Back to Journals » Journal of Inflammation Research » Volume 18
Caffeic Acid Alleviates Chronic Sleep Deprivation-Induced Intestinal Damage by Inhibiting the IMD Pathway in Drosophila
Authors Yang D, Xiu M, Jiang X, Kang Q, Fu J, Zhou S, Liu Y ![]() , He J
, He J ![]()
Received 6 November 2024
Accepted for publication 25 February 2025
Published 10 March 2025 Volume 2025:18 Pages 3485—3498
DOI https://doi.org/10.2147/JIR.S500892
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Tara Strutt
Dan Yang,1,* Minghui Xiu,2,* Xiaolin Jiang,1 Qian Kang,1 Jinyu Fu,1 Shihong Zhou,2 Yongqi Liu,1,2 Jianzheng He1– 3
1Provincial-Level Key Laboratory for Molecular Medicine of Major Diseases and The Prevention and Treatment with Traditional Chinese Medicine Research in Gansu Colleges and University, Gansu University of Chinese Medicine, Lanzhou, 730000, People’s Republic of China; 2College of Public Health, Gansu University of Chinese Medicine, Lanzhou, 730000, People’s Republic of China; 3Scientific Research and Experimental Center, Gansu University of Chinese Medicine, Lanzhou, 730000, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Jianzheng He, Email [email protected]
Background: Sleep is vital for maintaining the health of the organism. Chronic sleep deprivation (CSD) is a key contributor to significant health risks, including the induction of gastrointestinal disorders. However, the mechanism of CSD caused intestinal damage remains unclear.
Methods: Drosophila melanogaster as an in vivo model was used to investigate the mechanism of CSD-induced intestinal injury, as well as the ameliorative effect of caffeic acid.
Results: CSD resulted in reduced survival and severely affected intestinal homeostasis in flies, as evidenced by disruption of intestinal acid-base homeostasis, increased feeding, increased intestinal permeability and shortened intestinal length. Meanwhile, the expressions of the immune deficiency (IMD) pathway-related genes PGRP-SB1, Dpt, AttA, AttB and Mtk were significantly up-regulated in the intestine of CSD flies. On the other hand, Caffeic acid supplementation restored intestinal acid-base homeostasis and intake, while improving intestinal barrier permeability and intestinal length, and effectively reducing intestinal damage. In addition, administration of caffeic acid decreased the expressions of PGRP-SB1, Dpt, AttA and Mtk genes in the CSD flies gut.
Discussion: These results suggested that CSD could disrupt gut homeostasis in adult flies by overactivating the IMD pathway, while Caffeic acid has an obvious protective role on the gut homeostasis.
Keywords: thermogenetic CSD, intestine, Immune signaling, polyphenolic compounds, Drosophila melanogaster
Graphical Abstract:

Introduction
As people’s lifestyles change, sleep time has severely decreased, with nearly one-third of adults sleep less than 6 h per day.1 Chronic sleep deprivation (CSD) has a serious impact on human health and leads to various diseases, such as heart disease, diabetes, and cancer, even leading to death.2,3 Multiple studies have proved that the intestinal damage caused by CSD is the major factor for premature death.4–6 However, the molecular mechanism of CSD induced intestinal damage is still unclear. Therefore, it is necessary to dissect the mechanism of intestinal damage caused by CSD and find the effective therapeutic drugs.
CSD can cause serious intestinal damage, such as loss of intestinal mucosal integrity and the occurrence of intestinal inflammation.7,8 Current research has found that oxidative stress, inflammatory response, and imbalance of gut microbiota are the major causes of intestinal damage caused by CSD.9,10 After sleep deprivation, the levels of antioxidant enzymes (GSH-PX, SOD, and CAT) in the intestine are down regulated, the expression of inflammatory pathway NF-κB is up-regulated, and the diversity and abundance of gut microbiota are significantly decreased.6,11 Notably, innate immunity as the first line of defense is crucial for maintaining intestinal homeostasis.12,13 Immune deficiency (IMD) and Toll signaling pathways, two highly conserved recognition and signaling cascades, control the innate immune response in flies.14 When the intestinal homeostasis is imbalanced, the intestinal pathway and Toll pathway are activated, and increase the proliferation of intestinal stem cells, promoting the regeneration of flies intestinal epithelium and enhancing the systemic immune response.15,16 Conversely, there is an up-regulation of IMD pathway-associated AMP genes expression in the intestine of aged flies, leading to intestinal homeostasis imbalance and a reduction in fruit fly lifespan.17 However, the precise contribution of innate immunity to intestinal injury induced by CSD remains elusive.
Drosophila melanogaster serves as an exemplary model organism for investigating the intricate interplay among gut injury, sleep and health. The major signaling pathways associated with immunity and tissue regeneration are conserved in both flies and humans. Moreover, flies and humans share strikingly similar sleep patterns governed by their endogenous circadian clocks.18,19 Although previous studies have demonstrated that oxidative damage led to the gut hastens premature mortality in CSD flies, the underlying molecular mechanism linking gut injury with sleep deprivation remains largely unexplored.4
There is a relative dearth of research on treatment strategies for intestinal injury induced by CSD. Currently, antioxidants such as ebselen, glutathione, and methylene blue have been proven to be ineffective in rescuing the survival rate in CSD flies.4 In recent years, natural small molecules have garnered increasing attention due to their efficacy and low toxicity, exhibiting potential for treating various intestinal disorders.20–22 Caffeic acid is a polyphenolic compound with notable impact on diverse diseases, particularly in the context of intestinal ailments like ulcerative colitis and colon cancer treatment.23–25 Specifically, caffeic acid inhibits NF-κB pathway and chemokine biosynthesis, attenuating mucin content and restoring gut microbiota diversity in murine models of colitis, ultimately modulating intestinal inflammation and lesions.26,27 Additionally, caffeic acid inhibits the proliferation of HCT 15 colon cancer cells, induces apoptosis in these cells, and prevents metastasis by targeting MEK1 and TOPK.28,29 Nevertheless, the preventive and therapeutic effects of caffeic acid on sleep deprivation-induced intestinal damage remain elusive.
In this study, we investigated the underlying mechanism of intestinal injury induced by CSD and explored the therapeutic effect of caffeic acid using adult fly as a model organism. The results demonstrated that CSD led to excessive activation of the intestinal IMD pathway, while highlighting caffeic acid as a potential intervention for mitigating such injuries through inhibition of the IMD pathway. In summary, our study offers novel insights into the mechanisms and therapeutic strategies about intestinal injury induced by CSD.
