Back to Journals » Infection and Drug Resistance » Volume 13
Carbapenemase OXA-423: A Novel OXA-23 Variant in Acinetobacter baumannii
Authors Yang Z ![]() , Wang P, Song P
, Wang P, Song P ![]() , Li X
, Li X ![]()
Received 21 August 2020
Accepted for publication 26 October 2020
Published 10 November 2020 Volume 2020:13 Pages 4069—4075
DOI https://doi.org/10.2147/IDR.S277364
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor Suresh Antony
Zhenghai Yang,1 Peng Wang,2 Ping Song,3 Xiaoning Li1
1Department of Clinical Laboratory, Yijishan Hospital of Wannan Medical College, Wuhu, People’s Republic of China; 2College of Life Sciences, Anhui Normal University, Wuhu, People’s Republic of China; 3College of Biological and Chemical Engineering, Anhui Polytechnic University, Wuhu, People’s Republic of China
Correspondence: Xiaoning Li Department of Clinical Laboratory
Yijishan Hospital of Wannan Medical College, No. 2 Zheshan East Road, Wuhu, Anhui 241000, People’s Republic of China
Tel +86-553-5739411
Fax +86-553-5738279
Email [email protected]
Purpose: A novel variant of OXA-23, named OXA-423, was identified in an Acinetobacter baumannii clinical isolate. The aim of this study was to analyse the resistance phenotype of OXA-423.
Methods: The A. baumannii strain WY-0713 was isolated from an intensive care unit patient. PCR was used to detect the blaOXA-23-like genes. Amplifying, cloning and sequencing were performed for the complete blaOXA-23-like. The novel blaOXA-423 and its ancestor blaOXA-23 were cloned into the expression vector pET-28b(+), and transformed into E. coli Rosetta (DE3) for antibiotic susceptibility testing. SDS-PAGE, modified Hodge test and CarbaNP test were used for detecting the expression of OXA-423 and OXA-23.
Results: PCR screening of A. baumannii WY-0713 was positive for blaOXA-23-like genes. Sequencing of the PCR product identified a novel blaOXA-23-like, named blaOXA-423 which encoding OXA-423. OXA-423 differed from OXA-23 by a crucial amino acid substitution (Val128Ala). The V128A substitution was located at the conserved active-site motifs SAV of OXA-23. Antibiotic susceptibility testing performed using isogenic E. coli showed that the MICs of E. coli Rosetta (pET-OXA-423) for penicillins and carbapenems were lower (reduced MICs 4-fold to 16-fold) than that of E. coli Rosetta (pET-OXA-23). The MICs of cefotaxime, ceftazidime and aztreonam for both transformants remained the same as the acceptor strain. Moreover, OXA-423 was slightly inhibited by sulbactam, clavulanic acid and tazobactam. SDS-PAGE analysis showed that OXA-423 and OXA-23 were conspicuously expressed. Modified Hodge test and CarbaNP test were positive demonstrated both of them were functional.
Conclusion: OXA-423, the first report of an amino acid substitution located at conserved active-site motifs of OXA-23, conferred lower MIC values of penicillins and carbapenems as compared with OXA-23, while without affecting the resistance profiles of expanded-spectrum cephalosporins and aztreonam.
