")
Back to Journals » International Journal of Nanomedicine » Volume 18
Can Organoid Model Reveal a Key Role of Extracellular Vesicles in Tumors? A Comprehensive Review of the Literature
Authors Zhang Y , Lu A, Zhuang Z, Zhang S, Liu S, Chen H, Yang X, Wang Z
Received 7 June 2023
Accepted for publication 19 September 2023
Published 27 September 2023 Volume 2023:18 Pages 5511—5527
DOI https://doi.org/10.2147/IJN.S424737
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Farooq A. Shiekh
Yang Zhang,1,2,* Anqing Lu,2– 4,* Zixuan Zhuang,1,2 Su Zhang,5 Sicheng Liu,5 Haining Chen,1,2 Xuyang Yang,1,2 Ziqiang Wang1,2
1Colorectal Cancer Center, West China Hospital, Sichuan University, Chengdu, 610041, People’s Republic of China; 2Department of General Surgery, West China Hospital, Sichuan University, Chengdu, 610041, People’s Republic of China; 3Department of Central Transportation, West China Hospital, Sichuan University, Chengdu, 610041, People’s Republic of China; 4West China School of Nursing, Sichuan University, Chengdu, 610041, People’s Republic of China; 5Research Laboratory of Tumor Epigenetics and Genomics, Department of General Surgery, Frontiers Science Center for Disease-Related Molecular Network and National Clinical Research Center for Geriatrics, State Key Laboratory of Biotherapy and Cancer Center, West China Hospital, Sichuan University, Chengdu, 610041, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Ziqiang Wang; Xuyang Yang, Colorectal Cancer Center, West China Hospital, Sichuan University, Chengdu, 610041, People’s Republic of China, Email [email protected]; [email protected]
Abstract: Extracellular vesicles (EVs) are small membrane-bound vesicles that are released by cells into the extracellular environment. The role of EVs in tumors has been extensively studied, and they have been shown to play a crucial role in tumor growth, progression, and metastasis. Past research has mainly used 2D-cultured cell line models to investigate the role of EVs in tumors, which poorly simulate the tumor microenvironment. Organoid technology has gradually matured in recent years. Organoids are similar in composition and behavior to physiological cells and have the potential to recapitulate the architecture and function of the original tissue. It has been widely used in organogenesis, drug screening, gene editing, precision medicine and other fields. The integration of EVs and organoids has the potential to revolutionize the field of cancer research and represents a promising avenue for advancing our understanding of cancer biology and the development of novel therapeutic strategies. Here, we aimed to present a comprehensive overview of studies using organoids to study EVs in tumors.
Keywords: organoids, extracellular vesicles, tumors, model, review
Introduction
Extracellular vesicles (EVs) are small membrane-bound vesicles that are released by cells into the extracellular environment.1 They contain a variety of bioactive molecules, including proteins, lipids, and nucleic acids, and are involved in intercellular communication and the regulation of various physiological and pathological processes.2 EVs are a group of nanovesicles that can be mainly divided into 3 categories based on their cellular origin: exosomes, microvesicles and apoptotic bodies.3 It is difficult to distinguish them using common isolation methods (differential ultracentrifugation, density gradient centrifugation, size exclusion chromatography, filtration, polymer-based precipitation, immunological separation, etc.) due to the lack of specific markers and the fact that the sizes or diameters of these EV subtypes partially overlap. The minimal information for studies of extracellular vesicles 2018 (MISEV2018) guideline noted that EVs were more appropriate than other terms, such as exosomes or microvesicles.4 Therefore, we use the term EVs to unify the nomenclature throughout this review.
The role of EVs in tumors has been extensively studied, and they have been shown to play a crucial role in tumor growth, progression, and metastasis.5,6 Tumor-derived EVs can transfer oncogenic cargo to neighboring or distant cells, leading to the alteration of the tumor microenvironment and the promotion of cancer cell survival and proliferation. In addition, tumor cells generally secrete more EVs than their counterparts.7 For instance, colorectal cancer (CRC)-derived EVs can promote distant metastasis. Mechanistically, miR-25-3p can be transferred from CRC cells to endothelial cells via EVs, which target KLF2 and KLF4, resulting in the induction of vascular permeability and angiogenesis.8 Melanoma-derived exosomes can promote tumor growth and metastases. Mechanistically, melanoma-derived exosomes can reprogram bone marrow progenitors towards pro-vasculogenic and pro-metastatic phenotypes by transferring the receptor tyrosine kinase MET.9
In the field of medical research, many drugs or inventions have produced promising results in laboratories but have failed in clinical trials. It was reported that the success rate of clinical trials was as low as 3%.10 This is largely due to inappropriate modeling of tumors, as these models cannot recapitulate pathogenic processes in patients.11 Organoids, defined as “a collection of organ-specific cell types that develops from stem cells or organ progenitors and self-organizes through cell sorting and spatially restricted lineage commitment in a manner similar to in vivo”, have the potential to recapitulate the architecture and function of the original tissue.12 Organoid technology has gradually matured in recent years. Some standard experimental protocols or guidelines have been established,13,14 and organoids have been revealed to be a powerful tool for studying the biology of tumors and for drug discovery.15 In a recent study, lung cancer organoids were established with a success rate of 80%, and they can accurately predict the clinical response to specific chemotherapeutic or targeted therapy.16
The integration of EVs and organoids has the potential to revolutionize the field of cancer research. EVs can be used to transfer specific cargo to organoids, mimicking the interactions that occur in vivo and allowing for the study of complex signaling pathways and the identification of novel therapeutic targets. Organoids, on the other hand, can be used to model the response of tumors to EV-based therapies and to test the efficacy of new drugs in a more physiologically relevant context.17 Moreover, some studies have tried to uncover EVs secreted from tumor organoids. In short, the combination of EVs, tumors, and organoids represents a promising avenue for advancing our understanding of cancer biology and for the development of novel therapeutic strategies.
In this review, we aimed to present a comprehensive overview of studies using organoids to study EVs in tumors. We first briefly introduced EVs in tumors and organoids in tumors; then, the crosstalk between organoids and EVs in tumors was discussed from two aspects: 1) EVs secreted from cells and 2) EVs secreted from tumor organoids.
What are Extracellular Vesicles in Tumors
The human body is composed of trillions of cells, and how to govern their communication and interactions is of great importance. Extracellular vesicles (EVs) are composed of biological nanovesicles, including proteins, lipids and nucleic acids. Released from both prokaryotes and eukaryotes, EVs are key players in intercellular communication and are increasingly being studied in many fields, including tumors.1 In this section, we will briefly discuss EV biogenesis, subtypes, contents, functions, and roles in cancer development and progression. For more about the biogenesis, secretion, and intercellular interactions of EVs, please refer to the review written by Théry et al.2 For more specific mechanisms of EVs in tumors, please refer to the review written by Han et al.18
The Extracellular Vesicles and Their Biogenesis
Two major EV subtypes have been characterized, namely, exosomes and microvesicles (MVs). Exosomes, originating from multivesicular endosomes (MVEs), are 30–150 nm in size and have an average density of 1.13 g/mL.4,19 The complete process of exosome biogenesis is complex. Briefly, exosomes are first formed as intraluminal vesicles (ILVs) by budding into early endosomes and MVEs. Then, late endosomes are formed and these ILVs are released into the extracellular environment as exosomes when late endosomes fuse with the plasma membrane (PM). Late endosomes can also fuse with lysosomes to degrade their contents.20,21 MVs are 150–1000 nm vesicles released by direct budding from the PM and have a higher density of 1.18 g/mL2,22,23 (Figure 1).
 |
Figure 1 An illustration showed the subtypes and biogenesis of extracellular vesicles20,22 [created with BioRender.com (https://biorender.com/)]. Abbreviation: MVEs, multivesicular endosomes. |
The Extracellular Vesicles and Their Contents
EVs contain lipids, proteins, and nucleic acids (including DNAs, mRNAs and noncoding RNAs), which makes them an attractive platform for biomarker discovery and drug delivery.24 The nature and number of molecules present in an EV depend on the cellular context and cell type of origin.25 For example, exosomes isolated from cancer cells typically show higher levels of pro-oncogenic microRNAs and hence decreased expression of target tumor suppressive genes compared to their healthy counterparts.26 In addition, isolation methods or enrichment techniques can also affect the content of EVs in the actual experimental process.27
The Extracellular Vesicles and Their Functions
The biological function of EVs is multifaceted. EVs are widely involved in physiological and pathological processes, such as antigen presentation and the immune response, cell apoptosis and senescence, intercellular material transportation and signal transduction.1,28–30 For instance, they can act as paracrine messengers and transfer biomolecules from donor cells to target cells. This includes not only metabolic molecules but also proteins, lipids, mRNAs, lncRNAs, and microRNAs capable of altering the phenotype of target cells.24 Additionally, EVs can induce autocrine signaling in donor cells, as seen with platelet-derived EVs, which release thrombin to directly activate platelets.31
The Roles of Extracellular Vesicles in Tumors
EVs have long been studied for their potential implications in many diseases, including tumors. This is mainly because they carry information-rich biomolecules, which can influence gene expression in target cells.6 Numerous studies have shown that tumor-derived EVs facilitate tumor growth and metastasis by inducing cell proliferation, invasion, and resistance to chemotherapy and radiotherapy, as well as inducing angiogenesis.32–35 Research has also demonstrated that EVs secreted by tumor cells can recruit T cells and macrophages, contributing to tumor-associated inflammation and immunomodulatory phenotypes and furthering tumor progression and metastasis.36,37 Moreover, tumor-derived EVs can be useful for the early detection and diagnosis of cancer since they are much more abundant than their cellular counterparts and thus can be easily detected and quantified.38–42
In conclusion, these findings indicate that EVs can be instrumental in both tumorigenesis and tumor progression, thus necessitating further research into their clinical implications. Their potential as therapeutic agents and biomarkers might provide an unprecedented opportunity for personalized medicine for cancer patients, thus revolutionizing the field of oncology.
