tissues (Figure 9C). Through qRT-PCR assay, we confirmed that knockdown of CCDC103 in glioma cell lines significantly decreased cell proliferation (Figure 9D). Notably, the growth curve data revealed that CCDC103 knockdown considerably impeded glioma cell proliferation rates (Figure 9E). Knockdown of CCDC103 significantly suppressed colony formation ability, indicating the crucial role of gene CCDC103 in promoting clonogenic potential (Figure 9F and G). Furthermore, we employed Transwell and Wound Healing tests to evaluate the impact of CCDC103 knockdown on the migratory ability of glioma cells, and our findings indicated that it inhibited their migration (Figure 9H–K). Overall, these results suggest that CCDC103 is highly expressed in glioma cells and is significantly associated with glioma cell proliferation and migration.
 |
Figure 9 Knockdown CCDC103 expression inhibits the proliferation and migration of glioma cells. (A)The expression of CCDC103 in glioma cell line was analyzed by qRT-PCR assay. (B)The expression of CCDC103 in glioma cell line was analyzed by Western blotting. (C)Immunohistochemistry detection of CCDC103 in para-carcinoma tissues and glioma tissues.Scale bar, 50 μm. (D)Detection of CCDC103 knockdown efficiency in glioma cells by qRT-PCR assay. (E) Effect of knockdown CCDC103 expression on growth ability of glioma cells by GrowthCurve assay. (F and G) Effect of knockdown CCDC103 expression on colon information ability of glioma cells. (H–K) Effect of knockdown CCDC103 expression on migration of glioma cells by Transwell and wound healing assays.Scale bar, 100 μm. Each experiment were repeated at least three times. **P<0.01, ***P<0.001. |
Discussion
The prognosis of diffuse glioma is still poor even after thorough treatment, such as surgery, radiation or chemotherapy, and tumor electric field therapy. Glioblastomas account for more than 60% of diffuse gliomas, and their typical overall survival time is between 14 and 16 months.33 Because of the specific anatomical location, biological characteristics, and developmental, genetic, epigenetic, and microenvironmental properties of gliomas that cause them to acquire resistance to both conventional and novel therapies, the prognosis for gliomas has slightly improved over the past 15 years. Therefore, Patients with glioma urgently need more accurate and focused diagnostic biomarkers as well as possible treatment targets. Human CCDC103 is a compact protein consisting of 242 amino acids (AA). It comprises conserved central RPAP3_C domain flanked by N- and C-terminal coiled-coil domains.19 The pivotal function of CCDC103 lies in proper docking and assembly of outer dynein arms, which play a crucial role in promoting ciliary motion. Interestingly, CCDC103 has also been observed to localize throughout the cytoplasm of both ciliated and non-ciliated cells.34 In vitro investigations have demonstrated the ability of CCDC103 to form self-organizing oligomers. Additionally, it periodically binds to cytoplasmic microtubules, supporting the stability of assembled microtubules.18,19 Despite these extensive investigations, the precise role of CCDC103 in cancer remains unclear.
The current study, to our knowledge, is the first to thoroughly assess CCDC103 expression and its relationship with clinical and prognostic outcomes in glioma using a variety of open databases. We found that high expression of CCDC103 was associated with higher tumor grades, histological type, IDH mutation status,1p/19q chromosome co-deletion outcome, and poor OS in patients with glioma. We developed a nomogram based on multivariate Cox analysis to predict the prognosis of patients with glioma by detecting CCDC103 expression and stratifying glioma patients with better performance, and the outcomes showed that CCDC103 expression, WHO grade, IDH status, 1p/19q codeletion, and age were independent predictive factors for overall survival in glioma patients. ROC analysis also confirmed the diagnostic value of CCDC103. To explore more molecular mechanisms of CCDC103 in glioma. GO, KEGG, and GSEA enrichment studies revealed that most of the diverse pathways, including the MAPK signaling route, Ras signaling pathway, and cAMP signaling pathway, were connected to immune responses.
