")
Back to Journals » Infection and Drug Resistance » Volume 13
Carbapenemase Producing Gram-Negative Bacteria in Tunisia: History of Thirteen Years of Challenge
Authors Dziri O, Dziri R, Ali El Salabi A, Chouchani C
Received 23 April 2020
Accepted for publication 11 June 2020
Published 23 November 2020 Volume 2020:13 Pages 4177—4191
DOI https://doi.org/10.2147/IDR.S259562
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Suresh Antony
Olfa Dziri,1,2 Raoudha Dziri,1 Allaaeddin Ali El Salabi,3,4 Chedly Chouchani1,2
1Laboratory of Microorganisms and Active Biomolecules, Faculty of Sciences of Tunis, University of Tunis El Manar, Tunis, Tunisia; 2Laboratory of Research in Sciences and Technology of Environment, High Institute of Science and Technology of Environment, University of Carthage, Hammam-Lif, Tunisia; 3Department of Environmental Health, Faculty of Public Health, University of Benghazi, Benghazi, Libya; 4Infection Control and Patient Safety Office, New Marwa Hospital, Benghazi, Libya
Correspondence: Chedly Chouchani
ISSTE de Borj-Cedria, Université de Carthage, Hammam-Lif BP-1003, 2050, Tunisia
Tel +216 94114443
Fax +216 79325333
Email [email protected]
Abstract: The wide spread of multidrug-resistant bacteria, particularly carbapenem-resistant Gram-negative bacteria (CR-GNB), constitutes a major public health threat worldwide, owing to the limited therapeutic options. This review will describe and uncover the Tunisian experience in the challenge against carbapenem resistance. Indeed, we illuminate on the dissemination of CR-GNB in different hospitals, animals, and other natural environments in this country. We resumed the different carbapenemase variants detected from various bacterial species and mapped their regional distribution, basing on Tunisian published data during a period extended from 2006, the date of its first description in Tunisia, to February 2019. We also resumed the different mobile genetic elements implicated in their dissemination. This review shows that the majority of the research reports focused in the north and the coastal cities in spite of the fact that KPC and IMP carbapenemases were uncommonly detected in our country. However, VIM, NDM-1, and OXA-48 enzymes were usually reported with the predominance of OXA-48 among Enterobacteriaceae. Furthermore, OXA-23, OXA-51, and OXA-58 carbapenemases constituted the main mechanism conferring carbapenem resistance among Acinetobacter baumannii in Tunisia. Collaborative efforts and raising awareness of the threat of antibiotic resistance are required in order to minimize the spread of multidrug-resistant bacteria.
Keywords: Gram-negative bacteria, carbapenem resistance, carbapenemases, mobile genetic elements, Tunisia
Introduction
Global antibiotic consumption is steadily increasing, particularly in low-income countries, including Tunisia, compared to high-income countries that have managed to develop strategies for good control of antibiotic use.1–3 Beta-lactams are the most used antibiotics in antibiotherapy. Accordingly, the importance of their use is justified by their therapeutic efficacy, their broad spectrum of action, and their low toxicity.4,5 However, the use of antibiotics is not restricted to the human medicine but also in several sectors such as agriculture, animal husbandry, aquaculture, and water treatment.6 Moreover, non-medical use of antibiotics contributed significantly to the global challenge of antibiotic resistance and the wide spread of multidrug-resistant bacteria. In fact, a potential risk to human health was described as the transmission of multidrug-resistant bacteria to healthy people through the food chain was observed.7