The Role of Organoids in Tumors
The Advantages and Disadvantages of Organoids and Other Tumor Models
Tumor research has made significant progress due to the development of various tumor models. Since the establishment of the first patient-derived tumor cell line HeLa in 1951,43 tumor models have diversified into four common models: cell lines, animal models, patient-derived xenografts (PDXs), and 3D culture cell models, including organoids (Figure 2). We will discuss the advantages and limitations of these common models.
 |
Figure 2 Four common models used to study tumors.44–47 [created with BioRender.com (https://biorender.com/)]. |
Cell lines have been the mainstream models in biomedical laboratories. Typically, 2D-cultured immortalized cancer cell lines are used because they are easily accessible, easy to operate, and can be stored for a long time. Additionally, it is straightforward to study the downstream mechanism of a specific gene using cell lines. However, cell lines are limited in their ability to form multicellular structures and lose cell-to-cell connections, which poorly mimics the tumor microenvironment. Long-term cell culture can also cause cell lines to lose their unique features.44,48,49
Animal models are frequently used in biomedical research, especially in preclinical drug research. Basic knowledge of human anatomy and disease pathology can also be attributed to preclinical studies of animal models.45 As in vivo experimental models, animal models better mimic the clinical situation of specific diseases, such as cancer and predict the effectiveness of treatment strategies.50 Mouse cancer models are the most commonly used animal models in cancer research. Genetically engineered mice play a vital role in identifying novel oncogenes or tumor suppressor genes and tumor biomarkers and provide strong evidence to verify the function of genes in vivo.51 However, animal models have several limitations, including high cost, low throughput, long experimental period, interspecific differences, individual differences, and ethical issues.17,52
Patient-derived xenografts (PDXs) are generated by implanting patient-derived tissue into animal models, typically immunodeficient mice.46 PDXs provide a good model for studying the microenvironment of cancer cells, such as evaluating the response of fibroblasts or endothelial cells to treatment. They can maintain close similarities with the tumor of origin and the integrity of the primary tumor structure. However, PDXs have limitations, including the expense and time required to construct models, limited genetic and environmental manipulation, and reliance on immunodeficient mice, which makes it impossible to study the role of the adaptive immune system.53 Moreover, high-throughput screens are difficult to achieve, and they still involve animal ethics.54
3D cell culture models (including organoids) can provide a microenvironment closer to the living conditions in vivo for cells during cell culture.47 Compared with cell lines, organoids are similar in composition and behavior to physiological cells and are more suitable for high-throughput screening. Compared with animal models and PDXs, organoids have a simpler operation and lower cost and can be used to study the mechanism of tumorigenesis and tumor development. Therefore, it has been widely used in organogenesis, drug screening, gene editing, precision medicine and other fields.15 However, organoids have some drawbacks, including containing only the epithelial layer without a tumor microenvironment.55 The culture medium must be of high quality, and cytokines and other additive agents in the culture medium can affect gene expression, leading to different experimental results.47 Table 1 lists the major advantages and disadvantages of the different tumor models.
 |
Table 1 Major Advantages and Disadvantages of the Different Models in Tumors |
The Development of Organoids
Organoids have become a popular tool for modeling diseases in recent years, which are defined as “a collection of organ-specific cell types that develops from stem cells or organ progenitors and self-organizes through cell sorting and spatially restricted lineage commitment in a manner similar to in vivo”.12 Colloquially, organoids refer to a cluster of cells growing in a three-dimensional (3D) environment that self-organize and differentiate into various types of cells with specific functions, mimicking the structure and function of an organ in vivo.56 Organoids can be divided into four types based on the origin of stem cells, including pluripotent embryonic stem cells (ESCs), induced pluripotent stem cells (iPSCs), organ-restricted adult stem cells (aSCs), and cancer cells.47,57 Briefly, there are two major types of organoids: organoids from tissues and organoids from somatic cells or stem cells.
The concept of organoids can be traced back to 1907 when Wilson discovered that dissociated sponge cells could self-organize to regenerate a whole organism with normal functions.58 In 1960, Paul Weiss and A.C. Taylor performed dissociation-reaggregation experiments to generate several organs from chick embryos.59 Ten years later, Friedenstein et al discovered mesenchymal stem cell bone marrow in 1970,60 marking the rise of stem cell technologies. In 1998, James Thomson isolated human embryonic stem cells (ESCs) for the first time.61 Subsequently, induced pluripotent stem cells (iPSCs) were successfully generated from mouse embryonic and adult human fibroblasts.62,63 These events symbolize the thriving of stem cell technology, which has led to the further development of organoids. In 2009, Hans Clevers et al first established intestinal organoids using a single mouse intestinal stem cell.64 Organoid technology was rated as one of the top ten technologies of the year by Science magazine in 2013. In 2020, Lee et al successfully constructed heart organoids that could beat autonomously using mouse embryonic stem cell-derived embryoid bodies.65 Over the past few decades, organoid technology has matured rapidly, and it is now widely employed in biomedical research.
The Application of Organoids in Different Tumors
Recently, organoid research has mainly focused on patient-derived organoids (PDOs), particularly organoids derived from patients’ tumor tissues, also known as tumor organoids. These tumor organoids have been successfully established in various types of tumors, such as lung cancer,66,67 breast cancer,68,69 colorectal cancer (CRC),70–72 pancreatic ductal adenocarcinoma (PDAC),72–74 hepatocellular carcinoma (HCC)75,76 and cholangiocarcinoma (CC),76,77 gastric cancer,78 gliomas,79 glioblastoma,80,81 neuroendocrine neoplasms,82 bladder cancer,83 etc. (Figure 3). Several common tumors are discussed below, and some brief information is listed in Table 2. They are usually used for constructing living organoid biobanks,14,84 cancer modeling,47 evaluating the toxicity and efficacy of anticancer drugs,85 guiding precision medicine,86 and regeneration medicine87 (Figure 3).