Keywords: OXA-423, OXA-23, Acinetobacter baumannii, carbapenemase, CHDL
Introduction
Acinetobacter baumannii is one of the most troublesome opportunistic pathogens, especially for ICU patients.1 Carbapenem antibiotics are the most important treatment strategies for infections caused by A. baumannii, due to their efficacy, safety and the increased rate of A. baumannii resistance to other antibiotics.2 Anxiously, Carbapenem-resistant A. baumannii (CRAB) has been classified as one of the critical-priority pathogens that need investments in new antibiotics research and development by World Health Organization, due to its increasing prevalence and resistance.3 Carbapenem-hydrolyzing class D β-lactamases (CHDLs) have been considered as the primary factors responsible for carbapenem resistance in A. baumannii.4 Nowadays, six subclasses of CHDLs have been identified in A. baumannii: OXA-23-like, OXA-24/40-like, OXA-58-like, OXA-143-like, OXA-235-like, and the intrinsic chromosomal OXA-51-like.5 Among them, OXA-23-like have spread to many locations worldwide through plasmid-mediated transfer mechanism.5 So far, 40 OXA-23-like carbapenemases have been identified in Acinetobacter species (ftp://ftp.ncbi.nlm.nih.gov/pathogen/Antimicrobial_resistance/ Data, Accessed October 2, 2020). Previous studies on the structure of the CHDLs had showed that the active site of OXA-23-like is represented by three conserved motifs (STFK, SAV and KTG).6 In addition, the Val128 residue has been defined as a crucial residue for carbapenem binding and hydrolytic cleavage.7 Notably, all of the amino acid substitutions of the previously reported OXA-23-like variants were located outside the conserved active-site motifs. Here, we report a novel variant of OXA-23, named OXA-423, which differed from OXA-23 by a crucial amino acid substitution (Val128Ala) in the conserved active-site motif of SAV.
Materials and Methods
Bacterial Strain
Acinetobacter baumannii WY-0713 strain was isolated from the sputum of a hospitalized patient in ICU of Yijishan hospital in China on 13 July 2013. The isolate WY-0713 was initially identified as Acinetobacter calcoaceticus-baumannii complex using VITEK 2 Compact (bioMérieux, Durham, USA) and later species level was confirmed as A. baumannii by sequencing two hyper-variable zone sequences of the rpoB gene, as previously published.8 Multilocus sequence typing (MLST) was performed for the WY-0713 strain according to the Pasteur scheme.9
PCR, Gene Cloning and Sequencing
The blaOXA type genes (blaOXA-23-like, −24/40-like, −51-like, −58-like) were detected by PCR, respectively, with internal primers showed in Table 1. PCR was conducted using the ExTaq mix (Takara, Dalian, China) with the conditions of 95°C for 5 min, 35 cycles (95°C for 30 s, 55°C for 20 s, 72°C for 50 s) and a final elongation step at 72°C for 10 min. Then, in order to amplifying the complete blaOXA-23-like gene, the external primers were used (Table 1). The PCR conditions was 95°C for 5 min, 35 cycles (95°C for 30 s, 55°C for 30 s, 72°C for 1 min) and a final elongation for 10 min at 72°C. The amplicons of a 1008-bp fragment comprising the complete blaOXA-23-like and flanking regions (−46 to +131 bp) were purified using a gel extraction kit (AxyGen, Hangzhou, China) and then cloned into the cloning vector pMD18-T (ampicillin-resistance, Takara, Dalian, China). Subsequently, recombinant plasmids were reproduced in E. coli DH5α cells. The complete blaOXA-23-like was sequenced using an ABI 3730 DNA Analyzer at GenScript Inc. (Nanjing, China). The nucleotide sequences of blaOXA-23-like and deduced amino acid sequences were analyzed using BioEdit™ software and BLAST programs.
 |
Table 1 Primers Used in This Study |
Construction of E. coli Transformants
To analyse the resistance phenotype, blaOXA-423 and its ancestor blaOXA-23 were cloned into the expression vector pET-28b(+) (kanamycin-resistant, Novagen, USA) and overexpressed in E. coli Rosetta (DE3) (chloramphenicol resistance), respectively. Both genes were amplified using primers OXA-F-NdeI and OXA-R-XhoI (Table 1). The products of PCR were purified and digested with NdeI and XhoI, and then inserted into pET-28b(+). The ligation was performed at 16°C for 18 h with a 1:1 insert/vector ratio with T4 DNA ligase (Thermo Fisher Scientific, Pittsburgh, USA). Recombinant plasmids (pET-OXA-423 and pET-OXA-23) were introduced into E. coli DH5α cells. Transformant strains were selected on LB agar plates containing 20 μg/mL of kanamycin and checked by PCR and sequencing. The recombinant plasmids were then separately transformed into competent E. coli Rosetta (DE3) cells using chemical method, producing two E. coli transformants. Cells were selected on MH agar plates containing 20 μg/mL of kanamycin and 20 μg/mL of chloramphenicol. Selected transformants were inoculated in 200 mL of fresh MH broth supplemented with the same antibiotics. The transformants were cultured at 37°C with shaking (225 rpm) until the OD600 reached to 0.6. Then, IPTG was added to the cells at a final concentration of 0.1 mM to induce the expression of OXA-423 and OXA-23.