The crisis of antibiotic resistance was dramatically exacerbated by the global rise in consumption of the last resort of beta-lactam antibiotics, carbapenems which are considered as one of the treatments of choice against severe infections caused by multidrug-resistant bacteria.8 Bacterial resistance to carbapenems was mainly conferred by carbapenemase production.9 This mechanism was commonly described in Gram-negative bacteria. Indeed, more than 350 carbapenemase variants were currently identified (http://www.lahey.org/Studies/) and distributed in the entire world, noting the dominance of KPC, NDM-1, VIM, IMP, and OXA-48 enzyme types.10 Besides, OXA-23 carbapenemase was frequently detected among carbapenem resistant Acinetobacter baumannii.11
In Tunisia, carbapenem consumption was folded during the period of 2011–2015. This is directly related to the increasing prevalence of severe infections caused by multidrug-resistant Gram-negative bacteria (GNB) (http://www.dpm.tn/). Several carbapenemases producing Gram-negative species have been recorded in Tunisia, as well as numerous carbapenemase variants which have been detected since 2006, the date of the emergence of the first carbapenemase producing isolate in Tunisia.12
In this review, we aimed to illuminate on the dissemination of different carbapenemase variants detected among CR-GNB isolated from various origins in Tunisia, based on Tunisian data published over 13 years from 2006 to 2019. This study enabled us to gain deep insight into the carbapenem resistance problem in Tunisia, in addition to mapping the regional distribution of carbapenemase enzymes, which is strongly encouraged by epidemiologists in order to improve surveillance strategies to minimize the spread of CR-GNB such as previously described.13
Carbapenemase Producing Gram-Negative Bacteria from Hospitals
Since 2006, the date of the first description of carbapenemase producing Klebsiella pneumoniae clinical isolate in a Tunisian hospital,12 several types of carbapenemases have been detected in different species isolated from diverse clinical specimens and/or environmental hospitals, as shown in Table 1. KPC enzyme has recently been detected in Klebsiella pneumoniae and Escherichia coli clinical isolates and is still uncommon in Tunisia.14,15 This enzyme was firstly discovered in a clinical isolate of Klebsiella pneumoniae in North Carolina in 2001.16 Since then, it has dramatically disseminated around the world, with a high prevalence in Greece, Italy, and Latin America,17,18 whereas, it has rarely been detected in Africa.19 The KPC enzyme was described in various members of Enterobacteriaceae and non-fermenters such as Pseudomonas aeruginosa and Acinetobacter baumannii.18
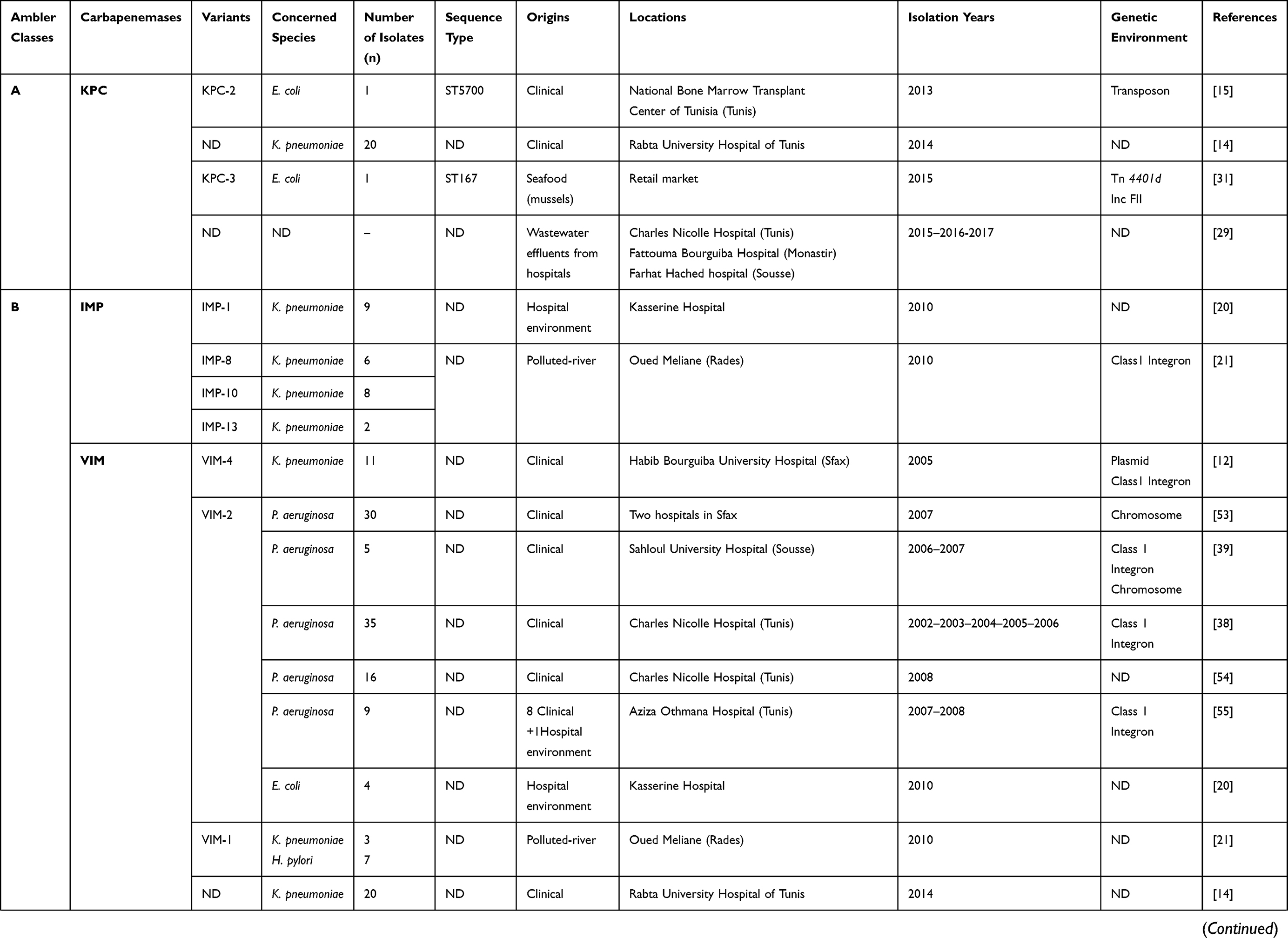 | 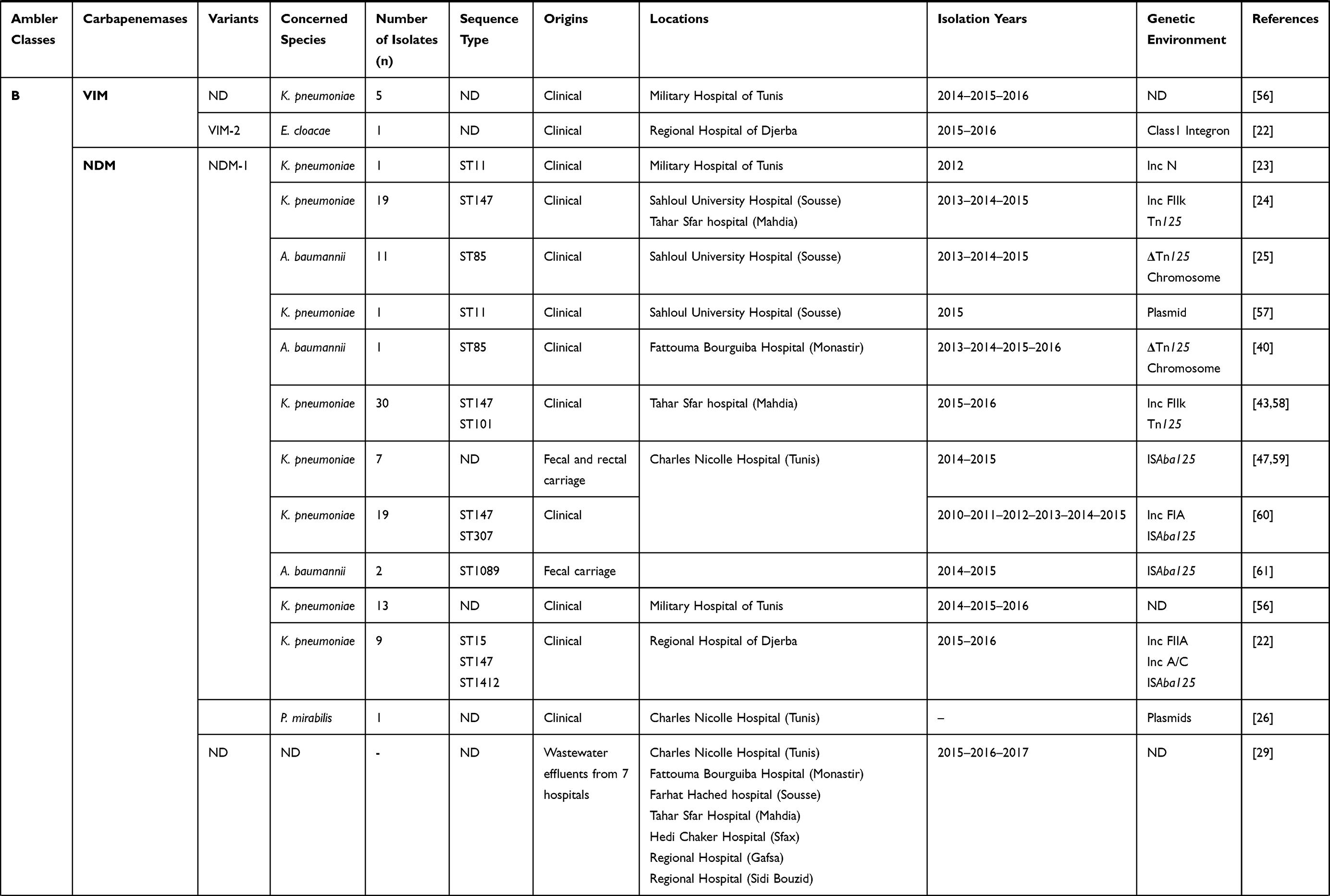 | 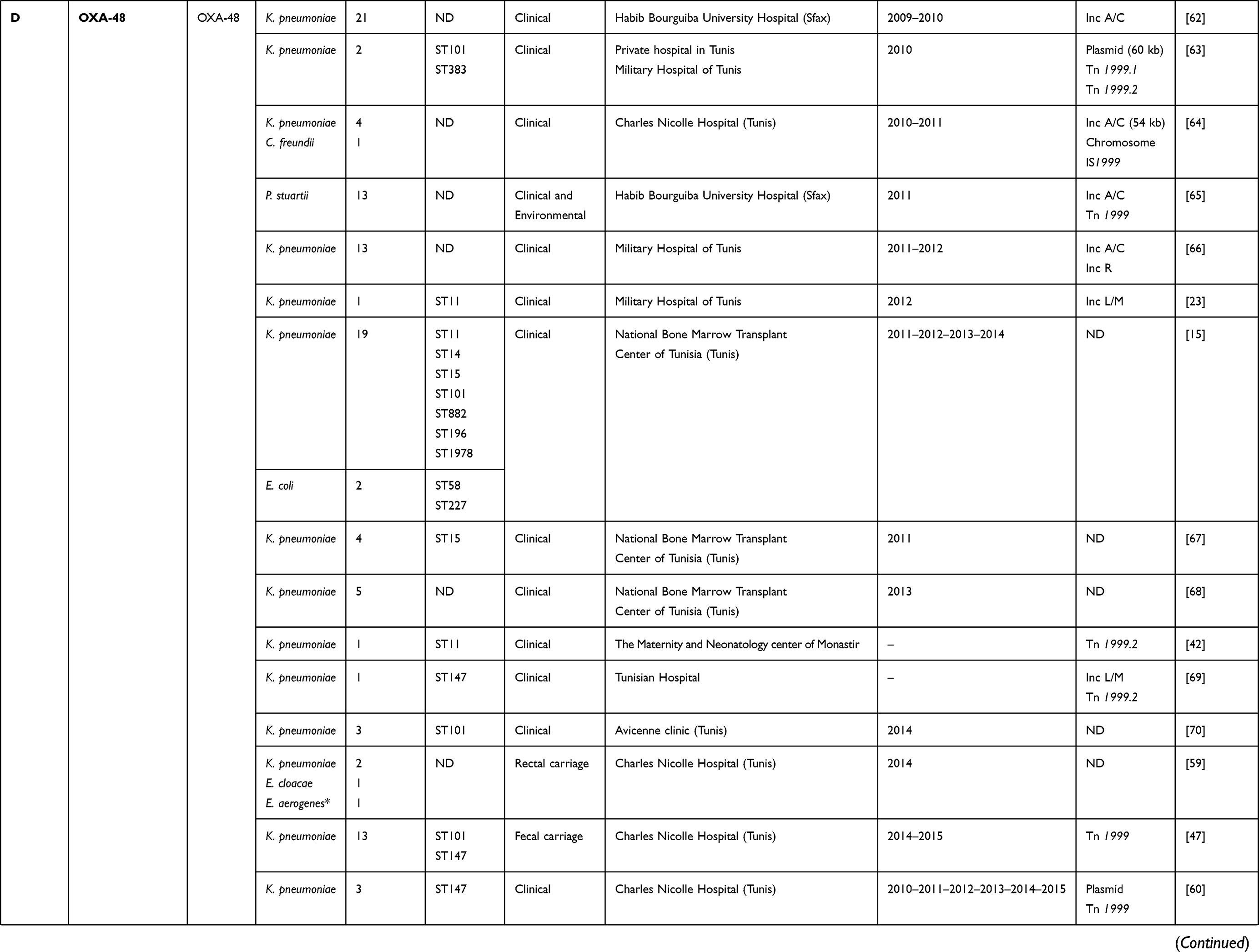 | 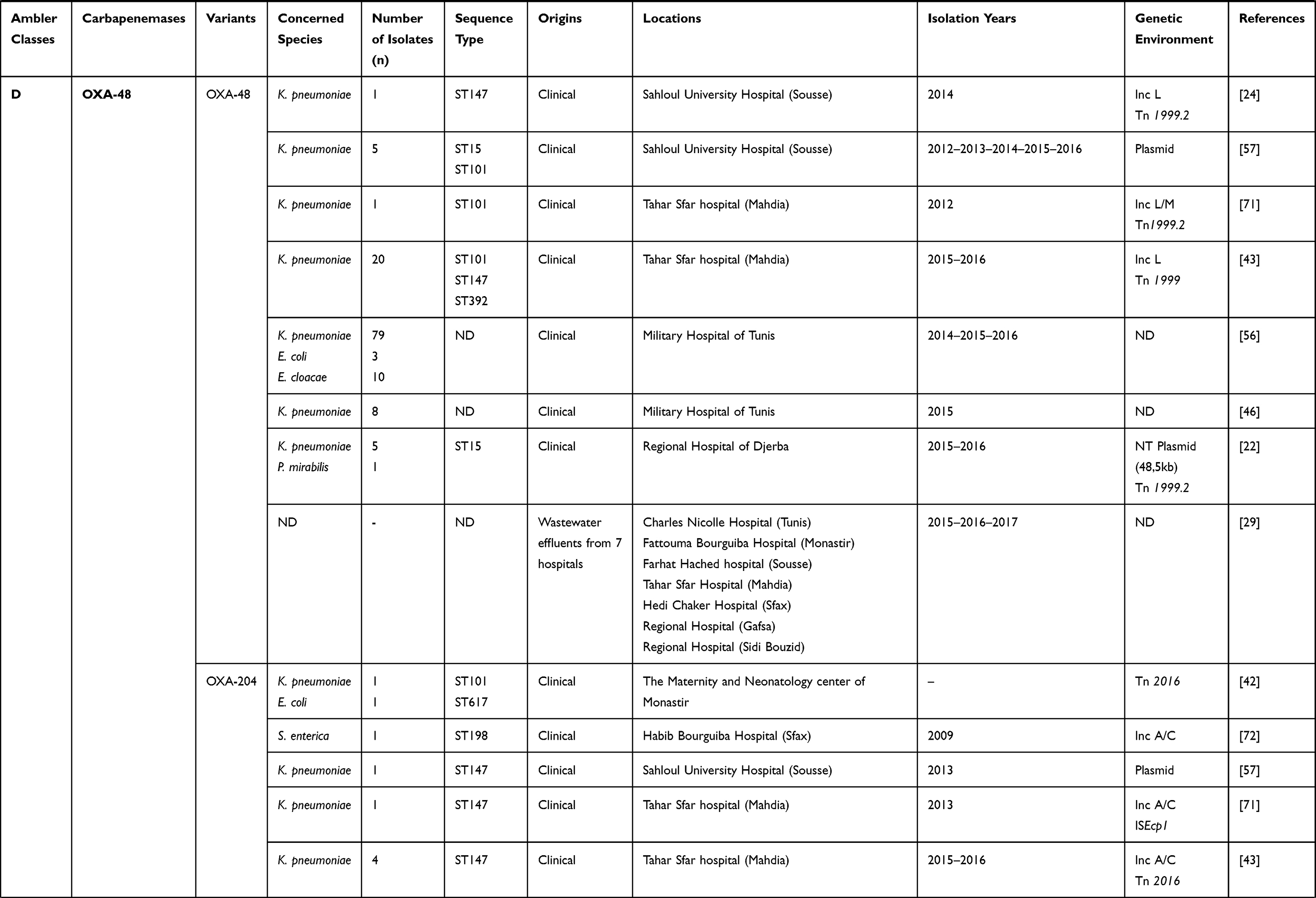 | 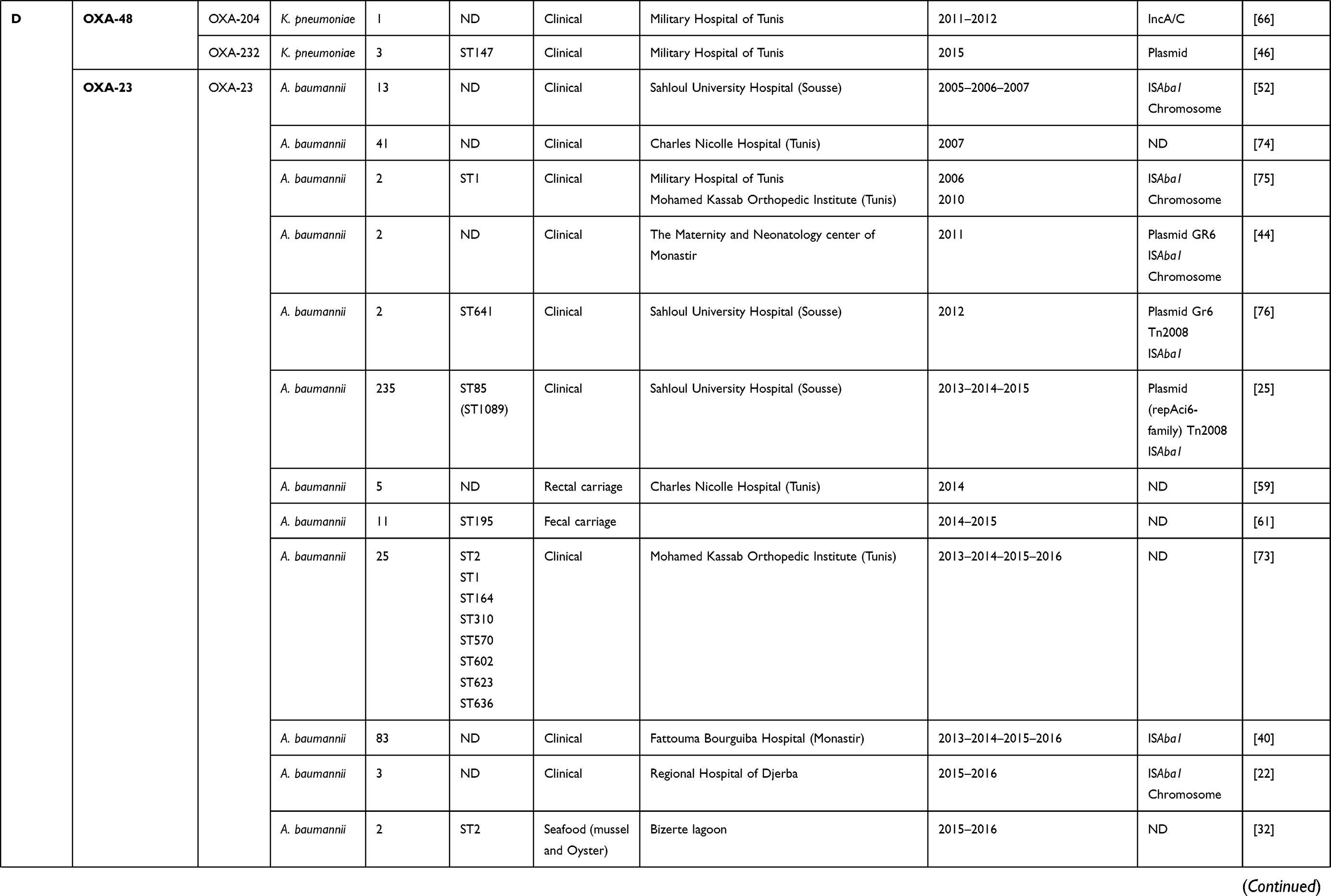 | 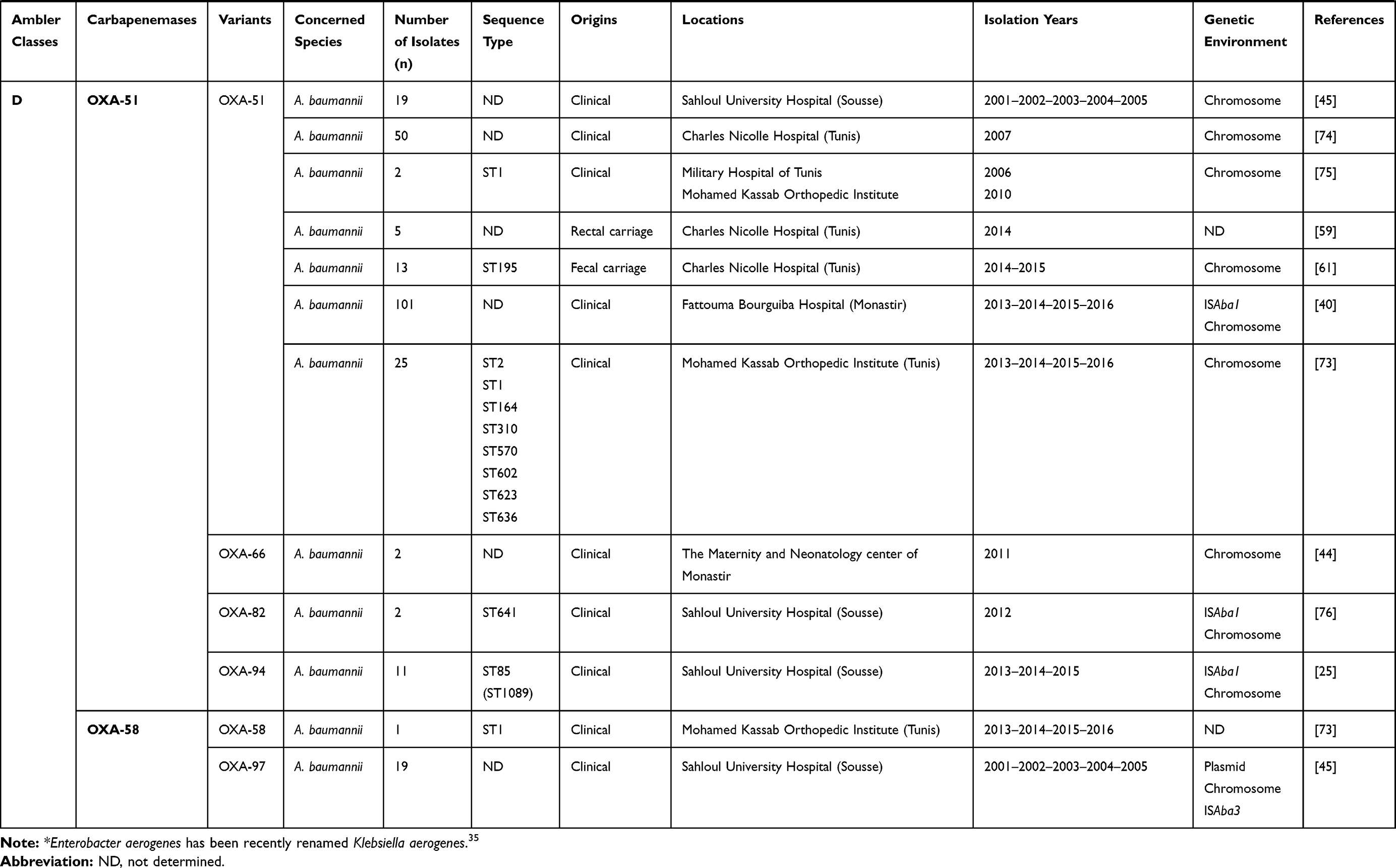 |