 |
Figure 3 Application of Organoids in different aspects (left) and tumors (right).66–83,88 [created with BioRender.com (https://biorender.com/)]. |
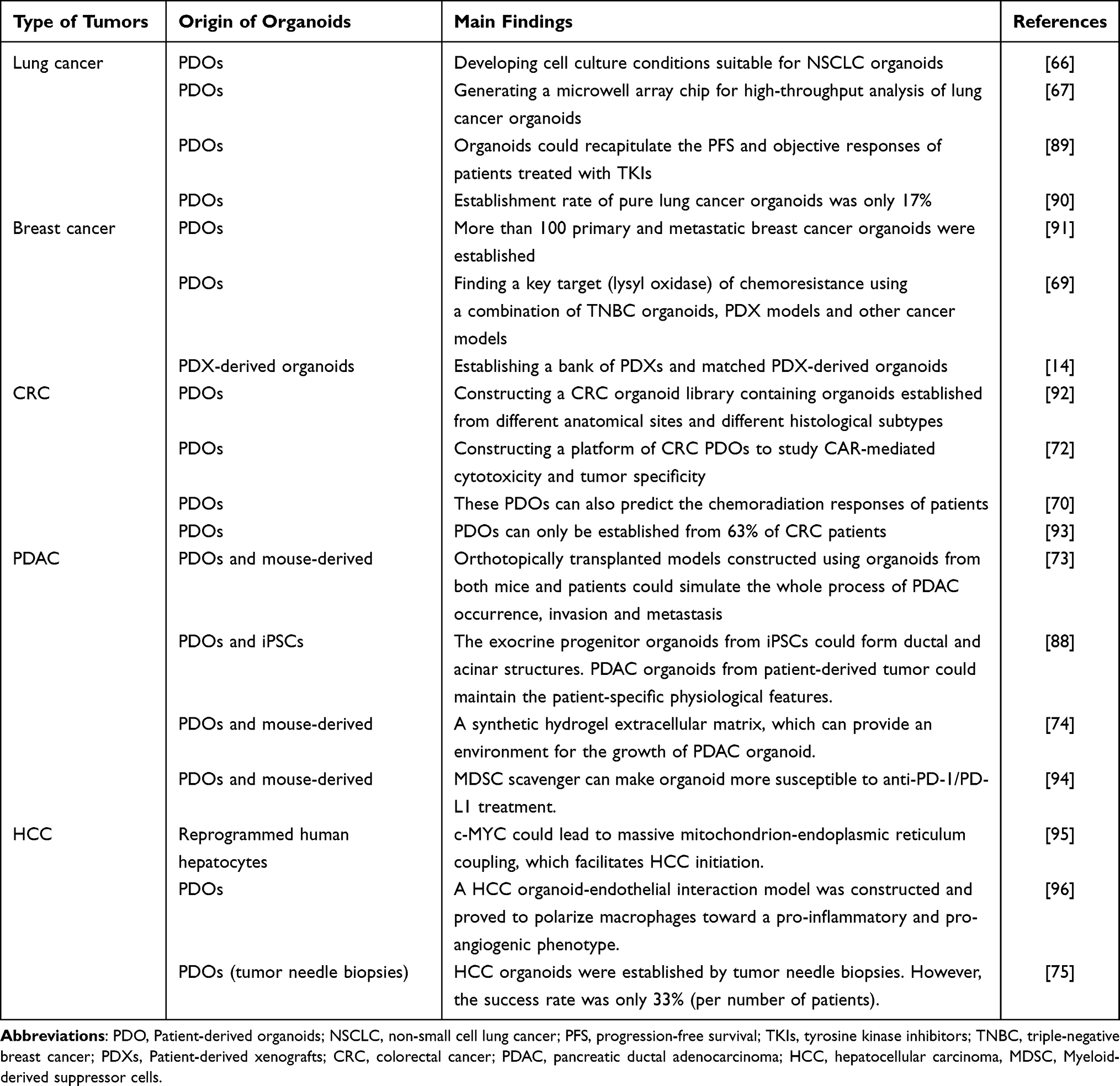 |
Table 2 The Application of Organoids in Different Tumors |
Lung Cancer
Lung cancer organoid models have been successfully applied to study the efficacy of chemotherapy drugs. It was reported that 87% of lung tumor samples could successfully generate tumor organoids.97 Researchers have developed cell culture conditions suitable for the establishment of NSCLC organoids. They also reported that these organoid models could preserve the sensitivity of the matched parental tumor to specific drugs.66 Another study further constructed advanced lung cancer organoids and found that these models could recapitulate the progression-free survival (PFS) and objective responses of NSCLC patients treated with tyrosine kinase inhibitors (TKIs).89 Hu et al developed an integrated superhydrophobic microwell array chip for high-throughput analysis of lung cancer organoids that can test the response to anticancer drugs within a week, and the results corresponded with clinical outcomes.67 However, one study used copy number profiling and immunohistochemistry to differentiate tumor and normal organoids and revealed that only 17% of pure lung cancer organoids were established.90 Although many studies on lung cancer organoids have been conducted, the accuracy of these models in reflecting the clinical situation is still debated due to purity issues.
Breast Cancer
Breast cancer is being modeled using organoid technology. A group of researchers led by Sachs N has developed over 100 primary and metastatic breast cancer organoids, which have been shown to recapitulate the genetic and histological features of breast cancer. These organoids can be used to perform high-throughput drug screening, which can aid in drug discovery for breast cancer treatment.14 In a study focused on triple-negative breast cancer (TNBC), a key target for chemoresistance (lysyl oxidase) was identified using TNBC organoids, PDX models, and other cancer models. The study also demonstrated that the expression of lysyl oxidase was associated with overall survival in chemotherapy-treated TNBC patients, indicating the potential clinical relevance of TNBC organoids.69 Additionally, another study involving TNBC reported the establishment of a bank of PDXs and matched PDX-derived organoids that accurately represented the clinical situation of TNBC patients treated with anticancer drugs.91
Colorectal Cancer
CRC PDOs have been established for a period of time. In 2009, Hans Clevers and his team first reported a groundbreaking achievement that successfully cultured colon epithelial-like organs in vitro.64 In this study, researchers digested colon epithelial tissue into colon crypt structures or individual Lgr5+ stem cells, and embedded them in a matrix gel (Matrigel), then cultured them in a medium containing epidermal growth factor (EGF), R-spondin 1 protein related to the Wnt signaling pathway, and Noggin protein. Due to the supporting properties of Matrigel and the proliferation and differentiation ability of Lgr5+ stem cells at the bottom of colon crypts, the obtained colon crypt structures or individual Lgr5+ stem cells were able to grow in all directions, forming 3D tissue spheres containing colon epithelial crypts and villus-like structures. Through immunofluorescence staining, immunohistochemistry, and electron microscopy observation, it was found that the structures contained various types of cells, including colon epithelial cells, goblet cells, Paneth cells, enteroendocrine cells, and Lgr5+ stem cells, among others, and had structures similar to colon epithelium. In 2015, Hans Clevers’ team established for the first time a biobank of colon cancer organoids.84 They successfully cultured 22 colon cancer organoids from 27 surgically resected colon cancer samples, with an overall success rate of 90%, and all organoids could be cryopreserved and revived with a survival rate of over 80% after thawing. By comparing fixed, sliced, and H&E-stained samples of donor tissue and cultured organoids, it was found that the organoids derived from normal intestinal epithelial tissue had a more regular shape with a hollow cavity and villus-like structures, while those derived from colon cancer tissue largely retained the morphological features of the donor tissue, such as thin-walled cystic structures or compact structures without cavities. Subsequently, genomic DNA was extracted from both organoids and corresponding donor biopsy tissues, and whole exome sequencing was performed. It was found that the average mutation rate per megabase of the organoids was similar to that of the donor biopsy tissues, with mutations mainly occurring as CpG to T transitions, consistent with the results of large-scale sequencing of colon cancer. Transcriptome analysis showed slight differences in gene expression between different organoids, indicating heterogeneity between organoids. Based on these findings, Hans Clevers’ team established the nonprofit organization Hubrecht Organoid Technology (HUB) to further expand the scale of biobanks for colon cancer and other organoid types. Masayuki Fujii successfully created a CRC organoid library with organoids from different anatomical sites (primary lesions and liver metastatic lesions) and different histological subtypes (adenocarcinoma and neuroendocrine carcinoma). These organoids displayed a high degree of similarity to primary tumors in terms of histopathological structures and gene expression profiling.92 Of all CRC sites, rectal cancer has a worse prognosis, especially locally advanced rectal cancer (LARC) and those with distant metastasis, where conventional chemoradiation is largely ineffective.98 Developing methods to predict chemoradiation response and achieve precision therapy is an area for future development. Another study focused specifically on LARC and developed an organoid biobank from LARC patients. The authors demonstrated that these organoids had a similar molecular spectrum to that of the primary tumors and can predict patients’ chemoradiation responses, implying their potential value for guiding LARC treatment.70 Tumor immunotherapy research has shown that chimeric antigen receptor-T (CAR-T) cell therapy is highly effective. However, the use of CAR-T-cell therapy in solid cancers such as CRC has been less studied. Schnalzger et al constructed a platform of CRC PDOs to investigate CAR-mediated cytotoxicity and tumor specificity, discovering a novel CAR target (FRIZZLED receptors) highly expressed in a subgroup of CRC tumors.72 Nevertheless, it is important to note that PDOs can only be established from 63% of CRC patients,93 indicating that much work is still needed before the clinical use of CRC organoids can be realized.
