Back to Journals » Journal of Inflammation Research » Volume 16
Cartilage Endplate-Derived Stem Cells for Regeneration of Intervertebral Disc Degeneration: An Analytic Study
Authors Jia Z ![]() , Liu D, Li X, Wen T, Li W
, Liu D, Li X, Wen T, Li W
Received 23 July 2023
Accepted for publication 28 November 2023
Published 4 December 2023 Volume 2023:16 Pages 5791—5806
DOI https://doi.org/10.2147/JIR.S431986
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor Ning Quan
Zhiwei Jia,1 Donghua Liu,1 Xingxuan Li,1 Tianlin Wen,1 Wei Li2
1Department of Orthopedics, Dongzhimen Hospital, Beijing University of Chinese Medicine, Beijing, People’s Republic of China; 2Department of Sports Medicine, Fourth Medical Center of PLA General Hospital, Beijing, People’s Republic of China
Correspondence: Tianlin Wen; Wei Li, Email [email protected]; [email protected]
Purpose: Intervertebral disc degeneration (IDD) is considered the predominant cause of low back pain (LBP) and accounts for global disability and a substantial socioeconomic burden. Given the unsatisfactory outcomes of current treatment strategies, cartilage endplate-derived stem cells (CESCs) are increasingly used in intervertebral disc regeneration. However, comprehensive analyses on CESCs remain rare. Herein, we examined the biological functions and applications of CESCs in IDD.
Methods: PubMed, Embase, and Cochrane Library databases were searched to identify studies focused on CESCs. Relevant information from included studies was extracted. Descriptive statistics were performed. Correlation analysis was conducted to determine the relationship among Web of Science (WoS) citations, Dimensions, and Altmetric Attention Score (AAS).
Results: Twenty-six studies were included in this study. Most studies (n=20) isolated CESCs from humans, followed by rats (n=5) and rabbits (n=1). Twenty studies were performed in vitro, and the remaining six were implemented both in vivo and in vitro. The findings of this study provide insight into the biological properties of CESCs and clarify their potential application for intervertebral disc regeneration. There was a very high correlation between WoS and Dimensions citation count (p< 0.001, r=0.988).
Conclusion: This study, for the first time, elaborates biological features of CESCs and analyzes their potential applications in regenerating intervertebral discs. CESCs may be promising candidates for protecting the intervertebral disc from degeneration and contributing to intervertebral disc regeneration.
Keywords: intervertebral disc degeneration, low back pain, cartilage endplate-derived stem cells, stem cells, intervertebral disc
Introduction
Low back pain (LBP), a prevalent musculoskeletal disorder, is encountered frequently in clinical settings.1–3 It is estimated that approximately 80% of the population will experience LBP at some point in their lifetime.3–5 LBP is a leading cause of sick leave and work-related disability, thereby imposing a significant economic impact on society.1,2,4,6 In the United States alone, the annual economic burden of LBP surpasses $100 billion.7,8
Although the pathogenesis of LBP is multifaceted and not fully understood, intervertebral disc degeneration (IDD) is commonly acknowledged as a primary etiological factor.3,5,6,8–12 The intervertebral disc, a fibrocartilaginous structure situated between adjacent vertebral bodies,9,10,13–16 is essential for spinal stability and mobility.11,13,16 This structure comprises three distinct tissues: the annulus fibrosus (AF), nucleus pulposus (NP), and cartilage endplate (CE).8,11,14,16 The NP, embryologically distinct from the AF and CE, originates from the notochord, whereas the latter two derive from sclerotomes.5,8–11 This complex architecture facilitates the absorption and distribution of physical forces.5,11,13–16 The CE, a thin hyaline cartilage layer, both separates the vertebral body from the intervertebral disc and protects it from NP invasion. Given the avascular nature of the intervertebral disc, nutrient and metabolite transport are primarily dependent on CE permeation.17–19 Degeneration-induced calcification or sclerosis of the CE can impede this critical nutritional and metabolic exchange,20,21 thus playing a pivotal role in the initiation and progression of IDD.22–25
Advancements in stem cell research have led to the discovery of cells with stem-like properties in various tissues and organs.26–31 These stem cells are integral to the repair or regeneration of damaged structures.5,8–10,15,16,32 In response to tissue damage or degeneration, endogenous repair mechanisms involve the recruitment of stem cells, which then proliferate and differentiate to restore structural and functional integrity.5,9 Stem cells in the intervertebral disc were initially isolated from degenerated human NP and AF cells, displaying mesenchymal stem cell (MSCs) characteristics and potential for osteogenic, adipogenic, and chondrogenic differentiation.5,8–10 Recently, cartilage endplate-derived stem cells (CESCs) have been isolated.33,34 Based on the distinct anatomical regions within the intervertebral disc, intervertebral disc-derived stem cells are typically categorized into three subsets, including NP-derived stem cells (NPSCs), AF-derived stem cells (AFSCs), and CESCs.5,8–10 CESCs are considered multipotent stem cells that likely play a significant role in maintaining the integrity and functionality of the cartilage endplate.8–10,33,34 Furthermore, CESCs have been shown to effectively mitigate IDD by promoting NP regeneration and regulating intervertebral disc homeostasis.5,9 Failure of endogenous repair by CESCs has been identified as a primary contributor to IDD.8,23,32–34
Current IDD interventions include both conservative and operative strategies.14,16,35,36 Despite significant progress in research and clinical studies, treatment efficacy remains suboptimal.35–38 This limitation is partly due to these strategies focusing more on alleviating pain than addressing the underlying cause of IDD.4,13–16,32,35–37,39 Stem cell transplantation has emerged as a novel therapeutic approach,13–15,37 but the challenging microenvironment of the intervertebral disc, characterized by high osmolarity, mechanical load, poor nutrition, low oxygen tension, and acidic pH, restricts its effectiveness.40–43 CESCs, as endogenous stem cells, are increasingly utilized in disc regeneration due to their ability to overcome transplantation barriers.5,8–10,44–46 Despite promising outcomes with CESCs in IDD regeneration, comprehensive studies on CESCs are still lacking. This study aimed to analyze the biological functions and applications of CESCs in IDD and provide insights into CESCs research, proposing future strategies for intervertebral disc regeneration.
Materials and Methods
Search Strategy
Given that human and animal subjects were not involved in the present study, approval from the Institutional Review Board was not required. We conducted a comprehensive search to identify papers focused on CESCs published from inception up to April 20, 2023. Two investigators independently retrieved data from the PubMed, Embase, and Cochrane Library databases. The keywords used for the search strategy included “cartilage endplate”, “stem cell”, “progenitor cell”, and “stromal cell”. Titles irrelevant to the topic were first removed. The abstracts of any papers potentially relevant to CESCs were further examined. The full text of each paper was reviewed if the abstract failed to provide sufficient information. Disagreements were settled through discussions, with a third investigator consulted to reach a consensus when necessary. References of included papers and related reviews were screened to identify potential publications.
Inclusion and Exclusion Criteria
The eligibility criteria for study inclusion/exclusion were as follows: (1) papers were published in English; (2) papers must involve basic research or clinical studies related to CESCs; (3) case reports, editorials, letters, and reviews were excluded.
Data Extraction and Management
Two investigators reviewed the selected studies separately and performed data extraction using a standardized form. Disagreements were resolved through discussion between investigators. A third investigator was consulted to reach a consensus when necessary. The following information was collected from included studies: authors, year of publication, journal, cell source, study type, CESCs isolation method, the method employed for surface marker detection, positive and negative markers, and key findings. Web of Science (WoS) citations of the included studies were identified in the WoS database. Dimensions citations and Altmetric Attention Score (AAS) were identified in the database of Dimensions (www.dimensions.ai).
Quality Assessment
Two investigators independently evaluated the methodological quality of eligible studies. Disagreements between investigators were resolved by discussion with a third investigator. The quality of included papers was assessed using a quality scoring system,47 which included eight questions and has been widely used in similar publications with “yes” and “no” answers assigned 1 and 0 points, respectively.48–51 The maximum achievable score is 8 points. The quality scores for included studies were independently determined. The quality of the studies was graded based on their scores. A score of 7–8 was interpreted as excellent, 5–6 as good, 3–4 as poor, and 0–2 as bad.48–51
Statistical Analysis
In this study, descriptive statistical methods, including total count and proportion, were employed to analyze the data. To examine the association between Web of Science (WoS) citations, Altmetric Attention Score (AAS), and Dimensions citations, Pearson’s correlation analysis was utilized. The Pearson’s correlation coefficient (r) was categorized as follows: a coefficient less than 0.3 indicated a poor correlation, 0.3–0.5 a low correlation, 0.5–0.7 a moderate correlation, 0.7–0.9 a high correlation, and greater than 0.9 a very high correlation.52,53 All statistical analyses were conducted using SPSS software (version 19.0; SPSS, Chicago, IL). A p-value of less than 0.05 was considered statistically significant.