Materials And Methods
Fly Stocks and Husbandry
The following flies’ were obtained from the Bloomington Drosophila stock center (BDSC; Indiana, USA): w1118 (#5905); 60D04-Gal4 (#45356); 11H05-Gal4 (#45016); UAS-TrpA1 (#26263). w1118 represents the genetic background and is often used as the appropriate controls. 11H05- and 60D04-Gal4 target neurons in the brain whose activation leads to loss of sleep in flies. Heat-activated cation channel TrpA1 activates neural circuits in flies under temperature control. Flies were raised at 25 °C, 60% humidity under a 12h light/12h dark cycle unless otherwise stated. Flies were maintained on a standard cornmeal-agar medium.
Sleep Deprivation
Flies genotypes 11H05>TrpA1 or 60D04>TrpA1 were obtained by crossing 11H05-Gal4 flies or 60D04-Gal4 flies with UAS-TrpA1 flies using the UAS/Gal4 system and cultured at 18°C. After eclosion, male flies were raised at multifunctional artificial climate chamber at a temperature of 29°C, 12:12 LD for 10 d.
Mechanical sleep deprivation was performed at 25°C using a multifunctional oscillator. After eclosion, w1118 flies were reared at room temperature for 2 d, then flies were placed into tubes containing standard food and fixed between shock-absorbing sponge plates. The multifunctional oscillator speed was 1100 rpm and delivered 10s-long vibrations at random intervals centered around 180s (±30s) (Supplementary Figure 1). Relevant tests were performed after 10 d of sleep deprivation.
Drug Treatment
All chemicals used are analytical grade. Caffeic acid was purchased from Shanghai Yuanye Biotechnology Co., Ltd, Shanghai, China, and purity levels exceeding 98%. Caffeic acid was dissolved in anhydrous ethanol for 20 mm and then was diluted in the standard food to achieve final concentrations of 0.2 mm for further experiments.
Sleep Monitoring
Adult male flies were individually inserted into 5×65 mm glass tubes, loaded into the Drosophila Activity Monitoring Systems (DAMS, Wuhan Yi Hong Technology Co., Ltd., China), and housed under 12:12 h light/dark cycle conditions.30 Flies were kept in this condition for 5 d, without sleep analysis for the first 2 d that allowed flies to acclimatize to the environmental conditions. Data were analyzed using ShinyR-DAM (https://karolcichewicz.shinyapps.io/shinyr-dam/) software to count sleep-wake cycle, total sleep time, daytime sleep time, nighttime sleep time, sleep bout number, sleep bout length, and sleep latency. For each genotype, a minimum of 15 males were tested.
Lifespan Assay
Adult (2–4 days-old) male and female flies were reared on food with or without 0.2 mm caffeic acid. Fresh food was changed every three days and the number of deaths was recorded daily until all flies died. Three independent replicates were set up for each group with a minimum of 3 bottles per replicate.
Bromophenol Blue Treatment
The acid–base homeostasis was observed using the bromophenol blue assay as previously described,31 200 μL of 2% Bromophenol blue sodium was added to a vial containing 1% agar medium, and several small holes were made in the surface of the food with a pipette to ensure adequate absorption. All flies were starved for 6 h followed by feeding of pH indicator Bromophenol Blue (Sigma, B5525) for 4 h. Images were taken immediately after gut dissection. The type of the GI tract of flies fed with Bromophenol blue include ‘Homeostasis’ (CCR area: yellow), CCR is acidic and the rest of the gut is alkaline; “Perturbed A” (CCR area: blue), the acidic region is lost and the whole gut is basic; “Perturbed B” (CCR area: light blue), the strongly acidic region is lost and the rest of the gut becomes less alkaline.32
Food Intake Assay
Flies were starved for 18 h and then transferred to staining medium for 3 h. The staining medium was composed of 2% sodium bromophenol blue, 5% sucrose and 1% agarose. Fly abdominal color was observed under a light microscope and scored.30 Abdominal score was recorded from 0 to 3, in which 0 (colorless abdomen), 1 (less than 1/3 the length of the abdomen), 2 (between 1/3 and 2/3 the length of the abdomen) and 3 (more than 2/3 the length of the abdomen). The experiment was repeated independently at least 3 times.
Excretion Measurement
The fly excretion measurement was performed as previous study.31 In brief, after starvation for 3 h, flies were placed in 2 mL centrifuge tube with small holes in tube wall and dyed medium in tube cover for 18 h. The dyed medium was composed of 5% sucrose, 2% Bromophenol blue sodium, and 1% agarose. The centrifuge tubes were then washed with phosphate buffered saline (PBS). Afterwards, the excretion was determined by measuring the absorbance at 625 nm using an enzyme meter. Three replicates were established for each group.
“Smurf” Assay
Smurf assay was performed as previously described.33 Briefly, flies were fed with or without 0.2 mm caffeic acid for 10 d. After starved for 3 h, flies were incubated on staining medium for 18 h. The dyed medium consisted of 2% Bromophenol blue sodium, 5% sucrose and 1% agar. Smurf flies with the blue colour visible outside the digestive tract were counted. Three biological replicates were used for the experiment.
Intestinal Length
Guts were dissected in cold PBS, and length of the intestines were observed under a light microscope. Length measurement tool is vernier caliper. The experiment was repeated independently more than 10 times.
RNA Sequencing and Analysis
Total RNA was extracted from tissues using TRIzol reagent according to the instructions. RNA purity was characterized and quantified using a NanoDrop 2000 spectrophotometer (Thermo Scientific, USA) and RNA integrity was assessed using an Agilent 2100 Bioanalyzer (Agilent Technologies, Santa Clara, CA, USA). Transcriptome libraries were constructed using the VAHTS Universal V5 RNA-seq Library Prep kit according to the instructions. Transcriptome sequencing and analysis were performed by Shanghai Ouyi Biotechnology Co.
Quantitative RT-PCR Analysis
Intestines were dissected in pre-cooled PBS, and total RNA was extracted from tissues dissected from 60 male or female flies using TRIzol (Invitrogen).30 Total RNA was processed by Hieff® reverse transcriptase (Shanghai YEASEN, China) to synthesize DNA (cDNA). Quantitative PCR was performed using the GFX 96 Connect™ Optics Module (Bio-Rad Laboratories) following the instructions of the Multiplex PCR Master Mix (Shanghai YEASEN, China). The relative expression of genes was calculated using the 2 −ΔΔct method and normalized to the relative expression of the housekeeping gene Rp49. The experiment was repeated independently at least three times. All primers are shown in Table 1.
 |
Table 1 List of the Primer Sequences Used in the Gene Expression Analysis |
Statistical Analysis
Statistical analysis was performed using Prism 9.3.0 (GraphPad software). Three or more experimental groups were analyzed using one-way ANOVA followed by Tukey’s multiple-comparison test. Differences between groups were tested using unpaired two-tailed Student’s t-test and Fisher’s exact test. The Log rank test was used to determine the statistical significance of survival rates between groups. The significance level was indicated as *p < 0.05, **p < 0.01, and ***p < 0.001.