Pancreatic Ductal Adenocarcinoma
Pancreatic ductal adenocarcinoma (PDAC) is a highly lethal cancer with a poor prognosis. Therefore, it is crucial to establish appropriate models to study the process and mechanism of tumor occurrence and development.99 Boj et al successfully established PDAC organoids from both mice and patients, which can simulate the entire process of PDAC occurrence, invasion, and metastasis.73 Similarly, Huang et al built pancreatic exocrine progenitor organoids from iPSCs that can form ductal and acinar structures. These organoids maintain the differentiation status and tissue architecture of the primary tumor and preserve patient-specific physiological features.88 Recently, Below et al designed a synthetic hydrogel extracellular matrix that can provide an environment for the growth of PDAC organoids from both human patients and murine models.74 Another study focused on the tumor immune microenvironment of PDAC and investigated potential immunotherapy. Although anti-programmed death 1 receptor (PD-1)/programmed death ligand 1 (PD-L1) therapy is a hot spot in the field of immune checkpoint inhibitor-mediated immunotherapy, there is still a great possibility of treatment failure.100 Myeloid-derived suppressor cells (MDSCs) can inhibit cellular immunity in cancers, including PDAC, which can lead to the failure of anti-PD1/PD-L1 therapy.101 Holokai et al found that when murine- or patient-derived organoids were cocultured with immune cells, MDSC scavengers could make organoids more susceptible to anti-PD-1/PD-L1 treatment. In other words, MDSC depletion combined with anti-PD-1/PD-L1 therapy enhanced efficacy.94
Hepatocellular Carcinoma
Hepatocellular carcinoma (HCC) is the most common primary liver cancer worldwide and has a poor prognosis. Despite this, the mechanisms underlying HCC development and progression remain unclear.102 The emergence of organoid technology has provided a powerful tool for studying the molecular mechanisms of HCC tumorigenesis and development. Using this technology, researchers have been able to build HCC organoid models by reprogramming human hepatocytes and inactivating p53 and RB. These models have revealed that c-MYC could lead to massive mitochondrion-endoplasmic reticulum coupling, which facilitates HCC initiation, and could potentially be targeted by pretreatment therapy. Therefore, HCC organoid models are useful in simulating the initiation of HCC and identifying potential preventive therapies.95 The tumor microenvironment plays a significant role in tumor occurrence and progression. Researchers have constructed HCC organoid-endothelial interaction models and found that these models can polarize macrophages towards a proinflammatory and proangiogenic phenotype. The results demonstrated that these models can serve as reliable tools to study the crosstalk between tumor cells and the tumor microenvironment.96 Nuciforo et al successfully used HCC organoids to test the sensitivity of tumors to sorafenib, a targeted therapeutic drug commonly used for advanced HCC. The HCC organoids in this study were established by tumor needle biopsies, providing a chance to guide personalized medication for HCC patients at their initial visit. However, the success rate was only 33% (per number of patients),75 and there is still a long way to go in improving the success rate of organoid construction.
Organoids as a Promising Model to Study Extracellular Vesicles in Tumors
Compared with 2D-cultured tumor cells and other cellular models, organoids have many unique advantages for studying the roles of EVs in tumors. By modeling specific tissues in terms of their composition and architecture, organoids can replicate the pathogenic processes that occur in human patients. This makes them an ideal tool for studying the tumor microenvironment and the altered molecular signaling pathways that occur in cancer.57 As an important component of the tumor microenvironment, the precise role of tumor-derived EVs has yet to be fully elucidated due to limitations in existing models. However, organoid technology has emerged as a promising solution to this dilemma. Organoids provide a valuable model for investigating the functions of EVs in tumors through two major approaches: 1) characterizing the changes in organoid morphology and gene expression profiling after EVs are secreted from cultured cells and absorbed by organoids, and 2) studying the factors that affect EV biogenesis, secretion, and contents within tumor organoids (Figure 4).
 |
Figure 4 Organoids as a promising model to study EVs in tumors.103–118 [created with BioRender.com (https://biorender.com/)]. Abbreviations: EVs, extracellular vesicles; NTA, nanoparticle tracking analysis. |
The Role of Extracellular Vesicles Secreted from Cells
Tumor cells can secrete EVs to regulate the function of other cells. Ke et al investigated the effects of EVs by studying their interaction with organoids in a coculture system. Specifically, they found that esophageal adenocarcinoma-derived EVs could enhance the neoplastic phenotype in gastric organoids.103 Despite recent advances, the complexity of tumor immunity still requires further research. Zhao et al used PDOs to investigate the influence of CRC cell-derived EVs on immune cells and found that CRC-derived exosomal miR-424 suppressed the T-cell-mediated antitumor immune response.104 The use of cell lines, organoids, and mouse models in this study enhanced the reliability of the results. Another investigation into tumor immunity, focusing on triple-negative breast cancer (TNBC), revealed that TNBC-derived exosomes can induce immunogenic cell death and promote tumor inhibition in PDOs.105
Organoids have shown promise as a tool not only for studying the function of tumor cell-derived EVs but also for investigating the roles of nontumor cell-derived EVs, such as fibroblasts and immune cells. One study used PDOs to examine the impact of fibroblast-derived EVs and found that they could enhance cell proliferation in epidermal growth factor (EGF)-dependent PDOs.106 Intestinal metaplasia is a known risk factor for gastric cancer, and it has been suggested that polypeptide-expressing metaplasia (SPEM) could be a contributing factor. Xu et al reported that deoxycholic acid-induced macrophage-derived EVs could be absorbed by gastric organoids in mice, promoting the expression of TFF2 and GSII lectin, both SPEM markers.107
Organoids can also be used to study the factors that affect EV absorption. Kelemen et al investigated EV uptake by PDOs and mouse tissue-derived organoids. They found that organoids derived from CRC cells with high expression of IFITM1 had a significantly reduced uptake of fibroblast EVs.108 EVs serve as carriers for cell-to-cell communication, which has unique advantages in drug delivery. Zhuang et al developed an EV-based Cas9 delivery platform. The HCC PDO model was used to evaluate the targeting efficacy of engineered EVs. These engineered EVs were found to efficiently accumulate in HCC-derived organoids in a tumor-specific manner.109
Organoids have special advantages in evaluating drug response and chemotherapy resistance. A recent study established CRC PDOs and found that adipocyte-derived exosomes could reduce the sensitivity of CRC organoids to oxaliplatin. Mechanistically, adipocyte-derived exosomes reduce ferroptosis susceptibility in obese CRC patients through the microsomal triglyceride transfer protein (MTTP)/proline-rich acidic protein 1 (PRAP1) complex.110
In general, organoids can be used to study the function of EVs derived from various cells, including tumor cells, fibroblasts, immune cells and adipocytes. In addition, organoids can also be used to study the factors that affect EV absorption, and organoids have special advantages in evaluating drug response and chemotherapy resistance.
The Role of Extracellular Vesicles Secreted from Tumor Organoids
Tumor-derived organoids can also secrete EVs to affect intercellular communication and tumor progression and metastasis. Various factors affect the secretion and contents of organoid-derived EVs. Pancreatic ductal adenocarcinoma (PDAC) and lung adenocarcinoma (LUAD) PDOs have shown that Wnt pathway activation promotes both cell proliferation and EV release in these organoids.111 The APC gene is a common tumor suppressor gene, and its mutation is associated with familial adenomatous polyposis (FAP) and sporadic colorectal tumors.112 Szvicsek et al investigated the effect of APC mutation on EV release in CRC organoids and found that it could activate the Wnt pathway and induce EV release. They also observed that hypoxic fibroblast-derived EVs could promote the colony formation of CRC organoids.113 Additionally, the distribution of EVs was studied using organoids. Namba et al discovered that ABCG1, a cholesterol lipid efflux pump, was upregulated in colon cancer organoids, and that its knockdown could lead to intracellular accumulation of EVs and regression of organoids.114
Several studies have investigated the content alteration of organoid-derived extracellular vesicles (EVs). One study demonstrated that organoids can regulate their proliferation and tumorigenesis by controlling EV secretion.115 The researchers knocked out MMP3 in LuM1 tumor cells and identified a protein profile that was significantly downregulated in organoid-derived EVs, resulting in a notable reduction in organoid size. In another study, Nagai et al generated colorectal adenoma (CRA) and colorectal cancer (CRC)-derived organoids and compared the microRNA profiles of their EVs. They found that the expression of microRNA-1246 was upregulated in EVs derived from CRC-derived organoids, and it was shown to promote the proliferation of HT29 cells (a CRC cell line).116 Moreover, researchers analyzed the protein profiles of EVs from pancreatic ductal adenocarcinoma (PDAC) organoids using mass spectrometry. They found that the proteins present in PDAC organoid-derived EVs were primarily involved in vesicular transport and tumorigenesis, in contrast to healthy control organoids.117 These studies suggest that EVs have the potential to serve as diagnostic biomarkers for tumors.
Numerous studies have investigated the molecular composition of extracellular vesicles (EVs) since they are believed to reflect the condition of the cells that release them. However, Kelemen et al found that CD44high CRC organoids release more EVs and induce the proliferation and activation of colon fibroblasts compared to CD44low CRC organoids. The study demonstrated that the proliferative effect was dependent on the dose of EVs rather than their miRNA cargo.118 This suggests that tumor cells can regulate the phenotype of other cells by altering the number of EVs they secrete, rather than changing the content of EVs in certain situations.
Various factors that affect EV secretion and contents have been studied, including the type of cancer cell used to generate organoids. Different cancer cells can produce different types of EVs with unique biophysical properties and gene expression profiles, which have been investigated in studies focusing on tumor intrinsic factors such as protein expression and noncoding RNAs.15 Culture conditions, such as oxygen levels, nutrient availability, and pH, can also affect EV production and cargo selection. However, more studies are needed to confirm these findings.
Overall, the current research on organoids as a model for studying EVs in tumors focuses on two aspects: EVs secreted from cells and EVs secreted from tumor organoids (Table 3). Nevertheless, there is a shortage of studies using organoids to examine body fluid-derived EVs (such as ascites, urine, and plasma).