Various studies have shown that immune cells play a very important role in tumors.35–38 The immunosuppressive tumor microenvironment (TME) of glioma facilitates the immune evasion of neoplastic cells and is a crucial element impeding the advancement of glioma therapy.39 Immunosuppressive cells, including GAMs, Tregs, and MDSCs, play a paramount role in maintaining an immunosuppressive TME.40–42 A recent investigation has revealed that the immunosuppressive glioma microenvironment is interrelated with the gut microbiota, which is implicated in the incidence, progression, and treatment of GBM.43 Deciphering the gut-immune-brain cancer axis and examining the composition of the gut microbiota can provide novel opportunities for developing efficacious immunotherapies for malignant brain cancer.44 Accumulating evidence indicates that immune cells are suppressed in the glioma microenvironment through various mechanisms, including the presence of immune checkpoints like PD-1/PD-L1 and CTLA-4.45,46 Some studies have shown that the expression of immune checkpoint genes limits the immune surveillance of TME.30 Studies have shown that antibodies targeting CD274, CTLA-4, and PDCD1 (immune checkpoint molecules) could promote anti-tumor T cell activity and improve clinical outcomes in a variety of cancers.47–49 In this research, We explored the correlation between CCDC103 and immune checkpoint molecules and found that CCDC103 was significantly correlated with CD274, CTLA4, HAVCR2, PDCD1, PDCD1LG2 and SIGLEC15. Importantly, there was a significant difference in the strength of the immune response in the high and low CCDC103 expression groups, and the weak immune response in the high expression group, indicating that the high expression group had a higher possibility of immune escape, while the low expression group was more beneficial to anti-PD1-CTAL4. Consequently, our findings validate that CCDC103 may modulate immune cell infiltration, thereby serving as a prognostic biomarker for immunotherapy in glioma patients. Moreover, both immunohistochemistry and Western blot analysis substantiated that CCDC103 was abundantly expressed in glioma tissue and glioma cell lines. The biological functions of CCDC103 in these cells were assessed by CCDC103 knockdown, which considerably suppressed glioma cell proliferation and migration. These outcomes propose that CCDC103 could function as an oncogene in glioma; nevertheless, further investigations are required to validate these observations.
Limitations
This study is subject to several limitations. Although the correlation between CCDC103 and immune cell infiltration in glioma patients was explored, the function of CCDC103 in regulating the tumor microenvironment in glioma was not determined. Moreover, although the study demonstrated that depletion of CCDC103 could inhibit the cell migration of glioma cells, the potential molecular mechanisms of CCDC103 in cancer metastasis remain unclear. Additionally, the present study only assessed the expression and biological roles of CCDC103 in databases of patients with glioma and cultured cells, not in vivo. Therefore, further studies are required to evaluate the function of CCDC103 in glioma metastasis and its role in regulating the glioma tumor microenvironment.
Conclusions
In summary, The present study unveiled that CCDC103 was expressed in glioma and its high expression level was associated with unfavorable patient prognosis. Silencing of CCDC103 in glioma cells resulted in decreased proliferation and migration capabilities. These findings suggest that CCDC103 exerts a crucial function in the progression of glioma and has the potential to serve as a novel prognostic biomarker for patients with gliomas.
Data Sharing Statement
The GSE4290 was downloaded from The Gene Expression Omnibus (GEO) database. We downloaded a uniformly standardized pan-cancer dataset from UCSC (https://xenabrowser.net/) database: TCGA TARGET GTEx (PANCAN,N=19131,G=60499), from which we further extracted the expression data of ENSG00000160446(CCDC103) gene in each sample.
