Back to Journals » Infection and Drug Resistance » Volume 16
Candida haemulonii Complex and Candida auris: Biology, Virulence Factors, Immune Response, and Multidrug Resistance
Authors Gómez-Gaviria M, Martínez-Álvarez JA ![]() , Chávez-Santiago JO, Mora-Montes HM
, Chávez-Santiago JO, Mora-Montes HM ![]()
Received 26 December 2022
Accepted for publication 6 March 2023
Published 14 March 2023 Volume 2023:16 Pages 1455—1470
DOI https://doi.org/10.2147/IDR.S402754
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Suresh Antony
Manuela Gómez-Gaviria,* José A Martínez-Álvarez, Joaquín O Chávez-Santiago, Héctor M Mora-Montes*
Departamento de Biología, División de Ciencias Naturales y Exactas, Campus Guanajuato, Universidad de Guanajuato, Guanajuato, Gto, México
*These authors contributed equally to this work
Correspondence: Manuela Gómez-Gaviria; Héctor M Mora-Montes, Departamento de Biología, División de Ciencias Naturales y Exactas, Campus Guanajuato, Universidad de Guanajuato, Noria Alta s/n, Col. Noria Alta, Guanajuato, Gto, C. P. 36050, México, Tel +52 473-7320006 Ext. 8193, Fax +52 473-7320006 Ext. 8153, Email [email protected]; [email protected]
Abstract: There is worldwide concern about the constant increase in infections caused by Candida species that are multiresistant to antifungal drugs. The most common candidiasis is caused by Candida albicans, however, the species of the Candida haemulonii complex and Candida auris are emerging opportunistic pathogens, which isolation from clinical samples has significantly increased in the past years. The special interest in the study of these species lies in their ability to evade the action of antifungal drugs, such as amphotericin B, azoles, and echinocandins. In addition, the phenotypic changes of these species have given them the ability to easily adapt to environmental changes, including the host milieu and immunity. In this paper, a detailed review of the current literature on the C. haemulonii complex and C. auris is shown, analyzing aspects such as biology, immune response, putative virulence factors, infection, treatment, and the current strategies for diagnosis.
Keywords: antifungal resistance, candidiasis, cryptic Candida species, emerging pathogens, virulence factors, Candida haemulonii complex, Candida auris
Introduction
Members of the genus Candida are common cause of invasive fungal infections in immunocompromised individuals,1 and are associated with high morbidity and mortality rates.2 A significant number of candidiasis cases have Candida albicans as the etiological agent, however, other Candida species, such as Candida krusei, Candida glabrata, Candida parapsilosis, and Candida tropicalis are frequently isolated from clinical samples.3
The Candida haemulonii complex comprises the opportunistic human pathogens Candida haemulonii sensu stricto, Candida duobushaemulonii, and C. haemulonii var. vulnera; but Candida auris, Candida pseudoheamulonii, and Candida vulturna are phylogenetically related species that often are included as part of the complex.4,5 Candidiasis has increased in frequency in the last years,6 and there is a concern because of the intrinsic antifungal resistance to amphotericin B, azoles, and echinocandins.7,8 The ability of C. haemulonii to undergo dimorphism has been recently demonstrated;9 along with phenotypic switching, a trait that other Candida species have to quickly adapt to changes in the environment and respond against antifungal compounds.9 Currently, two phenotypic switching systems have been described in this species. The first one is white-pink switching, and the second is pink-filament switching; thus defining three phenotypes: white yeasts, pink yeasts, and filament cells., When cells are grown in phloxine B-containing media, white and pink yeast differ in colony color, gene expression profile, secreted aspartyl protease activity, and virulence.9 Another complex member that can cause superficial and invasive infections in humans is C. duobushaemulonii.8,10,11 It was originally classified as C. haemulonii, which led to an underestimation of actual clinical cases associated with this species. The new molecular techniques allowed the correct identification of this complex member, placing the isolates as a new species since conventional biochemical tests led to misleading identification. Some isolates of C. duobushaemulonii are resistant to azoles and polyenes.12
C. auris, currently distributed worldwide, was identified for the first time in Japan, in 2009.4 To date, this specie has been isolated mainly in nosocomial outbreaks and the concern related to this pathogen is that the organism easily passes from one patient to another, especially in those who are immunocompromised.13 This fungal species is a haploid yeast, phylogenetically related to C. haemulonii and Candida lusitaniae, and relatively distant to the diploid species C. albicans and C. tropicalis, whose clinical occurrence is more common.14 C. auris can form biofilms more robust than those generated by other complex members, and undergoes dimorphism, generating true hyphae.13 A feature of clinical concern found in C. auris is the multidrug resistance, especially to the azoles, polyenes, and echinocandins.15–19
C. pseudohaemulonii has been isolated from bloodstream infections and molecular studies showed a close phylogenetic relationship with the C. haemulonii complex. C. pseudohaemulonii can be distinguished from this species by the assimilation pattern of esculine, melezitose, L-rhamnose, trehalose, and glycerol.20 Although the infections caused by this species are low, it is of clinical importance due to its capacity to form biofilms and its resistance to azoles and amphotericin B.15,20 C. vulturna, another related species to the C. haemulonii complex has a natural habitat in plants. However, it has also been found in human bloodstream infections.21–23 The occurrence of invasive candidiasis by this fungus makes it an emerging and opportunistic pathogen of clinical importance. Furthermore, it has been shown that some C. vulturna isolates are susceptible to echinocandins and azoles but resistant to amphotericin B.22,23
The emergence of C. auris and the C. haemulonii complex as causative agents of candidiasis has attracted attention in recent years and exposed the need to increase our knowledge on aspected related to their biology, pathogenesis, interaction with the host, and the correct identification of these organisms.
Epidemiology
Despite C. auris was isolated for the first time in 2009, retrospective studies found that the first isolates of this species date to 1996.14,24 Analysis of cultures collected between the years 2009, 2013, 2014, and 2015 identified four types of C. auris, which were grouped in clades (clade I of South Asia, clade II of East Asia, clade III of South Africa, and clade IV of South America).25 These findings indicate that C. auris may be a recently emerged pathogen.25,26 In 2016, the Center for Disease Control and Prevention, the European Center for Disease Prevention and Control, and Public Health England issued an alert, informing medical centers about C. auris as a new infectious agent.14 Thanks to genomic and epidemiological analyses, in 2017 different genetic populations of C. auris were found. These strains are known to have emerged almost simultaneously on three different continents.25 A decade after its discovery as a bloodstream pathogen, C. auris was recovered from 4000 blood isolates from patients from more than 40 countries.27–29
As of 2021, 47 countries have reported a single case or outbreak of C. auris infection. By the year 2022, according to the Center for Disease Control and Prevention of the United States of America, a large part of the territory was affected by this pathogen. The total number of clinical cases was 2377 and 5754 cases were screened (https://www.cdc.gov/fungal/candida-auris/tracking-c-auris.html#counted accessed February 17, 2023). The epidemiology of the infection caused by C. auris has changed over time, in the early years this pathogen was known to cause sporadic invasive infections. However, currently, the infection has been replaced by nosocomial outbreaks, which have been reported more frequently.27,28,30 This increase in the isolation frequency of this pathogen has meant that C. auris can affect susceptible patients in medical centers.31,32 In addition to the data already known for the United States of America, other countries such as Canada, Mexico, the United Kingdom, Spain, India, Pakistan, Russia, Saudi Arabia, Oman, Kuwait, Kenya, South Africa, and Colombia have also reported C. auris outbreaks.31,33–44
In the case of American countries, such as Mexico and the United States, in the years from 2019 to 2020, 12 patients with candidiasis caused by C. auris were reported. In Mexico, the mortality rate was 67% and in the United States, 40%44,45 Asian countries such as Kuwait, Oman, Russia, and Saudi Arabia, reported an increase in cases of candidiasis from 2018 to 2019. In Kuwait, there were a total of 71 patients infected by C. auris, of which the mortality rate was 51%, in Oman 32 cases were reported, and the mortality rate was 53.1%, in Russia the total number of cases in these years was 38 and the mortality rate was 55.3%. Finally, in Saudi Arabia, the reported cases were 35, with a mortality rate of 20%.39,40,42,46,47 The European continent has also been affected by C. auris, in Spain from 2017 to 2020 47 cases were reported, with a mortality rate of 23.4%.48
The concern around C. auris lies in the characteristics that possess, which include the ability to persist and remain viable for several months. This could be associated with the formation of biofilms on plastic surfaces, in the hospital environment, and in medical devices.49,50 Also, as mentioned below, rates of resistance to fluconazole and other drugs, such as amphotericin B make infection control challenging.51,52
Unlike C. auris, epidemiological data for the C. haemulonii complex are few. However, it is known that most cases caused by C. haemulonii and C. pseudohaemulonii are related to bloodstream infections and people with central venous catheters.53,54 In 2009, 8 cases of fungemia caused by C. haemulonii and C. pseudohaemulonii were reported in Korea. In 2011, the same country saw an increase in the number of cases in both species.15,53,55 In Belgium, of 142 isolates from patients, 27 of these (19%) were identified as C. haemulonii.56 In countries such as Brazil, Mexico, Kuwait, and Argentina, the presence of species belonging to the C. haemulonii complex has also been reported.7,54,57,58 In 2016, in Mexico, candidiasis by C. haemulonii was reported in a pediatric patient with congenital heart disease. To identify the isolate to a species level, it was necessary to carry out 28S and ITS sequencing analysis.59 This case drew attention because the epidemiological and etiological reports of candidiasis in Mexico had not reported yet the presence of C. haemulonii.59 Recently, C. duobushaemulonii was also identified in the country, which is responsible for superficial candidiasis in adults.12