Table 1 Characteristics of Carbapenemase Producing Gram-Negative Bacteria in Tunisia from 2006 to 2019 |
IMP metallo-β-lactamase type was scarcely reported in Tunisia, and it was observed only in Klebsiella pneumoniae isolated from the environment.20,21 Several investigations revealed that Asian countries, especially Japan and China, were the main reservoirs of this enzyme.9
Moreover, we have noted that the VIM enzyme was the main metallo-β-lactamase detected in Tunisia, and was frequently reported among Pseudomonas aeruginosa, followed by Klebsiella pneumoniae (Tables 1 and 2). This enzyme was scarcely reported among Escherichia coli and Enterobacter cloacae in Tunisia.20,22 In addition, the NDM-1 encoding gene was dramatically disseminated in Tunisia despite being lately reported comparing with other metallo-β-lactamases,23,24 and it was mainly detected among Klebsiella pneumoniae (Tables 1 and 2). However, recent studies reported the detection of NDM-1 in Acinetobacter baumannii and Proteus mirabilis.25,26
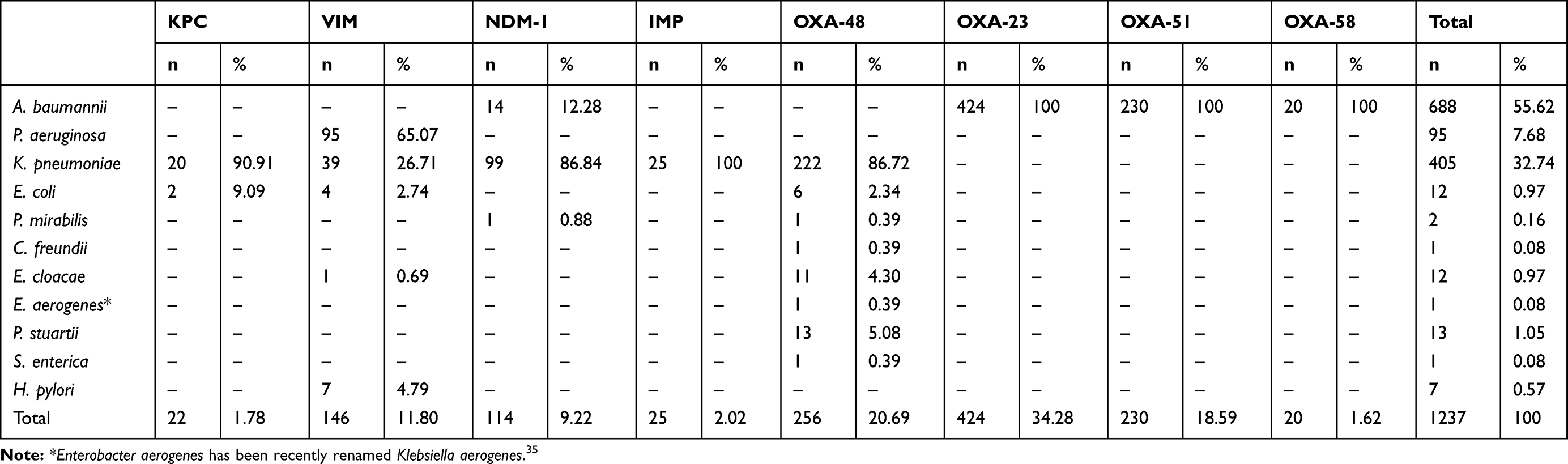 |
Table 2 The Frequencies of Carbapenemases Detected in Each Species in Tunisia from 2006 to 2019 |
Concerning class D carbapenemases, we have noted the circulation of four major subfamilies in Tunisia, namely; OXA-48-like, OXA-23-like, OXA-51-like, and OXA-58-like (Table 1). Besides, three OXA-48-like variants (OXA-48, OXA-204, and OXA-232) were found in Enterobacteriaceae, most commonly Klebsiella pneumoniae. OXA-48 production was considered as the main mechanism conferring resistance to carbapenems among Enterobacteriaceae in Tunisia, as well as Mediterranean countries where considered as endemic.27 However, carbapenem resistance among Acinetobacter baumannii species was usually associated with the production of OXA-23, OXA-58, and the intrinsic OXA-51 carbapenemases, as shown in Table 1. OXA-51 encoding gene is intrinsic to all Acinetobacter baumannii isolates. Therefore, a high prevalence of this gene is expected when this species is studied. This gene will only lead to carbapenem resistance when it is adjacent to the insertion sequence ISAba1 which promotes its expression. Indeed, the simple detection of the gene does not imply that the strain is resistant to carbapenems.28
On the other hand, carbapenemase encoding genes were also detected in the hospital effluents. A recent study has recorded the presence of KPC enzyme in the effluent of three hospitals characterized by a higher patient volume and higher volume of carbapenem consumption. One of these hospitals is located in the capital city, whereas the two others are in coastal cities.29 In contrast, NDM metallo-β-lactamase and OXA-48 class D enzyme were detected in the effluents of seven hospitals distributed throughout the country.29
In general, hospital effluents are discharged into the natural environment after treatment using wastewater treatment plants (WWTPs). As previously reported, The WWTPs are not effective for removing resistance genes, and microbial drug resistant bacteria, consequently, constitute a potential risk for the spread of carbapenemase producers in our natural ecosystems (waters, animals, soils, plants, …).29,30
Carbapenemase Producing Gram-Negative Bacteria from Other Origins