Results
Thermogenetic Sleep Deprivation Leads to Intestinal Homeostasis Imbalance in Adult Flies
CSD have severe detrimental effects on the body, potentially leading to mortality.4 To examine the effect of CSD on the gut, we used two specific genotypes 60D04>TrpA1 and 11H05>TrpA1 as SD model. The results showed that sleep was disrupted in both thermogenetic SD flies compared to the control flies (Figure 1A and B). Total sleep time, day sleep time, night sleep time, and sleep bout length in both thermogenetic SD flies at day 10 from continuous CSD were significantly decreased compared to control flies (Supplementary Figure S2A–H), while thermogenetic SD flies had increased sleep bout number and sleep latency (Supplementary Figure S2I–L). In addition, thermogenetic SD flies had considerably reduced survival rate compared to control flies, in which a much greater death rate was discovered when thermogenetic SD flies were cultured in 29°C environment after 10 d (Figure 1C and D).4 Therefore, the present study analyzed the intestinal homeostasis of flies at day 10 from continuous CSD. The gut tract in control flies exhibited acid-base homeostasis and maintained the integrity of the intestinal barrier to ensure proper physiological functioning.34,35 Thermogenetic CSD flies showed dramatically impaired intestinal digestive function as demonstrated by disruption of intestinal acid-base balance (Figure 1E–G) and increased food intake (Figure 1H and I). Nevertheless, thermogenetic CSD flies had the similar excretion compared to the control flies (Supplementary Figure S3A and B). An analysis of the intestinal length of flies also revealed no significant difference in intestinal length (Supplementary Figure S3C and D). Overall, thermogenetic CSD could impair gut homeostasis, resulting in premature death.
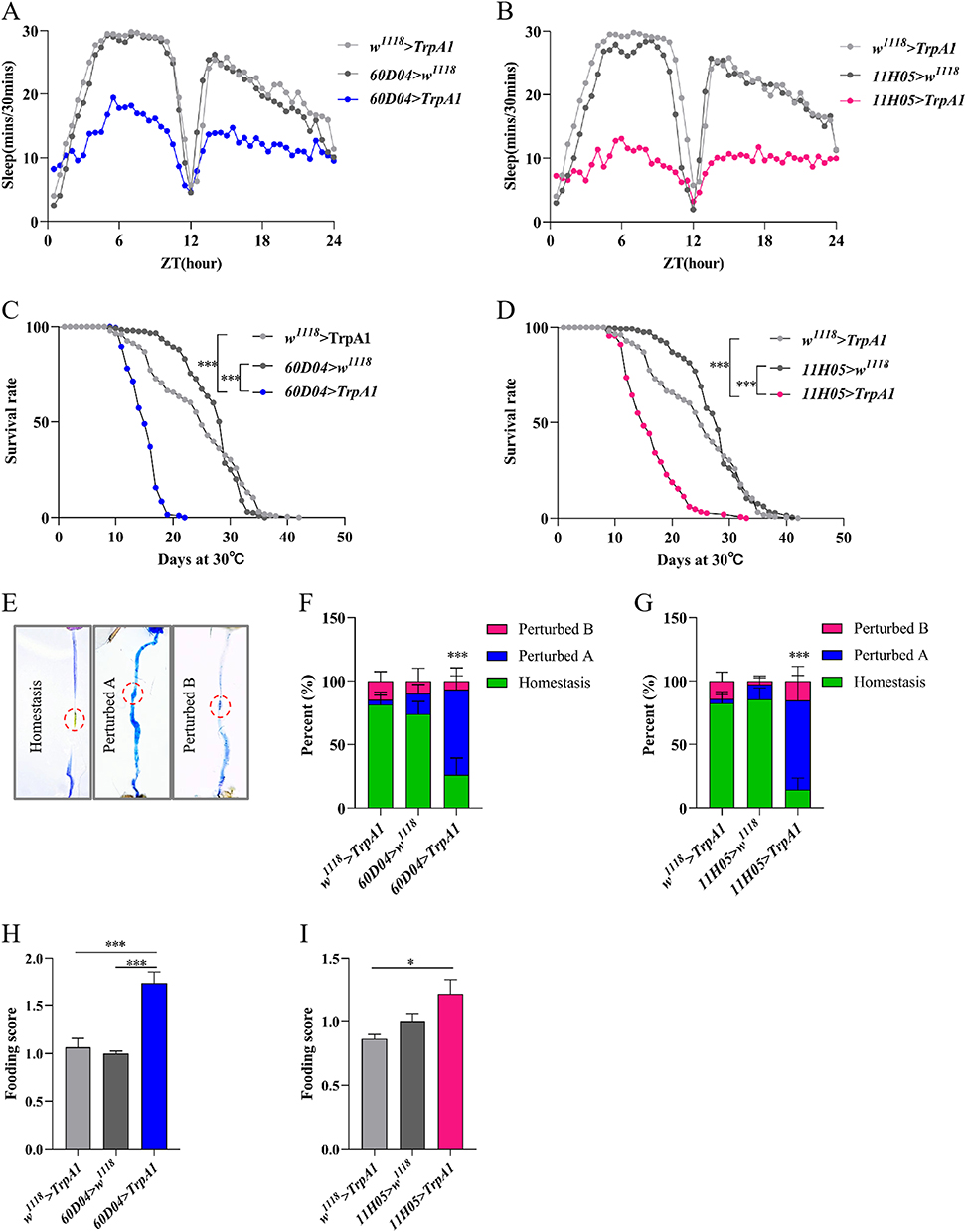 |
Figure 1 Thermogenetic CSD causes a decrease in intestinal function in flies. (A) Sleep traces in 60D04>TrpA1 flies and the control flies during a 24-h day (n=18-21). (B) Sleep traces in 11H05>TrpA1 flies and the control flies (n=21). (C) Survival curve of 60D04>TrpA1 flies and controls flies at 29°C (n=220-240). (D) Survival curve of 11H05>TrpA1 and controls flies at 29°C (n=220-240). (E) Representative images of intestinal acid- base homeostasis. (F and G) Quantification of the percentage of intestinal acid–base homeostasis of the flies (n=27-36). The copper cells region(CCR) is indicated by a circle. (H and I) Food intake of different genetic flies (n=24-30). Data are presented as means±S.E.M. *p< 0.05 and ***p< 0.001. |
Mechanical Sleep Deprivation Causes Intestinal Damage in Adult Flies
The sleep patterns of flies can be adversely affected by several environmental stimuli, such as vibration, sound, and light.36 Furthermore, mechanical shocks produced by a mechanical sleep deprivation apparatus were used to induce sleep deprivation in flies. Mechanical CSD was also found to dramatically diminish survival time in flies (Figure 2A). Mechanical CSD flies had disrupted intestinal acid-base homeostasis (Figure 2B) and increased food intake (Figure 2C) compared to control flies, but without impairing excretory function (Figure 2D). The integrity of the intestinal barrier was disrupted in adult male flies after mechanical CSD stimulation (Figure 2E). A considerable shorten of gut length in mechanical CSD flies was found compared to control flies (Figure 2F). Therefore, these results indicated that both forms of CSD could negatively affect gut function.