Antibiotic Susceptibility Testing
The induced bacterial cells were used for antibiotic susceptibility testing. The minimum inhibitory concentrations (MICs) for the clinical strain WY-0713 and transformants were determined by the agar dilution method, according to the protocols of the CLSI.10 The modified Hodge test, CarbaNP test and SDS-PAGE were used for detection of the expression of the two proteins.
Protein Modeling
The X-ray structure of OXA-23-meropenem complex (PDB code: 4JF4) was downloaded from the Protein Data Bank database. SWISS-MODEL server (http://swissmodel.expasy.org/) was employed to create the homology model of OXA-423 using OXA-23-meropenem complex structure as a template. Protein structural figures were conducted using PyMOL.11
Nucleotide Sequence Accession Number
The nucleotide sequence of blaOXA-423 we reported has been submitted to the GenBank database under accession number KM433672.
Results
PCR, Sequencing and Expressing
The blaOXA-23-like and blaOXA-51-like were detected in the A. baumannii WY-0713, but not blaOXA-24/40-like and blaOXA-58-like. DNA sequencing of the 1008-bp PCR product identified a novel variant that had two mutations (A327G and T383C) compared with the blaOXA-23 gene sequence. The nucleotide alteration at position 327 of the novel blaOXA-23 variant was a synonymous mutation, while mutation at position 383 led to the deduced amino acid substitution of a valine by an alanine at position 128 (DBL numbering system) (Figure 1).12 This novel OXA-23 variant was named OXA-423 by the Lahey clinic (https://www.lahey.org/). DNA sequencing of the amplicon of a 1008-bp fragment comprising the complete blaOXA-23-like and flanking regions (−46 to +131 bp) showed that ISAba1 sequence was identified upstream of blaOXA-23-like in the A. baumannii WY-0713 strain.
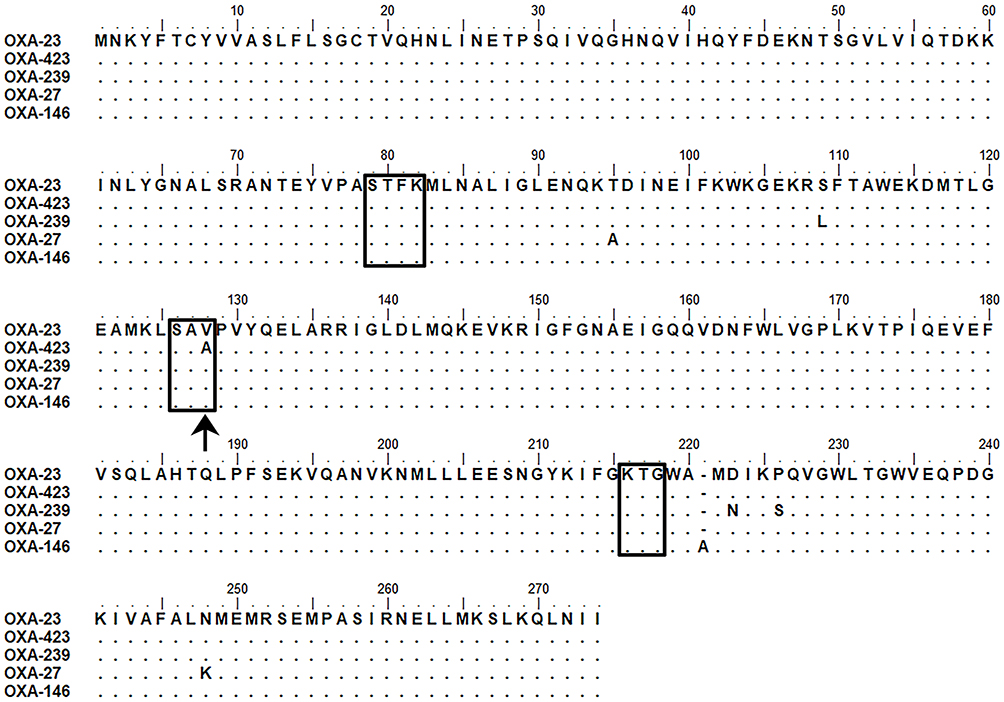 |