Detection of carbapenem-resistant bacteria in Tunisia was not limited to the hospitals, but also it was observed in natural sources (Table 1). Indeed, carbapenemase producing Gram-negative species were isolated from a polluted river, located in the north of the country, and exposed to urban, industrial and hospital effluents.21 In this river, carbapenemase production was observed among Klebsiella pneumoniae isolates harboring metallo-β-lactamases encoding genes, namely blaIMP-8, blaIMP-10, blaIMP-13, and blaVIM-1. This latter was also detected among Helicobacter pylori species.21 Moreover, a KPC-3 producing Escherichia coli, belonging to the ST167 clone, has been recently isolated from seafood bought from a Tunisian retail market.31 Besides, another study reported the emergence of OXA-23 producing Acinetobacter baumannii ST2 in seafood collected from Bizerte lagoon that was contaminated by hospital effluents.32 This wide spread of OXA-23 producing clone has been recently detected among two fish belonging to the Pagellus acarne species fished in the Mediterranean Sea of Algeria, near Bejaia, which is about 626.3 km away from Bizerte.33 The emergence and spread of carbapenem-resistant Gram-negative bacteria in the aquatic environment in Tunisia, as well as all over the world, poses an alarming concern. This is certainly related to the pollution, which is caused mainly by the discharge of hospitals effluents into the natural and aquatic environment. Several studies have shown that wastewater treatment plants (WWTP) playa major role in the release of antibiotics and antibiotic-resistant bacteria into the environment, particularly in low-income countries where many wastewater treatments plants functions are ineffective.34 Consequently, in order to preserve our natural wealth, it was strongly advised to control hospital effluents that constitute the main reservoir of these multidrug-resistant bacteria, especially carbapenemase producers.29
Regional Distribution of Carbapenemases
The emergence and rapid spread of carbapenem-resistant GNB in Tunisia encouraged scientific researchers and epidemiologists to investigate genes involved in resistance to carbapenems. As described in Figure 1A, the number of published articles per year was clearly increased, specifically after 2014, and indicated by the highest number recorded in 2018. This is closely related to the dissemination of CR-GNB throughout the country and the increased awareness of the importance of surveillance and control of multidrug-resistant bacteria in order to improve the health quality. A high prevalence of carbapenemase producers was detected in our country and they were mainly identified as Acinetobacter baumannii, followed by Klebsiella pneumoniae and Pseudomonas aeruginosa (Table 2). Besides, Figure 1B presents the number of published research papers per carbapenemase type and shows that the majority of the published data reported the detection of the OXA-48 enzyme. This latter is frequently found spread among Klebsiella pneumoniae, followed by Providencia stuartii, Enterobacter cloacae, and Escherichia coli. This enzyme was also detected in one isolate of each species of Proteus mirabilis, Citrobacter freundii, Salmonella enterica, and Enterobacter aerogenes, which was recently renamed Klebsiella aerogenes,35 as shown in Table 2. However, the low number of published articles was recorded for those which reported the detection of IMP metallo-β-lactamase type and OXA-58 class D carbapenemases (Figure 1B).
