Back to Journals » Journal of Pain Research » Volume 18
Borneol’s Pre-Clinical Analgesic Efficacy: Mediated by Receptor and Immune Mechanisms
Received 16 April 2025
Accepted for publication 5 August 2025
Published 13 August 2025 Volume 2025:18 Pages 4085—4104
DOI https://doi.org/10.2147/JPR.S533901
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor E Alfonso Romero-Sandoval
Yichen Zhou,1,* Xinyue Mei,1,* Changming Wang2
1School of Elderly Care Services and Management, Nanjing University of Chinese Medicine, Nanjing, Jiangsu, 210023, People’s Republic of China; 2School of Medicine, Nanjing University of Chinese Medicine, Nanjing, Jiangsu, 210023, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Changming Wang, Email [email protected]
Abstract: Chronic pain affects over 10% of the global population, yet current analgesics face limitations such as addiction and systemic toxicity. Borneol, a traditional Chinese medicine (TCM) with over a millennium history, has emerged as a promising alternative due to its multi-target mechanisms. This review synthesizes preclinical evidence demonstrating borneol’s dual analgesic pathways: (1) receptor-mediated actions (eg, TRPM8 activation, GABA agonism) and (2) immune modulation (eg, NF-κB suppression, ROS inhibition). Notably, comparative analysis with menthol reveals borneol’s superior TRPM8 specificity and lack of opioid-related side effects. By elucidating these mechanisms, we advocate for borneol’s integration into modern pain management strategies, highlighting its potential as a safe, multi-modal therapeutic.
Keywords: Borneol, analgesia, TRPM8, immune modulation, neuropathic pain, Traditional Chinese Medicine
Introduction
Over 10% of individuals globally experience chronic pain, significantly diminishing their quality of life and imposing substantial financial burdens.1 Presently, the majority of conventional analgesics are systemic and include non-steroidal anti-inflammatory drugs (NSAIDs) such as aspirin and ibuprofen, narcotic analgesics like morphine and pethidine, antispasmodic analgesics such as atropine and anisodamine, and central analgesics like tramadol and bucinnazine.2 The first two categories of analgesics, while potent, are highly addictive and are therefore subject to stringent regulation, being primarily approved for the management of severe pain associated with advanced cancer, major trauma, and surgical procedures.3 Antispasmodic analgesics alleviate visceral smooth muscle spastic pain, such as gastrointestinal spasms, biliary colic, and renal colic, by inhibiting M choline receptors on smooth muscle. However, they are associated with side effects including dry mouth, blurred vision, and increased heart rate.4 NSAIDs primarily mitigate inflammatory pain by inhibiting cyclooxygenase (COX) activity and reducing prostaglandin synthesis, although they can also produce adverse effects such liver and renal malfunction and gastrointestinal distress.5 Due to concerns regarding the addictive, abusive, and harmful systemic effects of analgesics, there is an ongoing search for safer and more effective medications and pain management approaches.
Traditional Chinese medicine (TCM) has been used in East Asia for millennia to address various pains. Among these, Borneol stands out with more than a millennium application history in analgesia, yet research on its analgesic mechanisms and effects remained scarce in the early stages due to the complexity of mechanisms and differences in theoretical systems, lack of standardization and normalization, cultural and cognitive barriers only being supplemented to a certain extent up to now.
The chemical composition of Borneol is identified as C10H18O, comprising four enantiomers: (+)-borneol, (−)-borneol, (+)-isoborneol, and (−)-isoborneol.6 Studies on the analgesic effects and mechanisms of (+)-borneol are relatively abundant, while research on other isomeric forms remains scarce. It should be noted that this article provides clear annotations whenever the included studies specifically describe borneol isomers; when the term “borneol” is used without qualification, it indicates that the original literature did not explicitly identify the isomeric form.