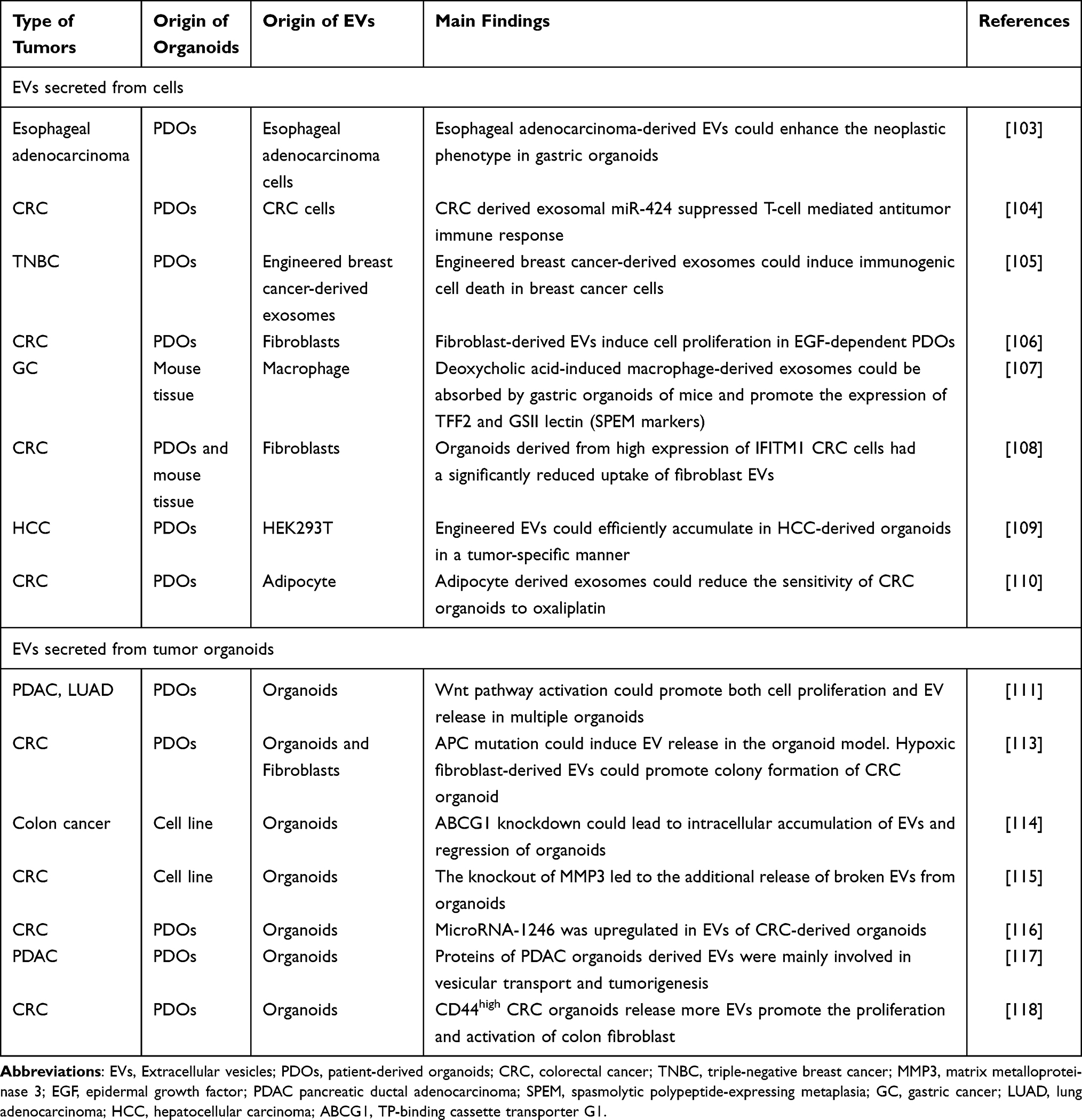 |
Table 3 Organoids as a Promising Model to Study Extracellular Vesicles in Tumors |
The Future Prospective
Extracellular vesicles (EVs) are key players in intercellular communication and a hot research topic in the field of tumors, as they play a crucial role in tumor growth, progression, and metastasis. However, past research has mainly used 2D-cultured cell line models, which poorly simulate the tumor microenvironment. Although many basic studies have been conducted in the field of cancer, the transition from preliminary basic research to clinical application remains a challenge. The emergence and development of organoid technology provide a promising model to study EVs in tumors, as organoids can mimic specific tissues in both composition and architecture, and recapitulate the real pathogenic process in the human body or patients.
It has been reported that equivalent amounts of cells in 3D cultures can produce 1.5–4.5 times more EVs than in 2D culture conditions, with overall downregulation of proteins and upregulation of microRNAs observed in 3D cultures.119 However, different tumor models might lead to different research results. As an in vitro model, organoids provide a vital opportunity for reducing the use of laboratory animals, with a study revealing that only approximately one-third of highly cited animal research could be proven by clinical randomized trials.120 Thus, organoid technology has built a bridge between 2D-cultured cell models and animal models, and is a good supplement to tumor models.
This review describes EVs and organoids distinctly and proposes the possible usefulness or advantages of organoids in the study of EVs in tumors. First, organoids have special advantages in evaluating the responses of anticancer drugs and studying the complex tumor immune microenvironment. The organoid/immune cell coculture model is the foundation when studying tumor immunity, which requires further research. In addition, organoid models can preliminarily verify the diagnostic efficacy of EVs, promoting the translation from basic cancer research to clinical practice. For instance, one study identified a group of eight ovarian cancer-related miRNAs based on overlapping dysfunctional miRNAs in ascites and plasma, which presented high diagnostic accuracy. They further constructed ascites-derived organoids and proved that malignant ascites-derived EVs could significantly promote the growth of these organoids.121 More importantly, organoids can be efficiently established from tumor tissues derived from individual patients, making it possible to develop personalized treatment or precision treatment of cancer. Meanwhile, engineered EVs have demonstrated the application potential in cancer-targeted therapy. The integration of organoids and EVs might revolutionize the field of cancer research and advance the development of novel therapeutic strategies.
The current research in this field still has many disadvantages, despite organoids offering excellent opportunities to study EVs in tumors. First, isolating and characterizing EVs secreted in 3D cultures is a problem to be solved. Second, growth factors or various small molecular inhibitors are needed for the culture of organoids, which might affect the secretion or contents of EVs. Third, some organoids still cannot be cultured as long as the immortalized cancer cell lines, due to the limitations of the culture medium. Fourth, organoids are composed of only an epithelial layer without a native microenvironment, and such studies rely on the establishment of coculture systems with other cells. Finally, when studying the effect of cell-derived EVs on organoids, the interference of organoid-secreted EVs needs to be eliminated.
Each tumor model has intrinsic limitations for recapitulating patient-specific tumors. With the development of organoid technology, it has become particularly important to standardize these models. Further exploration is needed to address the limitations and enhance the potential of organoids in the study of EVs in tumors.
Acknowledgments
This work was supported by Sichuan Science and Technology Program (2023YFS0277, 2021YFS0025), 1.3.5 project for disciplines of excellence, West China Hospital, Sichuan University (20HXJS003), 1.3.5 project for disciplines of excellence-Clinical Research Incubation Project, and West China Hospital, Sichuan University (22HXFH001, 2019HXFH031), China Postdoctoral Science Foundation (2022M712264), Post-Doctor Research Project, West China Hospital, Sichuan University (2021HXBH033).
Disclosure
The authors have no conflicts of interest in this work.