Ethics Statement
The study adheres to the principles outlined in the Declaration of Helsinki. All human tumor tissues were obtained following the Medical Ethics Committee of the Second Affiliated Hospital of Kunming Medical University’s approved protocol, and informed consent was obtained from all patients.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was subsidized by the National Nature Science Foundation of China (82160512) and the Kunming University of Science and Technology Medical Joint Special Project(- KUST-PE2022011Y). The authors would like to thank support from the second Affiliated Hospital of Kunming Medical University, Kunming, 650223, China and The Pu’er Poople’s Hospital, 665000, China.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Xu S, Tang L, Li X, et al. Immunotherapy for glioma: current management and future application. Cancer Lett. 2020;476:1–12. doi:10.1016/j.canlet.2020.02.002
2. Gusyatiner O, Hegi ME. Glioma epigenetics: from subclassification to novel treatment options. Semin Cancer Biol. 2018;51:50–58. doi:10.1016/j.semcancer.2017.11.010
3. Bianconi A, Palmieri G, Aruta G, et al. Updates in glioblastoma immunotherapy: an overview of the current clinical and translational scenario. Biomedicines. 2023;11(6):1520. doi:10.3390/biomedicines11061520
4. Masi I, Caprara V, Bagnato A, Rosanò L. Tumor cellular and microenvironmental cues controlling invadopodia formation. Front Cell Dev Biol. 2020;8:584181. doi:10.3389/fcell.2020.584181
5. Bonnans C, Chou J, Werb Z. Remodelling the extracellular matrix in development and disease. Nat Rev Mol Cell Bio. 2014;15(12):786–801. doi:10.1038/nrm3904
6. Dupont S, Morsut L, Aragona M, et al. Role of YAP/TAZ in mechanotransduction. Nature. 2011;474(7350):179–U212. doi:10.1038/nature10137
7. Heng YW, Koh CG. Actin cytoskeleton dynamics and the cell division cycle. Int J Biochem Cell Biol. 2010;42(10):1622–1633. doi:10.1016/j.biocel.2010.04.007
8. Kiss, A, Horvath P, Rothballer A, et al. Nuclear motility in glioma cells reveals a cell-line dependent role of various cytoskeletal components. PLoS One. 2014;9(4):e93431. doi:10.1371/journal.pone.0093431
9. Abe T, La TM, Miyagaki Y, et al. Phosphorylation of cortactin by cyclin-dependent kinase 5 modulates actin bundling by the dynamin 1-cortactin ring-like complex and formation of filopodia and lamellipodia in NG108-15 glioma-derived cells. Int J Oncol. 2019;54:1–10.
10. Krigers A, Demetz M, Moser P, et al. P12.05.B impact of GAP-43 and actin expression on the outcome and overall survival in diffuse and anaplastic gliomas neuro-oncology. Sci Rep. 2022;13:24.
11. Neubauer HA, Tea MN, Zebol JR, et al. Cytoplasmic dynein regulates the subcellular localization of sphingosine kinase 2 to elicit tumor-suppressive functions in glioblastoma. Oncogene. 2019;38(8):1151–1165. doi:10.1038/s41388-018-0504-9
12. Burkhard P, Stetefeld J, Strelkov SV. Coiled coils: a highly versatile protein folding motif. Trends Cell Biol. 2001;11(2):82–88. doi:10.1016/S0962-8924(00)01898-5
13. Yin DT, Xu J, Lei M, et al. Characterization of the novel tumor-suppressor gene CCDC67 in papillary thyroid carcinoma. Oncotarget. 2016;7(5):5830–5841. doi:10.18632/oncotarget.6709
14. Wang J, Wu X, Dai W, et al. The CCDC43-ADRM1 axis regulated by YY1, promotes proliferation and metastasis of gastric cancer. Cancer Lett. 2020;482:90–101. doi:10.1016/j.canlet.2020.03.026
15. Ning Y, Wang C, Liu X, et al. CK2-mediated CCDC106 phosphorylation is required for p53 degradation in cancer progression. J Exp Clin Cancer Res. 2019;38(1):131. doi:10.1186/s13046-019-1137-8
16. Gong Y, Qiu W, Ning X, et al. CCDC34 is up-regulated in bladder cancer and regulates bladder cancer cell proliferation, apoptosis a nd migration. Oncotarget. 2015;6(28):25856–25867. doi:10.18632/oncotarget.4624
17. Hu DD, Li PC, He YF, Jia W, Hu B. Overexpression of coiled-coil domain-containing protein 34 (CCDC34) and its correlation with angiogen esis in esophageal squamous cell carcinoma. Med Sci Monit. 2018;24:698–705. doi:10.12659/MSM.908335
18. King SM, Patel-King RS. The oligomeric outer dynein arm assembly factor CCDC103 is tightly integrated within the ciliary axoneme and exhibits periodic binding to microtubules. J Biol Chem. 2015;290(12):7388–7401. doi:10.1074/jbc.M114.616425
19. King SM, Patel-King RS. The outer dynein arm assembly factor CCDC103 forms molecular scaffolds through multiple self-interaction sites. Cytoskeleton. 2020;77(1–2):25–35. doi:10.1002/cm.21591
20. Zubair M, Khan R, Ma A, et al. A recurrent homozygous missense mutation in CCDC103 causes asthenoteratozoospermia due to disorganized dynein arms. Asian J Androl. 2022;24:1.