According to historical records such as Hui Yue Yi Jing (1818) and Ben Cao Gang Mu (1596), as well as modern standards outlined in the Chinese Pharmacopoeia (Chinese Pharmacopoeia Commission, 2015), borneol demonstrates therapeutic efficacy in topical applications. Specifically, it has been documented to reduce swelling, alleviate pain, and promote healing in cases of trauma (eg, falls, injuries), skin sores, and ulcers. Additionally, it can be administered orally to treat consciousness disorders, such as stroke-induced coma, and to relieve throat and gingival swelling and pain caused by the accumulation of heat toxins in the body.7 Borneol is frequently utilized as an adjunct component in traditional Chinese medicine for the treatment of cardiovascular and cerebrovascular diseases, primarily by facilitating the transmission of other medicinal constituents.8–10 Additionally, borneol exhibits anti-inflammatory, neuroprotective, and antibacterial properties.11–13 It has received approval from the FDA for use as a flavoring agent and excipient in food and toiletries.14 Given its various beneficial properties and uses, it is of great significance to explore and optimize its utilization in modern medicine including depth research into its molecular mechanisms of action, rigorous clinical trials to precisely determine its efficacy and safety profiles.
The diverse pharmacological effects of borneol appear to be mediated through its direct or indirect interactions with various receptors and the immune system. Current research indicates that borneol can exert analgesic effects by modulating several ion channels, including TRPM8, TRPV3, GABAA, 5-HT receptors, and acetylcholine receptors.14–16 Furthermore, studies have demonstrated that borneol can mitigate inflammatory pain by regulating immune-active substances such as NF-kB and nitric oxide synthase. This complex and distinctive role of borneol positions it as a promising candidate for analgesic drug development.
This article aims to provide a comprehensive review of borneol’s analgesic properties, focusing on preclinical findings and the molecular mechanisms underlying its effects. By exploring its potential as a modern analgesic drug, this review seeks to bridge the gap between TCM and contemporary drug development, offering insights into safer and more effective pain management strategies.
Clinical and Preclinical Tests of Borneol Analgesia
Pain is defined as an unpleasant sensory and emotional experience associated with actual or potential tissue damage, or described in terms of such damage.17 Chronic pain can be classified into three main categories based on etiology and clinical manifestations: nociceptive pain, neuropathic pain, and central sensitization pain.1,18 However, certain types of pain, such as cancer-related pain and spinal pain, exhibit mixed pain phenotypes. To date, the analgesic efficacy of borneol has been demonstrated in clinical studies in different modes of diseases and in various rodent models for acute pain, nociceptive pain associated with chronic pain, and neuropathic pain.
Clinical Applications of Borneol in Pain Management
A meta-analysis has demonstrated that the Compound Danshen Dripping Pill (CDDP), which includes borneol as a primary active ingredient, in conjunction with percutaneous coronary intervention (PCI), significantly reduces the incidence of recurrent angina (P = 0.0001) and alleviates angina symptoms.19 Furthermore, a randomized controlled clinical trial indicated that Kuanxiong Aerosol (KXA), which contains borneol, exhibited a significantly higher short-term efficacy in pain relief (72.2% pain relief rate within 30 minutes) compared to nitroglycerin aerosol for prehospital chest pain, while maintaining comparable safety.20 Additionally, another randomized controlled trial (RCT) revealed that the topical application of (+)-borneol (25%) can substantially alleviate postoperative pain in patients following spinal surgery.14 Nonetheless, current clinical trials involving borneol present considerable opportunities for enhancement, particularly regarding sample size, combination therapies, administration methods, dosage, frequency, disease models, and the specific stereochemical configuration of borneol utilized.