In Brazil, from 2010 to 2015, 14,642 patient samples were cultured in 5 hospitals affiliated with the University of São Paulo. Of all the samples, 40 isolates from 31 patients belonged to the C. haemulonii complex.60 Through molecular identification, it was possible to determine that of these 40 isolates, 14 corresponded to C. haemulonii, 8 to C. haemulonii var. vulnera, and 9 to C. duobushaemulonii.60 Among the morbidities found in the 31 patients, the most frequent were diabetes mellitus and vascular diseases.60 Studies carried out by the Laboratório Especial de Micologia, Escola Paulista de Medicina, Universidade Federal de São Paulo, São Paulo, Brazil, determined that from an 11 years collection of Brazilian yeasts, with a total of 3799 clinical isolates, there was a prevalence of 1.3% of the C. haemulonii complex members.6 Surprisingly, in the last five years there has been a substantial increase in these species. In this study, it was found that C. haemulonii var. vulnera and C. duobushaemulonii were found in 57% of all isolates in this analysis.6 In addition to the increase in the appearance of the species of this complex, through these studies it was possible to determine that these species exhibit high minimal inhibitory concentrations (MIC) against amphotericin B and fluconazole, similar to C. auris. This observation led to think that these fungal pathogens may be multi-resistant. Even though species of the C. haemulonii complex are considered rare human pathogens, concerns about incidence rates and resistance to antifungal drugs appear to be increasing worldwide.5,6,61
Candidiasis Caused by C. auris and the C. haemulonii Complex
The infections caused by Candida spp. can be superficial or systemic diseases. Cutaneous, mucocutaneous, and onychomycosis are some examples of superficial infections, while meningitis, sepsis, and candidemia are examples of systemic diseases.62 The C. haemulonii complex has been isolated from human infections; however, due to the isolates misidentification, the clinical information regarding the incidence and epidemiology of the complex members is limited.60 The clinical manifestations of infections caused by the C. haemulonii complex members are similar to that observed in the cases caused by C. albicans and other non-albicans Candida species.54,63–65 The members of this complex are frequently isolated from immunocompromised patients or individuals with some other aggravated illness, for instance, pulmonary or renal diseases. Another feature of the fungemia caused by this complex is that tends to infect neonates and elder individuals. Central venous catheter insertion and prolonged stays in intensive care units are risk factors to develop a systemic infection.7,57,64 Onychomycosis, peritonitis, leukocytosis, and high fever are features of systemic candidiasis caused by C. haemulonii sensu stricto and the infection is associated with peripheral vascular disease, lower extremity ulcers, and diabetes mellitus.57,64,66 C. duobushaemulonii causes onychomycosis, and vulvovaginal candidiasis, and a characteristic of this species is that infects diabetic patients.12,53,60,63,67 C. haemulonii var. vulnera causes onychomycosis, lower extremity ulcers, and fungemia;12,67 while candidemia caused by C. auris has been found in patients with severe lung and kidney diseases, and in those who suffer from diabetes mellitus.4,17
Identification of C. auris and the C. haemulonii Complex
Fungal isolate identification is of utmost importance from an epidemiological and clinical point of view. In addition to this, understanding the resistance to antifungal drugs is important when providing treatment. When C. auris is cultivated in a broth added with glucose, yeast extract, and peptone at 25℃, grows as ovoid, ellipsoidal cells of 2 to 3–2.5 to 5 μM. The culture on solid medium produced butyric to viscous colonies, in white and gray tones, smooth and shiny.4 Conventional biochemical methods are unreliable when trying to identify C. auris because they can be misidentified and confused with other Candida species. As an examples the VITEK2 system (bioMérieux, Marcy I’Etoile, France) misidentified 10 isolates as C. haemulonii sensu stricto and two as Candida famata. Similarly, the API20C system (bioMérieux) also misidentified them as Candida sake.17
An effective way to differentiate C. auris isolates from other Candida species is by growing them at 40–42°C because only C. auris can grow at this temperature.5,17 Another form of differentiation is taking advantage of its ability to tolerate high salt concentrations17,68 A culture in broth containing yeast nitrogen base, 10% NaCl supplemented with dextrose, ducitol, or mannitol, can differentiate C. auris from C. haemulonii sensu stricto, C. duobushaemulonii, C. albicans, and C. parapsilosis, which cannot grow under these conditions.69
Another method that is commonly used is the commercial CHROMOagar medium. When C. auris isolates are inoculated, they grow as white, pink, or dark purple colonies; while C. duobushaemulonii generates soft light pink colonies, and in the case of C. pseudohaemulonii, the colonies are violet.70 On the other hand, analysis of the new CHROMagar Candida Plus medium showed better performance in the C. auris identification, although C. pseudohaemulonii resembles C. auris in mixed culture colonies, it tends to be much smaller and produces a distinctive blue halo after 72 h. This makes the misidentification of C. pseudohaemulonii with C. auris less likely when found in clinical specimens.71
By using Matrix-Assisted Laser Desorption/Ionization and Time of Flight (MALDI-TOF MS) it is possible to accurately differentiate C. auris, C. haemulonii sensu stricto, and C. duobushaemulonii from other fungal species.13,14 Also, by using the sequencing of genetic loci such as the 28S rDNA D1/D2 region or the rDNA ITS region, they can accurately detect C. auris.10,14,55 C. duobushaemulonii can be identified by sequencing the rDNA ITS (ITS1-5.8S-ITS2) and the D1/D2 region of 26S rDNA.8,72 C. vulturna shows a great coincidence with C. duobushaemulonii, however, its sequence differs by 4% when using the D1/D2 domains, 7% when using ITS, and 1% when comparing it with the SSU 18S rRNA genes.73
Treatment of Candidiasis Caused by C. auris and the C. haemulonii Complex
The main antifungal drugs available to treat Candida infections are classified into four groups. The first are azoles, such as fluconazole, itraconazole, isavuconazole, posaconazole, and voriconazole; whilst the second group are polyenes, such as amphotericin B. The echinocandins anidulafungin, caspofungin, and micafungin are in the third group; and finally, in group four is the pyrimidine analog flucytosine, which is not licensed for monotherapy in Candida infection because the pathogen easily develops resistance. For this reason, its use is recommended in combination with some other antifungal drugs, such as amphotericin B, due to the synergistic effect observed.74,75 The group of azoles has been widely used against Candida infections; however, antifungal resistance to this type of compound has been increasing in recent decades, highlighting that one of the factors for this to occur is the prolonged use of antifungals, for example in patients with acquired immunodeficiency syndrome.76,77
C. auris has intrinsic resistance to fluconazole, voriconazole, amphotericin B, and echinocandins.78,79 Although the mechanism by which C. auris is resistant is unclear, it is known this organism contains ERG3 and ERG11 genes, whose products are enzymes involved in the ergosterol synthesis pathway. It has been found that substitutions in the amino acids Y132F and K143R of the C. albicans ERG11 gene product cause azoles have less interaction with these enzymes, leading to reduced susceptibility to the antifungal drugs.80 The same amino acid substitutions were found in the C. auris ERG11, and for C. pseudohaemulonii the substitution in the amino acid Y132F was also reported, which implies that the resistant mechanisms may be lower azoles binding to targeted enzyme.81–83 In addition, the ERG11 gene was found duplicated in some C. auris isolates.82 The C. auris genes FKS1, FKS2, and FKS3 are involved in glucan synthesis and are related to echinocandin resistance.84 A mutation with amino acid substitution S639F in the FKS1 gene has been reported in several caspofungin-resistant strains.85 In addition, although the main role of transport systems is to allow the uptake of essential nutrients and excretion of metabolic residues; these systems are also used for cellular detoxification, such as the ATP-binding cassette (ABC) family and major facilitator superfamily transporters, which are known to be important factors for C. albicans virulence. These genes have been recently found within the C. auris genome and are thought that contribute to its antifungal resistance.82,86–88
C. auris, C. haemulonii, C. duobushaemulonii, and C. pseudohaemulonii contain the glycosylphosphatidylinositol-anchored proteins Plb3, Iff4, Pga52, Pga20, Csa1, Hyr3, and Pga7, involved in C. albicans biofilm development and overexpressed in C. auris during biofilm formation; thus, these proteins may be related to antifungal resistance.82
The treatment options for patients with infections due to C. auris are complicated, however, the use of echinocandins or amphotericin B is the clinical recommendation,78,79 with the risk that the different isolates show a variable susceptibility. Recently, Ibrexafungerp has been used to treat infections by multidrug resistance Candida species. This drug is an enfumafungin-derived that inhibits β-1,3-D-glucan synthase and is structurally different from echinocandins; thus having different interactions with the targeted enzyme.89
C. haemulonii sensu stricto, C. haemulonii var. vulnera, and C. duobushaemulonii have intrinsic resistance to fluconazole and amphotericin B,4 and some isolates exhibit resistance to itraconazole, voriconazole, posaconazole, terbinafine, and echinocandins.8,10,11,65 Some patients with fungal infections due to C. haemulonii have received combined treatment with amphotericin B and fluconazole with evident improvements in the patient’s health, and another successful combination is amphotericin B deoxycholate plus fluconazole.90
Biological Attributes of C. auris and the C. haemulonii Complex