Results
Study Selection
Following the primary retrieval strategy, we identified 284 papers potentially related to CESCs. Twenty papers were removed because they were duplicates. After title and abstract screening, 41 papers progressed to full-text review; subsequently, 15 papers were excluded owing to failure to meet inclusion criteria. The remaining 26 papers were included.33,34,54–77 Figure 1 presents a flowchart of the screening process.
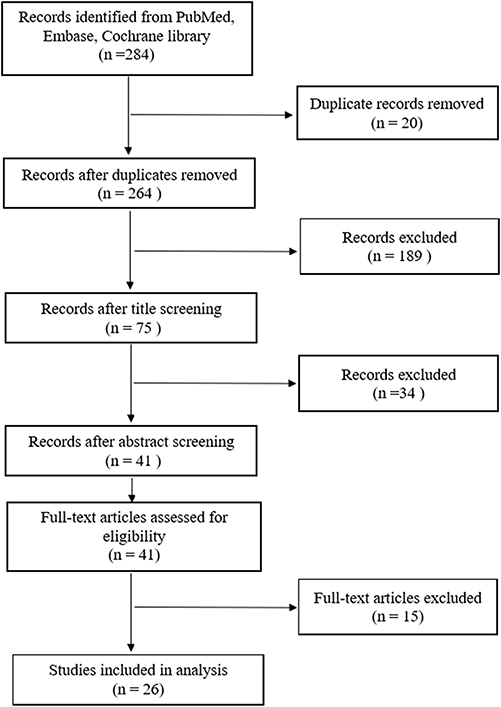 |
Figure 1 The flow chart of screening process. |
General Information on the Papers
The included studies were published between 2011 and 2022 (Table 1). The highest value of WoS citations, Dimensions citations and AAS were 145, 101 and 6, respectively. Twelve works (46%) had no AAS (Table 1). Nineteen journals contributed to 26 papers. Stem cells (3), Molecular Medicine Reports (3), European Spine Journal (2), PLoS ONE (2), and Acta Biochimica et Biophysica Sinica (2) published two or more papers. Most studies (n=20) isolated CESCs from humans, followed by rats (n=5) and rabbits (n=1) (Figure 2). Twenty papers were in vitro studies, while the remaining six involved both in vivo and in vitro experiments (Figure 2).
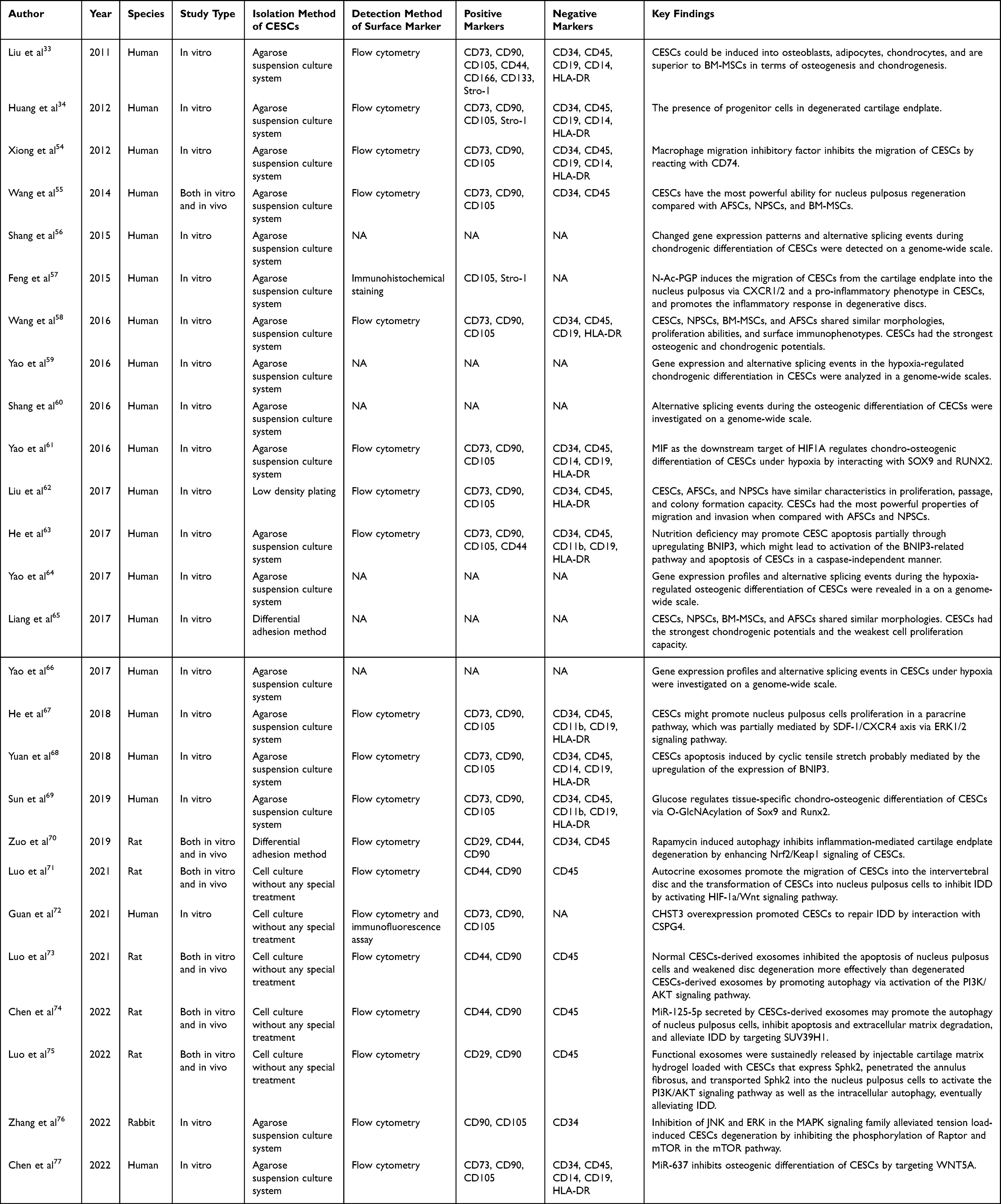 |
Table 1 Characteristics and Key Findings of the Included Studies |
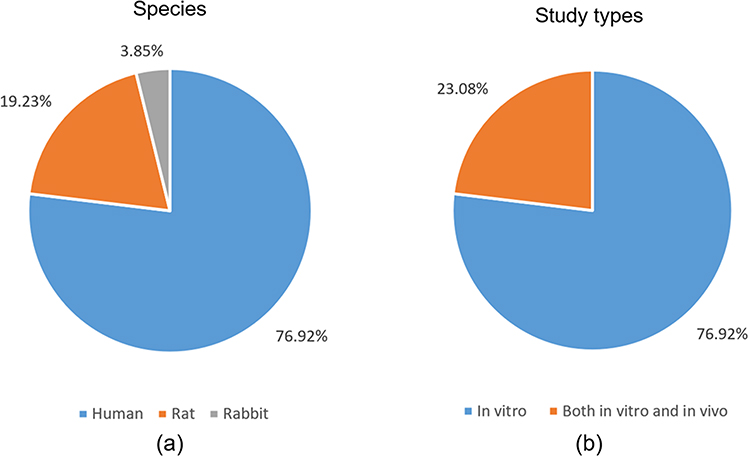 |
Figure 2 Experimental species (a) and included study types (b). |
Quality of Included Studies
Questions regarding the quality scoring systems are listed in Table 2. The methodological scores of included studies ranged from 5 to 8. Four studies achieved maximum scores. The overall quality of included studies ranged between good and excellent. In total, 17 studies received excellent, and the remaining nine were deemed good. Detailed methodological scores of included studies are presented in Table 3.