Preclinical Trials
Acute Pain
Capsaicin Model
Mice that received a local application of borneol at concentrations of 2% and 25% prior to capsaicin injection resulted in a significant reduction in paw licking and lifting time within five minutes post-injection.14,21
Formalin Model
Topical administration of (+)-borneol at concentrations of 1.5%, 2%, 4.5%, and 15% before formalin injection into the plantar surface significantly reduced the duration of paw licking and lifting within the initial five minutes and between fifteen to sixty minutes post-injection. The analgesic effect was dose-dependent.14,21 Additionally, the time spent licking the paws was consistently reduced by intraperitoneal injection of 5 mg/kg, 25 mg/kg, and 50 mg/kg borneol prior to formalin injection.22
AITC Model
The topical administration of (+)-borneol five minutes prior to AITC plantar injection significantly reduced the duration of paw licking in mice compared to the control group.21
Acetic Acid Model
In the context of intraperitoneal acetic acid injection, mice exhibited a dose-dependent reduction in convulsions when pre-treated with intraperitoneal injections of (+)-borneol at doses of 5 mg/kg, 25 mg/kg, or 50 mg/kg.22
Unmodeled Mouse
Mice demonstrated a significantly increased latency time compared to the control group when administered 50 mg/kg of (+)-borneol intraperitoneally prior to the hot plate test.22 Additionally, local application of 2% (+)-borneol prior to the hot plate test significantly extended latency time.21
The above researches highlight borneol as a promising candidate for acute pain management, with mechanisms likely involving modulation of pain pathways and anti-inflammatory effects.
Chronic Pain
Nociceptive Pain
Nociceptive pain arises from the activation of peripheral nociceptors by noxious stimuli, which may include mechanical, thermal, and chemical stimuli. Upon tissue damage, nociceptors are activated, generating nerve impulses that are transmitted to the spinal cord via nerve fibers and subsequently relayed to the brain, resulting in the perception of pain. In the context of inflammation, damaged tissues release chemical mediators such as prostaglandins and histamine, which increase nociceptor sensitivity, lower the pain threshold, and enhance pain signal transduction.23 The administration of 15% (+)-borneol significantly elevated the 50% paw withdrawal thresholds and nociceptive response latency in mice subjected to the CFA plantar injection model of inflammatory mechanical and thermal hyperalgesia.14 Additionally, oral administration of (+)-borneol at doses of 125 mg/kg, 250 mg/kg, and 500 mg/kg significantly increased the 50% paw withdrawal thresholds in a dose-dependent manner (P < 0.001), as did intrathecal injections of 15 µg, 30 µg, and 60 µg of borneol.24 In the inflammatory pain model induced by subcutaneous injection of P. acnes in the ear, the combination of a 40% borneol local ear application and photodynamic therapy (PDT) significantly enhanced the 50% paw withdrawal thresholds of the ipsilateral hind paw of the affected ear.25
Neuropathic Pain
The International Association for the Study of Pain (IASP) defines neuropathic pain as pain arising from damage or disease affecting the somatosensory nervous system.26 Neuropathic pain are primarily attributed to alterations in the structure and function of neurons following nerve injury, which lead to abnormal electrical activity and neurotransmitter release. The injured nerve exhibits ectopic discharges, resulting in spontaneous pain; concurrently, the remodeling of the nerve conduction pathway contributes to the aberrant amplification of pain signals.27,28 In the mouse spinal nerve ligation (SNL) model, intrathecal administration of (+)-borneol at doses of 15 µg, 30 µg, and 60 µg significantly increased the 50% paw withdrawal thresholds in a dose-dependent manner. Similarly, oral administration of (+)-borneol at doses of 125 mg/kg, 250 mg/kg, and 500 mg/kg also significantly elevated the 50% paw withdrawal thresholds.24
Overall, borneol can be administered through various routes, including local application, oral administration, intraperitoneal injection, and intrathecal injection. These methods have been evaluated in models of acute pain, such as capsaicin, formalin, AITC plantar injection, and acetic acid intraperitoneal injection, as well as in chronic pain models, including CFA-induced or acne pseudomonas infection-induced pain (Figure 1). These preclinical findings suggest that borneol may act through multiple mechanisms, such as modulating pain receptors, reducing inflammation, and normalizing neuronal activity.