Recently, C. auris has been recognized as the causative agent of invasive infections and outbreaks with high mortality rates in patients admitted to intensive care units or other special care centers.69 This organism is an oval yeast without pseudohyphae or germ tube formation in most of the isolates,17 and smaller than C. albicans cells.88 However, recent studies have shown that C. auris sometimes fails to release daughter cells after budding, resulting in the formation of pseudohyphal-like cell aggregates.69,91 C. auris colonies appear pink, white, and dark purple in CHROMagar Candida medium at 37°C and 42°C.17,69 Like other Candida species, C. auris can form biofilms, and undergo phenotypical switching, and some isolates are capable to generate true hyphae.14,92 It is hard to discriminate C. auris and C. haemulonii sensu stricto by conventional microbiological tests because have similar assimilation carbohydrate profiles and both are resistant to several antifungal drugs. They are closely related phylogenetically, are part of the Metschnikowiaceae family, and as mentioned, are resistant to some drugs.82,93
The optimal growth of C. haemulonii sensu stricto isolates is at 30°C and tends to decrease at 37°C and is absent at 42°C.93 When cultivated in YPD agar with phloxine B red dye at 25°C, white to pink colonies can be observed. Both phenotypes produce round and budding yeast cells but pink colonies produce two or three times larger cells than the white ones.9 When cultivated in a yeast-peptone-glycerol medium plate at 25°C, wrinkled colonies containing elongated filaments are observed.9 It has been shown that C. haemulonii sensu stricto isolates have glucose- and mannose-containing glycoconjugates on the cell surface.94 In general, these glycoconjugates participate in the interaction between the pathogen and host cell receptors and help in the evasion of host immune responses.95
In 2012, C. haemulonii was classified into three species: C. haemulonii sensu stricto, C. duobushaemulonii, and C. haemulonii var. vulnera, which conform the complex.5 In 2016, 14,642 yeast cultures from 5 hospitals in Brazil were analyzed, 40% were identified as species of the C. haemulonii complex and within these, 9 samples were identified as C. duobushaemulonii.60 The correlative analyses performed in this study concluded that patients with diabetes mellitus are more likely than healthy populations to have infections caused by C. duobushaemulonii.60,96 Furthermore, It has been reported as a causative agent of vulvovaginal candidiasis and wound infection in the lower extremities that can disseminate to the rest of the body.10,96
Also, in 2016, a taxonomic study of yeasts isolated from flowers on the island of Mindanao, Philippines, was carried out, and a species related to C. duobushaemulonii was identified, which was named Candida vulturna. Later, it was described in patients with systemic candidiasis from the Philippines and Malaysia.73 When cultured in yeast extract broth for 24 h at 25°C, it grows like ovoid cells (4–7 by 3–5 µm), alone or in pairs, with apical or subapical budding, forms sediments, and biofilms that contain pseudohyphae.73 On yeast extract agar, after 3 weeks at 25°C, their colonies are usually white and butyric, white in appearance with a complete margin.73
C. albicans, C. tropicalis, C. parapsilosis, and C. auris are part of the CTG clade; which means that these species translate the CTG codon into serine instead of leucine.97 C. albicans and C. tropicalis are clinically common diploid organisms, but C. auris and C. haemulonii sensu stricto are haploid fungi.14 The C. auris genome is approximately 12.3 to 12.5 Mbp in size with 8527 predicted genes.88,98 When compared to the C. albicans genome, a set of orthologs encoding for drug transporters, oligopeptides, proteinases, and mannosyltransferases was identified, and fungal virulence and drug resistance could be related to them. However, as most of the genome is not characterized, it can be expected that the hypothetical proteins may also contribute to cell virulence during interaction with the host.88 The C. haemulonii sensu stricto genome spans 13.3 Mbp with 6155 predicted genes;99 while C. duobushaemulonii is 12.5 Mbp long with 5943 predicted genes.99 When compared, 81 genes from C. haemulonii sensu stricto showed homology with 71 genes from C. duobushaemulonii, but with no other organism, being classified as unique genes of the C. haemulonii complex.99 In 2019, it was sequenced the C. vulturna genome, which contains 12.9 Mbp and 5560 predicted genes.21 Later, in 2020, C. haemulonii var. vulnera was sequenced and, using an average nucleotide identity analysis, it was found that contains 13.21 Mbp, with more than 5400 predicted genes, and showed high identity with C. haemulonii sensu stricto (99%), whilst when compared with C. duobushaemulonii, C. pseudohaemulonii, C. auris, and C. lusitaniae, the identity was 77%, 77%, 75%, and 72%, respectively.100
Virulence Factors in C. auris and the C. haemulonii Complex
Various virulence factors have been recognized in different Candida species, and among these are adhesins, hydrolytic enzymes, biofilm formation, dimorphism, immune evasion, and thermotolerance.14,82,101,102 For the study of virulence factors in C. auris, the species C. albicans has been used as a reference. Even though most of the virulence factors in C. auris and the members of the C. haemulonii complex have not been described, these could be predicted from the comparative genomic analysis between C. albicans and these species. The putative virulence factors found in C. auris, C. haemulonii sensu stricto, C. haemulonii var. vulnera, C. pseudohaemulonii, and C. duobushaemulonii are shown in Table 1.
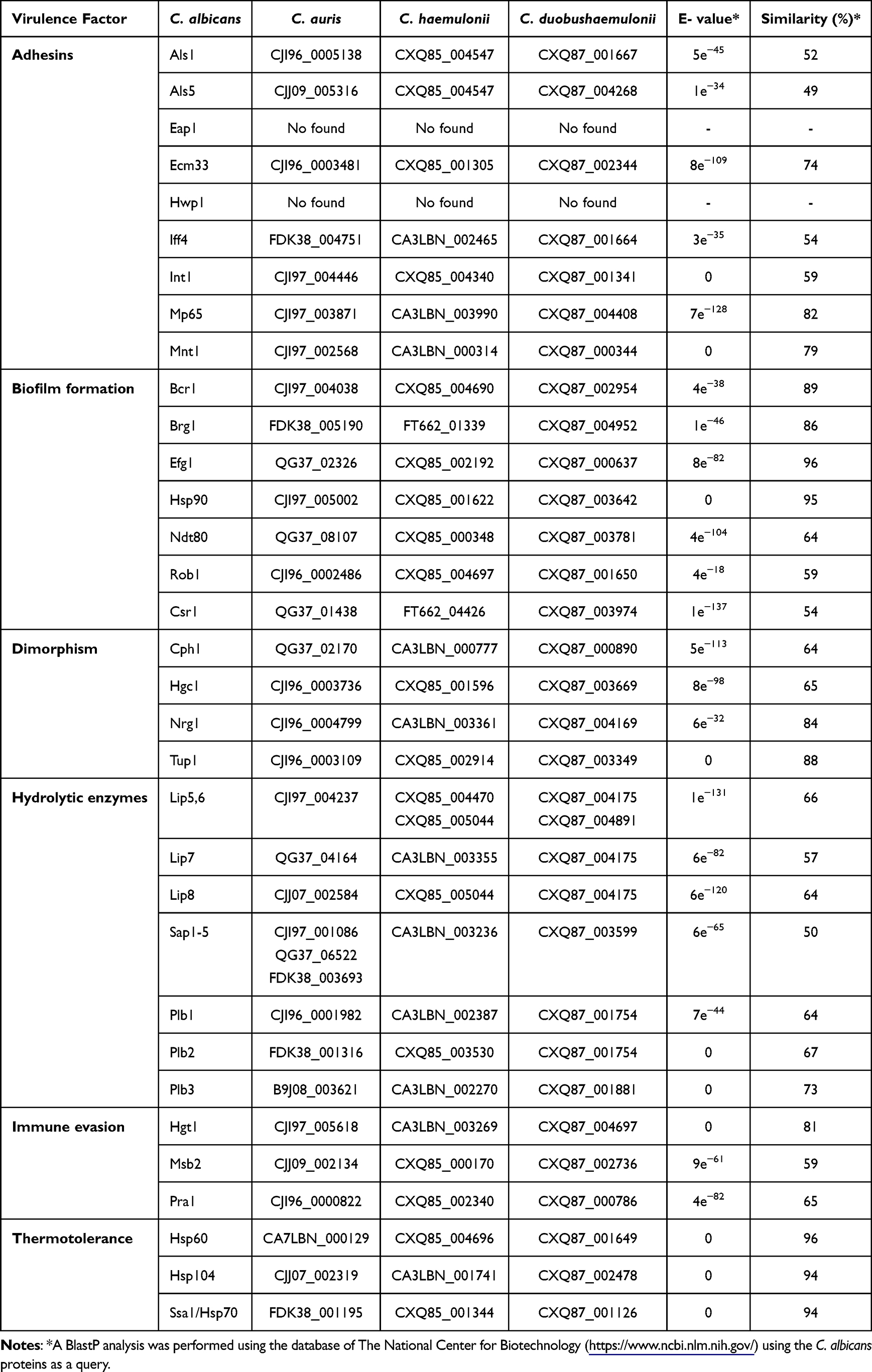 |
Table 1 Putative Virulence Factors in Candida auris, C. haemulonii, and C. duobushaemulonii |
Cell adhesion is an important trait that pathogens have to colonize host cells. Adherence gives pathogens the ability to form microbial communities or biofilms, an important virulence factor of Candida species.103 Biofilms are known to confer increased resistance to antifungal drugs to microorganisms.102,103 Different Candida species have been shown to have their machinery for adhesion, recognition, invasion, and colonization of host cells. In the case of C. auris, several studies have suggested that this species uses different mechanisms for cell adhesion.88 For example, one characteristic of C. auris is its ability to adhere to and persist on abiotic surfaces, such as steel devices in hospital environments, as well as human skin.49,104 In C. haemulonii sensu stricto, it has been reported that it can show variation in the cell wall when there is a phenotypic change between white and pink cells. This variation could be an important strategy used by this pathogen to adhere to the host cell surface, adapt to the environment, and escape the immune response.9
The genome of C. auris, C. haemulonii sensu stricto, C. haemulonii var. vulnera, C. pseudohaemulonii, and C. duobushaemulonii encodes several functional orthologs of C. albicans adhesins (Table 1). In C. auris, it has been reported that agglutinin-like sequence (ALS) proteins play an important role in fungal adherence.105 Previous work has shown that sera containing anti-Als3 antibodies can prevent biofilm formation in C. auris species, corroborating its important role in biofilm formation.106 Als4 is a well-characterized adhesin in C. albicans, and its ortholog is also differentially expressed during filamentous growth in C. auris.92 Although there are no reports for C. haemulonii sensu stricto, C. haemulonii var. vulnera, C. pseudohaemulonii, and C. duobushaemulonii adhesins, bioinformatic predictions indicate that these adhesins could be also found in these species (Table 1). Two orthologs of the ALS family, Als1 and Als5, are also expressed in C. auris biofilms.103 The different adhesins of the ALS family seem to play an important role in adhesion, biofilm formation, and persistence of C. auris in hospital settings.102
The IFF4 gene is highly conserved, and its transcription increases during biofilm production in isolates of C. auris, C. haemulonii sensu stricto, and C. duobushaemulonii.101 As observed during our Blastp analysis, this protein appears to be a functional ortholog of C. albicans Iff4, which is known to be associated with biofilm production and antifungal resistance mechanisms in this species (Table 1).103 Although there are few functional orthologs of the C. albicans adhesins that have been characterized in these three species, through bioinformatic analysis it is possible to hypothesize that they can be found within the genome of these species, so cell adhesion is likely taking place through the adhesins Als1,3-5, Eap1, Ecm33, Iff4, Int1, and Mp65 (Table 1). Interestingly, the Hwp1 adhesin present in C. albicans did not yield any match for any of the five species under analysis (Table 1). This could be because Hwp1 is only expressed in hyphae,107 and it is known that the species of the C. haemulonii complex cannot form true hyphae, and their formation also depends on the strain under study.19
C. albicans forms biofilms with a heterogeneous architecture, which combines the presence of blastoconidia and hyphae that are embedded within the extracellular matrix.108 However, C. auris produces thin biofilms composed of blastoconidia and in some cases pseudohyphae, which are embedded within a limited extracellular matrix.108 Studies of biofilm formation on polystyrene surfaces determined that species of the C. haemulonii complex could form biofilms to different degrees, which exhibit a specific isolate pattern. Through tests with crystal violet and safranin, the presence of biofilms formed only by a network of yeasts was observed.94,109 In the case of C. auris, the biofilms showed lower susceptibility to different antifungal drugs, such as polyenes, azoles, and echinocandins, compared to those of C. albicans. These findings would suggest that other mechanisms in the biofilms of this species are more important than having low biomass or limited extracellular matrix.87,88,110 Although the structure of these biofilms differs from those formed by C. albicans, biofilms formed by species of the C. haemulonii complex also contribute to fungal virulence, antifungal resistance, and survival.14 In C. albicans, biofilm formation is regulated by several genes, such as BCR1, BRG1, EFG1, HSP90, NDT80, ROB1, and CSR1. According to the bioinformatic analysis carried out, these genes could also be found in the species of the C. haemulonii complex and C. auris (Table 1). As has been reported, the three species can form biofilms, which could be an indicator that these genes could be part of this biological process, but could be regulated differently. Nevertheless, it is necessary to experimentally address this hypothesis.