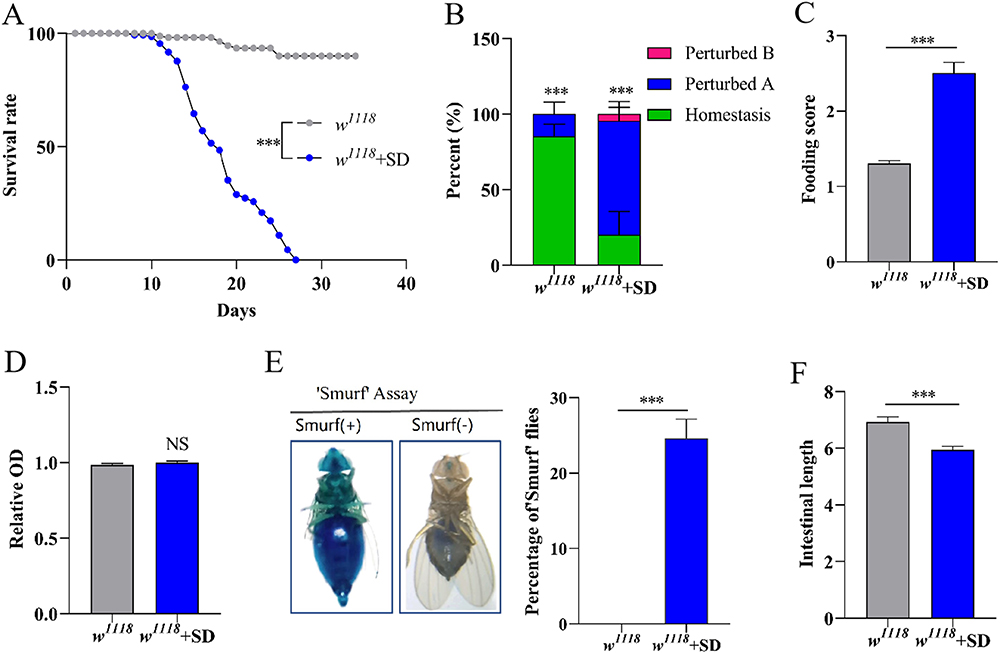 |
Figure 2 Mechanical CSD disrupts gut homeostasis in flies. (A) Survival rate of flies with mechanical CSD (n=110-120). (B) Quantification of the percentage of gut acid–base homeostasis of flies (n=27-36). (C) Food intake of flies (n=24-30). (D) Determination of excretion in flies (n=21-30). (E) Proportion of “Smurfs” in flies (n=21-30). (F) The relative length of the gut (n=15-20). ***p< 0.001 indicates significant difference between w1118 and w1118+SD. |
Analysis of CSD-Induced Genetic Changes in Flies
To determine the biological processes of CSD-induced intestinal damage, we performed RNA sequencing (RNA-seq) in the thermogenetic genotype 60D04>TrpA1 flies, and two control w1118>TrpA1 and 60D04>w1118 flies. Principal component analysis (PCA) showed that the principal components of three samples drove the differences among the samples, and all samples exhibited strong unique clustering in genotypes (Figure 3A). Next, the overlapping differential expressed genes were analyzed in these two pairwise comparisons (1) 60D04>TrpA1 vs w1118>TrpA1; (2) 60D04>TrpA1 vs 60D04> w1118. Compared to both control group flies, 60D04>TrpA1 flies had 31 up-regulated genes and 66 down-regulated genes (Figure 3B). Functional gene ontology (GO) analysis on the differential expressed genes revealed that various biological processes significantly altered in thermogenetic CSD flies. GO analysis showed a clear distinction between terms enriched in up-regulated gene categories (innate immunity, defense against bacteria, and humoral immunity) and down-regulated gene categories (metabolism and proteolysis) specific profiles (Figure 3C). The differential expression of genes was examined by the enrichment analysis of the Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway (Figure 3D). The most focused projects were mainly related to Toll and Imd signaling pathways, metabolic pathways, Hippo signaling pathways, and autophagy signaling pathways. Thus, the identified differential genes were mainly enriched in immune, apoptosis, and autophagy-related items.