References
1. Yanez-Mo M, Siljander PR, Andreu Z, et al. Biological properties of extracellular vesicles and their physiological functions. J Extracell Vesicles. 2015;4(1):27066. doi:10.3402/jev.v4.27066
2. Colombo M, Raposo G, Thery C. Biogenesis, secretion, and intercellular interactions of exosomes and other extracellular vesicles. Annu Rev Cell Dev Biol. 2014;30(1):255–289. doi:10.1146/annurev-cellbio-101512-122326
3. György B, Szabó TG, Pásztói M, et al. Membrane vesicles, current state-of-The-art: emerging role of extracellular vesicles. Cell Mol Life Sci. 2011;68(16):2667–2688. doi:10.1007/s00018-011-0689-3
4. Théry C, Witwer KW, Aikawa E, et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018). J Extracell Vesicles. 2018;7(1). doi:10.1080/20013078.2018.1535750
5. Tkach M, Thery C. Communication by extracellular vesicles: where we are and where we need to go. Cell. 2016;164(6):1226–1232. doi:10.1016/j.cell.2016.01.043
6. Becker A, Thakur BK, Weiss JM, Kim HS, Peinado H, Lyden D. Extracellular vesicles in cancer: cell-to-cell mediators of metastasis. Cancer Cell. 2016;30(6):836–848. doi:10.1016/j.ccell.2016.10.009
7. Xu R, Rai A, Chen M, Suwakulsiri W, Greening DW, Simpson RJ. Extracellular vesicles in cancer - implications for future improvements in cancer care. Nat Rev Clin Oncol. 2018;15(10):617–638. doi:10.1038/s41571-018-0036-9
8. Zeng Z, Li Y, Pan Y, et al. Cancer-derived exosomal miR-25-3p promotes pre-metastatic niche formation by inducing vascular permeability and angiogenesis. Nat Commun. 2018;9(1):5395. doi:10.1038/s41467-018-07810-w
9. Peinado H, Alečković M, Lavotshkin S, et al. Melanoma exosomes educate bone marrow progenitor cells toward a pro-metastatic phenotype through MET. Nat Med. 2012;18(6):883–891. doi:10.1038/nm.2753
10. Wong CH, Siah KW, Lo AW. Estimation of clinical trial success rates and related parameters. Biostatistics. 2019;20(2):273–286. doi:10.1093/biostatistics/kxx069
11. Caponigro G, Sellers WR. Advances in the preclinical testing of cancer therapeutic hypotheses. Nat Rev Drug Discov. 2011;10(3):179–187. doi:10.1038/nrd3385
12. Lancaster MA, Knoblich JA. Organogenesis in a dish: modeling development and disease using organoid technologies. Science. 2014;345(6194):1247125. doi:10.1126/science.1247125
13. Driehuis E, Kretzschmar K, Clevers H. Establishment of patient-derived cancer organoids for drug-screening applications. Nat Protoc. 2020;15(10):3380–3409. doi:10.1038/s41596-020-0379-4
14. Sachs N, de Ligt J, Kopper O, et al. A Living biobank of breast cancer organoids captures disease heterogeneity. Cell. 2018;172(1–2):373–86.e10. doi:10.1016/j.cell.2017.11.010
15. Drost J, Clevers H. Organoids in cancer research. Nat Rev Cancer. 2018;18(7):407–418. doi:10.1038/s41568-018-0007-6
16. Kim J, Minna JD. Moving toward precision medicine with lung cancer organoids. Cell Rep Med. 2023;4(2):100952. doi:10.1016/j.xcrm.2023.100952
17. Bordanaba-Florit G, Madarieta I, Olalde B, Falcón-Pérez JM, Royo F. 3D cell cultures as prospective models to study extracellular vesicles in cancer. Cancers. 2021;13(2):307. doi:10.3390/cancers13020307
18. Han L, Lam EWF, Sun Y. Extracellular vesicles in the tumor microenvironment: old stories, but new tales. Mol Cancer. 2019;18(1). doi:10.1186/s12943-019-0980-8
19. Raposo G, Stoorvogel W. Extracellular vesicles: exosomes, microvesicles, and friends. J Cell Biol. 2013;200(4):373–383. doi:10.1083/jcb.201211138
20. Thery C, Tkach M, Théry C. Biogenesis and secretion of exosomes. Curr Opin Cell Biol. 2014;29:116–125. doi:10.1016/j.ceb.2014.05.004
21. Thery C. Exosomes: secreted vesicles and intercellular communications. F1000 Biol Rep. 2011;3:15. doi:10.3410/B3-15
22. Ratajczak MZ, Ratajczak J. Extracellular microvesicles/exosomes: discovery, disbelief, acceptance, and the future? Leukemia. 2020;34(12):3126–3135. doi:10.1038/s41375-020-01041-z
23. van Niel G, D’Angelo G, Raposo G. Shedding light on the cell biology of extracellular vesicles. Nat Rev Mol Cell Biol. 2018;19(4):213–228. doi:10.1038/nrm.2017.125
24. Kourembanas S. Exosomes: vehicles of intercellular signaling, biomarkers, and vectors of cell therapy. Annu Rev Physiol. 2015;77(1):13–27. doi:10.1146/annurev-physiol-021014-071641
25. Thery C, Zitvogel L, Amigorena S. Exosomes: composition, biogenesis and function. Nat Rev Immunol. 2002;2(8):569–579. doi:10.1038/nri855
26. Zhao S, Mi Y, Guan B, et al. Tumor-derived exosomal miR-934 induces macrophage M2 polarization to promote liver metastasis of colorectal cancer. J Hematol Oncol. 2020;13(1):156. doi:10.1186/s13045-020-00991-2
27. Takov K, Yellon DM, Davidson SM. Comparison of small extracellular vesicles isolated from plasma by ultracentrifugation or size-exclusion chromatography: yield, purity and functional potential. J Extracell Vesicles. 2019;8(1):1560809. doi:10.1080/20013078.2018.1560809
28. Lindenbergh MFS, Stoorvogel W. Antigen presentation by extracellular vesicles from professional antigen-presenting cells. Annu Rev Immunol. 2018;36(1):435–459. doi:10.1146/annurev-immunol-041015-055700
29. Estévez-Souto V, Da Silva-álvarez S, Collado M. The role of extracellular vesicles in cellular senescence. Febs j. 2022;290(5):1203–1211. doi:10.1111/febs.16585
30. Kawahara H, Hanayama R. The role of exosomes/extracellular vesicles in neural signal transduction. Biol Pharm Bull. 2018;41(8):1119–1125. doi:10.1248/bpb.b18-00167
31. Vajen T, Mause SF, Koenen RR. Microvesicles from platelets: novel drivers of vascular inflammation. Thromb Haemost. 2015;114(08):228–236. doi:10.1160/TH14-11-0962
32. Yang X, Zhang Y, Zhang Y, et al. The key role of exosomes on the pre-metastatic niche formation in tumors. Front Mol Biosci. 2021;8:703640. doi:10.3389/fmolb.2021.703640
33. Marar C, Starich B, Wirtz D. Extracellular vesicles in immunomodulation and tumor progression. Nat Immunol. 2021;22(5):560–570. doi:10.1038/s41590-021-00899-0
34. Namee NM, O’Driscoll L. Extracellular vesicles and anti-cancer drug resistance. Biochimica Et Biophysica Acta Rev Cancer. 2018;1870(2):123–136. doi:10.1016/j.bbcan.2018.07.003
35. Aslan C, Maralbashi S, Salari F, et al. Tumor-derived exosomes: implication in angiogenesis and antiangiogenesis cancer therapy. J Cell Physiol. 2019;234(10):16885–16903. doi:10.1002/jcp.28374
36. Rabe DC, Walker ND, Rustandy FD, et al. Tumor extracellular vesicles regulate macrophage-driven metastasis through CCL5. Cancers. 2021;13(14):3459. doi:10.3390/cancers13143459
37. Njock M-S, O’Grady T, Nivelles O, et al. Endothelial extracellular vesicles promote tumour growth by tumour-associated macrophage reprogramming. J Extracell Vesicles. 2022;11(6):e12228. doi:10.1002/jev2.12228
38. De Rubis G, Rajeev Krishnan S, Bebawy M. Liquid biopsies in cancer diagnosis, monitoring, and prognosis. Trends Pharmacol Sci. 2019;40(3):172–186. doi:10.1016/j.tips.2019.01.006
39. Baassiri A, Nassar F, Mukherji D, Shamseddine A, Nasr R, Temraz S. Exosomal non coding RNA in LIQUID biopsies as a promising biomarker for colorectal cancer. Int J Mol Sci. 2020;21(4):1398. doi:10.3390/ijms21041398
40. Yang B, Xiong WY, Hou HJ, et al. Exosomal miRNAs as biomarkers of cancer: a meta-analysis. Clin Lab. 2019;65. doi:10.7754/Clin.Lab.2018.181011
41. Mu H, Zhang S, Yao Z, et al. The diagnostic and prognostic value of exosome-derived long non-coding RNAs in cancer patients: a meta-analysis. Clin Exp Med. 2020;20(3):339–348. doi:10.1007/s10238-020-00638-z
42. Zhang Y, Yang X, Zhuang Z, et al. The diagnostic value of exosomal circular RNAs in cancer patients: a systematic review and meta-analysis. Cancer Med. 2022;12(2):1709–1720. doi:10.1002/cam4.5012