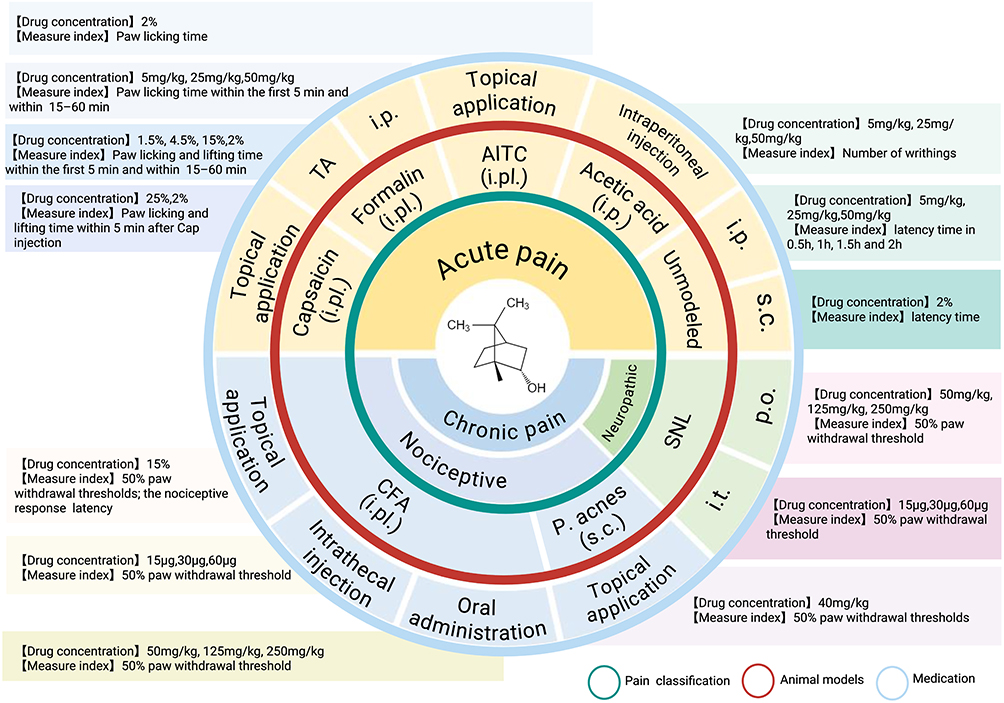 |
Figure 1 Analgesic efficacy of borneol across various administration routes and pain models. Borneol can be administered through various routes, including topical application (with concentrations of 1.5%, 2%, 4.5%, and 15%, and dosages of 25 mg/kg and 40 mg/kg), oral administration (at dosages of 125 mg/kg, 250 mg/kg, and 500 mg/kg), intraperitoneal injection (at dosages of 5 mg/kg, 25 mg/kg, and 50 mg/kg), and intrathecal injection (at dosages of 15 μg, 30 μg, and 60 μg), among others. The figure illustrates the application of borneol across different pain models, including acute pain induced by plantar injection of capsaicin, formalin, AITC, intraperitoneal injection of acetic acid, and the hot-plate test, as well as chronic pain induced by CFA, Pseudomonas acnes infection, and spinal nerve ligation. The data indicate that borneol exhibits significant analgesic effects in these pain models. Created in BioRender. Z, (Y) (2025) https://BioRender.com/m33h644. Abbreviations: TA, topical application; i.p., intraperitoneal injection; s.c., subcutaneous injection; i.pl., intraplantar injection; i.t., intrathecal injection; P. acnes, Propionibacterium acnes; SNL, spinal nerve ligation; p.o., oral administration. |
Receptors Related Analgesic Mechanism of Borneol
Borneol gives us a cooling sensation and makes us feel extremely happy when in pain. Preclinical test of borneol analgesia indicates that borneol may directly or un-directly functioned on those pain related receptors. The Transient Receptor Potential (TRP) family is the most concerned receptors for hot and cold stimuli. TRP family comprises non-selective cation channels that are ubiquitously present in various biological cell membranes.29 These channels serve as sensors for a wide range of endogenous and exogenous stimuli. They are capable of initially detecting harmful signals that induce pain and subsequently converting these signals into neural electrical activity by mediating intracellular and extracellular ion transport. This neural activity is then transmitted to the cerebral cortex via the spinal cord, subthalamic region, and brainstem, ultimately resulting in the sensation of pain. The TRP channels associated with pain perception include several subfamilies, such as ankyrin (A), vanillin (V), melanostatin (M), and classical (C). Each subfamily’s ion channels play distinct roles in nociceptive, inflammatory, or neuropathic pain.30 Currently, borneol is recognized for its regulatory effects on TRPM8, TRPA1, and TRPV3 channels within the TRP family.