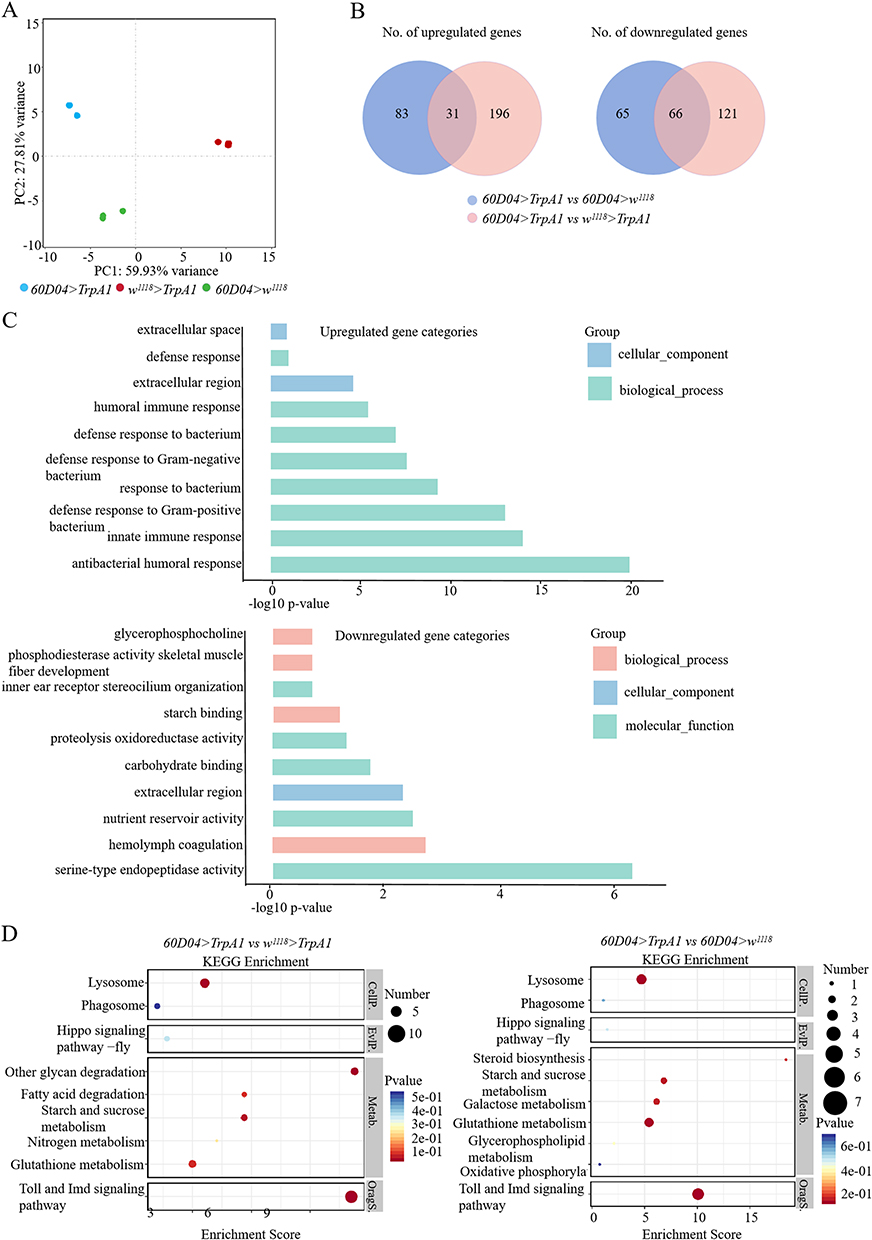 |
Figure 3 Thermogenetic CSD causes upregulation of immune response-related genes. (A) Principal component analysis (PCA) plot showing the variance among the genotypes used in RNA-seq. (B) Venn diagram showing the overlap of differentially regulated genes in 60D04>TrpA1 versus control w1118>TrpA1, 60D04>TrpA1 versus control 60D04>w1118 in RNA-seq analysis. The numbers of up- and down-regulated genes shown in the Venn diagrams are based on >1.5 fold-change. (C) GO analysis of biological processes of up-regulated (up) and down-regulated (down) genes based on their–log10 of FDR. (D) KEGG enrichment analysis of differentially regulated genes in RNA-seq analysis of 60D04>TrpA1 versus control w1118>TrpA1, 60D04>TrpA1 versus control 60D04>w1118. |
CSD Activates the IMD Signaling in the Intestine
To explore the effect of CSD on the innate immunity, the transcript levels of IMD pathway-related genes PGRP-SB1, AttA, AttB, Dpt and Mtk were analyzed. CSD resulted in significantly up-regulated expression of these genes based on the transcriptome result (Figure 4A). To further confirm whether CSD could activate the IMD pathway, qRT-PCR was utilized to investigate the expression of these genes. The expression levels of these immune genes were dramatically enhanced in the intestine of thermogenetic CSD flies (Figure 4B and C). Therefore, IMD pathway in the intestine could be activated by CSD.
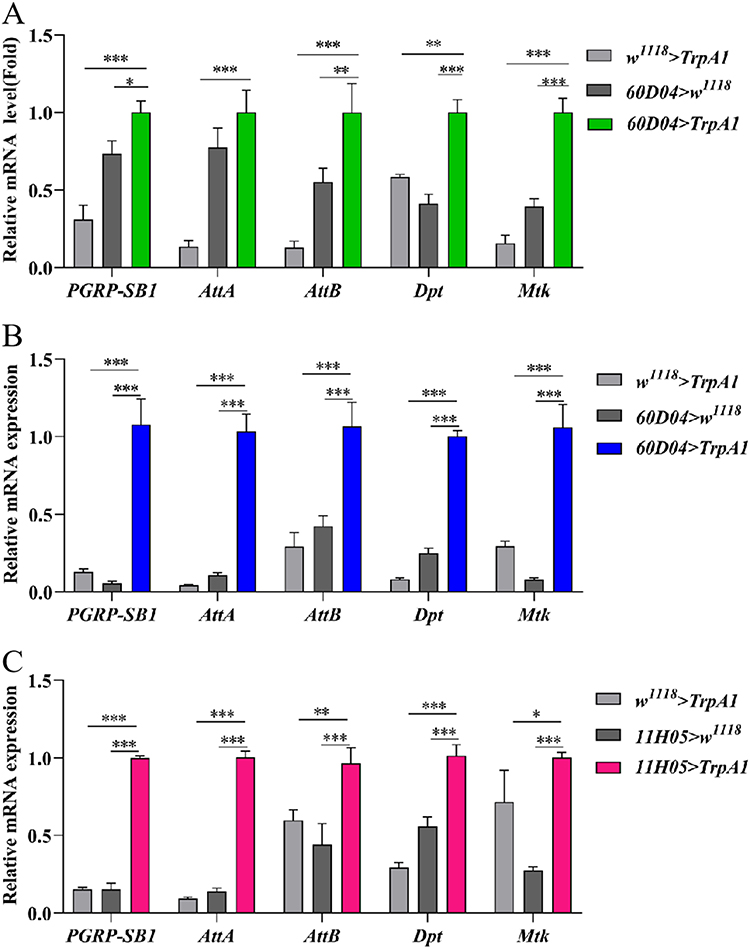 |
Figure 4 CSD activates the intestinal IMD signaling pathway. (A) Gene expression of innate immune-related genes in thermogenetic CSD flies (n=3). (B and C) mRNA levels of IMD pathway-related genes in thermogenetic CSD flies (n=3). Data are presented as means±S.E.M. *p< 0.05, **p< 0.01, ***p< 0.001. |
Caffeic Acid Rescues Mechanical CSD-Induced Death and Intestinal Damage in Adult Flies
Next, we determined the protective role of caffeic acid against mechanical CSD-induced injury. Administration of 0.2 mm caffeic acid remarkably extended the survival rate of mechanical SD flies (Figure 5A). Meanwhile, caffeic acid supplementation could rescue the intestinal acid-base homeostasis and reduce the food consumption in mechanical CSD flies (Figure 5B and C). In addition, the disrupted intestinal barrier was significantly improved in mechanical CSD flies (Figure 5D). Overall, these results demonstrated that caffeic acid could increase the survival duration and alleviate the disrupted intestine function caused by mechanical CSD.