43. Culliton BJ. HeLa (for Henrietta Lacks). Science. 1974;184(4143):1268. doi:10.1126/science.184.4143.1268
44. Ryan SL, Baird AM, Vaz G, et al. Drug discovery approaches utilizing three-dimensional cell culture. Assay Drug Dev Technol. 2016;14(1):19–28. doi:10.1089/adt.2015.670
45. Robinson NB, Krieger K, Khan FM, et al. The current state of animal models in research: a review. Int J Surg. 2019;72:9–13. doi:10.1016/j.ijsu.2019.10.015
46. Invrea F, Rovito R, Torchiaro E, Petti C, Isella C, Medico E. Patient-derived xenografts (PDXs) as model systems for human cancer. Curr Opin Biotechnol. 2020;63:151–156. doi:10.1016/j.copbio.2020.01.003
47. Xu H, Lyu X, Yi M, Zhao W, Song Y, Wu K. Organoid technology and applications in cancer research. J Hematol Oncol. 2018;11(1):116. doi:10.1186/s13045-018-0662-9
48. Joshi P, Lee MY. High Content Imaging (HCI) on miniaturized three-dimensional (3D) cell cultures. Biosensors. 2015;5(4):768–790. doi:10.3390/bios5040768
49. Jurj A, Pasca S, Braicu C, Rusu I, Korban SS, Berindan-Neagoe I. Focus on organoids: cooperation and interconnection with extracellular vesicles - is this the future of in vitro modeling? Semin Cancer Biol. 2021;86(Pt 3):367–381. doi:10.1016/j.semcancer.2021.12.002
50. Cekanova M, Rathore K. Animal models and therapeutic molecular targets of cancer: utility and limitations. Drug Des Devel Ther. 2014;8:1911–1921. doi:10.2147/DDDT.S49584
51. Cheon DJ, Orsulic S. Mouse models of cancer. Annu Rev Pathol. 2011;6(1):95–119. doi:10.1146/annurev.pathol.3.121806.154244
52. Seok J, Warren HS, Cuenca AG, et al. Genomic responses in mouse models poorly mimic human inflammatory diseases. Proc Natl Acad Sci U S A. 2013;110(9):3507–3512. doi:10.1073/pnas.1222878110
53. LeSavage BL, Suhar RA, Broguiere N, Lutolf MP, Heilshorn SC. Next-generation cancer organoids. Nat Mater. 2022;21(2):143–159. doi:10.1038/s41563-021-01057-5
54. Byrne AT, Alférez DG, Amant F, et al. Interrogating open issues in cancer precision medicine with patient-derived xenografts. Nat Rev Cancer. 2017;17(4):254–268. doi:10.1038/nrc.2016.140
55. Jabs J, Zickgraf FM, Park J, et al. Screening drug effects in patient-derived cancer cells links organoid responses to genome alterations. Mol Syst Biol. 2017;13(11):955. doi:10.15252/msb.20177697
56. Corrò C, Novellasdemunt L, Vsw L. A brief history of organoids. Am J Physiol Cell Physiol. 2020;319(1):C151–C65. doi:10.1152/ajpcell.00120.2020
57. Clevers H. Modeling development and disease with organoids. Cell. 2016;165(7):1586–1597. doi:10.1016/j.cell.2016.05.082
58. Wilson HV. A new method by which sponges may be artificially reared. Science. 1907;25(649):912–915. doi:10.1126/science.25.649.912
59. Weiss P, Taylor AC. Reconstitution of complete organs from single-cell suspensions of chick embryos in advanced stages of differentiation. Proc Natl Acad Sci U S A. 1960;46(9):1177–1185. doi:10.1073/pnas.46.9.1177
60. Friedenstein A J, Chailakhjan R K and Lalykina K S. (1970). THE DEVELOPMENT OF FIBROBLAST COLONIES IN MONOLAYER CULTURES OF GUINEA-PIG BONE MARROW AND SPLEEN CELLS. Cell Prolif, 3(4), 393–403. 10.1111/j.1365-2184.1970.tb00347.x
61. Charbord P. Bone marrow mesenchymal stem cells: historical overview and concepts. Hum Gene Ther. 2010;21(9):1045–1056. doi:10.1089/hum.2010.115
62. Takahashi K, Yamanaka S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell. 2006;126(4):663–676. doi:10.1016/j.cell.2006.07.024
63. Takahashi K, Tanabe K, Ohnuki M, et al. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell. 2007;131(5):861–872. doi:10.1016/j.cell.2007.11.019
64. Sato T, Vries RG, Snippert HJ, et al. Single Lgr5 stem cells build crypt-villus structures in vitro without a mesenchymal niche. Nature. 2009;459(7244):262–265. doi:10.1038/nature07935
65. Lee J, Sutani A, Kaneko R, et al. In vitro generation of functional murine heart organoids via FGF4 and extracellular matrix. Nat Commun. 2020;11(1):4283. doi:10.1038/s41467-020-18031-5
66. Shi R, Radulovich N, Ng C, et al. Organoid cultures as preclinical models of non-small cell lung cancer. Clin Cancer Res. 2020;26(5):1162–1174. doi:10.1158/1078-0432.CCR-19-1376
67. Hu Y, Sui X, Song F, et al. Lung cancer organoids analyzed on microwell arrays predict drug responses of patients within a week. Nat Commun. 2021;12(1):2581. doi:10.1038/s41467-021-22676-1
68. Dekkers JF, Whittle JR, Vaillant F, et al. Modeling breast cancer using CRISPR-Cas9-mediated engineering of human breast organoids. J Natl Cancer Inst. 2020;112(5):540–544. doi:10.1093/jnci/djz196
69. Saatci O, Kaymak A, Raza U, et al. Targeting lysyl oxidase (LOX) overcomes chemotherapy resistance in triple negative breast cancer. Nat Commun. 2020;11(1):2416. doi:10.1038/s41467-020-16199-4
70. Yao Y, Xu X, Yang L, et al. Patient-derived organoids predict chemoradiation responses of locally advanced rectal cancer. Cell Stem Cell. 2020;26(1):17–26.e6. doi:10.1016/j.stem.2019.10.010
71. Matano M, Date S, Shimokawa M, et al. Modeling colorectal cancer using CRISPR-Cas9-mediated engineering of human intestinal organoids. Nat Med. 2015;21(3):256–262. doi:10.1038/nm.3802
72. Schnalzger TE, de Groot MH, Zhang C, et al. 3D model for CAR -mediated cytotoxicity using patient-derived colorectal cancer organoids. EMBO j. 2019;38(12). doi:10.15252/embj.2018100928
73. Boj SF, Hwang CI, Baker LA, et al. Organoid models of human and mouse ductal pancreatic cancer. Cell. 2015;160(1–2):324–338. doi:10.1016/j.cell.2014.12.021
74. Below CR, Kelly J, Brown A, et al. A microenvironment-inspired synthetic three-dimensional model for pancreatic ductal adenocarcinoma organoids. Nat Mater. 2022;21(1):110–119. doi:10.1038/s41563-021-01085-1
75. Nuciforo S, Fofana I, Matter MS, et al. Organoid models of human liver cancers derived from tumor needle biopsies. Cell Rep. 2018;24(5):1363–1376. doi:10.1016/j.celrep.2018.07.001
76. Broutier L, Mastrogiovanni G, Verstegen MM, et al. Human primary liver cancer-derived organoid cultures for disease modeling and drug screening. Nat Med. 2017;23(12):1424–1435. doi:10.1038/nm.4438
77. Maier CF, Zhu L, Nanduri LK, et al. Patient-derived organoids of cholangiocarcinoma. Int J Mol Sci. 2021;22(16):8675. doi:10.3390/ijms22168675
78. Koh V, Chakrabarti J, Torvund M, et al. Hedgehog transcriptional effector GLI mediates mTOR-Induced PD-L1 expression in gastric cancer organoids. Cancer Lett. 2021;518:59–71. doi:10.1016/j.canlet.2021.06.007
79. Golebiewska A, Hau AC, Oudin A, et al. Patient-derived organoids and orthotopic xenografts of primary and recurrent gliomas represent relevant patient avatars for precision oncology. Acta Neuropathol. 2020;140(6):919–949. doi:10.1007/s00401-020-02226-7
80. Linkous A, Balamatsias D, Snuderl M, et al. Modeling patient-derived glioblastoma with cerebral organoids. Cell Rep. 2019;26(12):3203–11.e5. doi:10.1016/j.celrep.2019.02.063
81. Jacob F, Ming GL, Song H. Generation and biobanking of patient-derived glioblastoma organoids and their application in CAR T cell testing. Nat Protoc. 2020;15(12):4000–4033. doi:10.1038/s41596-020-0402-9
82. Kawasaki K, Toshimitsu K, Matano M, et al. An organoid biobank of neuroendocrine neoplasms enables genotype-phenotype mapping. Cell. 2020;183(5):1420–35.e21. doi:10.1016/j.cell.2020.10.023
83. Mullenders J, de Jongh E, Brousali A, et al. Mouse and human urothelial cancer organoids: a tool for bladder cancer research. Proc Natl Acad Sci U S A. 2019;116(10):4567–4574. doi:10.1073/pnas.1803595116
84. van de Wetering M, Francies HE, Francis JM, et al. Prospective derivation of a living organoid biobank of colorectal cancer patients. Cell. 2015;161(4):933–945. doi:10.1016/j.cell.2015.03.053