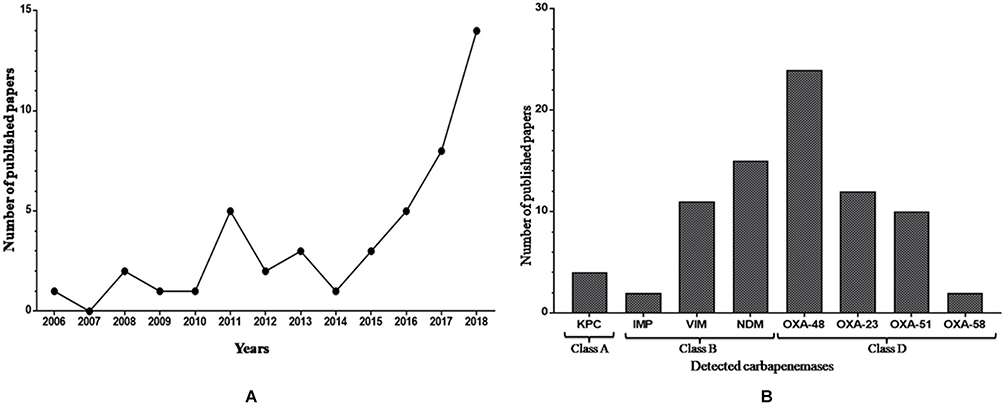 |
Figure 1 (A) Number of published papers reporting carbapenem-resistant Gram-negative bacteria in Tunisia from 2006 to 2019. (B) Number of published papers per carbapenemase classes in Tunisia from 2006 to 2019. |
We have mapped the regional distribution of the different carbapenemases detected in this country since its first description, but we have noticed that epidemiological studies and investigations of resistance mechanisms were not systematically studied either, certainly because of the absence of surveillance programs or even diagnostic microbiology laboratories with sophisticated assays in some regions. As presented in Figure 2, epidemiological studies published to date in Tunisia did not exemplify the situation in Tunisia and just focused on the north and the coastal cities. In addition, we have noted that KPC enzyme was detected only in the northern cities, in contrast to metallo-β-lactamases and class D carbapenemases, which were detected also in the south and center of the country.
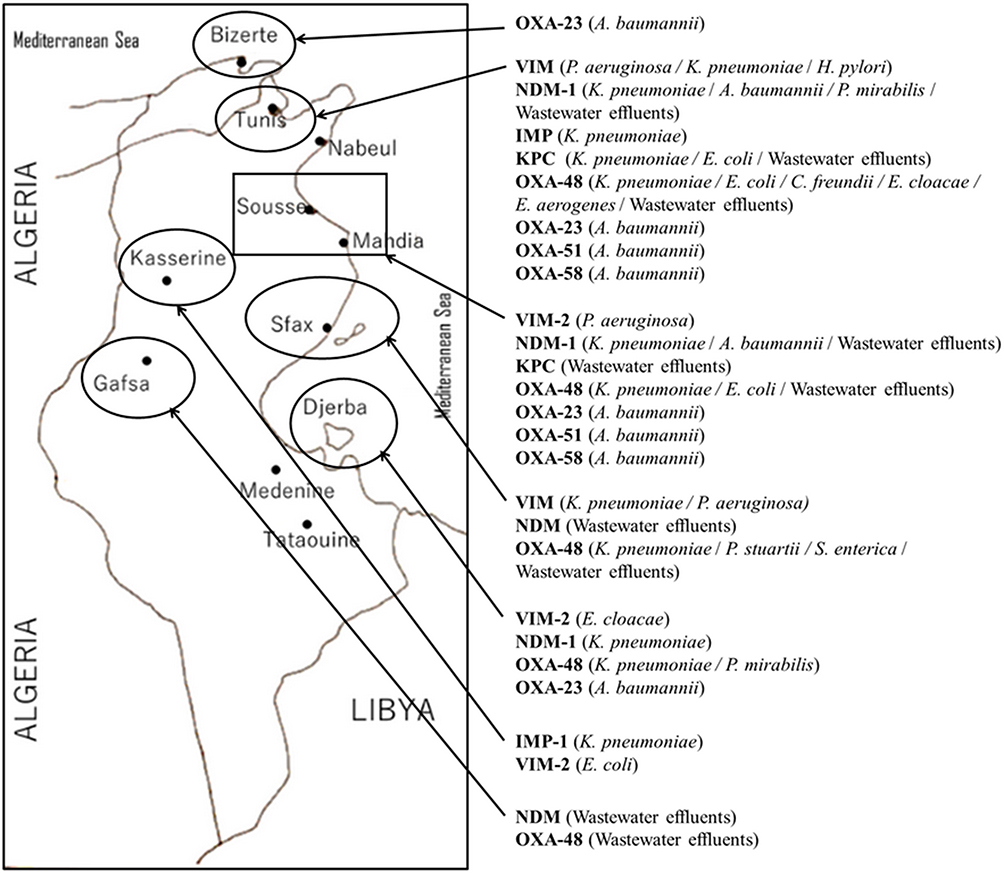 |
Figure 2 Regional distribution of carbapenemase producing Gram-negative bacteria in Tunisia from 2006 to 2019. |
Genetic Environment
Generally, carbapenemase encoding genes are carried by mobile genetic elements such as plasmids, insertion sequences, transposons, and integrons that are responsible for their rapid dissemination.36 As shown in Table 1, several reports focused on studying the genetic environment of detected carbapenemase encoding genes. The blaKPC gene was located in a conjugative plasmid belonging to the Inc FII group and inserted in the Tn3-like transposon, identified as Tn4401d and recognized as an active transposon enhancing the rapid spread of KPC encoding gene.31,37 We cannot be certain that the blaKPC gene is usually harbored by Inc FII plasmid because not all studies determined this. In fact, according to Table 1, only one Escherichia coli isolate was determined to carry the KPC encoding gene in this plasmid replicon type. Concerning class B metallo-β-lactamases encoding genes, it was demonstrated that the blaIMP gene was inserted within the variable region of class 1 integrons.21 This genetic structure was also found to be harboring the blaVIM gene.22,38,39 This latest was further found to be associated with the conjugative plasmids.12 However, the blaNDM-1 gene was mainly found in plasmids which belonged to several incompatibility groups, such as Inc N, Inc FIIk, Inc FIA, Inc FIIA, and Inc A/C (Table 1). The characterization of the genetic structure surrounding the NDM-1 encoding gene showed that it was located in association with transposable elements, known as Tn125, flanked by two insertion sequences ISAba125 conferring a high potential of NDM-1 diffusion.24,40,41
Different plasmids were identified as harboring the blaOXA-48 gene; Inc A/C, Inc L/M, and Inc L, as described in Table 1. In addition, it was noted that the OXA-48 encoding gene was usually associated with the composite transposon Tn1999.2. In contrast, the blaOXA-204 gene was inserted into the Tn2016 transposon structure.42,43 Besides, it was also revealed that the blaOXA-23 gene was carried on the Tn2008 transposon and associated with an upstream located insertion sequence ISAba1. This gene was mainly detected on the chromosome, whereas few data described its plasmidic location.25,44 According to the literature and as shown in Table 1, it was noted that the blaOXA-51-like gene, known as the intrinsic gene of Acinetobacter baumannii, was often detected on the chromosome backbone and sometimes linked to the insertion sequence ISAba1.40 OXA-58-like encoding gene was found to be carried either by chromosome or plasmids and associated with the insertion sequence ISAba3.45
Sequence Types
As shown in Table 1, diverse sequence types (STs) were involved in the dissemination of carbapenemase encoding genes in Tunisia. Indeed, it was noted that KPC producing Escherichia coli isolates belonged to ST5700 and ST167, in contrast to those producing OXA-48-like which belonged to ST58, ST227, and ST617 proving the non-clonal dissemination of carbapenemase producing Escherichia coli in Tunisia.15,31,42 Several sequence types of NDM-1 producing Klebsiella pneumoniae were recorded, including ST11, ST15, ST147, ST101, ST307, and ST1412, noting the predominance of ST147.22–24,46 This latest was also described as the main producer of OXA-48 enzyme in this area, despite the detection of other sequence types such as ST11, ST15, ST14, ST101, ST383, and ST392 (Table 1). Only one OXA-204 producing S. enterica isolate, which belonged to ST198, was hitherto reported.