Trpm8
TRPM8 is a non-selective calcium channel expressed in a limited number of nociceptors located in the dorsal root ganglion (DRG) and trigeminal ganglion (TG).31 These channels can be activated by low temperatures (8–28°C),32 menthol, icilin,33 and voltage changes,34 which can not only provide benign thermal stimulation that initiates a cooling sensation and exerts analgesic effects, but also to transmit nociceptive signals that can induce hyperalgesia.35 Research indicates that (+)-borneol modulates analgesia through its interaction with the TRPM8 channel. With a half-maximal effective concentration (EC50) of 65 μM, borneol activates human TRPM8 (hTRPM8) in HEK293 cells expressing TRPM8, as demonstrated by patch-clamp recordings.14 By activating TRPM8 and increasing the opening frequency of TRPM8 channels, (+)-borneol facilitates the influx of calcium ions into primary sensory neurons, thereby promoting the propagation of action potentials.36 Furthermore, studies have confirmed that the analgesic effects of (+)-borneol are evident at both behavioral and cytological levels. At the cytological level, it has been shown that 100 μM borneol enhances Ca2+ influx in HEK293 cells expressing TRPM8 and in the DRG of wild-type mice. However, no calcium influx was observed in HEK293 cells lacking TRPM8 expression or in the DRG of TRPM8-deficient mice.14,36 Additionally, the calcium influx induced by (+)-borneol can be inhibited by the specific TRPM8 inhibitor AMTB (N-(4-tert-butylphenyl)-4-(3-chloropyridin-2-yl)piperazine-1-carboxamide) and the less selective TRPM8 inhibitor BCTC (N-(4-butylphenyl)-4-(3-chloropyridin-2-yl)piperazine-1-carboxamide).14,36
From a behavioral perspective, in the capsaicin, formalin, and CFA models, the analgesic effect of local (+)-borneol application was almost negligible in TRPM8-deficient mice compared to wild-type (WT) mice.14 Furthermore, the analgesic effect induced by (+)-borneol was significantly counteracted by administering AMTB into the plantar region prior to the local application of (+)-borneol or ethanol.14 Borneol modulates analgesia by activating TRPM8 channels in sensory neurons, with its effects confirmed cytologically and behaviorally in TRPM8-expressing systems and inhibited by TRPM8 antagonists.
The analgesic efficacy of topically applied (+)-borneol was notably diminished following intrathecal injection of LY341495, a selective antagonist of group II metabotropic glutamate receptors (mGluRs).14 This observation suggests that borneol may share similarities with other TRPM8 agonist analgesic pathways, such as icillin, by competitively inhibiting postsynaptic membrane glutamate receptor activation-mediated pain transmission. This inhibition occurs through the reduction of glutamate levels in the synaptic cleft, achieved by upregulating the expression of metabolic glutamate receptors in the presynaptic spinal cord group II/III.37
Overall, borneol’s analgesic action is mediated by TRPM8 activation and subsequent calcium influx, with additional involvement of glutamate receptor modulation.