Figure 1 Sequence alignment of OXA-423 with several OXA-23-like carbapenemases. Conserved motifs are boxed. The Val128Ala substitution in OXA-423 was indicated by a black arrow. |
SDS-PAGE analysis of the crude extracts of the two transformants, E. coli Rosetta (pET-OXA-423 and pET-OXA-23), showed conspicuously expression of the two proteins. The modified Hodge test and CarbaNP test to carbapenemases were positive for both transformants, demonstrating that the expressed OXA-423 and OXA-23 were functional.
Antibiotic Susceptibility Testing
Antibiotic susceptibility testing performed using isogenic E. coli showed that the MICs of E. coli Rosetta (pET-OXA-423) for penicillins and carbapenems were lower (reduced MICs 4-fold to 16-fold) than that of E. coli Rosetta (pET-OXA-23), suggesting that OXA-423 conferred a weaker hydrolytic capacity towards those substrates, as compared to OXA-23 (Table 2). Moreover, OXA-423 was slightly inhibited by sulbactam, clavulanic acid, and tazobactam. Noteworthy, the MICs of cefotaxime, ceftazidime and aztreonam for both transformants remained the same as the acceptor strain, demonstrating the OXA-423 resistance profile to expanded-spectrum cephalosporins and aztreonam was similar to OXA-23 which do not hydrolyze expanded-spectrum cephalosporins or aztreonam (Table 2).
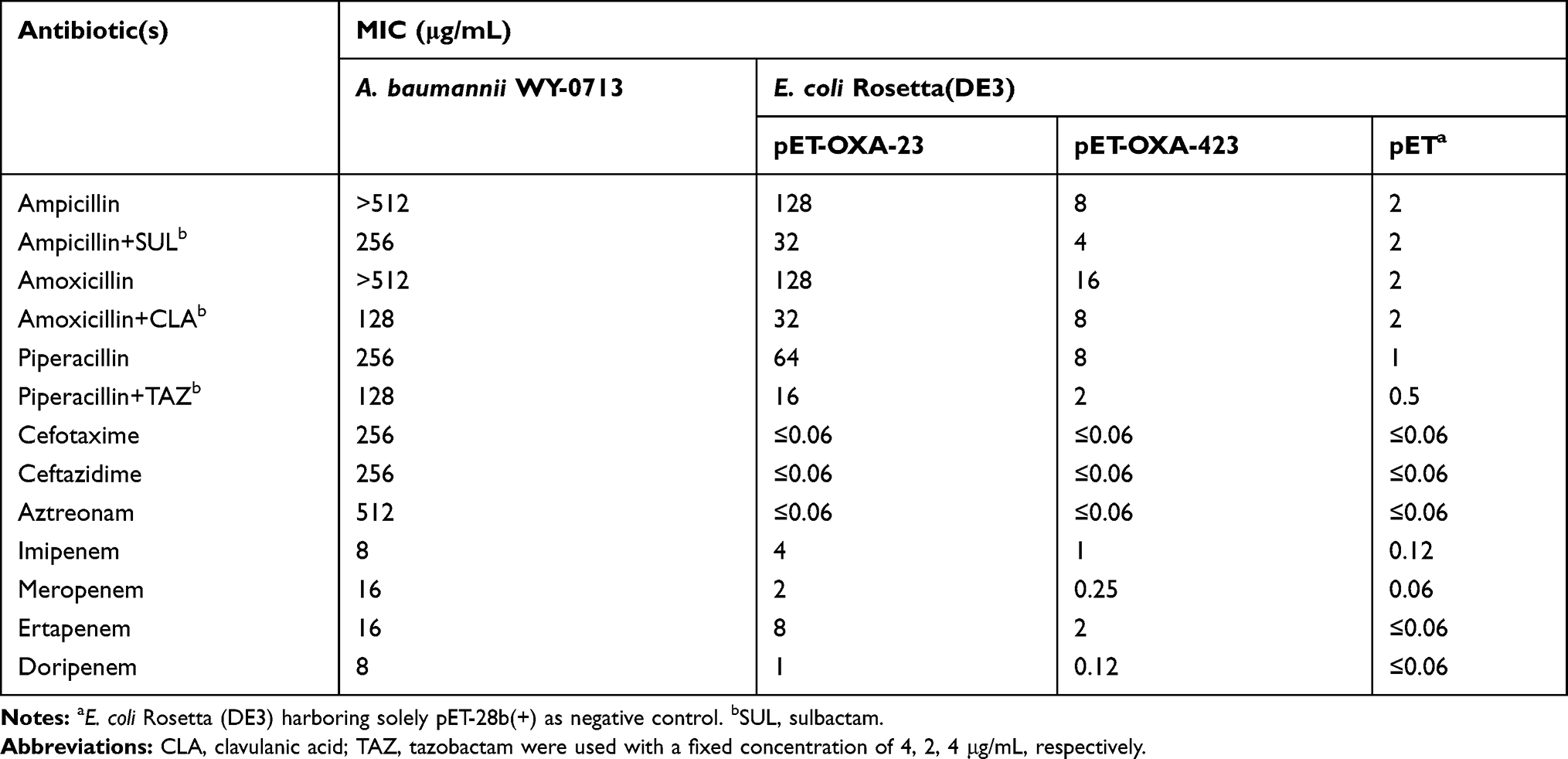 |
Table 2 Antibiotic Susceptibilities of A. baumannii WY-0713 and E. coli Transformants |
Discussion
Currently, 40 further variants of OXA-23 have been identified in Acinetobacter spp. of which differed by one or several amino acids. Previous studies on the structure of the CHDLs had showed that the active site of OXA-23-like is represented by three conserved motifs (STFK positions 79 to 82, SAV positions 126 to 128 and KTG positions 216 to 218) (Figure 1).6 Notably, for the previously reported OXA-23-like variants, all of the amino acid substitutions were located outside the conserved active-site motifs. However, for OXA-423, the Val128Ala substitution was located