 |
Table 2 The Criteria of Quality Assessment |
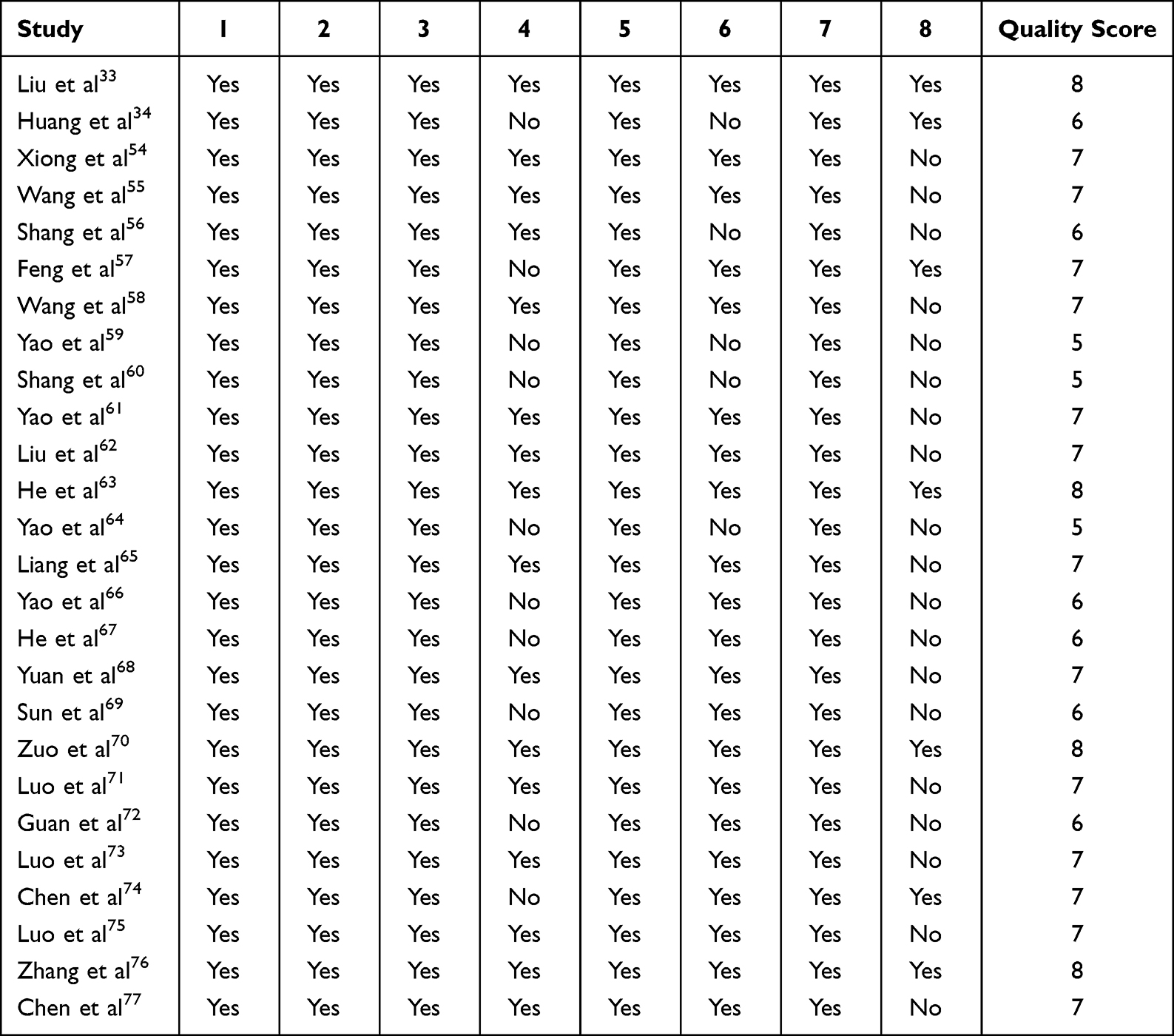 |
Table 3 Methodological Scores of the Included Studies |
CESCs Isolation
All studies provided a detailed description of the method employed for CESCs isolation (Figure 3). Among included studies, 18 isolated CESCs using the agarose suspension culture system, 5 employed cell culture without any special treatment, 2 used the differential adhesion method, and 1 used low-density plating.
 |
Figure 3 Isolation method (a) and surface marker detection of CESCs (b). |
Method Employed for Surface Marker Detection
Twenty studies performed surface marker detection. Of these, 18 studies used flow cytometry, 1 used immunohistochemical staining, and 1 used both flow cytometry and immunofluorescence assays. Figure 3 presents detailed information regarding the detection of surface markers in CESCs.
Surface Markers of CESCs
Twenty papers reported surface markers for isolated CESCs; surface markers were not reported in the other six papers. CD90 was the most common positive marker detected in 19 studies (95%), followed by CD105 (15, 75%), and CD73 (13, 65%). The majority of the studies (n=13; 65%) detected CD73, CD90, and CD105. C45 (17, 75%) was the most common negative marker, followed by CD34 (14, 70%) and CD19 (10, 50%). CD45 and CD34 (n=13; 65%) were the most common negative markers reported in the included studies. Table 1 lists CESCs surface markers detected in the included studies.
Key Findings of CESCs
Table 1 summarizes key CESCs findings of included studies. Two studies described the discovery of CESCs.33,34 Five studies compared CESCs with NPSCs, AFSCs, and bone marrow-derived stem cells (BMSCs) considering their biological properties.33,55,58,62,65 In addition, five studies reported genome-wide analyses of gene expression profiling and alternative splicing events in CESCs under normoxic and hypoxic conditions.56,59,60,64,66 Eight studies discussed the fate of CESCs in IDD.54,57,61,63,68–70,76 Eight studies elaborated on the biological potential of CESCs in intervertebral disc regeneration.55,67,71–75,77
Correlation Analysis
The citation counts of included studies in WoS were very high correlated with their citations in Dimensions (r = 0.988, p < 0.001). Figure 4 described a clear linear correlation between WoS citation and Dimensions citations. In addition, a poor correlation was demonstrated between the number of citations in WoS and AAS (r = 0.280, p = 0.220).
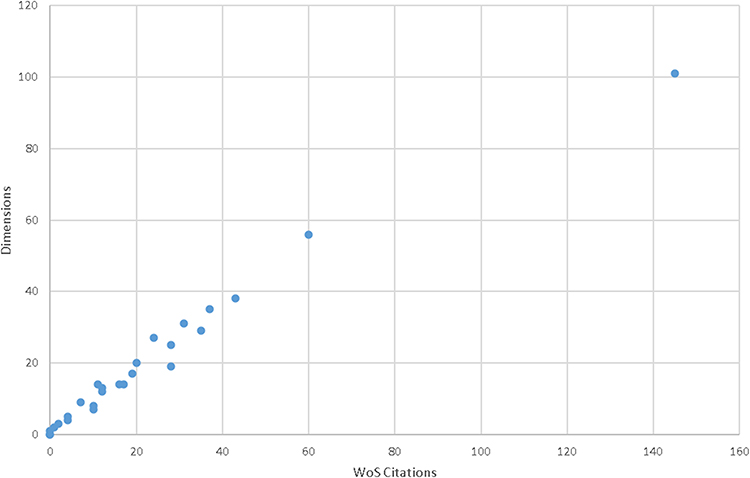 |
Figure 4 Scatter plots showing association of WoS and Dimensions citations. |
Discussion
The expanding comprehension of stem cell biology has underscored the potential of stem cells as formidable therapeutic agents for IDD.5,9,15,16,37 Among these, CESCs are regarded as a promising cellular resource for IDD. However, the biological characteristics of CESCs have not been extensively investigated, and their potential applications in disc regeneration warrant further exploration. To our knowledge, this study is the first to concentrate on the biological attributes and potential uses of CESCs.
CESCs were first identified by Liu et al in 2011.33 They discovered a cell population within the degenerated human CE of intervertebral discs that exhibited stem cell properties, including the capacity for adipogenic, osteogenic, and chondrogenic differentiation.33 While CE cells are differentiated cells within the CE, CESCs are stem cells characterized by their self-renewal ability and differentiation potential.9,10 CESCs have the potential to replenish the depleting differentiated cell population in the CE.9,10,14 However, it is crucial to note that not all cells isolated from the cartilage endplate are stem cells. CESCs meet the criteria for the definition of MSCs, as defined by the International Society for Cellular Therapy (ISCT), and should be sorted into MSCs.78 According to ISCT, MSCs must adhere to plastic in standard culture conditions, express specific surface markers (CD105, CD73, and CD90), lack the expression of others (CD45, CD34, CD14, CD11b, CD79a, CD19, and HLA-DR), and differentiate into osteoblasts, adipocytes, and chondroblasts in vitro. As endogenous stem cells, CESCs play an integral role in maintaining a dynamic equilibrium between tissue repair and damage, modulating their proliferation and differentiation. Furthermore, Huang et al revealed the existence of progenitor cells in degenerated human CE.34 These cells expressed stem cell-associated genes such as OCT-4, NANOG, and SOX-2. These findings provide substantial evidence of CESCs in the CE. Subsequent research has identified CESCs in other species, including rats70,71,73–75 and rabbits,76 further broadening the scope of knowledge in this field.