The species of the C. haemulonii complex are found within the members of the Candida genus that are not capable of forming true hyphae, in some cases only pseudohyphae, as has been reported for other species, such as C. lusitaniae.94,111 Dimorphism in Candida species is an important factor that has been related to tissue invasion, pathogenicity, and the virulence factors expression that are morphology specific.112,113 In C. albicans, several transcriptional regulators of dimorphism have been studied, including Cph1, Hgc1, Nrg1, and Tup1.114 According to our bioinformatic analysis, the genome of C. auris, C. haemulonii sensu stricto, C. haemulonii var. vulnera, C. pseudohaemulonii, and C. duobushaemulonii contain possible functional orthologs of these genes (Table 1). Although these five species cannot form true hyphae, the process that controls dimorphism could be regulated by these genes, but with some differences when compared with C. albicans. Yeast morphology also plays an important role in species that cannot make dimorphism, this morphology is the one that is directly involved in fungal spread.112,115
The production of secreted hydrolytic enzymes, such as SAPs, lipases, phospholipases, and hemolysins are important for Candida pathogenicity since they contribute to host adhesion and invasion.101,116 Comparative analyzes of the genome of C. auris with C. albicans and C. dubliniensis found that there is a similar amount of lipases between these species.82 In C. auris, it has been shown that the ability to produce lytic enzymes depends on the different strains and isolates.109 In addition, in vitro studies using different C. auris isolates from different geographical regions showed that 37.5% of the strains had phospholipase-type activity and 64% of these were positive for proteinases.109 Recent studies showed that the levels of SAPs secreted by a C. auris isolate at a temperature of 42 °C were higher than the levels of SAPs in C. albicans at the same temperature.68 These results would indicate that C. auris isolates adapt to temperature stress and can maintain their pathogenicity at high temperatures.101 Several studies have demonstrated that C. auris expresses lytic enzymes, such as SAPs, phospholipases, and hemolysin; however, there is not much information on how the expression profiles of these are compared with the orthologs of C. albicans in vitro and in vivo.67,92,109 In C. auris, it has been reported that filamentous cells produce lower levels of SAPs than yeast cells. The filamentous form could be better adapted to colonize the host skin, and according to this, it is suggested that C. auris is a primary colonizer of the skin, unlike C. albicans, which may be a primary colonizer of the gastrointestinal tract.14,25,92
Comparative studies in a murine model of disseminated infection and a model of Galleria mellonella infection showed that C. auris is less virulent than C. albicans, however, C. auris is more virulent than C. glabrata and C. haemulonii sensu stricto in the same models.101,117 These differences in virulence, when compared to C. albicans, may probably be because C. auris, C. glabrata, and C. haemulonii sensu stricto are unable to develop hyphae in the mammalian hosts.91,92 Through our bioinformatic analysis, it was possible to find the putative orthologs of C. albicans genes that code for phospholipases, and lipases, in the species of the C. haemulonii complex and C. auris (Table 1). The gene products could be used to carry out specific functions, such as invasion of host cells and evasion of the immune response, as has been reported in C. albicans.
Other virulence factors that play an important role in Candida species are thermotolerance and immune evasion.14 Thermotolerance is a characteristic that contributes to the persistence and survival of C. auris on biotic and abiotic surfaces for a long time.14,103,118 C. auris is thermotolerant because it grows optimally at 37 °C, but it can remain viable at 42 °C. However, C. haemulonii and C. duobushaemulonii cannot grow at this temperature.4,91 The C. auris thermotolerance gives the option to cause invasive candidiasis, including tolerance to fever.119 This thermotolerance is believed to be related to climate change and global temperature changes, and this pathogen could be the first example of a fungus emerging from human-induced global warming.120 Bioinformatic analysis suggests that the genome of C. auris, C. haemulonii sensu stricto, C. haemulonii var. vulnera, C. pseudohaemulonii, and C. duobushaemulonii contains orthologs of genes involved in thermotolerance, such as HSP60, HSP70, HSP90, and HSP104, which code for heat shock proteins (Table 1). Hsp90 is a chaperone that controls temperature-dependent filamentation in C. albicans, and in C. auris, it was reported that treatment with an Hsp90 inhibitor resulted in the formation of pseudohyphae.121 However, when the treatment was administered to C. albicans, it showed a filamentous growth, which could suggest that in both species certain filamentation mechanisms are conserved.14
In the case of immune evasion, C. albicans uses different strategies, which involve biofilm formation, protease production, morphological changes, and protein synthesis.122 In C. albicans, there are three genes involved in the immune evasion process, named HGT1, MSB2, and PRA1. In the C. haemulonii complex and C. auris possible orthologs of these genes were found (Table 1). These results could indicate that both the mechanisms of thermotolerance and immune evasion in these non-albicans species could be similar to the mechanisms used by C. albicans. However, more studies are required to corroborate these hypotheses. For the species C. vulturna its genome is not yet available, therefore it was not possible to carry out comparative Blastp analyzes.
Immune Response Against C. auris and the C. haemulonii Complex
Taking into account the importance of new emerging pathogens such as C. auris, C. haemulonii sensu stricto, C. haemulonii var. vulnera, C. vulturna, C. pseudohaemulonii, and C. duobushaemulonii, it is fundamental to understand the host’s defense mechanisms and the fungal strategies to evade immunity. The mechanisms related to the immune sensing of these species after infection are still not fully understood. However, recent works have shed some light on this subject. The immune response against fungi is based on two classical immune branches, the innate and adaptive responses. Innate immune cells, such as monocytes, neutrophils, and macrophages recognize Candida cells through pattern recognition receptors (PRRs), which interact with pathogen-associated molecular patterns (PAMPs). These PAMPs are, in most cases, cell wall components such as β-glucans, chitin, mannose-based glycans, named mannans, and phospholipomannan.95,123 For adaptive immunity, which contributes to the long-term host protection against Candida infection, the release of immunoglobulins, and activation of T cells are essential events, along with antigen presentation from myeloid cells.124
Thus far, anti-Candida immunity has been thoroughly studied in C. albicans. However, recent progress has been reported in the study of C. auris immune sensing. It is known that most of the C. auris strains that are resistant to different antifungal drugs show susceptibility when interacting with the antimicrobial peptide histatin 5, which is known to have significant antifungal activity against C. albicans.125 Immune response cells, such as neutrophils, play an important role in controlling Candida infections, via the release of neutrophil extracellular traps (NETs).126 To determine whether these cells effectively kill C. auris, fungal viability was measured in a time-course experiment.127 After 4 h of interaction, a 75% growth inhibition of the reference species C. albicans was observed; however, C. auris was not affected.127 In addition, it was also documented that human neutrophils were not capable of recruiting C. auris cells.127 Previous studies have shown that C. auris has a higher tolerance to oxidative stress than C. albicans, which may be correlated with survival within neutrophils if engulfed.125
It has been reported that species such as C. albicans, C. tropicalis, C. guilliermondii, C. krusei, and C. auris can differentially stimulate cytokine production by human peripheral blood mononuclear cells (PBMCs).21 Incubation of yeast cells with human PBMCs and quantification of secreted cytokines determined that C. auris and C. albicans are barely able to stimulate cytokine production, such as TNFα, IL-6, IL-1β, and IL-10.21 For C. albicans, it had already been reported that heat-inactivated cells stimulated the production of higher levels of TNFα, IL-6, IL-1β, and IL-10 than live cells. In the case of C. auris, it was reported that similar to C. albicans exposure of inner wall layers by heat inactivation positively affected the ability to stimulate both pro- and anti-inflammatory cytokines.21 When interacting with human monocyte-derived macrophages, both C. auris and C. albicans were poorly phagocytosed, when compared with the ability of the human cells to uptake C. tropicalis, C. guilliermondii, and C. krusei yeast cells.21 However, its interaction with neutrophils is different from that shown by C. albicans, which could suggest that once C. auris evades the neutrophil response, the next defensive line, including PBMCs and macrophages, is activated and could control the entry of this pathogen. However, more experiments are needed to help reinforce this hypothesis.128
Recently, in vivo and in vitro studies were carried out with the C. auris isolate BJCA001, to elucidate more about the immune response against this pathogen.129 To study the possibility that C. auris cells could be efficiently eliminated by the host’s innate immune response, immunocompetent female C57BL/6 mice were used, which were intravenously infected with yeast cells of C. auris BJCA001. In this experiment, the fungal load was also determined in different mice organs, such as the kidney, spleen, and brain. The results suggested that after infection with the fungus, the yeast cells could remain in the host, avoiding being recognized and eliminated by the innate immune system.129 In addition, when the fungal load in the different murine organs was determined, abundant tissue colonization by yeast cells was found; however, they did not undergo dimorphism.129 Inflammation and tissue damage were less severe than that observed in mice infected with C. albicans. From these experiments, it was hypothesized that C. auris could be developing an uncharacterized immune evasion strategy to combat destruction by immune cells. To test it, in vitro studies were carried out, examining the expression patterns of different proinflammatory cytokines in murine bone marrow-derived macrophages, which were treated with live cells of either C. auris isolate BJCA001 or C. albicans.129 It was observed that the expression levels of different proinflammatory cytokines, such as IL-1β, IL-6, TNF-α, CXCL1, and CXCL2 were significantly upregulated after stimulation by C. albicans; however, the expression of these was modest when immune cells interacted with C. auris. These results could suggest that C. auris, compared to C. albicans, is a less potent inducer of the MAPK signaling pathway, which plays an important role in controlling the expression of proinflammatory cytokines in macrophages.129 The reduction of the proinflammatory processes by C. auris could also be related to the exposure of the cell wall’s outer layer, which seems to be different. This mannan-rich outer layer has been reported to help mask the inner β-1,3-glucan layer, thus, it is not easily recognized by host immune cell receptors.129 Such masking could play a key role in protecting C. auris. Other Candida species, such as C. albicans and C. glabrata, are known to use this mechanism to evade the host attack.130–132
In vitro adhesion assays, using different types of human epithelial cells, such as human skin keratinocyte cells, and umbilical vein endothelial cells showed that C. auris has a reduced cell adhesion activity, compared to C. albicans.129 However, when the experiment was carried out in vivo using murine skin, it was observed that C. auris was able to colonize the skin surface. The reason for these differences in both experiments is not clear, but the authors propose that it could be related to the structural variations of the monocultures and skin, in addition to the fact that C. auris could have different growth rates in the different experimental conditions.129
For a long time, C. auris, unlike C. albicans, has been recognized as a pathogen that does not tend to undergo cell dimorphism.4,29,91 However, the isolate BJCA001 can transit from yeast to filamentous cells.92 When the adhesion capacity was analyzed in both types of cells, no significant differences were found in cell adhesion, nor changes in cytokine production, and in the same way as reported for yeasts, the filamentous cells also failed to induce MAPK pathway activation.129 Therefore, the C. auris innate immune evasion is morphology-independent.