at the conserved active-site motif SAV (Figure 1). In addition, the Val128 residue has been defined as a crucial residue for carbapenem binding and hydrolytic cleavage.7 Therefore, the Val128Ala substitution will possibly affect the activity to hydrolyze carbapenems, and thus affects the resistance level. This speculation was supported by susceptibility tests, OXA-423 conferred lower MIC values of carbapenems in comparison to OXA-23 when expressed in an isogenic E. coli. Furthermore, DNA sequencing showed that ISAba1 sequence was identified upstream of blaOXA-23 in the A. baumannii WY-0713 strain. Previous studies have demonstrated that the presence of ISAba1 upstream of the blaOXA-23 can increase the expression of OXA-23 and contributes to a high level of carbapenem resistance in A. baumannii.13 Surprisingly, despite other resistance mechanisms existed in the A. baumannii WY-0713 strain, the MIC of imipenem for this clinical strain was still relatively lower when compared with the A. baumannii ATCC 15,151 which harboring ISAba1-blaOXA-23 structure.14 Overall, the strain harboring OXA-423 displayed lower MIC values of carbapenems than that harboring OXA-23, which were both observed in clinical strain and in vitro studies. For the compromised resistance to carbapenems by OXA-423, the most likely explanation was that the Val128Ala substitution conferred reduced carbapenemase hydrolytic activity. To illustrate the potential effect of the substitution, we built a structural model of OXA-423 using the solved OXA-23-meropenem complex structure (PDB code: 4JF4) as a template. Comparison of the homology model of OXA-423 with OXA-23 suggested that the Val128Ala mutation increased the distance of the active site for the substrate as Ala has a smaller side chain than Val (Figure 2). Therefore, the Val128Ala substitution most likely reduces the affinity of OXA-423 for the substrate, and thus weakens the hydrolytic capacity.