Trpa1
TRPA1 is a non-selective cation channel belonging to the TRP family. Unlike TRPM8, which is crucial for both external and endogenous functions, TRPA1 is exclusively expressed in the nociceptive neurons of DRG and TG.38 Bradykinin and other inflammatory mediators,39 formalin, cinnamon aldehyde,40,41 and other irritating substances can all activate it, which in turn causes pain. Consequently, TRPA1 inhibitors might be a viable analgesic target. Borneol has been shown to inhibit TRPA1, mediating its analgesic effects. This inhibitory action on TRPA1 is potentially attributable to interactions between the hydroxyl group on the six-membered ring of borneol and the S873, T874, and Y812 residues of human TRPA1 (hTRPA1).42 In patch-clamp recordings, borneol demonstrated a dose-dependent inhibition of AITC (20 μM)-induced hTRPA1 currents, with an IC50 of 0.20 ± 0.06 mM.42 Behavioral studies have corroborated these findings, in hot plate assays, intrathecal administration of (+)-borneol increased the pain threshold in mice with oxaliplatin-induced hyperalgesia in a dose-dependent manner. Furthermore, (+)-borneol was able to counteract the effects of the TRPA1 agonist cinnamaldehyde in mechanical and cold plate tests without impairing motor function in mice.43 However, some studies have indicated that the analgesic effect of locally applied (+)-borneol is similar in TRPA1-deficient and wild-type mice in capsaicin and formalin models, suggesting that while borneol may interact with TRPA1, it may not significantly contribute to the analgesic effects of local application.14 This suggests that borneol may act on TRPA1 but contributes less to the analgesic effect of local borneol. Borneol’s analgesic effects are likely mediated through multiple pathways, including its well-documented activation of TRPM8 and glutamate signaling. While TRPA1 inhibition contributes to borneol’s analgesic properties in specific conditions, its role may be secondary to other mechanisms, such as TRPM8 activation, in mediating the compound’s overall analgesic efficacy. Further research is needed to fully elucidate the relative contributions of TRPA1 and other pathways to borneol’s multifaceted analgesic effects.
Trpv3
The thermosensitive ion channel TRPV3 is predominantly expressed in the brain, spinal cord, DRG, and keratinocytes.44,45 TRPV3 is activated not only by monoterpene compounds such as camphor, carvacrol, and thymol but also at temperatures ranging from approximately 34 to 38 °C.46 Although the precise physiological role of TRPV3 remains unclear, research has demonstrated that its activation can lead to discomfort. In murine models with keratinocytes overexpressing TRPV3, there is an increased production of prostaglandin E2 (PGE2), resulting in sensations of heat-induced damage and hyperalgesia.47 The administration of TRPV3-specific inhibitors, including resolvin D1 (RvD1) and isopentenyl pyrophosphate, has been shown to induce peripheral antinociception.48,49 Patch clamp studies indicate that TRPV3 can be activated by 2 mM (+)-borneol during 2-APB sensitization, with an EC50 of 3.45 ± 0.13 mM.50 However, it remains unclear whether borneol’s activation of TRPV3 contributes to its analgesic effects or whether it might indirectly modulate pain pathways by influencing other channels, such as TRPV1 and TRPA1. Overall, borneol activation with TRPV3 represents a complex and underexplored component of pain signaling, with potential implications for understanding and treating pain through modulation of thermosensitive ion channels.