The discovery of resident stem cells has underscored the importance of their successful isolation. For the isolation of cartilage endplate-derived stem cells (CESCs), several methodologies have been developed. Initially, Liu et al introduced the agarose suspension culture system for this purpose.33 This method has been widely adopted in subsequent studies due to its efficacy in selectively culturing chondrocytes.79 Additionally, some researchers have utilized the differential adhesion method for CESCs isolation, leveraging the inherent properties of stem cells such as plastic adherence and rapid proliferation.65,70 This technique takes advantage of the unique characteristics of stem cells to segregate CESCs from other cell types in the intervertebral disc. Another distinctive feature of stem cells is colony formation, which can be exploited for CESCs isolation through low-density plating,62 based on the premise that stem cells can thrive at lower seeding densities compared to other cells.10,80 Moreover, several studies have reported the direct isolation of CESCs using standard cell culture techniques without specific interventions.71–75 Despite these varied techniques, a consensus on the optimal method for CESCs isolation remains elusive, particularly in the absence of comparative studies. Therefore, further investigation is required to develop strategies for the simple and economical isolation of pure CESCs.10 However, once cells are confirmed as CESCs, the method of isolation may be considered of secondary importance.
Regarding surface markers, CESCs share similarities with MSCs, such as the expression of CD73, CD90, and CD105, and the absence of CD11b, CD14, CD19, CD34, CD45, or HLA-DR, as per the criteria established by the ISCT.33,34,78 However, the presence of these markers does not unequivocally identify a cell as a CESC,9,32 as intervertebral disc cells, including AF and NP cells, also exhibit MSC-like surface markers.81,82 To date, no standardized consensus exists regarding specific surface markers for CESCs,5,9,10,32 complicating their differentiation from other cells with similar marker profiles.9,32 Consequently, there is a pressing need to identify unique surface markers for CESCs. Single-cell sequencing is emerging as a potent technique for this purpose, offering the ability to precisely identify distinct CESCs populations at the individual cell level.9,52
Advancements in research concerning cartilage endplate-derived stem cells (CESCs) have led to a thorough understanding of their biological properties. Comparative analyses have shown that CESCs possess superior chondrogenic and osteogenic potentials compared to NPSCs, AFSCs, and BMSCs.55,58 Liu et al62 conducted a comparative study of CESCs, NPSCs, and AFSCs, revealing that CESCs exhibit enhanced migratory and invasive capabilities. These findings suggest the potential of CESCs as effective cellular sources for stem cell therapy and as seed cells for tissue engineering in the context of IDD.5,9,10 However, conflicting results have been observed in other studies. Liang et al65 reported that AFSCs exhibited the most pronounced stem cell-like characteristics, particularly in terms of cell proliferation and trilineage differentiation capabilities. Such discrepancies in biological properties across different studies could be ascribed to variations in species, isolation techniques, passaging, and culture methods.5,9,10,55,58,62,65 Moreover, it is noteworthy that most studies on CESCs have been conducted in vitro, raising questions about the translatability of these findings to in vivo contexts.5,9,10 Therefore, there is a pressing need for additional in vivo research to elucidate the biological characteristics of CESCs more accurately. Such studies would be instrumental in validating the potential applications of CESCs in the treatment of IDD.
Recent advancements in genome-wide analyses have facilitated in-depth investigation into CESCs. Given the CE’s role as a primary channel for nutrient supply, its degeneration, primarily characterized by ossification rather than chondrification, is implicated in the initiation and progression of IDD.20,21,23,75,83,84 CESCs, with their inherent chondro-osteogenic differentiation capabilities, may play a crucial role in maintaining the balance between chondrification and ossification in the CE.33,66 Alternative splicing, a ubiquitous and essential mechanism in eukaryotic cells, leads to gene isoform diversity and protein complexity. Understanding the mechanisms of alternative splicing during stem cell differentiation is vital. High-throughput screening has become the primary method for analyzing transcription profiles and alternative splicing events. Shang et al conducted genome-wide analyses of gene expression profiling and alternative splicing during chondrogenic and osteogenic differentiation of CESCs, identifying various genes with distinct expression and splicing patterns and investigating related molecular functions and pathways.56,60 Moreover, hypoxia is known to regulate stem cell proliferation and multi-differentiation potential, suggesting its significant role in modulating CESCs’ biological activities.85,86 Yao et al investigated the regulatory effects of hypoxia on CESCs, including differential gene expression and alternative splicing during hypoxia-regulated chondrogenic and osteogenic differentiation.59,64,66 Overall, these investigations indicate a strong association between alternative splicing events and the regulatory mechanisms of CESCs. Genome-wide analyses, therefore, could provide a valuable reference framework for future studies aimed at delineating the biological functions of CESCs. These analyses are instrumental in enhancing our understanding of the underlying mechanisms of IDD and in aiding the development of novel therapeutic approaches for IDD regeneration. This insight into the molecular intricacies of CESCs could pave the way for innovative strategies in regenerative medicine, potentially transforming the current paradigm of IDD treatment.
CESCs, as resident endogenous stem cells, are pivotal in the context of IDD.5,41 However, the mechanisms underlying the compromise of CESCs during IDD remain elusive. One hypothesis posits that CESCs may undergo exhaustion following repeated cycles of proliferation and differentiation in an attempt to regenerate the IDD-affected disc. Alternatively, the adverse microenvironmental conditions prevalent in intervertebral discs during IDD could detrimentally impact CESC function.5,32 This microenvironment is characterized by hypoxia, acidic pH, high mechanical stress, low glucose concentration, hypertonicity, and suboptimal nutrition supply.10,12,14,18,87,88 Such environmental stressors could disrupt the physiological milieu of CESCs, thereby adversely affecting their activity and challenging their survival.9,10,14,40,44 This understanding highlights the need for further research into the resilience and adaptability of CESCs within the challenging microenvironment of the degenerated intervertebral disc.
Recent research has extensively explored the fate of CESCs in the context of IDD.54,57,61,63,68–70,76 Yao et al discovered that normoxia promotes osteogenesis but inhibits chondrogenesis in CESCs.61 Additionally, they identified the macrophage migration inhibitory factor (MIF) as a key nuclear transcriptional regulator influencing the chondrogenic and osteogenic differentiation of CESCs under hypoxic conditions. The correlation between IDD and inflammatory responses has also been a focal point of investigation.57,63,68,69,89,90 Xiong et al reported that MIF, a proinflammatory cytokine, was expressed in degenerated discs and hinders the migration of CESCs to inflammatory sites, potentially exacerbating IDD.54 Further studies have underscored the impact of inflammatory cytokines on CESCs. Zuo et al demonstrated that tumor necrosis factor (TNF)-α reduces the proliferation, senescence, and osteogenic differentiation of CESCs.70 They also found that rapamycin-induced autophagy mitigates inflammation-mediated cartilage endplate degeneration by enhancing nuclear factor erythroid 2-related factor 2 (Nrf2)/Kelch-like ECH-associated protein 1 (Keap1) signaling in CESCs. Feng et al revealed that N-acetylated proline-glycine-proline (N-Ac-PGP), produced by NP cells within the complex microenvironment of the degenerated disc, prompts CESCs to migrate from the CE into the NP and differentiate into a pro-inflammatory phenotype, accelerating IDD progression.57 Yuan et al elucidated that cyclic tensile stress induces CESC apoptosis through the Bcl-2/adenovirus E1B 19-kDa-interacting protein 3 (BNIP3)/Bcl-2 pathway.68 Moreover, Zhang et al observed that tension load decreases cell viability, enhances apoptosis, and reduces the expression of ACAN, COL-2A, and SOX9 in CESCs, indicative of CESC degeneration. They also showed that inhibiting c-Jun N-terminal kinase (JNK) and extracellular signal-regulated kinase (ERK) within the MAPK signaling family can alleviate tension load-induced CESC degeneration by suppressing the phosphorylation of Raptor and mammalian target of rapamycin (mTOR) in the mTOR pathway.76 He et al demonstrated that nutritional deficiency leads to caspase-dependent apoptosis of CESCs via the BNIP3-related pathway.63 Furthermore, Sun et al found that high glucose levels favor osteogenic over chondrogenic differentiation of CESCs, leading to CE calcification in IDD,69 possibly due to the intrusion of blood vessels into CE during IDD and the subsequent disruption of the low-glucose microenvironment of CESCs.91,92 Overall, the studies underscore that the microenvironment of degenerated discs significantly influences the viability of CESCs, affecting their quantity, quality, and overall fate. These findings highlight the need for a deeper understanding of potential interventions that can enhance the beneficial effects on CESCs and mitigate the negative impacts, thereby aiding in the development of effective regeneration strategies for IDD.