Regarding non-fermentative GNB, NDM-1 dissemination is associated to only one sequence type Acinetobacter baumannii, which designed ST85 according to the Pasteur database and ST1089 according to Oxford schema.25,40,47 This sequence type was also detected as a producer of OXA-23 carbapenemase type.25 In addition, several sequence types of carbapenemase class D enzyme producing Acinetobacter baumannii were further detected, including ST1, ST2, ST195, ST641, ST164, ST310, ST570, ST602, ST623, and ST636, noting the dominance of ST2 (Table 1).
Treatments
Infections caused by carbapenemase producing Gram negative bacteria, usually qualified as multidrug resistant, constitute a real concern. Indeed, these bacteria are characterized by a high frequency of co-resistance leading to the restriction of treatment choices and presenting real challenges in therapeutic decision-making.48 Few antibiotics were destined for the treatment of these severe infections; polymyxin B (colimycine), polymyxin E (colistin), tigecycline, fosfomycin and some selected aminoglycosides that could be used in combination with carbapenems.49 The clinical treatment of some carbapenemase producers seems to be extremely difficult, quoting the example of P. mirabilis which is intrinsically resistant to colistin and tigecycline;26,50 emergence of such isolates constitutes a real threat.
Several studies addressed the effectiveness of the chosen therapy,49 but herein the Tunisian experience in this field will be reviewed. Regardless of the treatment failure of some cases that ended in patient death, it was very interesting to note the treatment success of several patients. This is closely related to the infection sites and patient response with medication. Indeed, a successful treatment of carbapenemase producing Klebsiella pneumoniae, causing bloodstream infections and co-harboring blaVIM-2, blaOXA-48, blaSHV, and blaCMY-2, with high dose combination of two antibiotics (Imipenem + Amikacin) was recorded.51 This success was also observed when treating patients, suffering from urinary infections caused by VIM-4 producing Klebsiella pneumoniae, with a combination of Imipenem and colimycine that was not effective for those suffering from bacteremia.12 Moreover, it was reported that successive treatments with tigecycline/amikacin and fosfomycin of carbapenemase producing Klebsiella pneumoniae, showing colistin resistance, was also required.43 Another study reported the successful treatment of NDM-1 producing Klebsiella pneumoniae with a combination of (fosfomycin/amikacin), (imipenem/fosfomycin/colistin), or (cefotaxime/tigecycline/imipenem).24 Moreover, a successful treatment of OXA-23 producing Acinetobacter baumannii infection was observed using an association of ceftazidime with amikacin or gentamicin.52
Conclusion
The spread of multidrug-resistant Gram-negative bacteria throughout Tunisia, especially carbapenemase producers, constitutes a real concern. Indeed, we have noted the dissemination of several carbapenemase types among various Gram-negative species, showing a geographical variation that could be explained by the variability of hospital hygiene measures, epidemiological factors, and antibiotic use policies between different Tunisian healthcare settings. Our review is limited to the published data since the first description of carbapenemase production in Tunisia to February 2019, and we should also note the absence of reporting in several regional hospitals because of the absence of surveillance programs or even diagnostic microbiology laboratories with sophisticated assays. Furthermore, the present study reveals the absence of published Tunisian data reporting the carbapenems consumption in the veterinary field as well as the detection of carbapenemases producing Gram-negative bacteria among breeding animals and farms in our country. The lack of such data in this field could be explained by the use of other antibiotics in farms, quoting the example of colistin. This latter has been considered as a successful solution for treatment of carbapenem resistant bacterial infections. However, the spread of colistin resistant encoding genes among breeding animals, patients, and community could exacerbate the crisis, conducting to the pandrug resistance. Intensifying efforts of clinicians, scientists, veterinaries, and ecologists are very required in order to curb the antibiotic resistance challenge which is considered one of the top priorities to improve the health system and to preserve the natural wealth in Tunisia.
Acknowledgments
The authors wish to thank researchers of human health, animal health, environmental sectors, and scientific researchers for their serious and continuous work in the surveillance of carbapenemase producing Gram-negative bacteria.
Author Contributions
All authors contributed to the data analysis, drafting, or revising of the article, gave final approval of the version to be published, and agree to be accountable for all aspects of the work.
The author Olfa DZIRI designed the study and was interested in extracting all Tunisian published data, processing them, and drafting the text, tables, and figures.
The author Raoudha DZIRI revised the draft paper and helped in the preparation of the study’s analytical strategy.
The two authors, Allaaeddin Ali EL SALABI and Chedly CHOUCHANI revised the paper, ensuring the quality and the scientific information of the article.
Funding
This research received no external funding.
Disclosure
The authors declare no conflicts of interest.
References
1. Klein E, Van Boeckel T, Martinez E, et al. Global increase and geographic convergence in antibiotic consumption between 2000 and 2015. Proc Natl Acad Sci U S A. 2018;115(15):E3463–E3470. doi:10.1073/pnas.1717295115
2. Mölstad S, Löfmark S, Carlin K, et al. Lessons learnt during 20 years of the Swedish strategic programme against antibiotic resistance. Bull World Health Organ. 2017;95(11):764–773. doi:10.2471/BLT.16.184374
3. Pulcini C, Gyssens I. How to educate prescribers in antimicrobial stewardship practices. Virulence. 2013;4(2):192–202. doi:10.4161/viru.23706
4. Georgopapadakou N. Penicillin-binding proteins and bacterial resistance to beta-lactams. Antimicrob Agents Chemother. 1993;37(10):2045–2053. doi:10.1128/AAC.37.10.2045
5. Poole K. Resistance to β-lactam antibiotics. Cell Mol Life Sci. 2004;61(17):2200–2223. doi:10.1007/s00018-004-4060-9
6. Meek R, Vyas H, Piddock L. Nonmedical uses of antibiotics: time to restrict their use? PLoS Biol. 2015;13(10):e1002266. doi:10.1371/journal.pbio.1002266
7. Founou L, Founou R, Essack S. Antibiotic resistance in the food chain: a developing country-perspective. Front Microbiol. 2016;7:1881.
8. Hawkey P, Livermore D. Carbapenem antibiotics for serious infections. BMJ. 2012;344:e3236. doi:10.1136/bmj.e3236
9. Walsh T. Emerging carbapenemases: a global perspective. Int J Antimicrob Agents. 2010;36:S8–S14. doi:10.1016/S0924-8579(10)70004-2
10. Logan L, Weinstein R. The epidemiology of carbapenem-resistant Enterobacteriaceae: the impact and evolution of a global menace. J Infect Dis. 2017;215(suppl_1):S28–S36. doi:10.1093/infdis/jiw282
11. Mugnier P, Poirel L, Naas T, et al. Worldwide dissemination of the blaOXA-23 carbapenemase gene of Acinetobacter baumannii. Emerg Infect Dis. 2010;16(1):35–40.