85. Vlachogiannis G, Hedayat S, Vatsiou A, et al. Patient-derived organoids model treatment response of metastatic gastrointestinal cancers. Science. 2018;359(6378):920–926. doi:10.1126/science.aao2774
86. Pauli C, Hopkins BD, Prandi D, et al. Personalized in vitro and in vivo cancer models to guide precision medicine. Cancer Discov. 2017;7(5):462–477. doi:10.1158/2159-8290.CD-16-1154
87. Aloia L, McKie MA, Vernaz G, et al. Epigenetic remodelling licences adult cholangiocytes for organoid formation and liver regeneration. Nat Cell Biol. 2019;21(11):1321–1333. doi:10.1038/s41556-019-0402-6
88. Huang L, Holtzinger A, Jagan I, et al. Ductal pancreatic cancer modeling and drug screening using human pluripotent stem cell- and patient-derived tumor organoids. Nat Med. 2015;21(11):1364–1371. doi:10.1038/nm.3973
89. Kim SY, Kim SM, Lim S, et al. Modeling clinical responses to targeted therapies by patient-derived organoids of advanced lung adenocarcinoma. Clin Cancer Res. 2021;27(15):4397–4409. doi:10.1158/1078-0432.CCR-20-5026
90. Dijkstra KK, Monkhorst K, Schipper LJ, et al. Challenges in establishing pure lung cancer organoids limit their utility for personalized medicine. Cell Rep. 2020;31(5):107588. doi:10.1016/j.celrep.2020.107588
91. Guillen KP, Fujita M, Butterfield AJ, et al. A human breast cancer-derived xenograft and organoid platform for drug discovery and precision oncology. Nat Cancer. 2022;3(2):232–250. doi:10.1038/s43018-022-00337-6
92. Fujii M, Shimokawa M, Date S, et al. A colorectal tumor organoid library demonstrates progressive loss of niche factor requirements during tumorigenesis. Cell Stem Cell. 2016;18(6):827–838. doi:10.1016/j.stem.2016.04.003
93. Ooft SN, Weeber F, Dijkstra KK, et al. Patient-derived organoids can predict response to chemotherapy in metastatic colorectal cancer patients. Sci Transl Med. 2019;11(513). doi:10.1126/scitranslmed.aay2574
94. Holokai L, Chakrabarti J, Lundy J, et al. Murine- and human-derived autologous organoid/immune cell co-cultures as pre-clinical models of pancreatic ductal adenocarcinoma. Cancers. 2020;12(12):3816. doi:10.3390/cancers12123816
95. Sun L, Wang Y, Cen J, et al. Modelling liver cancer initiation with organoids derived from directly reprogrammed human hepatocytes. Nat Cell Biol. 2019;21(8):1015–1026. doi:10.1038/s41556-019-0359-5
96. Lim JTC, Kwang LG, Ncw H, et al. Hepatocellular carcinoma organoid co-cultures mimic angiocrine crosstalk to generate inflammatory tumor microenvironment. Biomaterials. 2022;284:121527. doi:10.1016/j.biomaterials.2022.121527
97. Kim M, Mun H, Sung CO, et al. Patient-derived lung cancer organoids as in vitro cancer models for therapeutic screening. Nat Commun. 2019;10(1):3991. doi:10.1038/s41467-019-11867-6
98. Dekker E, Tanis PJ, Vleugels JLA, Kasi PM, Wallace MB. Colorectal cancer. Lancet. 2019;394(10207):1467–1480. doi:10.1016/S0140-6736(19)32319-0
99. McGuigan A, Kelly P, Turkington RC, Jones C, Coleman HG, McCain RS. Pancreatic cancer: a review of clinical diagnosis, epidemiology, treatment and outcomes. World J Gastroenterol. 2018;24(43):4846–4861. doi:10.3748/wjg.v24.i43.4846
100. Topalian SL, Taube JM, Anders RA, Pardoll DM. Mechanism-driven biomarkers to guide immune checkpoint blockade in cancer therapy. Nat Rev Cancer. 2016;16(5):275–287. doi:10.1038/nrc.2016.36
101. Highfill SL, Cui Y, Giles AJ, et al. Disruption of CXCR2-mediated MDSC tumor trafficking enhances anti-PD1 efficacy. Sci Transl Med. 2014;6(237). doi:10.1126/scitranslmed.3007974
102. Forner A, Reig M, Bruix J. Hepatocellular carcinoma. Lancet. 2018;391(10127):1301–1314. doi:10.1016/S0140-6736(18)30010-2
103. Ke X, Yan R, Sun Z, et al. Esophageal adenocarcinoma-derived extracellular vesicle MicroRNAs Induce a neoplastic phenotype in gastric organoids. Neoplasia. 2017;19(11):941–949. doi:10.1016/j.neo.2017.06.007
104. Zhao X, Yuan C, Wangmo D, Subramanian S. Tumor-secreted extracellular vesicles regulate T-cell costimulation and can be manipulated to induce tumor-specific T-cell responses. Gastroenterology. 2021;161(2):560–74.e11. doi:10.1053/j.gastro.2021.04.036
105. Huang L, Rong Y, Tang X, et al. Engineered exosomes as an in situ DC-primed vaccine to boost antitumor immunity in breast cancer. Mol Cancer. 2022;21(1):45. doi:10.1186/s12943-022-01515-x
106. Oszvald Á, Szvicsek Z, Pápai M, et al. Fibroblast-derived extracellular vesicles induce colorectal cancer progression by transmitting amphiregulin. Front Cell Dev Biol. 2020;8:558. doi:10.3389/fcell.2020.00558
107. Xu X, Cheng J, Luo S, et al. Deoxycholic acid-stimulated macrophage-derived exosomes promote spasmolytic polypeptide-expressing metaplasia in the stomach. Biochem Biophys Res Commun. 2020;524(3):649–655. doi:10.1016/j.bbrc.2020.01.159
108. Kelemen A, Carmi I, Oszvald Á, et al. IFITM1 expression determines extracellular vesicle uptake in colorectal cancer. Cell Mol Life Sci. 2021;78(21–22):7009–7024. doi:10.1007/s00018-021-03949-w
109. Zhuang J, Tan J, Wu C, et al. Extracellular vesicles engineered with valency-controlled DNA nanostructures deliver CRISPR/Cas9 system for gene therapy. Nucleic Acids Res. 2020;48(16):8870–8882. doi:10.1093/nar/gkaa683
110. Zhang Q, Deng T, Zhang H, et al. Adipocyte-derived exosomal MTTP suppresses ferroptosis and promotes chemoresistance in colorectal cancer. Adv Sci. 2022;9(28):e2203357. doi:10.1002/advs.202203357
111. Sándor GO, Soós A, Lörincz P, et al. Wnt activity and cell proliferation are coupled to extracellular vesicle release in multiple organoid models. Front Cell Dev Biol. 2021;9:670825. doi:10.3389/fcell.2021.670825
112. Fodde R, Smits R, Clevers H. APC, signal transduction and genetic instability in colorectal cancer. Nat Rev Cancer. 2001;1(1):55–67. doi:10.1038/35094067
113. Szvicsek Z, Oszvald Á, Szabó L, et al. Extracellular vesicle release from intestinal organoids is modulated by Apc mutation and other colorectal cancer progression factors. Cell Mol Life Sci. 2019;76(12):2463–2476. doi:10.1007/s00018-019-03052-1
114. Namba Y, Sogawa C, Okusha Y, et al. Depletion of lipid efflux pump ABCG1 triggers the intracellular accumulation of extracellular vesicles and reduces aggregation and tumorigenesis of metastatic cancer cells. Front Oncol. 2018;8:376. doi:10.3389/fonc.2018.00376
115. Taha EA, Sogawa C, Okusha Y, et al. Knockout of MMP3 weakens solid tumor organoids and cancer extracellular vesicles. Cancers. 2020;12(5):1260. doi:10.3390/cancers12051260
116. Nagai H, Kuroha M, Handa T, et al. Comprehensive analysis of microRNA profiles in organoids derived from human colorectal adenoma and cancer. Digestion. 2021;102(6):860–869. doi:10.1159/000513882
117. Buenafe AC, Dorrell C, Reddy AP, Klimek J, Marks DL. Proteomic analysis distinguishes extracellular vesicles produced by cancerous versus healthy pancreatic organoids. Sci Rep. 2022;12(1):3556. doi:10.1038/s41598-022-07451-6
118. Kelemen A, Carmi I, Seress I, et al. CD44 expression intensity marks colorectal cancer cell subpopulations with different extracellular vesicle release capacity. Int J Mol Sci. 2022;23(4):2180. doi:10.3390/ijms23042180
119. Rocha S, Carvalho J, Oliveira P, et al. 3D cellular architecture affects MicroRNA and protein cargo of extracellular vesicles. Adv Sci. 2019;6(4):1800948. doi:10.1002/advs.201800948
120. Hackam DG, Redelmeier DA. Translation of research evidence from animals to humans. JAMA. 2006;296(14):1731–1732. doi:10.1001/jama.296.14.1731
121. Wang W, Jo H, Park S, et al. Integrated analysis of ascites and plasma extracellular vesicles identifies a miRNA-based diagnostic signature in ovarian cancer. Cancer Lett. 2022;542:215735. doi:10.1016/j.canlet.2022.215735
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.