The immune response against C. haemulonii sensu stricto and C. duobushaemulonii has not been studied as that described for C. auris; however, recent works have elucidated some characteristics of the C. haemulonii sensu stricto immune response.93 To understand this process, gene expression analyzes were carried out to observe the host immune response against this pathogen, using the zebrafish as an experimental model. Changes in gene expression levels in animals infected with C. haemulonii sensu stricto and C. auris species were compared with C. albicans.93 It was found that most pro- and anti-inflammatory cytokine expression genes were upregulated in infection caused by C. haemulonii sensu stricto and C. auris. However, changes in the expression levels of the different cytokines genes (TNF-α, IL-8, IL-10) were found in the early times of infection with C. auris, but in the case of C. haemulonii sensu stricto those genes showed the top expression at late times of infection.93 The activation of IL-17α, which is related to the neutrophils recruitment, showed a slow response once the infection by C. auris was carried out, in the same way, a significant reduction of IL-8 and leukocyte myeloperoxidase was observed in the later phase of infection by this pathogen.93 These results suggest that the function of neutrophils in a host infected with C. auris may be diminished, due to a lower ability to recognize this pathogen, as has been reported in previous works.93,127,133 In animals infected with C. haemulonii sensu stricto or C. albicans, neutrophils readily recognize both species. These findings could explain part of the virulence of C. auris and C. haemulonii sensu stricto, and how they are recognized by host immune cells.93 Experiments carried out in immunocompetent mice and Galleria mellonella larvae showed that C. haemulonii sensu stricto is a fungus of low virulence, compared to C. albicans and C. auris.102,117,134
When evaluating the expressions of matrix metalloproteinases as possible mediators of leukocyte recruitment, it was found that the genes of these metalloproteinases were significantly regulated in zebrafish infected with C. haemulonii sensu stricto or C. auris.135 In the case of MMP9, the animal group that was infected with C. auris showed a high expression in the late infection phase, while animals infected with C. haemulonii sensu stricto increased the expression in the early phase.93 Regarding JAK2 expression, it was increased in the animal group infected with C. auris at the early time point and NF-κβ, a key transcription factor for proinflammatory cytokine production, showed higher expressions after C. auris and C. haemulonii sensu stricto infection, than with C. albicans.93 Because of the importance of the development and function of regulatory T cells during the immune response, the expression of two genes that are involved in this process, FOXP3a and FOXP3b, was determined.136 C. haemulonii sensu stricto infection revealed that the expression of these genes had similar levels to C. albicans infection; however, C. auris infection led to significantly higher expression of both genes.93 A different time point of gene expression was found between the different species, for C. auris it increased at the early time point, but for C. haemulonii sensu stricto, it increased at the late time point.93 Based on these results, it could be inferred that C. auris, unlike C. haemulonii sensu stricto, could reduce the number of regulatory T cells at the late time point and thus increase the yeast population to continue the infectious process.
Although C. auris and C. haemulonii sensu stricto have a close phylogenetic relationship, the immune response seems to be different. Regarding C. duobushaemulonii, C. pseudohaemulonii, and C. vulturna, there are currently no reports on how the pathogen-host interaction takes place. Even though these species are phylogenetically related to C. auris, it is not feasible to extrapolate the information about the immune response of this species to other ones, because it is clear that this crosstalk with the host immunity is species-specific.
Conclusions
In recent years, the epidemiology of infections caused by Candida has changed, and the emergency of non-albicans species has been increasing, as is the case of C. auris, C. haemulonii sensu stricto, C. haemulonii var. vulnera, C. vulturna, C. pseudohaemulonii, and C. duobushaemulonii. These have been recognized as pathogens of concern, presenting intrinsic resistance to commonly used antifungal drugs. The infections caused by these species, especially C. auris, which is the most studied, can be fatal in immunocompromised patients.
The use of bioinformatics tools has become a key strategy to understand the biology of neglected organisms, such as the C. haemulonii complex and C. auris. These tools allow the predictions of genes and gene products, which can be used to build up working models to establish differences and similarities in virulence factors in species mentioned in this work.
From the information revised, it was evident that more information and studies are needed on the C. haemulonii complex and C. auris, especially on species identification and immune response. These could be opportunity areas to develop new methodologies that allow us to easily distinguish between species and learn more about pathogen-host interaction. Knowledge of resistance to one or more classes of antifungals can be of great help in selecting a therapy to eliminate the causative agent of mycosis, for which, the correct identification of the pathogen and the determination of antifungal susceptibility is paramount. It is also vital to avoid the administration of the drug in subtherapeutic doses and for prolonged periods, as these are factors that contribute to the development of drug resistance.
Acknowledgments
This work was supported by Consejo Nacional de Ciencia y Tecnología (ref. FC 2015-02-834 and CF-2019-6380), and Red Temática Glicociencia en Salud (CONACYT-México).
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Disclosure
The authors declare no conflicts of interest in this work.
References
1. Brown GD, Denning DW, Gow NAR, Levitz SM, Netea MG, White TC. Hidden killers: human fungal infections. Sci Transl Med. 2012;4(165):165rv13. doi:10.1126/scitranslmed.3004404
2. Fisher MC, Gurr SJ, Cuomo CA, et al. Threats posed by the fungal kingdom to humans, wildlife, and agriculture. mBio. 2020;11(3):e00449–20. doi:10.1128/mBio.00449-20
3. Pappas PG, Lionakis MS, Arendrup MC, Ostrosky-Zeichner L, Kullberg BJ. Invasive candidiasis. Nat Rev Dis Primers. 2018;4(1):18026. doi:10.1038/nrdp.2018.26
4. Satoh K, Makimura K, Hasumi Y, Nishiyama Y, Uchida K, Yamaguchi H. Candida auris sp. nov., a novel ascomycetous yeast isolated from the external ear canal of an inpatient in a Japanese hospital. Microbiol Immunol. 2009;53(1):41–44. doi:10.1111/j.1348-0421.2008.00083.x
5. Cendejas-Bueno E, Kolecka A, Alastruey-Izquierdo A, et al. Reclassification of the Candida haemulonii complex as Candida haemulonii (C. haemulonii group I), C. duobushaemulonii sp. nov. (C. haemulonii group II), and C. haemulonii var. vulnera var. nov.: three multiresistant human pathogenic yeasts. J Clin Microbiol. 2012;50(11):3641–3651. doi:10.1128/jcm.02248-12
6. Lima SL, Francisco EC, de Almeida Júnior JN, et al. Increasing prevalence of multidrug-resistant Candida haemulonii species complex among all yeast cultures collected by a reference laboratory over the past 11 Years. J Fungi. 2020;6(3). doi:10.3390/jof6030110
7. Khan ZU, Al-Sweih NA, Ahmad S, et al. Outbreak of fungemia among neonates caused by Candida haemulonii resistant to amphotericin B, itraconazole, and fluconazole. J Clin Microbiol. 2007;45(6):2025–2027. doi:10.1128/jcm.00222-07
8. Ramos LS, Figueiredo-Carvalho MH, Barbedo LS, et al. Candida haemulonii complex: species identification and antifungal susceptibility profiles of clinical isolates from Brazil. J Antimicrob Chemother. 2015;70(1):111–115. doi:10.1093/jac/dku321
9. Deng Y, Li S, Bing J, Liao W, Tao L. Phenotypic switching and filamentation in Candida haemulonii, an emerging opportunistic pathogen of humans. Microbiol Spectr. 2021;9(3):e0077921. doi:10.1128/Spectrum.00779-21
10. Hou X, Xiao M, Chen SC-A, et al. Identification and antifungal susceptibility profiles of Candida haemulonii species complex clinical isolates from a multicenter study in China. J Clin Microbiol. 2016;54(11):2676–2680. doi:10.1128/jcm.01492-16
11. Ramos LS, Figueiredo-Carvalho MHG, Silva LN, et al. The threat called Candida haemulonii species complex in Rio de Janeiro State, Brazil: focus on antifungal resistance and virulence attributes. J Fungi. 2022;8(6):574. doi:10.3390/jof8060574
12. Frías-de-león MG, Martínez-Herrera E, Acosta-Altamirano G, Arenas R, Rodríguez-Cerdeira C. Superficial candidosis by Candida duobushaemulonii: an emerging microorganism. Infect Genet Evol. 2019;75:103960. doi:10.1016/j.meegid.2019.103960
13. Jeffery-Smith A, Taori SK, Schelenz S, et al. Candida auris: a review of the literature. Clin Microbiol Rev. 2018;31(1):e00029–17. doi:10.1128/cmr.00029-17
14. Du H, Bing J, Hu T, Ennis CL, Nobile CJ, Huang G. Candida auris: epidemiology, biology, antifungal resistance, and virulence. PLoS Pathog. 2020;16(10):e1008921. doi:10.1371/journal.ppat.1008921
15. Oh BJ, Shin JH, Kim MN, et al. Biofilm formation and genotyping of Candida haemulonii, Candida pseudohaemulonii, and a proposed new species (Candida auris) isolates from Korea. Med Mycol. 2011;49(1):98–102. doi:10.3109/13693786.2010.493563
16. Sherry L, Ramage G, Kean R, et al. Biofilm-forming capability of highly virulent, multidrug-resistant Candida auris. Emerg Infect Dis. 2017;23(2):328–331. doi:10.3201/eid2302.161320
17. Chowdhary A, Sharma C, Duggal S, et al. New clonal strain of Candida auris, Delhi, India. Emerg Infect Dis. 2013;19(10):1670–1673. doi:10.3201/eid1910.130393