Current therapeutic approaches for IDD predominantly focus on symptom management and palliation.12,14,35,88 Notably, these treatments do not address the fundamental etiology of IDD.9,10,26,45 Understanding the mechanisms underlying the degeneration of CESCs could be instrumental in both preventing IDD and developing targeted therapeutic strategies. Recent research efforts have been directed towards elucidating the impact of CESCs on IDD pathogenesis. Chen et al discovered that miR-637 is downregulated in degenerated CESCs, and they posited that overexpression of miR-637 might inhibit the osteogenic differentiation of CESCs by targeting the WNT5A gene.77 Similarly, Guan et al identified that overexpression of carbohydrate sulfotransferase 3 (CHST3) enhances the proliferation and migration capabilities of CESCs.72 Insights gleaned from these studies offer a promising avenue for identifying potential molecular targets, which could inform the development of novel, more effective treatments for IDD. These advances in understanding the molecular dynamics of CESCs contribute significantly to the evolving landscape of IDD research, potentially leading to breakthroughs in regenerative medicine and therapeutics.
In the rapidly evolving domain of biological therapy, there is an escalating interest in developing novel interventions for IDD that aim to limit, arrest, or potentially reverse the condition, while restoring the biological functions of intervertebral discs.13,39,45,93–96 The field of regenerative medicine offers promising biological repair strategies for degenerated discs.13,39,45,93–95 Pioneering this approach, Sakai et al in 2003 investigated the feasibility of MSCs transplantation in IDD using a rabbit model.97,98 This was followed by Haufe and Mork’s 2006 groundbreaking human trial involving intradiscal injections of hematopoietic stem cells.99,100 In the realm of CESCs, the application of chemokines and exosomes for recruiting endogenous stem cells or stimulating endogenous repair processes has gained significant traction.9,10,32,41,44 He et al demonstrated that CESCs enhance the proliferation of NP cells through paracrine interactions, particularly via the stromal cell-derived factor 1 (SDF-1)/chemokine receptor type 4 (CXCR4) axis.67 This suggests the potential of the SDF-1/CXCR4 axis to augment the recruitment and interaction of endogenous stem cells with NP cells, thereby contributing to the regeneration of degenerated discs. Moreover, exosomes, as potent mediators of intercellular communication derived from stem cells, have exhibited remarkable regenerative capabilities for tissues and organs.96 Luo et al conducted studies to assess the therapeutic effects of CESCs-derived exosomes on IDD.73 They found that exosomes from normal CESCs were more effective in inhibiting NP cell apoptosis and mitigating IDD than those from degenerated CESCs, primarily by enhancing autophagy through the PI3K/AKT signaling pathway. Further exploration in a rat model of IDD revealed that CESCs-derived exosomes could activate HIF-1a/Wnt signaling, promoting CESCs migration into the intervertebral disc and their transformation into NP cells, thus alleviating disc disease.71 Additionally, Chen et al explored the molecular mechanisms through which CESCs-derived exosomes impact NP cell function.74 They discovered that miR-125-5p, secreted by CESCs-derived exosomes, facilitates NP cell autophagy, inhibits apoptosis and extracellular matrix degradation, and attenuates IDD by targeting SUV39H1. These findings underscore the significant potential of CESCs-derived exosomes in IDD therapy and highlight the need for further research to optimize their application in clinical settings.
Tissue engineering in the context of IDD provides innovative approaches for functional repair. Wang et al established stem cell-alginate constructs for NP tissue engineering, evaluating the regenerative potential of four different stem cell types, including CESCs, NPSCs, AFSCs, and BMSCs.55 Their findings revealed that alginate constructs seeded with CESCs demonstrated superior regenerative capabilities for IDD, while AFSCs exhibited the least regenerative capacity. NPSCs and BM-MSCs displayed intermediate potential.55 This evidence positions CESCs as a promising candidate for seeding in intervertebral disc tissue engineering, offering an effective biological approach to IDD treatment. Furthermore, Luo et al investigated an innovative noninvasive strategy involving the injection of an injectable hydrogel combined with lentivirus-engineered CESCs adjacent to the CE.75 This study confirmed the sustained release of functional exosomes from the hydrogel, loaded with CESCs expressing sphingosine kinase 2 (Sphk2). These exosomes penetrated the AF, delivering Sphk2 to NP cells and activating the PI3K/AKT signaling pathway and intracellular autophagy, thereby contributing to the alleviation of IDD.75 Despite these promising advancements, the application of CESCs in regenerative strategies for IDD is still in its infancy. Several challenges remain, such as limited understanding of the biological characteristics of CESCs, the complex pathophysiology of IDD, and the need for scaffolds with optimal biocompatibility, injectability, and sustained-release capabilities.5,8–10,14,88 Therefore, further research is imperative to explore endogenous repair mechanisms for IDD. Focusing on endogenous repair could potentially overcome several limitations associated with exogenous therapeutic approaches, paving the way for more effective and less invasive treatments for IDD.8,9,14,32,88,94
This study possesses certain limitations. Primarily, the sample size was relatively limited, both in terms of the number of studies reviewed and the animal samples considered within these studies. Despite this constraint, the study offers valuable insights into the research surrounding CESCs and their role in the regeneration of IDD since their initial discovery. Furthermore, although a comprehensive search strategy was employed, encompassing a review of references in the included papers and related review articles to identify pertinent literature on CESCs, it is important to acknowledge that no search strategy is flawless. Consequently, the possibility of selection bias in the study cannot be completely eliminated.
Conclusion
This study represents the inaugural comprehensive analysis of the biological characteristics of CESCs and evaluates their prospective utility in the regeneration of intervertebral discs. CESCs appear to be promising candidates for mitigating degeneration and fostering regeneration within the intervertebral disc. However, current research is limited by the absence of specific surface markers and purification methodologies for CESCs. Furthermore, there is no established consensus on the most effective approach for employing CESCs in the treatment of IDD. Despite these limitations, CESCs have emerged as a compelling cellular resource for IDD regeneration. Future investigations should focus on further elucidating the underlying biological mechanisms of CESCs and refining their therapeutic potential.
Data Sharing Statement
The datasets used and/or analyzed during the present study are available from the corresponding author on reasonable request.
Ethics Approval and Informed Consent
No approval of Institutional Reviewed Board was needed due to not involving human and animals in this study.