21. Chandran UR, Medvedeva OP, Barmada MM, et al. TCGA expedition: a data acquisition and management system for TCGA data. PLoS One. 2016;11(10):e0165395. doi:10.1371/journal.pone.0165395
22. Edgar R, Domrachev M, Lash AE. Gene Expression Omnibus: NCBI gene expression and hybridization array data repository. Nucleic Acids Res. 2002;30(1):207–210. doi:10.1093/nar/30.1.207
23. Zhao Z, Zhang KN, Wang Q, et al. Chinese Glioma Genome Atlas (CGGA): a comprehensive resource with functional genomic data from Chinese glioma patients. Genomics Proteomics Bioinformatics. 2021;19(1):1–12. doi:10.1016/j.gpb.2020.10.005
24. Tanabe M, Kanehisa M. Using the KEGG database resource. Curr Protoc Bioinformatics Chapter. 2012;1:
25. Chen L, Zhang YH, Zheng M, Huang T, Cai YD. Identification of compound-protein interactions through the analysis of gene ontology, KEGG enrichmen t for proteins and molecular fragments of compounds. Mol Genet Genomics. 2016;291(6):2065–2079. doi:10.1007/s00438-016-1240-x
26. Subramanian A, Tamayo P, Mootha VK, et al. Gene set enrichment analysis: a knowledge-based approach for interpreting genome-wide expression prof iles. Proc Natl Acad Sci U S A. 2005;102(43):15545–15550. doi:10.1073/pnas.0506580102
27. Xiang S, Li J, Shen J, et al. Identification of prognostic genes in the tumor microenvironment of hepatocellular carcinoma. Front Immunol. 2021;12:653836. doi:10.3389/fimmu.2021.653836
28. Ke ZB, Wu YP, Huang P, et al. Identification of novel genes in testicular cancer microenvironment based on ESTIMATE algorithm-deriv ed immune scores. J Cell Physiol. 2021;236(1):706–713. doi:10.1002/jcp.29898
29. Sturm G, Finotello F, Petitprez F, et al. Comprehensive evaluation of transcriptome-based cell-type quantification methods for immuno-oncology. Bioinformatics. 2019;35(14):i436–i445. doi:10.1093/bioinformatics/btz363
30. Bao Y, Jiang A, Dong K, et al. DDX39 as a predictor of clinical prognosis and immune checkpoint therapy efficacy in patients with cl ear cell renal cell carcinoma. Int J Biol Sci. 2021;17(12):3158–3172. doi:10.7150/ijbs.62553
31. Sahasrabudhe NM, van der Horst JC, Spaans V, et al. MGL ligand expression is correlated to lower survival and distant metastasis in cervical squamous Cel l and adenosquamous carcinoma. Front Oncol. 2019;9:29. doi:10.3389/fonc.2019.00029
32. Sangro B, Chan SL, Meyer T, Reig M, El-Khoueiry A, Galle PR. Diagnosis and management of toxicities of immune checkpoint inhibitors in hepatocellular carcinoma. J Hepatol. 2020;72(2):320–341. doi:10.1016/j.jhep.2019.10.021
33. Hanif F, Muzaffar K, Perveen K, Malhi SM. Glioblastoma multiforme: a review of its epidemiology and pathogenesis through clinical presentation and treatment. Asian Pac J Cancer P. 2017;18:3–9.