18. Kathuria S, Singh PK, Sharma C, et al. Multidrug-resistant Candida auris misidentified as Candida haemulonii: characterization by Matrix-Assisted Laser Desorption Ionization-Time of Flight Mass Spectrometry and DNA Sequencing and Its antifungal susceptibility profile variability by Vitek 2, CLSI broth microdilution, and Etest method. J Clin Microbiol. 2015;53(6):1823–1830. doi:10.1128/jcm.00367-15
19. Healey KR, Kordalewska M, Jiménez Ortigosa C, et al. Limited ERG11 mutations identified in isolates of Candida auris directly contribute to reduced azole susceptibility. Antimicrob Agents Chemother. 2018;62(10). doi:10.1128/aac.01427-18
20. Sugita T, Takashima M, Poonwan N, Mekha N. Candida pseudohaemulonii Sp. Nov., an amphotericin B-and azole-resistant yeast species, isolated from the blood of a patient from Thailand. Microbiol Immunol. 2006;50(6):469–473. doi:10.1111/j.1348-0421.2006.tb03816.x
21. Navarro-Muñoz JC, de Jong AW, Gerrits van den Ende B, et al. The high-quality complete genome sequence of the opportunistic fungal pathogen Candida vulturna CBS 14366(T). Mycopathologia. 2019;184(6):731–734. doi:10.1007/s11046-019-00404-0
22. Gade L, Muñoz JF, Sheth M, et al. Understanding the emergence of multidrug-resistant Candida: using whole-genome sequencing to describe the population structure of Candida haemulonii species complex. Original Research. Front Genet. 2020;11:554. doi:10.3389/fgene.2020.00554
23. Muthusamy A, Rao M, Chakrabarti A, Velayuthan RD. Case report: catheter related blood stream infection caused by Candida vulturna. Med Mycol Case Rep. 2022;36:27–30. doi:10.1016/j.mmcr.2022.04.001
24. Lee WG, Shin JH, Uh Y, et al. First three reported cases of nosocomial fungemia caused by Candida auris. J Clin Microbiol. 2011;49(9):3139–3142. doi:10.1128/jcm.00319-11
25. Lockhart SR, Etienne KA, Vallabhaneni S, et al. Simultaneous emergence of multidrug-resistant Candida auris on 3 continents confirmed by whole-genome sequencing and epidemiological analyses. Clin Infect Dis. 2017;64(2):134–140. doi:10.1093/cid/ciw691
26. Lamoth F, Kontoyiannis DP. The Candida auris alert: facts and perspectives. J Infect Dis. 2018;217(4):516–520. doi:10.1093/infdis/jix597
27. Chowdhary A, Sharma C, Meis JF. Candida auris: a rapidly emerging cause of hospital-acquired multidrug-resistant fungal infections globally. PLoS Pathog. 2017;13(5):e1006290. doi:10.1371/journal.ppat.1006290
28. Khan Z, Ahmad S. Candida auris: an emerging multidrug-resistant pathogen of global significance. Curr Med Res Pract. 2017;7(6):240–248. doi:10.1016/j.cmrp.2017.11.004
29. Saris K, Meis JF, Voss A. Candida auris. Curr Opin Infect Dis. 2018;31(4):334–340. doi:10.1097/qco.0000000000000469
30. Zhu Y, O’Brien B, Leach L, et al. Laboratory analysis of an outbreak of Candida auris in New York from 2016 to 2018: impact and lessons learned. J Clin Microbiol. 2020;58(4):e01503–19. doi:10.1128/jcm.01503-19
31. Chowdhary A, Anil Kumar V, Sharma C, et al. Multidrug-resistant endemic clonal strain of Candida auris in India. Eur J Clin Microbiol Infect Dis. 2014;33(6):919–926. doi:10.1007/s10096-013-2027-1
32. Ahmad S, Khan Z, Al-Sweih N, Alfouzan W, Joseph L. Candida auris in various hospitals across Kuwait and their susceptibility and molecular basis of resistance to antifungal drugs. Mycoses. 2020;63(1):104–112. doi:10.1111/myc.13022
33. Schelenz S, Hagen F, Rhodes JL, et al. First hospital outbreak of the globally emerging Candida auris in a European hospital. Antimicrob Resist Infect Control. 2016;5:35. doi:10.1186/s13756-016-0132-5
34. Govender NP, Magobo RE, Mpembe R, et al. Candida auris in South Africa, 2012–2016. Emerg Infect Dis. 2018;24(11):2036–2040. doi:10.3201/eid2411.180368
35. Rhodes J, Abdolrasouli A, Farrer RA, et al. Genomic epidemiology of the UK outbreak of the emerging human fungal pathogen Candida auris. Emerg Microbes Infect. 2018;7(1):43. doi:10.1038/s41426-018-0045-x
36. Ruiz-Gaitán A, Moret AM, Tasias-Pitarch M, et al. An outbreak due to Candida auris with prolonged colonisation and candidaemia in a tertiary care European hospital. Mycoses. 2018;61(7):498–505. doi:10.1111/myc.12781
37. Adam RD, Revathi G, Okinda N, et al. Analysis of Candida auris fungemia at a single facility in Kenya. Int J Infect Dis. 2019;85:182–187. doi:10.1016/j.ijid.2019.06.001
38. Armstrong PA, Rivera SM, Escandon P, et al. Hospital-associated multicenter outbreak of emerging fungus Candida auris, Colombia, 2016. Emerg Infect Dis. 2019;25(7):1339–1346. doi:10.3201/eid2507.180491
39. Barantsevich NE, Vetokhina AV, Ayushinova NI, Orlova OE, Barantsevich EP. Candida auris bloodstream infections in Russia. Antibiotics. 2020;9(9):557. doi:10.3390/antibiotics9090557
40. Alfouzan W, Ahmad S, Dhar R, et al. Molecular epidemiology of Candida auris outbreak in a major secondary-care hospital in Kuwait. J Fungi. 2020;6(4):307. doi:10.3390/jof6040307
41. Farooqi JQ, Soomro AS, Baig MA, et al. Outbreak investigation of Candida auris at a tertiary care hospital in Karachi, Pakistan. J Infect Prev. 2020;21(5):189–195. doi:10.1177/1757177420935639
42. Alshamrani MM, El-Saed A, Mohammed A, et al. Management of Candida auris outbreak in a tertiary-care setting in Saudi Arabia. Infect Control Hosp Epidemiol. 2021;42(2):149–155. doi:10.1017/ice.2020.414
43. Eckbo EJ, Wong T, Bharat A, et al. First reported outbreak of the emerging pathogen Candida auris in Canada. Am J Infect Control. 2021;49(6):804–807. doi:10.1016/j.ajic.2021.01.013
44. Villanueva-Lozano H, Treviño-Rangel RJ, González GM, et al. Outbreak of Candida auris infection in a COVID-19 hospital in Mexico. Clin Microbiol Infect. 2021;27(5):813–816. doi:10.1016/j.cmi.2020.12.030
45. Arensman K, Miller JL, Chiang A, et al. Clinical outcomes of patients treated for Candida auris infections in a multisite health system, Illinois, USA. Emerg Infect Dis. 2020;26(5):876–880. doi:10.3201/eid2605.191588
46. Al Maani A, Paul H, Al-Rashdi A, et al. Ongoing challenges with healthcare-associated Candida auris outbreaks in Oman. J Fungi. 2019;5(4):101. doi:10.3390/jof5040101
47. Mohsin J, Weerakoon S, Ahmed S, et al. A cluster of Candida auris blood stream infections in a tertiary care hospital in Oman from 2016 to 2019. Antibiotics. 2020;9(10):638. doi:10.3390/antibiotics9100638
48. Mulet Bayona JV, Tormo Palop N, Salvador García C, et al. Characteristics and management of candidaemia episodes in an established Candida auris outbreak. Antibiotics. 2020;9(9):558. doi:10.3390/antibiotics9090558
49. Welsh RM, Bentz ML, Shams A, et al. Survival, persistence, and isolation of the emerging multidrug-resistant pathogenic yeast Candida auris on a plastic health care surface. J Clin Microbiol. 2017;55(10):2996–3005. doi:10.1128/jcm.00921-17
50. Kumar J, Eilertson B, Cadnum JL, et al. Environmental contamination with Candida species in multiple hospitals including a tertiary care hospital with a Candida auris outbreak. Pathog Immun. 2019;4(2):260–270. doi:10.20411/pai.v4i2.291
51. Chen J, Tian S, Han X, et al. Is the superbug fungus really so scary? A systematic review and meta-analysis of global epidemiology and mortality of Candida auris. BMC Infect Dis. 2020;20(1):827. doi:10.1186/s12879-020-05543-0
52. Chaabane F, Graf A, Jequier L, Coste AT. Review on antifungal resistance mechanisms in the emerging pathogen Candida auris. Front Microbiol. 2019;10:2788. doi:10.3389/fmicb.2019.02788
53. Kim S, Ko KS, Moon SY, Lee MS, Lee MY, Son JS. Catheter-related candidemia caused by Candida haemulonii in a patient in long-term hospital care. J Korean Med Sci. 2011;26(2):297–300. doi:10.3346/jkms.2011.26.2.297
54. Almeida JN, Motta AL, Rossi F, et al. First report of a clinical isolate of Candida haemulonii in Brazil. Clinics. 2012;67(10):1229–1231. doi:10.6061/clinics/2012(10)18
55. Kim MN, Shin JH, Sung H, et al. Candida haemulonii and closely related species at 5 university hospitals in Korea: identification, antifungal susceptibility, and clinical features. Clin Infect Dis. 2009;48(6):e57–e61. doi:10.1086/597108
56. Dewaele K, Lagrou K, Frans J, Hayette MP, Vernelen K. Hospital laboratory survey for identification of Candida auris in Belgium. J Fungi. 2019;5(3):84. doi:10.3390/jof5030084
57. Rodero L, Cuenca-Estrella M, Córdoba S, et al. Transient fungemia caused by an amphotericin B-resistant isolate of Candida haemulonii. J Clin Microbiol. 2002;40(6):2266–2269. doi:10.1128/jcm.40.6.2266-2269.2002
58. Giusiano G, Mangiaterra M, Garcia Saito V, Rojas F, Gómez V, Díaz MC. Fluconazole and itraconazole resistance of yeasts isolated from the bloodstream and catheters of hospitalized pediatric patients. Chemotherapy. 2006;52(5):254–259. doi:10.1159/000094867
59. Reséndiz-Sánchez J, Ortiz-álvarez J, Casimiro-Ramos A, Hernández-Rodríguez C, Villa-Tanaca L. First report of a catheter-related bloodstream infection by Candida haemulonii in a children’s hospital in Mexico City. Int J Infect Dis. 2020;92:123–126. doi:10.1016/j.ijid.2019.12.037
60. de Almeida JN, Assy JG, Levin AS, et al. Candida haemulonii complex species, Brazil, January 2010-March 2015. Emerg Infect Dis. 2016;22(3):561–563. doi:10.3201/eid2203.151610
61. Kumar A, Prakash A, Singh A, et al. Candida haemulonii species complex: an emerging species in India and its genetic diversity assessed with multilocus sequence and amplified fragment-length polymorphism analyses. Emerg Microbes Infect. 2016;5(5):e49. doi:10.1038/emi.2016.49
62. Guarana M, Nucci M. Acute disseminated candidiasis with skin lesions: a systematic review. Clin Microbiol Infect. 2018;24(3):246–250. doi:10.1016/j.cmi.2017.08.016
63. Goswami R, Dadhwal V, Tejaswi S, et al. Species-specific prevalence of vaginal candidiasis among patients with diabetes mellitus and its relation to their glycaemic status. J Infect. 2000;41(2):162–166. doi:10.1053/jinf.2000.0723
64. Ruan SY, Kuo YW, Huang CT, Hsiue HC, Hsueh PR. Infections due to Candida haemulonii: species identification, antifungal susceptibility and outcomes. Int J Antimicrob Agents. 2010;35(1):85–88. doi:10.1016/j.ijantimicag.2009.08.009
65. Oberoi JK, Wattal C, Goel N, Raveendran R, Datta S, Prasad K. Non-albicans Candida species in blood stream infections in a tertiary care hospital at New Delhi, India. Indian J Med Res Dec. 2012;136(6):997–1003.