Funding
This work was supported by the National Natural Science Foundation of China (82074452, 81601948).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Andersson GB. Epidemiological features of chronic low-back pain. Lancet. 1999;354(9178):581–585. doi:10.1016/S0140-6736(99)01312-4
2. Vos T, Flaxman AD, Naghavi M, et al. Years lived with disability (YLDs) for 1160 sequelae of 289 diseases and injuries 1990–2010: a systematic analysis for the Global Burden of Disease Study 2010. Lancet. 2012;380(9859):2163–2196. doi:10.1016/S0140-6736(12)61729-2
3. Walker BF. The prevalence of low back pain: a systematic review of the literature from 1966 to 1998. J Spinal Disord. 2000;13(3):205–217. doi:10.1097/00002517-200006000-00003
4. Hoy D, March L, Brooks P, et al. The global burden of low back pain: estimates from the Global Burden of Disease 2010 study. Ann Rheum Dis. 2014;73(6):968–974. doi:10.1136/annrheumdis-2013-204428
5. Lyu FJ, Cheung KM, Zheng Z, Wang H, Sakai D, Leung VY. IVD progenitor cells: a new horizon for understanding disc homeostasis and repair. Nat Rev Rheumatol. 2019;15(2):102–112. doi:10.1038/s41584-018-0154-x
6. Roh JS, Teng AL, Yoo JU, Davis J, Furey C, Bohlman HH. Degenerative disorders of the lumbar and cervical spine. Orthop Clin North Am. 2005;36(3):255–262. doi:10.1016/j.ocl.2005.01.007
7. Dagenais S, Caro J, Haldeman S. A systematic review of low back pain cost of illness studies in the United States and internationally. Spine J. 2008;8(1):8–20. doi:10.1016/j.spinee.2007.10.005
8. Liu Y, Li Y, Nan LP, et al. Insights of stem cell-based endogenous repair of intervertebral disc degeneration. World J Stem Cells. 2020;12(4):266–276. doi:10.4252/wjsc.v12.i4.266
9. Du Y, Wang Z, Wu Y, Liu C, Zhang L, Hu B. Intervertebral disc stem/progenitor cells: a promising “Seed” for intervertebral disc regeneration. Stem Cells Int. 2021;2021:2130727. doi:10.1155/2021/2130727
10. Hu B, He R, Ma K, et al. Intervertebral disc-derived stem/progenitor cells as a promising cell source for intervertebral disc regeneration. Stem Cells Int. 2018;2018:7412304. doi:10.1155/2018/7412304
11. Hadjipavlou AG, Tzermiadianos MN, Bogduk N, Zindrick MR. The pathophysiology of disc degeneration: a critical review. J Bone Joint Surg Br. 2008;90(10):1261–1270. doi:10.1302/0301-620X.90B10.20910
12. Adams MA, Roughley PJ. What is intervertebral disc degeneration, and what causes it? Spine. 2006;31(18):2151–2161. doi:10.1097/01.brs.0000231761.73859.2c
13. Li B, Yang Y, Wang L, Liu G, Hu B. Stem cell therapy and exercise for treatment of intervertebral disc degeneration. Stem Cells Int. 2021;2021:7982333. doi:10.1155/2021/7982333
14. Sakai D, Andersson GB. Stem cell therapy for intervertebral disc regeneration: obstacles and solutions. Nat Rev Rheumatol. 2015;11(4):243–256. doi:10.1038/nrrheum.2015.13
15. Huang S, Tam V, Cheung KM, et al. Stem cell-based approaches for intervertebral disc regeneration. Curr Stem Cell Res Ther. 2011;6(4):317–326. doi:10.2174/157488811797904335
16. Vadala G, Ambrosio L, Russo F, Papalia R, Denaro V. Stem cells and intervertebral disc regeneration overview-what they can and can’t do. Int J Spine Surg. 2021;15(s1):40–53. doi:10.14444/8054
17. Magnier C, Boiron O, Wendling-Mansuy S, Chabrand P, Deplano V. Nutrient distribution and metabolism in the intervertebral disc in the unloaded state: a parametric study. J Biomech. 2009;42(2):100–108. doi:10.1016/j.jbiomech.2008.10.034
18. Huang YC, Urban JP, Luk KD. Intervertebral disc regeneration: do nutrients lead the way? Nat Rev Rheumatol. 2014;10(9):561–566. doi:10.1038/nrrheum.2014.91
19. Urban JP, Smith S, Fairbank JC. Nutrition of the intervertebral disc. Spine. 2004;29(23):2700–2709. doi:10.1097/01.brs.0000146499.97948.52
20. Peng B, Hou S, Shi Q, Jia L. The relationship between cartilage end-plate calcification and disc degeneration: an experimental study. Chin Med J. 2001;114(3):308–312.
21. Roberts S, Urban JP, Evans H, Eisenstein SM. Transport properties of the human cartilage endplate in relation to its composition and calcification. Spine. 1996;21(4):415–420. doi:10.1097/00007632-199602150-00003
22. Benneker LM, Heini PF, Alini M, Anderson SE, Ito K. 2004 Young Investigator Award Winner: vertebral endplate marrow contact channel occlusions and intervertebral disc degeneration. Spine. 2005;30(2):167–173. doi:10.1097/01.brs.0000150833.93248.09
23. Rajasekaran S, Venkatadass K, Naresh Babu J, Ganesh K, Shetty AP. Pharmacological enhancement of disc diffusion and differentiation of healthy, ageing and degenerated discs: results from in-vivo serial post-contrast MRI studies in 365 human lumbar discs. Eur Spine J. 2008;17(5):626–643. doi:10.1007/s00586-008-0645-6
24. Ariga K, Miyamoto S, Nakase T, et al. The relationship between apoptosis of endplate chondrocytes and aging and degeneration of the intervertebral disc. Spine. 2001;26(22):2414–2420. doi:10.1097/00007632-200111150-00004
25. Rajasekaran S, Babu JN, Arun R, Armstrong BR, Shetty AP, Murugan S. ISSLS prize winner: a study of diffusion in human lumbar discs: a serial magnetic resonance imaging study documenting the influence of the endplate on diffusion in normal and degenerate discs. Spine. 2004;29(23):2654–2667. doi:10.1097/01.brs.0000148014.15210.64
26. de Sousa EB, Casado PL, Moura Neto V, Duarte ME, Aguiar DP. Synovial fluid and synovial membrane mesenchymal stem cells: latest discoveries and therapeutic perspectives. Stem Cell Res Ther. 2014;5(5):112. doi:10.1186/scrt501
27. Jiang D, Yang S, Gao P, et al. Combined effect of ligament stem cells and umbilical-cord-blood-derived CD34+ cells on ligament healing. Cell Tissue Res. 2015;362(3):587–595. doi:10.1007/s00441-015-2250-4
28. Ito Y, Fitzsimmons JS, Sanyal A, Mello MA, Mukherjee N, O’Driscoll SW. Localization of chondrocyte precursors in periosteum. Osteoarthritis Cartilage. 2001;9(3):215–223. doi:10.1053/joca.2000.0378
29. Debnath S, Yallowitz AR, McCormick J, et al. Discovery of a periosteal stem cell mediating intramembranous bone formation. Nature. 2018;562(7725):133–139. doi:10.1038/s41586-018-0554-8
30. Williams R, Khan IM, Richardson K, et al. Identification and clonal characterisation of a progenitor cell sub-population in normal human articular cartilage. PLoS One. 2010;5(10):e13246. doi:10.1371/journal.pone.0013246
31. Yin H, Price F, Rudnicki MA. Satellite cells and the muscle stem cell niche. Physiol Rev. 2013;93(1):23–67. doi:10.1152/physrev.00043.2011
32. Zhang Y, Hu Y, Wang W, et al. Current Progress in the Endogenous Repair of Intervertebral Disk Degeneration Based on Progenitor Cells. Front Bioeng Biotechnol. 2020;8:629088. doi:10.3389/fbioe.2020.629088
33. Liu LT, Huang B, Li CQ, Zhuang Y, Wang J, Zhou Y. Characteristics of stem cells derived from the degenerated human intervertebral disc cartilage endplate. PLoS One. 2011;6(10):e26285. doi:10.1371/journal.pone.0026285
34. Huang B, Liu LT, Li CQ, et al. Study to determine the presence of progenitor cells in the degenerated human cartilage endplates. Eur Spine J. 2012;21(4):613–622. doi:10.1007/s00586-011-2039-4
35. Chen BL, Guo JB, Zhang HW, et al. Surgical versus non-operative treatment for lumbar disc herniation: a systematic review and meta-analysis. Clin Rehabil. 2018;32(2):146–160. doi:10.1177/0269215517719952
36. Meisel HJ, Agarwal N, Hsieh PC, et al. Cell therapy for treatment of intervertebral disc degeneration: a systematic review. Global Spine J. 2019;9(1 Suppl):39S–52S. doi:10.1177/2192568219829024
37. Gou S, Oxentenko SC, Eldrige JS, et al. Stem cell therapy for intervertebral disk regeneration. Am J Phys Med Rehabil. 2014;93(11 Suppl 3):S122–S131. doi:10.1097/PHM.0000000000000152
38. Epstein NE. Adjacent level disease following lumbar spine surgery: a review. Surg Neurol Int. 2015;6(Suppl 24):S591–S599. doi:10.4103/2152-7806.170432
39. Krut Z, Pelled G, Gazit D, Gazit Z. Stem cells and exosomes: new therapies for intervertebral disc degeneration. Cells. 2021;10(9):2241. doi:10.3390/cells10092241
40. Binch ALA, Fitzgerald JC, Growney EA, Barry F. Cell-based strategies for IVD repair: clinical progress and translational obstacles. Nat Rev Rheumatol. 2021;17(3):158–175. doi:10.1038/s41584-020-00568-w
41. Ma K, Chen S, Li Z, et al. Mechanisms of endogenous repair failure during intervertebral disc degeneration. Osteoarthritis Cartilage. 2019;27(1):41–48. doi:10.1016/j.joca.2018.08.021
42. Wu H, Zeng X, Yu J, et al. Comparison of nucleus pulposus stem/progenitor cells isolated from degenerated intervertebral discs with umbilical cord derived mesenchymal stem cells. Exp Cell Res. 2017;361(2):324–332. doi:10.1016/j.yexcr.2017.10.034
43. Johnson WE, Stephan S, Roberts S. The influence of serum, glucose and oxygen on intervertebral disc cell growth in vitro: implications for degenerative disc disease. Arthritis Res Ther. 2008;10(2):R46. doi:10.1186/ar2405
44. Lee CH, Lee FY, Tarafder S, et al. Harnessing endogenous stem/progenitor cells for tendon regeneration. J Clin Invest. 2015;125(7):2690–2701. doi:10.1172/JCI81589
45. Clouet J, Fusellier M, Camus A, Le Visage C, Guicheux J. Intervertebral disc regeneration: from cell therapy to the development of novel bioinspired endogenous repair strategies. Adv Drug Deliv Rev. 2019;146:306–324. doi:10.1016/j.addr.2018.04.017
46. Zhang W, Sun T, Li Y, et al. Application of stem cells in the repair of intervertebral disc degeneration. Stem Cell Res Ther. 2022;13(1):70. doi:10.1186/s13287-022-02745-y
47. Wells K, Littell JH. Study quality assessment in systematic reviews of research on intervention effects. Res Soc Work Pract. 2009;19(1):52–62. doi:10.1177/1049731508317278
48. Shin RL, Lee CW, Shen OY, Xu H, Lee OK, Jones EA. The crosstalk between mesenchymal stem cells and macrophages in bone regeneration: a systematic review. Stem Cells Int. 2021;2021:8835156. doi:10.1155/2021/8835156
49. Lin Y, Tang Z, Jin L, Yang Y. The expression and regulatory roles of long non-coding RNAs in periodontal ligament cells: a systematic review. Biomolecules. 2022;12(2):304. doi:10.3390/biom12020304
50. Xu H, Lee CW, Wang YF, et al. The role of paracrine regulation of mesenchymal stem cells in the crosstalk with macrophages in musculoskeletal diseases: a systematic review. Front Bioeng Biotechnol. 2020;8:587052. doi:10.3389/fbioe.2020.587052
51. Xu HT, Lee CW, Li MY, Wang YF, Yung PS, Lee OK. The shift in macrophages polarisation after tendon injury: a systematic review. J Orthop Translat. 2020;21:24–34. doi:10.1016/j.jot.2019.11.009
52. Martelli AJ, Machado RA, Martelli DRB, Neves LTD, Martelli Junior H. The 100 most-cited papers in oral medicine and pathology. Braz Oral Res. 2020;35:e020.