 |
Figure 5 Caffeic acid prolongs survival time and protects intestinal homeostasis in mechanically CSD flies. (A) caffeic acid improves survival of flies with mechanical CSD (n=110-120). (B) Caffeic acid restores acid-base homeostasis in the intestinal tract of flies with mechanical CSD (n=27-36). (C) Reduced food intake in mechanical CSD flies after fed with caffeic acid (n=24-30). (D) Mechanical CSD flies fed with caffeic acid reduce proportion of “Smurf” flies (n=21-30). Compared to w1118+SD, **p< 0.01 and ***p< 0.001 indicate significant difference. |
Caffeic Acid Alleviates Thermogenetic CSD Induced Intestinal Damage via the Inhibition of the IMD Pathway
Flies survival and intestinal function were evaluated in thermogenetic CSD flies after feeding 0.2 mm caffeic acid. Administration of caffeic acid significantly extended the survival rate of the thermogenetic CSD flies (Figure 6A and B). Meanwhile, in both thermogenetic CSD flies, caffeic acid supplementation remarkably rescued the disrupted intestinal acid-base balance (Figure 6C and D). Additionally, it was seen that the food intake was also reduced in thermogenetic CSD flies after fed with 0.2 mm caffeic acid (Figure 6E and F). These results further indicated that caffeic acid could alleviate CSD-induced intestinal injury. Next, we detected whether caffeic acid could regulate IMD signaling pathway in the intestine of CSD flies. The results showed that the expressions of IMD pathway-related genes PGRP-SB1, Dpt, AttA, AttB and Mtk were significantly reduced in the intestine of thermogenetic CSD flies after fed with 0.2 mm caffeic acid for 10 d (Figure 6G). Therefore, these findings suggested that caffeic acid has ability to ameliorate CSD-induced intestinal damage by reducing IMD pathway overactivation.
 |
Figure 6 Caffeic acid improves survival and maintains intestinal homeostasis in thermogenetic CSD flies by inhibiting the IMD pathway. (A and B) Caffeic acid improves survival in CSD flies (n=110–120). (C and D) Caffeic acid restores acid-base homeostasis in the fly gut (n=27-36). (E and F) Caffeic acid reduces food intake in CSD flies (n=24-30). (G) Caffeic acid reduces mRNA levels of genes related to the intestinal IMD pathway in CSD flies (n=3). *p< 0.05, **p< 0.01, and ***p< 0.001. |
Discussion
In recent years, there are a lot of discussion regarding how CSD affects the gut. Increased oxidative stress and catabolism in the gut have been documented in rodent models and clinical trials.4,37 However, the complicated link between sleep and gut immunity is still largely unknown. To investigate the mechanism of CSD damage in the intestinal tract and to clarify the potential protective effect of caffeic acid, this study utilized adult flies as a model organism for experimental evaluation. Here, we found that CSD could damage gut homeostasis and intestinal function in flies. Furthermore, IMD signaling pathway in the gut was overactivated during the course of CSD. Finally, caffeic acid partially rescued gut function in CSD flies by blocking the IMD pathway.
Previous study has found that CSD could lead to early death mainly via oxidative stress damage in the gut of animals, suggesting that gut homeostasis is linked to organism health.4 In this study, the thermogenetic sleep deprivation and mechanical CSD fly models were constructed to detect the damaging effect of CSD on the gut. Intestinal function was disrupted in both CSD flies species, as evidenced by an imbalance in intestinal acid-base homeostasis and an increase in food intake. We found that mechanical CSD had a more significant injury on gut function compared to thermogenetic CSD, which may be because mechanical stimulation could engage the homeostat and also induce the serious body injury except gut. It is important to note that mechanical CSD could lead to an increase in fly intestinal permeability and a significant shortening of intestinal length. This suggested that physical shocks may cause additional damage to flies. Previous studies found that sleepless (sssΔ)38 mutant flies with sleep deprivation had an imbalance in intestinal acid-base homeostasis, increased food intake, and disruption of the intestinal barrier, compared to control flies.38 Another study found that CSD led to increased intestinal permeability and decreased colon length in mice.10 These studies confirm that sleep deprivation severely disrupts intestinal homeostasis, laying the foundation for the next steps in exploring its molecular mechanisms.
It has been proved that CSD could induce intestinal injury by promoting the development of intestinal oxidative stress and activation of the NF-κB pathway in mice and adult flies.6,7,39,40 Here, the results of transcriptomics testing revealed that CSD-induced intestinal damage may be associated with biological processes including innate immunity. Analysis of KEGG enrichment of up-regulated genes revealed that CSD induced hyperactivation of the IMD pathway in flies. It has been shown that over-activation of the IMD signaling pathway in flies leads to an imbalance in intestinal homeostasis, which further exacerbates intestinal damage and even death.41 The Drosophila gene encodes 13 PGRPs, in which the expression of PGRP-SB1 is highly induced upon infection, far more than any other PGRP.42 PGRP-SB1 massively activates the IMD signaling pathway by recognizing the diaminopimelic acid peptidoglycan (DAP) of Gram-negative bacterial cell walls.43 We found that CSD led to massive expression of PGRP-SB1, Dpt, AttA, AttB and Mtk in flies by using transcriptomics and RT-PCR. The results of our preliminary experiments found that CSD led to the accumulation of ROS in the fly gut. The expression of genes involved in the immune response could increase in response to oxidative stress.44 Thus, CSD-induced oxidative stress in the gut may underlie the hyperactivation of the IMD pathway.
Caffeic acid is a polyphenolic compound extensively present in various plants.26,45 Numerous pharmacological activities of caffeic acid have been reported, including antioxidative, anti-inflammatory, and antibacterial activities, and it has also the potential to attenuate intestinal injury.24,46,47 It was found that caffeic acid could ameliorate DSS-induced ulcerative colitis in mice, significantly reducing intestinal mucosal inflammation.48 Here, administration of caffeic acid could significantly reduce mortality of CSD flies. Meanwhile, it ameliorated CSD-induced intestinal damage, including restoration of intestinal acid-base homeostasis, decreased food intake, and decreased intestinal permeability. Previous studies have proved that caffeic acid ameliorated DSS-induced intestinal injury by affecting the intestinal microbiota and activating the Nrf-2/HO-1 pathway to reduce intestinal oxidative stress and inflammatory responses in mice.49 Here, we found that caffeic acid restored intestinal function and homeostasis by inhibiting the IMD pathway in the fly gut, suggesting that caffeic acid has a protective effect against CSD-induced intestinal immune injury. However, the possibility that caffeic acid ameliorates CSD intestinal damage by affecting intestinal oxidative stress and gut microbiota lacks demonstration and warrants further consideration in subsequent studies.