12. Ktari S, Arlet G, Mnif B, et al. Emergence of multidrug-resistant Klebsiella pneumoniae isolates producing VIM-4 metallo-β-lactamase, CTX-M-15 extended-spectrum β-lactamase, and CMY-4 AmpC β-lactamase in a Tunisian university hospital. Antimicrob Agents Chemother. 2006;50(12):4198–4201. doi:10.1128/AAC.00663-06
13. Berrazeg M, Diene S, Medjahed L, et al. New Delhi Metallo-beta-lactamase around the world: an eReview using Google Maps. Euro Surveill. 2014;19(20):
14. Battikh H, Harchay C, Dekhili A, et al. Clonal spread of colistin-resistant Klebsiella pneumoniae coproducing KPC and VIM carbapenemases in neonates at a Tunisian University Hospital. Microb Drug Resist. 2017;23(4):468–472. doi:10.1089/mdr.2016.0175
15. Ben Tanfous F, Alonso CA, Achour W, Ruiz-Ripa L, Torres C, Ben Hassen A. First description of KPC-2-producing Escherichia coli and ST15 OXA-48-positive Klebsiella pneumoniae in Tunisia. Microb Drug Resist. 2017;23(3):365–375. doi:10.1089/mdr.2016.0090
16. Yigit H, Queenan A, Anderson G, et al. Novel carbapenem-hydrolyzing β-lactamase, KPC-1, from a carbapenem-resistant strain of Klebsiella pneumoniae. Antimicrob Agents Chemother. 2001;45(4):1151–1161. doi:10.1128/AAC.45.4.1151-1161.2001
17. Grundmann H, Glasner C, Albiger B, et al. Occurrence of carbapenemase-producing Klebsiella pneumoniae and Escherichia coli in the European survey of carbapenemase-producing Enterobacteriaceae (EuSCAPE): a prospective, multinational study. Lancet Infect Dis. 2017;17(2):153–163. doi:10.1016/S1473-3099(16)30257-2
18. Robledo I, Aquino E, Vázquez G. Detection of the KPC gene in Escherichia coli, Klebsiella pneumoniae, Pseudomonas aeruginosa, and Acinetobacter baumannii during a PCR-based nosocomial surveillance study in Puerto Rico. Antimicrob Agents Chemother. 2011;55(6):2968–2970. doi:10.1128/AAC.01633-10
19. Manenzhe R, Zar H, Nicol M, et al. The spread of carbapenemase-producing bacteria in Africa: a systematic review. J Antimicrob Chemother. 2015;70(1):23–40. doi:10.1093/jac/dku356
20. Chouchani C, Marrakchi R, Ferchichi L, et al. VIM and IMP metallo‐β‐lactamases and other extended‐spectrum β‐lactamases in Escherichia coli and Klebsiella pneumoniae from environmental samples in a Tunisian hospital. APMIS. 2011;119(10):725–732. doi:10.1111/j.1600-0463.2011.02793.x
21. Chouchani C, Marrakchi R, Henriques I, et al. Occurrence of IMP-8, IMP-10, and IMP-13 metallo-β-lactamases located on class 1 integrons and other extended-spectrum β-lactamases in bacterial isolates from Tunisian rivers. Scand J Infect Dis. 2013;45(2):95–103. doi:10.3109/00365548.2012.717712
22. Dziri O, Alonso CA, Dziri R, et al. Metallo-β-lactamases and class D carbapenemases in south-east Tunisia: implication of mobile genetic elements in their dissemination. Int J Antimicrob Agents. 2018;52(6):871–877. doi:10.1016/j.ijantimicag.2018.06.002
23. Ben Nasr A, Decré D, Compain F, et al. Emergence of NDM-1 in association with OXA-48 in Klebsiella pneumoniae from Tunisia. Antimicrob Agents Chemother. 2013;57(8):4089–4090. doi:10.1128/AAC.00536-13
24. Messaoudi A, Haenni M, Mansour W, et al. ST147 NDM-1-producing Klebsiella pneumoniae spread in two Tunisian hospitals. J Antimicrob Chemother. 2017;72(1):315–316. doi:10.1093/jac/dkw401
25. Jaidane N, Naas T, Oueslati S, et al. Whole-genome sequencing of NDM-1-producing ST85 Acinetobacter baumannii isolates from Tunisia. Int J Antimicrob Agents. 2018;52(6):916–921. doi:10.1016/j.ijantimicag.2018.05.017
26. Kanzari L, Ferjani S, Saidani M, et al. First report of extensively-drug-resistant Proteus mirabilis isolate carrying plasmid-mediated blaNDM-1 in a Tunisian intensive care unit. Int J Antimicrob Agents. 2018;52(6):906–909. doi:10.1016/j.ijantimicag.2018.06.009
27. Potron A, Poirel L, Rondinaud E, et al. Intercontinental spread of OXA-48 beta-lactamase-producing Enterobacteriaceae over a 11-year period, 2001 to 2011. Euro Surveill. 2013;18(31):
28. Evans BA, Amyes SGB. OXA β-Lactamases. Clin Microbial Rev. 2014;27(2):241–263. doi:10.1128/CMR.00117-13
29. Nasri E, Subirats J, Sànchez-Melsió A, et al. Abundance of carbapenemase genes (blaKPC, blaNDM and blaOXA-48) in wastewater effluents from Tunisian hospitals. Environ Pollut. 2017;229:371–374. doi:10.1016/j.envpol.2017.05.095
30. Khan F, Söderquist B, Jass J. Prevalence and diversity of antibiotic resistance genes in Swedish aquatic environments impacted by household and hospital wastewater. Front Microbiol. 2019;10:688.
31. Mani Y, Mansour W, Mammeri H, et al. KPC-3-producing ST167 Escherichia coli from mussels bought at a retail market in Tunisia. J Antimicrob Chemother. 2017;72(8):2403–2404. doi:10.1093/jac/dkx124
32. Mani Y, Mansour W, Lupo A, et al. Spread of blaCTX-M-15-producing Enterobacteriaceae and OXA-23-producing Acinetobacter baumannii sequence type 2 in Tunisian seafood. Antimicrob Agents Chemother. 2018;62(9):e00727–18. doi:10.1128/AAC.00727-18
33. Brahmi S, Touati A, Cadière A, et al. First description of two sequence type 2 Acinetobacter baumannii isolates carrying OXA-23 carbapenemase in Pagellus acarne fished from the Mediterranean Sea near Bejaia, Algeria. Antimicrob Agents Chemother. 2016;60(4):2513–2515.
34. Lien L, Hoa N, Chuc N, et al. Antibiotics in wastewater of a rural and an urban hospital before and after wastewater treatment, and the relationship with antibiotic use—a one year study from Vietnam. Int J Environ Res Public Health. 2016;13(6):
35. Tindall BJ, Sutton G, Garrity GM. Enterobacter aerogenes Hormaeche and Edwards 1960 (Approved Lists 1980) and Klebsiella mobilis Bascomb et al. 1971 (Approved Lists 1980) share the same nomenclatural type (ATCC 13048) on the Approved Lists and are homotypic synonyms, with consequences for the name Klebsiella mobilis Bascomb et al. 1971 (Approved Lists 1980). Int J Syst Evol Microbiol. 2017;67(2):502–504.
36. Diene S, Rolain J. Carbapenemase genes and genetic platforms in Gram-negative bacilli: Enterobacteriaceae, Pseudomonas and Acinetobacter species. Clin Microbiol Infect. 2014;20(9):831–838.
37. Cuzon G, Naas T, Nordmann P. Functional characterization of Tn4401, a Tn3-based transposon involved in blaKPC gene mobilization. Antimicrob Agents Chemother. 2011;55(11):5370–5373.
38. Hammami S, Gautier V, Ghozzi R, et al. Diversity in VIM-2-encoding class 1 integrons and occasional blaSHV2a carriage in isolates of a persistent, multidrug-resistant Pseudomonas aeruginosa clone from Tunis. Clin Microbiol Infect. 2010;16(2):189–193. doi:10.1111/j.1469-0691.2009.03023.x
39. Mansour W, Poirel L, Bettaieb D, et al. Metallo-β-lactamase-producing Pseudomonas aeruginosa isolates in Tunisia. Diagn Microbiol Infect Dis. 2009;64(4):458–461.
40. Cheikh H, Domingues S, Silveira E, et al. Molecular characterization of carbapenemases of clinical Acinetobacter baumannii–calcoaceticus complex isolates from a University Hospital in Tunisia. 3 Biotech. 2018;8(7):297.
41. Wailan A, Paterson D. The spread and acquisition of NDM-1: a multifactorial problem. Expert Rev Anti Infect Ther. 2014;12(1):91–115.
42. Charfi K, Mansour W, Khalifa A, et al. Emergence of OXA-204 β-lactamase in Tunisia. Diagn Microbiol Infect Dis. 2015;82(4):314–317. doi:10.1016/j.diagmicrobio.2015.04.003
43. Mansour W, Haenni M, Saras E, et al. Outbreak of colistin-resistant carbapenemase-producing Klebsiella pneumoniae in Tunisia. J Glob Antimicrob Resist. 2017;10:88–94. doi:10.1016/j.jgar.2017.03.017
44. Charfi-Kessis K, Mansour W, Ben Haj Khalifa A, et al. Multidrug-resistant Acinetobacter baumannii strains carrying the blaOXA-23 and the blaGES-11 genes in a neonatology center in Tunisia. Microb Pathog. 2014;74:20–24. doi:10.1016/j.micpath.2014.07.003
45. Poirel L, Mansour W, Bouallegue O, et al. Carbapenem-resistant Acinetobacter baumannii isolates from Tunisia producing the OXA-58-like carbapenem-hydrolyzing oxacillinase OXA-97. Antimicrob Agents Chemother. 2008;52(5):1613–1617. doi:10.1128/AAC.00978-07
46. Lahlaoui H, Bonnin R, Moussa M, et al. First report of OXA-232-producing Klebsiella pneumoniae strains in Tunisia. Diagn Microbiol Infect Dis. 2017;88(2):195–197.