53. Garcovich D, Marques Martinez L, Adobes Martin M. Citation classics in paediatric dentistry: a bibliometric study on the 100 most-cited articles. Eur Arch Paediatr Dent. 2020;21(2):249–261. doi:10.1007/s40368-019-00483-z
54. Xiong CJ, Huang B, Zhou Y, et al. Macrophage migration inhibitory factor inhibits the migration of cartilage end plate-derived stem cells by reacting with CD74. PLoS One. 2012;7(8):e43984. doi:10.1371/annotation/a5edef40-e46d-4810-9008-dbda429ccc2c
55. Wang H, Zhou Y, Huang B, et al. Utilization of stem cells in alginate for nucleus pulposus tissue engineering. Tissue Eng Part A. 2014;20(5–6):908–920. doi:10.1089/ten.tea.2012.0703
56. Shang J, Fan X, Shangguan L, Liu H, Zhou Y. Global gene expression profiling and alternative splicing events during the chondrogenic differentiation of human cartilage endplate-derived stem cells. Biomed Res Int. 2015;2015:604972. doi:10.1155/2015/604972
57. Feng C, Zhang Y, Yang M, Huang B, Zhou Y. Collagen-derived N-acetylated proline-glycine-proline in intervertebral discs modulates CXCR1/2 expression and activation in cartilage endplate stem cells to induce migration and differentiation toward a pro-inflammatory phenotype. Stem Cells. 2015;33(12):3558–3568. doi:10.1002/stem.2200
58. Wang H, Zhou Y, Chu TW, et al. Distinguishing characteristics of stem cells derived from different anatomical regions of human degenerated intervertebral discs. Eur Spine J. 2016;25(9):2691–2704. doi:10.1007/s00586-016-4522-4
59. Yao Y, Shang J, Song W, Deng Q, Liu H, Zhou Y. Global profiling of the gene expression and alternative splicing events during hypoxia-regulated chondrogenic differentiation in human cartilage endplate-derived stem cells. Genomics. 2016;107(5):170–177. doi:10.1016/j.ygeno.2016.03.003
60. Shang J, Wang H, Fan X, Shangguan L, Liu H. A genome wide analysis of alternative splicing events during the osteogenic differentiation of human cartilage endplate-derived stem cells. Mol Med Rep. 2016;14(2):1389–1396. doi:10.3892/mmr.2016.5359
61. Yao Y, Deng Q, Song W, et al. MIF plays a key role in regulating tissue-specific chondro-osteogenic differentiation fate of human cartilage endplate stem cells under hypoxia. Stem Cell Rep. 2016;7(2):249–262. doi:10.1016/j.stemcr.2016.07.003
62. Liu S, Liang H, Lee SM, Li Z, Zhang J, Fei Q. Isolation and identification of stem cells from degenerated human intervertebral discs and their migration characteristics. Acta Biochim Biophys Sin. 2017;49(2):101–109. doi:10.1093/abbs/gmw121
63. He Z, Pu L, Yuan C, Jia M, Wang J. Nutrition deficiency promotes apoptosis of cartilage endplate stem cells in a caspase-independent manner partially through upregulating BNIP3. Acta Biochim Biophys Sin. 2017;49(1):25–32. doi:10.1093/abbs/gmw111
64. Yao Y, Deng Q, Sun C, Song W, Liu H, Zhou Y. A genome-wide analysis of the gene expression profiles and alternative splicing events during the hypoxia-regulated osteogenic differentiation of human cartilage endplate-derived stem cells. Mol Med Rep. 2017;16(2):1991–2001. doi:10.3892/mmr.2017.6846
65. Liang L, Li X, Li D, et al. The characteristics of stem cells in human degenerative intervertebral disc. Medicine. 2017;96(25):e7178. doi:10.1097/MD.0000000000007178
66. Yao Y, Song W, Deng Q, et al. General regulatory effects of hypoxia on human cartilage endplate-derived stem cells: a genomewide analysis of differential gene expression and alternative splicing events. Mol Med Rep. 2017;16(3):3001–3009. doi:10.3892/mmr.2017.6907
67. He Z, Jia M, Yu Y, Yuan C, Wang J. Roles of SDF-1/CXCR4 axis in cartilage endplate stem cells mediated promotion of nucleus pulposus cells proliferation. Biochem Biophys Res Commun. 2018;506(1):94–101. doi:10.1016/j.bbrc.2018.10.069
68. Yuan C, Pu L, He Z, Wang J. BNIP3/Bcl-2-mediated apoptosis induced by cyclic tensile stretch in human cartilage endplate-derived stem cells. Exp Ther Med. 2018;15(1):235–241. doi:10.3892/etm.2017.5372
69. Sun C, Lan W, Li B, et al. Glucose regulates tissue-specific chondro-osteogenic differentiation of human cartilage endplate stem cells via O-GlcNAcylation of Sox9 and Runx2. Stem Cell Res Ther. 2019;10(1):357. doi:10.1186/s13287-019-1440-5
70. Zuo R, Wang Y, Li J, et al. Rapamycin induced autophagy inhibits inflammation-mediated endplate degeneration by enhancing Nrf2/Keap1 signaling of cartilage endplate stem cells. Stem Cells. 2019;37(6):828–840. doi:10.1002/stem.2999
71. Luo L, Gong J, Zhang H, et al. Cartilage endplate stem cells transdifferentiate into nucleus pulposus cells via autocrine exosomes. Front Cell Dev Biol. 2021;9:648201. doi:10.3389/fcell.2021.648201
72. Guan Y, Sun C, Zou F, et al. Carbohydrate sulfotransferase 3 (CHST3) overexpression promotes cartilage endplate-derived stem cells (CESCs) to regulate molecular mechanisms related to repair of intervertebral disc degeneration by rat nucleus pulposus. J Cell Mol Med. 2021;25(13):6006–6017. doi:10.1111/jcmm.16440
73. Luo L, Jian X, Sun H, et al. Cartilage endplate stem cells inhibit intervertebral disc degeneration by releasing exosomes to nucleus pulposus cells to activate Akt/autophagy. Stem Cells. 2021;39(4):467–481. doi:10.1002/stem.3322
74. Chen D, Jiang X. Exosomes-derived miR-125-5p from cartilage endplate stem cells regulates autophagy and ECM metabolism in nucleus pulposus by targeting SUV38H1. Exp Cell Res. 2022;414(1):113066. doi:10.1016/j.yexcr.2022.113066
75. Luo L, Gong J, Wang Z, et al. Injectable cartilage matrix hydrogel loaded with cartilage endplate stem cells engineered to release exosomes for non-invasive treatment of intervertebral disc degeneration. Bioact Mater. 2022;15:29–43. doi:10.1016/j.bioactmat.2021.12.007
76. Zhang Y, Liu C, Li Y, Xu H. Mechanism of the mitogen-activated protein kinases/mammalian target of rapamycin pathway in the process of cartilage endplate stem cell degeneration induced by tension load. Global Spine J. 2023;13(8):2396–2408. doi:10.1177/21925682221085226
77. Chen Y, Chen Q, Zhong M, Xu C, Wu Y, Chen R. miR-637 inhibits osteogenic differentiation of human intervertebral disc cartilage endplate stem cells by targeting WNT5A. J Invest Surg. 2022;35(6):1313–1321. doi:10.1080/08941939.2022.2050857