This study reveals that CSD induces hyperactivation of the IMD pathway in the gut, providing a new perspective on the relationship between CSD and the gut. The results showed that CSD could disrupt the intestine, including disruption of intestinal acid-base homeostasis, increased uptake, increased intestinal permeability, and shortened intestinal length. Consumption of a diet rich in caffeic acid has a highly significant beneficial effect on CSD-induced intestinal injury. In conclusion, our findings suggest that the IMD pathway is a promising avenue for the treatment of CSD gut injury. Caffeic acid significantly reduced mortality and ameliorated CSD-induced intestinal injury in flies with CSD by inhibiting the IMD pathway. In summary, caffeic acid is a new and promising compound for ameliorating CSD-induced intestinal damage.
Acknowledgments
We thank the Bloomington Stock Center (BSC) for providing the fly stocks. The authors appreciate the financial support received from the National Natural Science Foundation of China (No. 82104562), Gansu Natural Science Foundation (No. 23JRRA1202), Gansu Higher Education Industry Support Plan Project (No. 2025cYzc-47), Foundation of Key Laboratory of Dunhuang Medicine and Translational Education Ministry of Education (No. DHYX24-02).
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Disclosure
The authors declare that they have no conflicts of interest in this work.
References
1. Ford ES, Cunningham TJ, Croft JB. Trends in Self-Reported Sleep Duration among US Adults from 1985 to 2012. Sleep. 2015;38(5):829–832. doi:10.5665/sleep.4684
2. Garbarino S, Lanteri P, Bragazzi NL, Magnavita N, Scoditti E. Role of sleep deprivation in immune-related disease risk and outcomes. Commun Biol. 2021;4(1):1304. doi:10.1038/s42003-021-02825-4
3. Liew SC, Aung T. Sleep deprivation and its association with diseases- a review. Sleep Med. 2021;77:192–204. doi:10.1016/j.sleep.2020.07.048
4. Vaccaro A, Kaplan DY, Nambara K, et al. Sleep Loss Can Cause Death through Accumulation of Reactive Oxygen Species in the Gut. Cell. 2020;181(6):1307–1328. doi:10.1016/j.cell.2020.04.049
5. Nakada Y, Sadoshima J. Sleep deficiency and mortality: is the solution in the gut? Cardiovasc Res. 2021;117(2):e26–e28. doi:10.1093/cvr/cvaa309
6. Gao T, Wang Z, Dong Y, et al. Role of melatonin in sleep deprivation-induced intestinal barrier dysfunction in mice. J Pineal Res. 2019;67(1):e12574. doi:10.1111/jpi.12574
7. Gao T, Wang Z, Dong Y, Cao J, Chen Y. Butyrate Ameliorates Insufficient Sleep-Induced Intestinal Mucosal Damage in Humans and Mice. Microbiol Spectr. 2023;11:e200022. doi:10.1128/spectrum.02000-22
8. Gao T, Wang Z, Cao J, Dong Y, Chen Y. The Role of Aeromonas-Goblet Cell Interactions in Melatonin-Mediated Improvements in Sleep Deprivation-Induced Colitis. Oxid Med Cell Longev. 2022;2022:8133310. doi:10.1155/2022/8133310
9. Wang T, Wang Z, Cao J, Dong Y, Chen Y. Melatonin prevents the dysbiosis of intestinal microbiota in sleep-restricted mice by improving oxidative stress and inhibiting inflammation. Saudi J Gastroenterol. 2022;28(3):209–217. doi:10.4103/sjg.sjg_110_21
10. Gao T, Wang Z, Cao J. Melatonin Ameliorates Corticosterone-Mediated Oxidative Stress-Induced Colitis in Sleep-Deprived Mice Involving Gut Microbiota. Oxid Med Cell Longev. 2021;2021:9981480.
11. Yang DF, Huang WC, Wu CW, Huang CY, Yang Y-CSH, Tung Y-T. Tung, Acute sleep deprivation exacerbates systemic inflammation and psychiatry disorders through gut microbiota dysbiosis and disruption of circadian rhythms. Microbiol Res. 2023;268:127292. doi:10.1016/j.micres.2022.127292
12. You XY, Zhang HY, Han X, Wang F, Zhuang PW, Zhang YJ. Intestinal Mucosal Barrier Is Regulated by Intestinal Tract Neuro-Immune Interplay. Front Pharmacol. 2021;12:659716. doi:10.3389/fphar.2021.659716
13. Ansaldo E, Slayden LC, Ching KL, et al. Akkermansia muciniphila induces intestinal adaptive immune responses during homeostasis. Science. 2019;364(6446):1179–1184. doi:10.1126/science.aaw7479
14. Williams JA, Sathyanarayanan S, Hendricks JC, Sehgal A. Interaction between sleep and the immune response in Drosophila: a role for the NFkappaB relish. Sleep. 2007;30(4):389–400. doi:10.1093/sleep/30.4.389
15. Wang Z, Hang S, Purdy AE, Watnick PI. Mutations in the IMD pathway and mustard counter Vibrio cholerae suppression of intestinal stem cell division in Drosophila. Mbio. 2013;4:e313–e337. doi:10.1128/mBio.00337-13
16. Kosakamoto H, Yamauchi T, Akuzawa-Tokita Y, et al. Local Necrotic Cells Trigger Systemic Immune Activation via Gut Microbiome Dysbiosis in Drosophila. Cell Rep. 2020;32(3):107938. doi:10.1016/j.celrep.2020.107938
17. Cai Q, Ji S, Li M, et al. Theaflavin-regulated Imd condensates control Drosophila intestinal homeostasis and aging. iScience. 2021;24(3):102150. doi:10.1016/j.isci.2021.102150
18. Zhang R, Du J, Zhao X, Wei L, Zhao Z. Regulation of circadian behavioural output via clock-responsive miR −276b. Insect mol Biol. 2021;30(1):81–89. doi:10.1111/imb.12679
19. Hendricks JC. Shaking up sleep research. Nat Neurosci. 2005;8(6):703–705. doi:10.1038/nn0605-703
20. Hou Y, Zou L, Li Q, et al. Supramolecular assemblies based on natural small molecules: union would be effective. Mater Today Bio. 2022;15:100327. doi:10.1016/j.mtbio.2022.100327
21. Zu M, Ma Y, Cannup B, et al. Oral delivery of natural active small molecules by polymeric nanoparticles for the treatment of inflammatory bowel diseases. Adv Drug Deliv Rev. 2021;176:113887. doi:10.1016/j.addr.2021.113887
22. Narayanankutty A. PI3K/Akt/mTOR Pathway as a Therapeutic Target for Colorectal Cancer: a Review of Preclinical and Clinical Evidence. Curr Drug Targets. 2019;20(12):1217–1226. doi:10.2174/1389450120666190618123846
23. Pavlikova N. Caffeic Acid and Diseases-Mechanisms of Action. Int J mol Sci. 2022;24(1):588. doi:10.3390/ijms24010588
24. Wen X, Wan F, Wu Y, et al. Caffeic acid supplementation ameliorates intestinal injury by modulating intestinal microbiota in LPS-challenged piglets. Food Funct. 2023;14(16):7705–7717. doi:10.1039/D3FO02286B
25. Esmeeta A, Adhikary S, Dharshnaa V, et al. Plant-derived bioactive compounds in colon cancer treatment: an updated review. Biomed Pharmacother. 2022;153:113384.