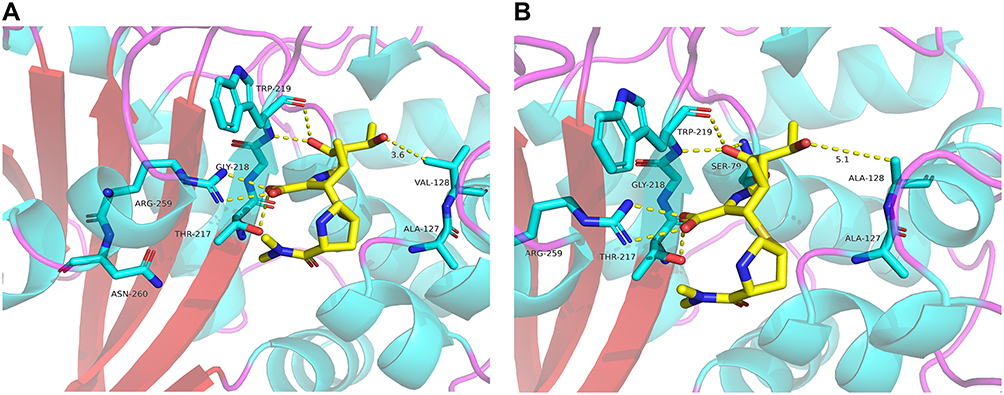 |
Figure 2 Active-site tertiary structure comparison of OXA-23-meropenem complex (A) with the model of OXA-423-meropenem complex (B). Distances (Å) are displayed as yellow dashed lines. |
It is worth noting that as it was described in other OXA-23 variants, such as OXA-239 and OXA-27, decreased hydrolytic activity against some β-lactams may be compensated by increased activity against other β-lactams.15,16 OXA-239, an OXA-23 subfamily variant differed from OXA-23 by three amino acid substitutions (Figure 1), was been identified and horizontally transferred among Acinetobacter spp. in Mexico.17,18 OXA-239 exhibited lower activity against carbapenems, but higher activity against a number of extended-spectrum cephalosporins and aztreonam compared to OXA-23.15 For OXA-27, another OXA-23 variant differed from OXA-23 by two amino acid substitutions (Figure 1), was more active than OXA-23 against imipenem, whereas cephaloridine-hydrolyzing activity was relatively weaker.16 Furthermore, maybe mutation impairs the hydrolytic activity not only against carbapenems but also against other β-lactams drugs, even they were not classic substrates of OXA-23-like enzymes. One example was OXA-146, another OXA-23 variant, differed from OXA-23 by a single residue insertion (A220). This insertion conferred OXA-146 to hydrolyze extended-spectrum cephalosporins and aztreonam more efficiently than OXA-23, while preserving activity to carbapenems.19 Hence, in order to thoroughly understand the resistance phenotype of OXA-423, we determined the MICs of some other drugs: penicillins and β-lactamase inhibitors. In addition, we also detected the MICs of extended-spectrum cephalosporins and aztreonam, although they were not classic substrates of OXA-23-like enzymes. Interestingly, according to the antibiotic susceptibility testing results, the decreased hydrolytic activity against carbapenemases conferred by OXA-423 seemed not been compensated by increased activity against other β-lactams. A particular interest is rapid carbapenemase detection tests such as Modified Hodge test and CarbaNP test are able to detect this variant, although OXA-423 conferred a weaker hydrolytic capacity towards those substrates. It is unknown whether the OXA-423 variant was caused by the selective pressure under carbapenem therapy, as the strain WY-0713 was isolated after received treatment with imipenem (1 g q8h, iv) for 7 days. Curiously, no additional OXA-423 was been detected among Acinetobacter spp. (n=39) during a 2-month investigation period in our hospital since this first identification. Furthermore, database searches carried out through the National Center for Biotechnology Information using the BLAST program showed that OXA-423 was not present in other A. baumannii strains and other species. The reason for no additional OXA-423 has been detected might be because it is an evolutionary dead-end as it does not confer a resistance advantage over OXA-23, which is by large the primary factor responsible for carbapenem resistance in CRAB, as well as being intrinsic to A. radioresistens.20 MLST analysis revealed the strain WY-0713 belonged to ST2, the dominant of International Clone 2 which is distributed worldwide.21 Although only a single case of OXA-423 was identified, ongoing surveillance is important because mutations located at conserved amino-acid motifs are very rare.