78. Dominici M, Le Blanc K, Mueller I, et al. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy. 2006;8(4):315–317. doi:10.1080/14653240600855905
79. Lin L, Jia Z, Zhao Y, et al. Use of limiting dilution method for isolation of nucleus pulposus mesenchymal stem/progenitor cells and effects of plating density on biological characteristics and plasticity. Biomed Res Int. 2017;2017:9765843. doi:10.1155/2017/9765843
80. Sekiya I, Larson BL, Smith JR, Pochampally R, Cui JG, Prockop DJ. Expansion of human adult stem cells from bone marrow stroma: conditions that maximize the yields of early progenitors and evaluate their quality. Stem Cells. 2002;20(6):530–541. doi:10.1634/stemcells.20-6-530
81. Feng G, Yang X, Shang H, et al. Multipotential differentiation of human anulus fibrosus cells: an in vitro study. J Bone Joint Surg Am. 2010;92(3):675–685. doi:10.2106/JBJS.H.01672
82. Sakai D, Nakamura Y, Nakai T, et al. Exhaustion of nucleus pulposus progenitor cells with ageing and degeneration of the intervertebral disc. Nat Commun. 2012;3:1264. doi:10.1038/ncomms2226
83. Li FC, Zhang N, Chen WS, Chen QX. Endplate degeneration may be the origination of the vacuum phenomenon in intervertebral discs. Med Hypotheses. 2010;75(2):169–171. doi:10.1016/j.mehy.2010.02.012
84. Antoniou J, Goudsouzian NM, Heathfield TF, et al. The human lumbar endplate. Evidence of changes in biosynthesis and denaturation of the extracellular matrix with growth, maturation, aging, and degeneration. Spine. 1996;21(10):1153–1161. doi:10.1097/00007632-199605150-00006
85. Zhu LL, Wu LY, Yew DT, Fan M. Effects of hypoxia on the proliferation and differentiation of NSCs. Mol Neurobiol. 2005;31(1–3):231–242.
86. Chen W, Zhuo Y, Duan D, Lu M. Effects of hypoxia on differentiation of mesenchymal stem cells. Curr Stem Cell Res Ther. 2020;15(4):332–339. doi:10.2174/1574888X14666190823144928
87. Vadala G, Ambrosio L, Russo F, Papalia R, Denaro V. Interaction between mesenchymal stem cells and intervertebral disc microenvironment: from cell therapy to tissue engineering. Stem Cells Int. 2019;2019:2376172. doi:10.1155/2019/2376172
88. Wang F, Shi R, Cai F, Wang YT, Wu XT. Stem cell approaches to intervertebral disc regeneration: obstacles from the disc microenvironment. Stem Cells Dev. 2015;24(21):2479–2495. doi:10.1089/scd.2015.0158
89. Takahashi H, Suguro T, Okazima Y, Motegi M, Okada Y, Kakiuchi T. Inflammatory cytokines in the herniated disc of the lumbar spine. Spine. 1996;21(2):218–224. doi:10.1097/00007632-199601150-00011
90. Shamji MF, Setton LA, Jarvis W, et al. Proinflammatory cytokine expression profile in degenerated and herniated human intervertebral disc tissues. Arthritis Rheum. 2010;62(7):1974–1982. doi:10.1002/art.27444
91. Walsh DA, McWilliams DF, Turley MJ, et al. Angiogenesis and nerve growth factor at the osteochondral junction in rheumatoid arthritis and osteoarthritis. Rheumatology. 2010;49(10):1852–1861. doi:10.1093/rheumatology/keq188
92. Nerlich AG, Schaaf R, Walchli B, Boos N. Temporo-spatial distribution of blood vessels in human lumbar intervertebral discs. Eur Spine J. 2007;16(4):547–555. doi:10.1007/s00586-006-0213-x
93. O’Connell GD, Leach JK, Klineberg EO. Tissue engineering a biological repair strategy for lumbar disc herniation. Biores Open Access. 2015;4(1):431–445. doi:10.1089/biores.2015.0034
94. Buckley CT, Hoyland JA, Fujii K, Pandit A, Iatridis JC, Grad S. Critical aspects and challenges for intervertebral disc repair and regeneration-Harnessing advances in tissue engineering. JOR Spine. 2018;1(3):e1029. doi:10.1002/jsp2.1029
95. Melrose J. Strategies in regenerative medicine for intervertebral disc repair using mesenchymal stem cells and bioscaffolds. Regen Med. 2016;11(7):705–724. doi:10.2217/rme-2016-0069
96. Zhang Y, Bi J, Huang J, Tang Y, Du S, Li P. Exosome: a review of its classification, isolation techniques, storage, diagnostic and targeted therapy applications. Int J Nanomedicine. 2020;15:6917–6934. doi:10.2147/IJN.S264498
97. Sakai D, Mochida J, Yamamoto Y, et al. Transplantation of mesenchymal stem cells embedded in Atelocollagen gel to the intervertebral disc: a potential therapeutic model for disc degeneration. Biomaterials. 2003;24(20):3531–3541. doi:10.1016/S0142-9612(03)00222-9
98. Drazin D, Rosner J, Avalos P, Acosta F. Stem cell therapy for degenerative disc disease. Adv Orthop. 2012;2012:961052. doi:10.1155/2012/961052
99. Haufe SM, Mork AR. Intradiscal injection of hematopoietic stem cells in an attempt to rejuvenate the intervertebral discs. Stem Cells Dev. 2006;15(1):136–137. doi:10.1089/scd.2006.15.136
100. Sakai D, Schol J. Cell therapy for intervertebral disc repair: clinical perspective. J Orthop Translat. 2017;9:8–18. doi:10.1016/j.jot.2017.02.002
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Pyroptosis and Intervertebral Disc Degeneration: Mechanistic Insights and Therapeutic Implications
Ge Y, Chen Y, Guo C, Luo H, Fu F, Ji W, Wu C, Ruan H
Journal of Inflammation Research 2022, 15:5857-5871
Published Date: 17 October 2022
Adipose-Derived Stem Cells (ASCs) for Regeneration of Intervertebral Disc Degeneration: Review Article
Romaniyanto F, Mahyudin F, Prakoeswa CRS, Notobroto HB, Tinduh D, Ausrin R, Rantam FA, Suroto H, Utomo DN, Rhatomy S
Stem Cells and Cloning: Advances and Applications 2022, 15:67-76
Published Date: 4 November 2022
Trends of Low Back Pain Research in Older and Working-Age Adults from 1993 to 2023: A Bibliometric Analysis
Zheng DKY, Kawchuk GN, Bussières AE, Al Zoubi FM, Hartvigsen J, Fu SN, de Luca K, Weiner DK, Karppinen J, Samartzis D, Ferreira ML, Wu J, Dennett L, Wong AY
Journal of Pain Research 2023, 16:3325-3341
Published Date: 2 October 2023
Detrimental Effects of Space Flight on the Lumbar Spine May Be Correlated to Baseline Degeneration: Insights From an Advanced MR Imaging Study
Bokhari R, Bisson DG, Fortin M, Vigouroux M, Cata JP, Hwang KP, Chen MM, Ceniza-Bordallo G, Ouellet JA, Ingelmo PM
Journal of Pain Research 2025, 18:1375-1385
Published Date: 19 March 2025
Impact of Socioeconomic Status on the Risk of Intervertebral Disc Degeneration and Low Back Pain in the European Population and the Mediating Role of Obesity-Related Traits: Evidence from a Mendelian Randomization Study
Long Z, Qin Q
Journal of Pain Research 2025, 18:1627-1641
Published Date: 26 March 2025