26. Zielinska D, Zielinski H, Laparra-Llopis JM, Szawara-Nowak D, Honke J, Giménez-Bastida JA. Caffeic Acid Modulates Processes Associated with Intestinal Inflammation. Nutrients. 2021;13(2):554. doi:10.3390/nu13020554
27. Zhang Z, Wu X, Cao S, et al. Caffeic acid ameliorates colitis in association with increased Akkermansia population in the gut microbiota of mice. Oncotarget. 2016;7(22):31790–31799. doi:10.18632/oncotarget.9306
28. Jaganathan SK. Growth inhibition by caffeic acid, one of the phenolic constituents of honey, in HCT 15 colon cancer cells. Sci World J. 2012;2012:372345. doi:10.1100/2012/372345
29. Kang NJ, Lee KW, Kim BH, et al. Coffee phenolic phytochemicals suppress colon cancer metastasis by targeting MEK and TOPK. Carcinogenesis. 2011;32:921–928.
30. Li X, Yang S, Wang S, et al. Regulation and mechanism of Astragalus polysaccharide on ameliorating aging in Drosophila melanogaster. Int J Biol Macromol. 2023;234:123632. doi:10.1016/j.ijbiomac.2023.123632
31. He J, Han S, Wang Y, et al. Irinotecan cause the side effects on development and adult physiology, and induces intestinal damage via innate immune response and oxidative damage in Drosophila. Biomed Pharmacother. 2023;169:115906.
32. Li H, Qi Y, Jasper H. Preventing Age-Related Decline of Gut Compartmentalization Limits Microbiota Dysbiosis and Extends Lifespan. Cell Host Microbe. 2016;19(2):240–253. doi:10.1016/j.chom.2016.01.008
33. Han S, Xiu M, Li S, et al. Exposure to cytarabine causes side effects on adult development and physiology and induces intestinal damage via apoptosis in Drosophila. Biomed Pharmacother. 2023;159:114265. doi:10.1016/j.biopha.2023.114265
34. Abu F, Ohlstein B. Monitoring Gut Acidification in the Adult Drosophila Intestine. J Vis Exp. 2021;11(176):e63141. doi:10.3791/63141.
35. Staats S, Luersen K, Wagner AE, Rimbach G. Drosophila melanogaster as a Versatile Model Organism in Food and Nutrition Research. J Agric Food Chem. 2018;66(15):3737–3753. doi:10.1021/acs.jafc.7b05900
36. Wang YY, Ma WW, Peng IF. Screening of sleep assisting drug candidates with a Drosophila model. PLoS One. 2020;15:e236318.
37. Li Y, Zhou X, Cheng C, et al. Gut AstA mediates sleep deprivation-induced energy wasting in Drosophila. Cell Discov. 2023;9(1):49. doi:10.1038/s41421-023-00541-3
38. Zhou J, He L, Liu M, et al. Sleep loss impairs intestinal stem cell function and gut homeostasis through the modulation of the GABA signalling pathway in Drosophila. Cell Prolif. 2023;56(9):e13437. doi:10.1111/cpr.13437
39. Gao T, Wang Z, Cao J, Dong Y, Chen Y. Melatonin alleviates oxidative stress in sleep deprived mice: involvement of small intestinal mucosa injury. Int Immunopharmacol. 2020;78:106041. doi:10.1016/j.intimp.2019.106041
40. Su H, Zhang C, Zou X, et al. Jiao-tai-wan inhibits inflammation of the gut-brain-axis and attenuates cognitive impairment in insomnic rats. J Ethnopharmacol. 2020;250:112478. doi:10.1016/j.jep.2019.112478
41. Zhai Z, Huang X, Yin Y. Beyond immunity: the Imd pathway as a coordinator of host defense, organismal physiology and behavior. Dev Comp Immunol. 2018;83:51–59. doi:10.1016/j.dci.2017.11.008
42. Zaidman-Remy A, Poidevin M, Hervé M, et al. Drosophila immunity: analysis of PGRP-SB1 expression, enzymatic activity and function. PLoS One. 2011;6(2):e17231. doi:10.1371/journal.pone.0017231
43. Mellroth P, Steiner H. PGRP-SB1: an N-acetylmuramoyl L-alanine amidase with antibacterial activity. Biochem Biophys Res Commun. 2006;350(4):994–999. doi:10.1016/j.bbrc.2006.09.139
44. Landis GN, Abdueva D, Skvortsov D, et al. Similar gene expression patterns characterize aging and oxidative stress in Drosophila melanogaster. Proc Natl Acad Sci U S A. 2004;101(20):7663–7668. doi:10.1073/pnas.0307605101
45. Mirzaei S, Gholami MH, Zabolian A, et al. Caffeic acid and its derivatives as potential modulators of oncogenic molecular pathways: new hope in the fight against cancer. Pharmacol Res. 2021;171:105759. doi:10.1016/j.phrs.2021.105759
46. Hao R, Ge J, Ren Y, et al. Caffeic acid phenethyl ester mitigates cadmium-induced hepatotoxicity in mice: role of miR-182-5p/TLR4 axis. Ecotoxicol Environ Saf. 2021;207:111578. doi:10.1016/j.ecoenv.2020.111578
47. Kolgazi M, Cilingir S, Yilmaz O, et al. Caffeic acid attenuates gastric mucosal damage induced by ethanol in rats via nitric oxide modulation. Chem Biol Interact. 2021;334:109351. doi:10.1016/j.cbi.2020.109351
48. Xiang C, Liu M, Lu Q, et al. Blockade of TLRs-triggered macrophage activation by caffeic acid exerted protective effects on experimental ulcerative colitis. Cell Immunol. 2021;365:104364. doi:10.1016/j.cellimm.2021.104364
49. Wan F, Zhong R, Wang M, et al. Caffeic Acid Supplement Alleviates Colonic Inflammation and Oxidative Stress Potentially Through Improved Gut Microbiota Community in Mice. Front Microbiol. 2021;12:784211. doi:10.3389/fmicb.2021.784211
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Caffeic Acid Protects Against Ulcerative Colitis via Inhibiting Mitochondrial Apoptosis and Immune Overactivation in Drosophila

Xiu M, Li B, He L, Shi Y, Zhang Y, Zhou S, Liu Y, Wang N, He J
Drug Design, Development and Therapy 2025, 19:2157-2172
Published Date: 22 March 2025