Conclusion
OXA-423 reported in this study is a novel variant of OXA-23, conferred lower MICs values of penicillins and carbapenems as compared with OXA-23, while without affecting the resistance profiles of expanded-spectrum cephalosporins and aztreonam. This conclusion needs further studies on catalytic properties and crystal structure of OXA-423. Furthermore, from an evolutionary perspective, OXA-423 might be an evolutionary dead-end as it does not confer a resistance advantage over OXA-23.
Ethics Statement
The isolation of clinical strain was part of the routine hospital laboratory procedure, without the need for informed consent and ethics committee approval. The study complied with the Declaration of Helsinki.
Acknowledgments
Special thanks to George Jacoby for assistance with the numbering of OXA-423. This work was funded by Anhui Province Key Laboratory of Active Biological Macro-molecules Research (LAB201607), and Natural Science Foundation of Anhui Province of China (1808085QC85).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Abdi SN, Ghotaslou R, Ganbarov K, et al. Acinetobacter baumannii efflux pumps and antibiotic resistance. Infect Drug Resist. 2020;13:423–434. doi:10.2147/IDR.S228089
2. Vivas R, Barbosa AAT, Dolabela SS, Jain S. Multidrug-resistant bacteria and alternative methods to control them: an overview. Microb Drug Resist. 2019;25(6):890–908. doi:10.1089/mdr.2018.0319
3. Tacconelli E, Carrara E, Savoldi A; for WHO Pathogens Priority List Working Group. Discovery, research, and development of new antibiotics: the WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect Dis. 2018;18(3):318–327. doi:10.1016/S1473-3099(17)30753-3
4. Ramirez MS, Bonomo RA, Tolmasky ME. Carbapenemases: transforming Acinetobacter baumannii into a yet more dangerous menace. Biomolecules. 2020;10(5):720. doi:10.3390/biom10050720
5. Hamidian M, Nigro SJ. Emergence, molecular mechanisms and global spread of carbapenem-resistant Acinetobacter baumannii. Microb Genom. 2019;5(10):e000306.
6. Santillana E, Beceiro A, Bou G, Romero A. Crystal structure of the carbapenemase OXA-24 reveals insights into the mechanism of carbapenem hydrolysis. Proc Natl Acad Sci U S A. 2007;104(13):5354–5359. doi:10.1073/pnas.0607557104
7. Toth M, Smith CA, Antunes NT, Stewart NK, Maltz L, Vakulenko SB. The role of conserved surface hydrophobic residues in the carbapenemase activity of the class D β-lactamases. Acta Crystallogr D Struct Biol. 2017;73(8):692–701. doi:10.1107/S2059798317008671
8. Gundi VAKB, Dijkshoorn L, Burignat S, Raoult D, La Scola B. Validation of partial rpoB gene sequence analysis for the identification of clinically important and emerging Acinetobacter species. Microbiology. 2009;155(7):2333–2341. doi:10.1099/mic.0.026054-0
9. Diancourt L, Passet V, Nemec A, Dijkshoorn L, Brisse S. The population structure of Acinetobacter baumannii: expanding multiresistant clones from an ancestral susceptible genetic pool. PLoS One. 2010;5(4):e10034. doi:10.1371/journal.pone.0010034
10. Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Susceptibility Testing: Twenty-Ninth Informational Supplement M100-S29. Wayne, PA, USA: Clinical and Laboratory Standards Institute (CLSI); 2019.