34. Panizzi JR, Becker-Heck A, Castleman VH, et al. CCDC103 mutations cause primary ciliary dyskinesia by disrupting assembly of ciliary dynein arms. Nat Genet. 2012;44(6):714–719. doi:10.1038/ng.2277
35. Ye W, Luo C, Liu F, et al. CD96 correlates with immune infiltration and impacts patient prognosis: a pan-cancer analysis. Front Oncol. 2021;11:634617. doi:10.3389/fonc.2021.634617
36. Zhang QF, Li J, Jiang K, et al. CDK4/6 inhibition promotes immune infiltration in ovarian cancer and synergizes with PD-1 blockade in a B cell-dependent manner. Theranostics. 2020;10(23):10619–10633. doi:10.7150/thno.44871
37. Tran TO, Vo TH, Lam LHT, Le NQK. ALDH2 as a potential stem cell-related biomarker in lung adenocarcinoma: comprehensive multi-omics analysis. Comput Struct Biotechnol J. 2023;21:1921–1929. doi:10.1016/j.csbj.2023.02.045
38. Dang HH, Ta HDK, Nguyen TTT, et al. Prospective role and immunotherapeutic targets of sideroflexin protein family in lung adenocarcinoma: evidence from bioinformatics validation. Funct Integr Genomic. 2022;22(5):1057–1072. doi:10.1007/s10142-022-00883-3
39. Polyak K, Haviv I, Campbell I. Co-Evolution of tumor cells and their microenvironment. Trends Genet. 2009;25(1):30–38. doi:10.1016/j.tig.2008.10.012
40. Ginhoux F, Lim S, Hoeffel G, Low D, Huber T. Origin and differentiation of microglia. Front Cell Neurosci. 2013;7:45. doi:10.3389/fncel.2013.00045
41. Wing K, Onishi Y, Prieto-Martin P, et al. CTLA-4 control over Foxp3+ regulatory T cell function. Science. 2008;322(5899):271–275. doi:10.1126/science.1160062
42. Chai E, Zhang L, Li C. LOX-1+ PMN-MDSC enhances immune suppression which promotes glioblastoma multiforme progression. Cancer Manag Res. 2019;11:7307–7315. doi:10.2147/CMAR.S210545
43. Ferreiro A, Crook N, Gasparrini AJ, Dantas G. Multiscale evolutionary dynamics of host-associated microbiomes. Cell. 2018;172(6):1216–1227. doi:10.1016/j.cell.2018.02.015
44. D’Alessandro G, Antonangeli F, Marrocco F, et al. Gut microbiota alterations affect glioma growth and innate immune cells involved in tumor immunosurveillance in mice. Eur J Immunol. 2020;50(5):70511.
45. Ghouzlani A, Kandoussi S, Tall M, et al. Immune checkpoint inhibitors in human glioma microenvironment. Front Immunol. 2021;12:679425. doi:10.3389/fimmu.2021.679425
46. Liu F, Huang J, Liu X, et al. CTLA-4 correlates with immune and clinical characteristics of glioma. Cancer Cell Int. 2020;20(1):7. doi:10.1186/s12935-019-1085-6
47. Shoji T, Saito R, Chonan M, et al. Local convection-enhanced delivery of an anti-CD40 agonistic monoclonal antibody induces antitumor ef fects in mouse glioma models. Neuro Oncol. 2016;18(8):1120–1128. doi:10.1093/neuonc/now023
48. Wang Q, Li M, Yang M, et al. Analysis of immune-related signatures of lung adenocarcinoma identified two distinct subtypes: implic ations for immune checkpoint blockade therapy. Aging. 2020;12(4):3312–3339. doi:10.18632/aging.102814
49. Stirling ER, Bronson SM, Mackert JD, Cook KL, Triozzi PL, Soto-Pantoja DR. Metabolic implications of immune checkpoint proteins in cancer. Cells. 2022;11(1):179. doi:10.3390/cells11010179
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.