66. Yuvaraj A, Rohit A, Koshy PJ, Nagarajan P, Nair S, Abraham G. Rare occurrence of fatal Candida haemulonii peritonitis in a diabetic CAPD patient. Ren Fail. 2014;36(9):1466–1467. doi:10.3109/0886022x.2014.944067
67. Kumar D, Banerjee T, Pratap CB, Tilak R. Itraconazole-resistant Candida auris with phospholipase, proteinase and hemolysin activity from a case of vulvovaginitis. J Infect Dev Ctries. 2015;9(4):435–437. doi:10.3855/jidc.4582
68. Wang X, Bing J, Zheng Q, et al. The first isolate of Candida auris in China: clinical and biological aspects. Emerg Microbes Infect. 2018;7(1):93. doi:10.1038/s41426-018-0095-0
69. Ahmad S, Alfouzan W. Candida auris: epidemiology, diagnosis, pathogenesis, antifungal susceptibility, and infection control measures to combat the spread of infections in healthcare facilities. Microorganisms. 2021;9(4):807. doi:10.3390/microorganisms9040807
70. Jurado-Martín I, Marcos-Arias C, Tamayo E, et al. Candida duobushaemulonii: an old but unreported pathogen. J Fungi. 2020;6(4). doi:10.3390/jof6040374
71. de Jong AW, Dieleman C, Carbia M, Mohd Tap R, Hagen F. Performance of two novel chromogenic media for the identification of multidrug-resistant Candida auris compared with other commercially available. J Clin Microbiol. 2021;59(4):e03220. doi:10.1128/jcm.03220-20
72. Fang SY, Wei KC, Chen WC, et al. Primary deep cutaneous candidiasis caused by Candida duobushaemulonii in a 68-year-old man: the first case report and literature review. Mycoses. 2016;59(12):818–821. doi:10.1111/myc.12540
73. Sipiczki M, Tap RM. Candida vulturna pro tempore sp. nov., a dimorphic yeast species related to the Candida haemulonis species complex isolated from flowers and clinical sample. Int J Syst Evol Microbiol. 2016;66(10):4009–4015. doi:10.1099/ijsem.0.001302
74. Arendrup MC, Patterson TF. Multidrug-resistant Candida: epidemiology, molecular mechanisms, and treatment. J Infect Dis. 2017;216(suppl_3):S445–S451. doi:10.1093/infdis/jix131
75. Delma FZ, Al-Hatmi AMS, Brüggemann RJM, et al. Molecular mechanisms of 5-fluorocytosine resistance in yeasts and filamentous fungi. J Fungi. 2021;7(11):909. doi:10.3390/jof7110909
76. Pristov KE, Ghannoum MA. Resistance of Candida to azoles and echinocandins worldwide. Clin Microbiol Infect. 2019;25(7):792–798. doi:10.1016/j.cmi.2019.03.028
77. Marak MB, Dhanashree B. Antifungal susceptibility and biofilm production of Candida spp. isolated from clinical samples. Int J Microbiol. 2018;2018:7495218. doi:10.1155/2018/7495218
78. Vallabhaneni S, Kallen A, Tsay S, et al. Investigation of the first seven reported cases of Candida auris, a globally emerging invasive, multidrug-resistant fungus-United States, May 2013-August 2016. Am J Transplant. 2017;17(1):296–299. doi:10.1111/ajt.14121
79. Chowdhary A, Voss A, Meis JF. Multidrug-resistant Candida auris: ‘new kid on the block’ in hospital-associated infections? J Hosp Infect. 2016;94(3):209–212. doi:10.1016/j.jhin.2016.08.004
80. Perea S, López-Ribot JL, Kirkpatrick WR, et al. Prevalence of molecular mechanisms of resistance to azole antifungal agents in Candida albicans strains displaying high-level fluconazole resistance isolated from human immunodeficiency virus-infected patients. Antimicrob Agents Chemother. 2001;45(10):2676–2684. doi:10.1128/aac.45.10.2676-2684.2001
81. Chowdhary A, Prakash A, Sharma C, et al. A multicentre study of antifungal susceptibility patterns among 350 Candida auris isolates (2009–17) in India: role of the ERG11 and FKS1 genes in azole and echinocandin resistance. J Antimicrob Chemother. 2018;73(4):891–899. doi:10.1093/jac/dkx480
82. Muñoz JF, Gade L, Chow NA, et al. Genomic insights into multidrug-resistance, mating and virulence in Candida auris and related emerging species. Nat Commun. 2018;9(1):5346. doi:10.1038/s41467-018-07779-6
83. Feng W, Yang J, Xi Z, et al. Regulatory role of ERG3 and Efg1 in azoles-resistant strains of Candida albicans isolated from patients diagnosed with vulvovaginal candidiasis. Indian J Microbiol. 2019;59(4):514–524. doi:10.1007/s12088-019-00833-x
84. Suwunnakorn S, Wakabayashi H, Kordalewska M, Perlin DS, Rustchenko E. FKS2 and FKS3 genes of opportunistic human pathogen Candida albicans influence echinocandin susceptibility. Antimicrob Agents Chemother. 2018;62(4):e02299–17. doi:10.1128/aac.02299-17
85. Kordalewska M, Perlin DS. Identification of drug resistant Candida auris. Front Microbiol. 2019;10:1918. doi:10.3389/fmicb.2019.01918
86. Ramage G, Bachmann S, Patterson TF, Wickes BL, López-Ribot JL. Investigation of multidrug efflux pumps in relation to fluconazole resistance in Candida albicans biofilms. J Antimicrob Chemother. 2002;49(6):973–980. doi:10.1093/jac/dkf049
87. Sharma C, Kumar N, Pandey R, Meis JF, Chowdhary A. Whole genome sequencing of emerging multidrug resistant Candida auris isolates in India demonstrates low genetic variation. New Microbes New Infect. 2016;13:77–82. doi:10.1016/j.nmni.2016.07.003
88. Chatterjee S, Alampalli SV, Nageshan RK, Chettiar ST, Joshi S, Tatu US. Draft genome of a commonly misdiagnosed multidrug resistant pathogen Candida auris. BMC Genomics. 2015;16(1):686. doi:10.1186/s12864-015-1863-z
89. Jallow S, Govender NP. Ibrexafungerp: a first-in-class oral triterpenoid glucan synthase inhibitor. J Fungi. 2021;7(3):163. doi:10.3390/jof7030163
90. Muro MD, Motta Fde A, Burger M, Melo AS, Dalla-Costa LM. Echinocandin resistance in two Candida haemulonii isolates from pediatric patients. J Clin Microbiol. 2012;50(11):3783–3785. doi:10.1128/jcm.01136-12
91. Borman AM, Szekely A, Johnson EM. Comparative pathogenicity of United Kingdom isolates of the emerging pathogen Candida auris and other key pathogenic Candida species. mSphere. 2016;1(4). doi:10.1128/mSphere.00189-16
92. Yue H, Bing J, Zheng Q, et al. Filamentation in Candida auris, an emerging fungal pathogen of humans: passage through the mammalian body induces a heritable phenotypic switch. Emerg Microbes Infect. 2018;7(1):1–13. doi:10.1038/s41426-018-0187-x
93. Pharkjaksu S, Boonmee N, Mitrpant C, Ngamskulrungroj P. Immunopathogenesis of emerging Candida auris and Candida haemulonii strains. J Fungi. 2021;7(9):725. doi:10.3390/jof7090725
94. Ramos LS, Oliveira SSC, Silva LN, et al. Surface, adhesiveness and virulence aspects of Candida haemulonii species complex. Med Mycol. 2020;58(7):973–986. doi:10.1093/mmy/myz139
95. Mora-Montes HM, Ponce-Noyola P, Villagomez-Castro JC, Gow NA, Flores-Carreon A, Lopez-Romero E. Protein glycosylation in Candida. Future Microbiol. 2009;4(9):1167–1183. doi:10.2217/fmb.09.88
96. Boatto HF, Cavalcanti SD, Del negro GM, et al. Candida duobushaemulonii: an emerging rare pathogenic yeast isolated from recurrent vulvovaginal candidiasis in Brazil. Mem Inst Oswaldo Cruz. 2016;111(6):407–410. doi:10.1590/0074-02760160166
97. Santos MAS, Gomes AC, Santos MC, Carreto LC, Moura GR. The genetic code of the fungal CTG clade. C R Biol. 2011;334(8–9):607–611. doi:10.1016/j.crvi.2011.05.008
98. Bidaud AL, Chowdhary A, Dannaoui E. Candida auris: an emerging drug resistant yeast – a mini-review. J Mycol Med. 2018;28(3):568–573. doi:10.1016/j.mycmed.2018.06.007
99. Zhang H, Niu Y, Tan J, et al. Global screening of genomic and transcriptomic factors associated with phenotype differences between multidrug-resistant and -susceptible Candida haemulonii strains. mSystems. 2019;4(6). doi:10.1128/mSystems.00459-19