11. Bramucci E, Paiardini A, Bossa F, Pascarella S. PyMod: sequence similarity searches, multiple sequence-structure alignments, and homology modeling within PyMOL. BMC Bioinform. 2012;13(Suppl 4):S2. doi:10.1186/1471-2105-13-S4-S2
12. Couture F, Lachapelle J, Levesque RC. Phylogeny of LCR-1 and OXA-5 with class A and class D β-lactamases. Mol Microbiol. 1992;6(12):1693–1705. doi:10.1111/j.1365-2958.1992.tb00894.x
13. Wong MH, Chan BK, Chan EW, Chen S. Over-expression of ISAba1-linked intrinsic and exogenously acquired OXA type carbapenem-hydrolyzing-class D-ß-lactamase-encoding genes is key mechanism underlying carbapenem resistance in Acinetobacter baumannii. Front Microbiol. 2019;10:2809. doi:10.3389/fmicb.2019.02809
14. Lin YC, Hsia KC, Chen YC, et al. Genetic basis of multidrug resistance in Acinetobacter clinical isolates in Taiwan. Antimicrob Agents Chemother. 2010;54(5):2078–2084. doi:10.1128/AAC.01398-09
15. Harper TM, June CM, Taracila MA, Bonomo RA, Powers RA, Leonard DA. Multiple substitutions lead to increased loop flexibility and expanded specificity in Acinetobacter baumannii carbapenemase OXA-239. Biochem J. 2018;475(1):273–288. doi:10.1042/BCJ20170702
16. Afzal-Shah M, Woodford N, Livermore DM. Characterization of OXA-25, OXA-26, and OXA-27, molecular class D β-lactamases associated with carbapenem resistance in clinical isolates of Acinetobacter baumannii. Antimicrob Agents Chemother. 2001;45(2):583–588. doi:10.1128/AAC.45.2.583-588.2001
17. Tamayo-Legorreta EM, Garza-Ramos U, Barrios-Camacho H, et al. Identification of OXA-23 carbapenemases: novel variant OXA-239 in Acinetobacter baumannii ST758 clinical isolates in Mexico. New Microbes New Infect. 2014;2(6):173–174. doi:10.1002/nmi2.60
18. Graña-Miraglia L, Evans BA, López-Jácome LE, et al. Origin of OXA-23 variant OXA-239 from a recently emerged lineage of Acinetobacter baumannii international clone V. mSphere. 2020;5(1):e00801–19. doi:10.1128/mSphere.00801-19
19. Kaitany KC, Klinger NV, June CM, et al. Structures of the class D carbapenemases OXA-23 and OXA-146: mechanistic basis of activity against carbapenems, extended-spectrum cephalosporins, and aztreonam. Antimicrob Agents Chemother. 2013;57(10):4848–4855. doi:10.1128/AAC.00762-13
20. Poirel L, Figueiredo S, Cattoir V, Carattoli A, Nordmann P. Acinetobacter radioresistens as a silent source of carbapenem resistance for Acinetobacter spp. Antimicrob Agents Chemother. 2008;52(4):1252–1256. doi:10.1128/AAC.01304-07
21. Gaiarsa S, Batisti Biffignandi G, Esposito EP, et al. Comparative analysis of the two Acinetobacter baumannii Multilocus Sequence Typing (MLST) schemes. Front Microbiol. 2019;10:930. doi:10.3389/fmicb.2019.00930
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.