47. Maamar E, Ferjani S, Jendoubi A, et al. High prevalence of gut microbiota colonization with broad-spectrum cephalosporin resistant Enterobacteriaceae in a Tunisian intensive care unit. Front Microbiol. 2016;7:1859. doi:10.3389/fmicb.2016.01859
48. Fritzenwanker M, Imirzalioglu C, Herold S, et al. Treatment options for carbapenem-resistant Gram-negative infections. Dtsch Arztbl Int. 2018;115(20–21):345–352.
49. Lee C, Lee J, Park K, et al. Global dissemination of carbapenemase-producing Klebsiella pneumoniae: epidemiology, genetic context, treatment options, and detection methods. Front Microbiol. 2016;7:895.
50. Qin S, Qi H, Zhang Q, et al. Emergence of extensively drug-resistant Proteus mirabilis harboring a conjugative NDM-1 plasmid and a novel Salmonella genomic island 1 variant, SGI1-Z. Antimicrob Agents Chemother. 2015;59(10):6601–6604. doi:10.1128/AAC.00292-15
51. Hajjej Z, Gharsallah H, Naija H, et al. Successful treatment of a Carbapenem-resistant Klebsiella pneumoniae carrying blaOXA-48, blaVIM-2, blaCMY-2 and blaSHV-with high dose combination of imipenem and amikacin. IDCases. 2016;4:10–12. doi:10.1016/j.idcr.2016.01.003
52. Mansour W, Poirel L, Bettaieb D, et al. Dissemination of OXA-23–producing and carbapenem-resistant Acinetobacter baumannii in a University Hospital in Tunisia. Microb Drug Resist. 2008;14(4):289–292.
53. Ktari S, Mnif B, Znazen A, et al. Diversity of β-lactamases in Pseudomonas aeruginosa isolates producing metallo-β-lactamase in two Tunisian hospitals. Microb Drug Resist. 2011;17(1):25–30. doi:10.1089/mdr.2010.0104
54. Hammami S, Boutiba-Ben Boubaker I, Ghozzi R, et al. Nosocomial outbreak of imipenem-resistant Pseudomonas aeruginosa producing VIM-2 metallo-β-lactamase in a kidney transplantation unit. Diagn Pathol. 2011;6:106. doi:10.1186/1746-1596-6-106
55. Belotti P, Thabet L, Laffargue A, et al. Description of an original integron encompassing blaVIM-2, qnrVC1 and genes encoding bacterial group II intron proteins in Pseudomonas aeruginosa. J Antimicrob Chemother. 2015;70(8):2237–2240. doi:10.1093/jac/dkv103
56. Ben Helal R, Dziri R, Chedly M, et al. Occurrence and characterization of carbapenemase-producing Enterobacteriaceae in a Tunisian Hospital. Microb Drug Resist. 2018;24(9):1361–1367.
57. Jaidane N, Bonnin R, Mansour W, et al. Genomic insights into colistin-resistant Klebsiella pneumoniae from a Tunisian teaching hospital. Antimicrob Agents Chemother. 2018;62(2):
58. Mansour W, Grami R, Jaidane N, et al. Epidemiology and Whole-Genome analysis of NDM-1-producing Klebsiella pneumoniae KP3771 from Tunisia. Microb Drug Resist. 2019;25(5):644–651. doi:10.1089/mdr.2018.0204
59. Hammami S, Dahdeh C, Mamlouk K, et al. Rectal carriage of extended-spectrum beta-lactamase and carbapenemase producing Gram-negative bacilli in intensive care units in Tunisia. Microb Drug Resist. 2017;23(6):695–702. doi:10.1089/mdr.2016.0205
60. Hamzaoui Z, Ocampo-Sosa A, Maamar E, et al. An outbreak of NDM-1-producing Klebsiella pneumoniae, associated with OmpK35 and OmpK36 porin loss in Tunisia. Microb Drug Resist. 2018;24(8):1137–1147. doi:10.1089/mdr.2017.0165
61. Maamar E, Alonso CA, Ferjani S, et al. NDM-1-and OXA-23-producing Acinetobacter baumannii isolated from intensive care unit patients in Tunisia. Int J Antimicrob Agents. 2018;52(6):910–915.
62. Ktari S, Mnif B, Louati F, et al. Spread of Klebsiella pneumoniae isolates producing OXA-48 β-lactamase in a Tunisian university hospital. J Antimicrob Chemother. 2011;66(7):1644–1646. doi:10.1093/jac/dkr181
63. Lahlaoui H, Poirel L, Barguellil F, et al. Carbapenem-hydrolyzing class D β-lactamase OXA-48 in Klebsiella pneumoniae isolates from Tunisia. Eur J Clin Microbiol Infect Dis. 2012;31(6):937–939.
64. Saïdani M, Hammami S, Kammoun A, et al. Emergence of carbapenem-resistant OXA-48 carbapenemase-producing Enterobacteriaceae in Tunisia. J Med Microbiol. 2012;61(12):1746–1749. doi:10.1099/jmm.0.045229-0
65. Mnif B, Ktari S, Chaari A, et al. Nosocomial dissemination of Providencia stuartii isolates carrying blaOXA-48, blaPER-1, blaCMY-4 and qnrA6 in a Tunisian hospital. J Antimicrob Chemother. 2013;68(2):329–332.
66. Ouertani R, Ben Jomàa-Jemili M, Gharsa H, et al. Prevalence of a new variant OXA-204 and OXA-48 carbapenemases plasmids encoded in Klebsiella pneumoniae clinical isolates in Tunisia. Microb Drug Resist. 2018;24(2):142–149. doi:10.1089/mdr.2016.0236
67. Ben Tanfous F, Achour W, Raddaoui A, et al. Molecular characterisation and epidemiology of extended-spectrum β-lactamase-producing Klebsiella pneumoniae isolates from immunocompromised patients in Tunisia. J Glob Antimicrob Resist. 2018;13:154–160.
68. Tanfous F, Raddaoui A, Chebbi Y, et al. Epidemiology and molecular characterisation of colistin-resistant Klebsiella pneumoniae isolates from immunocompromised patients in Tunisia. Int J Antimicrob Agents. 2018;52(6):861–865. doi:10.1016/j.ijantimicag.2018.08.022
69. Ouertani R, Limelette A, Guillard T, et al. First report of nosocomial infection caused by Klebsiella pneumoniae ST147 producing OXA-48 and VEB-8 β-lactamases in Tunisia. J Glob Antimicrob Resist. 2016;4:53–56. doi:10.1016/j.jgar.2015.10.002
70. Mathlouthi N, Al-Bayssari C, El Salabi A, et al. Carbapenemases and extended-spectrum β-lactamases producing Enterobacteriaceae isolated from Tunisian and Libyan hospitals. J Infect Dev Ctries. 2016;10(7):718–727. doi:10.3855/jidc.7426
71. Grami R, Mansour W, Ben Haj Khalifa A, et al. Emergence of ST147 Klebsiella pneumoniae producing OXA-204 carbapenemase in a University Hospital, Tunisia. Microb Drug Resist. 2016;22(2):137–140. doi:10.1089/mdr.2014.0278
72. Ktari S, Le Hello S, Ksibi B, et al. Carbapenemase-producing Salmonella enterica serotype Kentucky St198, North Africa. J Antimicrob Chemother. 2015;70(12):3405–3407.
73. Mathlouthi N, Ben Lamine Y, Somai R, et al. Incidence of OXA-23 and OXA-58 carbapenemases coexpressed in clinical isolates of Acinetobacter baumannii in Tunisia. Microb Drug Resist. 2018;24(2):136–141. doi:10.1089/mdr.2016.0306
74. Hammami S, Ghozzi R, Saidani M, et al. Carbapenem-resistant Acinetobacter baumannii producing the carbapenemase OXA-23 in Tunisia. Tunis Med. 2011;89(7):638–643.
75. Chihi H, Bonnin R, Bourouis A, et al. GES-11-producing Acinetobacter baumannii clinical isolates from Tunisian hospitals: long-term dissemination of GES-type carbapenemases in North Africa. J Glob Antimicrob Resist. 2016;5:47–50. doi:10.1016/j.jgar.2016.03.005
76. Jaidane N, Naas T, Mansour W, et al. Genomic analysis of in vivo acquired resistance to colistin and rifampicin in Acinetobacter baumannii. Int J Antimicrob Agents. 2018;51(2):266–269. doi:10.1016/j.ijantimicag.2017.10.016
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.