100. Rodrigues LS, Gazara RK, Passarelli-Araujo H, et al. First genome sequences of two multidrug-resistant Candida haemulonii var. vulnera isolates from pediatric patients with candidemia. Front Microbiol. 2020;11:1535. doi:10.3389/fmicb.2020.01535
101. Rossato L, Colombo AL. Candida auris: what have we learned about its mechanisms of pathogenicity? Front Microbiol. 2018;9:3081. doi:10.3389/fmicb.2018.03081
102. Chybowska AD, Childers DS, Farrer RA. Nine things genomics can tell us about Candida auris. Front Genet. 2020;11:351. doi:10.3389/fgene.2020.00351
103. Kean R, Delaney C, Sherry L, et al. Transcriptome assembly and profiling of candida auris reveals novel insights into biofilm-mediated resistance. mSphere. 2018;3(4):e00334–18. doi:10.1128/mSphere.00334-18
104. Piedrahita CT, Cadnum JL, Jencson AL, Shaikh AA, Ghannoum MA, Donskey CJ. Environmental surfaces in healthcare facilities are a potential source for transmission of candida auris and other Candida species. Infect Control Hosp Epidemiol. 2017;38(9):1107–1109. doi:10.1017/ice.2017.127
105. Singh S, Uppuluri P, Mamouei Z, et al. The NDV-3A vaccine protects mice from multidrug resistant Candida auris infection. PLoS Pathog. 2019;15(8):e1007460. doi:10.1371/journal.ppat.1007460
106. Pushpakom S, Iorio F, Eyers PA, et al. Drug repurposing: progress, challenges and recommendations. Nat Rev Drug Discov. 2019;18(1):41–58. doi:10.1038/nrd.2018.168
107. Nobile CJ, Nett JE, Andes DR, Mitchell AP. Function of Candida albicans Adhesin Hwp1 in biofilm formation. Eukaryot Cell. 2006;5(10):1604–1610. doi:10.1128/ec.00194-06
108. Nayak AP, Green BJ, Beezhold DH. Fungal hemolysins. Med Mycol. 2013;51(1):1–16. doi:10.3109/13693786.2012.698025
109. Larkin E, Hager C, Chandra J, et al. The emerging pathogen Candida auris: growth phenotype, virulence factors, activity of antifungals, and effect of SCY-078, a novel glucan synthesis inhibitor, on growth morphology and biofilm formation. Antimicrob Agents Chemother. 2017;61(5):e02396–16. doi:10.1128/aac.02396-16
110. Billamboz M, Fatima Z, Hameed S, Jawhara S. Promising drug candidates and new strategies for fighting against the emerging superbug Candida auris. Microorganisms. 2021;9(3):634. doi:10.3390/microorganisms9030634
111. Mendoza-Reyes DF, Gómez-Gaviria M, Mora-Montes HM. Candida lusitaniae: biology, pathogenicity, virulence factors, diagnosis, and treatment. Infect Drug Resist. 2022;15:5121–5135. doi:10.2147/idr.S383785
112. Mayer FL, Wilson D, Hube B. Candida albicans pathogenicity mechanisms. Virulence. 2013;4(2):119–128. doi:10.4161/viru.22913
113. Gómez-Gaviria M, Mora-Montes HM. Current aspects in the biology, pathogeny, and treatment of Candida krusei, a neglected fungal pathogen. Infect Drug Resist. 2020;13:1673–1689. doi:10.2147/idr.S247944
114. Braun BR, Johnson AD. TUP1, CPH1 and EFG1 make independent contributions to filamentation in Candida albicans. Genetics. 2000;155(1):57–67. doi:10.1093/genetics/155.1.57
115. Saville SP, Lazzell AL, Monteagudo C, Lopez-Ribot JL. Engineered control of cell morphology in vivo reveals distinct roles for yeast and filamentous forms of Candida albicans during infection. Eukaryot Cell. 2003;2(5):1053–1060. doi:10.1128/ec.2.5.1053-1060.2003
116. Polke M, Hube B, Jacobsen ID. Candida survival strategies. Adv Appl Microbiol. 2015;91:139–235. doi:10.1016/bs.aambs.2014.12.002
117. Fakhim H, Vaezi A, Dannaoui E, et al. Comparative virulence of Candida auris with Candida haemulonii, Candida glabrata and Candida albicans in a murine model. Mycoses. 2018;61(6):377–382. doi:10.1111/myc.12754
118. Biswal M, Rudramurthy SM, Jain N, et al. Controlling a possible outbreak of Candida auris infection: lessons learnt from multiple interventions. J Hosp Infect. 2017;97(4):363–370. doi:10.1016/j.jhin.2017.09.009
119. Jackson BR, Chow N, Forsberg K, et al. On the origins of a species: what might explain the rise of Candida auris? J Fungi. 2019;5(3):58. doi:10.3390/jof5030058
120. Casadevall A, Kontoyiannis DP, Robert V. On the emergence of Candida auris: climate change, azoles, swamps, and birds. mBio. 2019;10(4):e01397–19. doi:10.1128/mBio.01397-19
121. Kim SH, Iyer KR, Pardeshi L, et al. Genetic analysis of Candida auris implicates Hsp90 in morphogenesis and azole tolerance and Cdr1 in azole resistance. mBio. 2019;10(1):e02529–18. doi:10.1128/mBio.02529-18
122. Hernández-Chávez MJ, Pérez-García LA, Niño-Vega GA, Mora-Montes HM. Fungal strategies to evade the host immune recognition. J Fungi. 2017;3(4):51. doi:10.3390/jof3040051
123. Netea MG, Brown GD, Kullberg BJ, Gow NA. An integrated model of the recognition of Candida albicans by the innate immune system. Nat Rev Microbiol. 2008;6(1):67–78. doi:10.1038/nrmicro1815
124. Richardson JP, Moyes DL. Adaptive immune responses to Candida albicans infection. Virulence. 2015;6(4):327–337. doi:10.1080/21505594.2015.1004977
125. Pathirana RU, Friedman J, Norris HL, et al. Fluconazole-resistant Candida auris is susceptible to salivary histatin 5 killing and to intrinsic host defenses. Antimicrob Agents Chemother. 2018;62(2):e01872–17. doi:10.1128/aac.01872-17
126. Urban CF, Reichard U, Brinkmann V, Zychlinsky A. Neutrophil extracellular traps capture and kill Candida albicans yeast and hyphal forms. Cell Microbiol. 2006;8(4):668–676. doi:10.1111/j.1462-5822.2005.00659.x
127. Johnson CJ, Davis JM, Huttenlocher A, Kernien JF, Nett JE. Emerging fungal pathogen Candida auris evades neutrophil attack. mBio. 2018;9(4). doi:10.1128/mBio.01403-18
128. Navarro-Arias MJ, Hernández-Chávez MJ, García-Carnero LC, et al. Differential recognition of Candida tropicalis, Candida guilliermondii, Candida krusei, and Candida auris by human innate immune cells. Infect Drug Resist. 2019;12:783–794. doi:10.2147/idr.S197531
129. Wang Y, Zou Y, Chen X, et al. Innate immune responses against the fungal pathogen Candida auris. Nat Commun. 2022;13(1):3553. doi:10.1038/s41467-022-31201-x
130. Pérez-García LA, Csonka K, Flores-Carreón A, et al. Role of protein glycosylation in Candida parapsilosis cell wall integrity and host interaction. Original Research. Front Microbiol. 2016;7. doi:10.3389/fmicb.2016.00306
131. McKenzie CG, Koser U, Lewis LE, et al. Contribution of Candida albicans cell wall components to recognition by and escape from murine macrophages. Infect Immun. 2010;78(4):1650–1658. doi:10.1128/IAI.00001-10
132. West L, Lowman DW, Mora-Montes HM, et al. Differential virulence of Candida glabrata glycosylation mutants. J Biol Chem. 2013;288(30):22006–22018. doi:10.1074/jbc.M113.478743
133. Bruno M, Kersten S, Bain JM, et al. Transcriptional and functional insights into the host immune response against the emerging fungal pathogen Candida auris. Nat Microbiol. 2020;5(12):1516–1531. doi:10.1038/s41564-020-0780-3
134. Gandra RM, McCarron P, Viganor L, et al. In vivo activity of copper(II), manganese(II), and silver(I) 1,10-phenanthroline chelates against Candida haemulonii using the Galleria mellonella model. Front Microbiol. 2020;11:470. doi:10.3389/fmicb.2020.00470
135. Xu S, Webb SE, Lau TCK, Cheng SH. Matrix metalloproteinases (MMPs) mediate leukocyte recruitment during the inflammatory phase of zebrafish heart regeneration. Sci Rep. 2018;8(1):7199. doi:10.1038/s41598-018-25490-w
136. Fontenot JD, Gavin MA, Rudensky AY. Foxp3 programs the development and function of CD4+CD25+ regulatory T cells. Nat Immunol. 2003;4(4):330–336. doi:10.1038/ni904
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.