Gabaa
In the central nervous system, gamma-aminobutyric acid (GABA) serves as a prevalent inhibitory neurotransmitter. Approximately 40% of neurons in the mammalian central nervous system contain this neurotransmitter.51 Its receptors are classified into three subtypes: GABAA, GABAB, and GABAC. Among these, GABAA is the most prevalent and is primarily located on the postsynaptic membrane within the central nervous system. Upon activation, this ligand-gated ion channel facilitates the influx of extracellular chloride ions, resulting in membrane hyperpolarization and subsequent neuronal inhibition. This process plays a crucial role in sensory and motor conduction pathways.24 The administration of GABA receptor antagonists via intrathecal injection, which inhibits GABAergic neurotransmission in the spinal cord, can lead to allergic reactions to otherwise harmless stimuli.52,53 It has been proposed that drugs activating the GABAA receptor (GABAAR), such as benzodiazepines or GABAAR agonists, serve as effective analgesics in various models of inflammatory and neuropathic pain.54,55
Research has demonstrated that (+)-borneol functions as a full agonist of GABAA receptors. Patch-clamp experiments have shown its dose-dependent enhancement of chloride currents in human recombinant GABAA receptors expressed in Xenopus laevis oocytes, with effects observed at low (EC5-14), moderate (EC15-24), and saturating (EC100) concentrations of GABA.51 Furthermore, (+)-borneol has been found to increase the expression of brain-derived neurotrophic factor (BDNF) by upregulating GABAA expression. In excitatory synapses activated by NMDARs, BDNF can facilitate the binding of PSD-95 to TrkB, potentially alleviating neuropathic pain associated with the binding of PSD95 to nNOS in the presence of ZL006.56 Additionally, studies have shown a progressive increase in brain GABA levels in mice, ranging from 0.167 to 5 hours post-oral administration of (+)-borneol, indicating that borneol may exert analgesic effects through the modulation of brain GABA levels.57
Although studies have indicated that the application of strychnine (a glycine antagonist) and picrotoxin (a GABAA antagonist) in the substantia gelatinosa (SG) neurons of the caudal trigeminal subnucleus (Vc) in mice nearly abolished the inward current induced by -(-)borneol,58 it is essential to consider the differential distribution of TRPM8 and GABAA receptors. This raises questions regarding whether GABAA is the primary target of borneol. The evidence predominantly suggests this at the advanced central level, as TRPM8 is infrequently present there. It is plausible that GABAA is indeed the principal target of borneol.
At the behavioral level, research has demonstrated that in the SNL and CFA models, the analgesic effect of orally administered borneol can be almost entirely negated by the intrathecal injection of bicuculline (a selective GABAA antagonist, 0.1 µg) 30 minutes following oral administration of (+)-borneol.24 Conversely, intrathecal administration of bicuculline had minimal effect on the analgesic efficacy of locally administered (+)-borneol in the capsaicin model.14
Overall, borneol may act on GABAA receptors, showing dose- and stereo-selectivity, and affects pain models with mixed evidence on its main target.
5-Ht
Serotonin (5-HT), a monoamine extensively distributed within the nervous system, is located in 5-hydroxytryptaminergic neurons in the brain and spinal cord, where it acts as a neurotransmitter to regulate mood, sleep, and pain. Upon inflammation or tissue damage, 5-HT is released from mast cells and platelets in the surrounding system, potentially alleviating or exacerbating pain.59 The role of 5-HT in influencing the pain process has been recognized for a considerable time.60,61 Research conducted in both fundamental and clinical contexts has established the critical role of serotonin in pain modulation. For the management of various pain syndromes, a range of 5-HT receptor subtype agonists, antagonists, and 5-HT reuptake inhibitors have been utilized.61,62
High-performance liquid chromatography-electrochemical detection (HPLC-ECD) has demonstrated a significant increase in 5-HT levels in the hypothalamus of rats 45 minutes following the administration of high-dose oral borneol (P < 0.05).63 Nevertheless, it remains unclear whether borneol facilitates the action of 5-HT on receptors such as 5-HT1B and 5-HT1D in spinal dorsal horn neurons, thereby inhibiting pain signal transmission to produce analgesic effects.
Acetylcholine Receptors
Nicotinic acetylcholine receptors (nAChRs), members of the cysteine-loop ligand-gated ion channel superfamily, respond to endogenous acetylcholine (ACh) and other cholinergic ligands. These receptors are extensively expressed in both the central nervous system (CNS) and peripheral nervous system (PNS), where they are located at presynaptic and postsynaptic sites and participate in various functions, including learning and memory, arousal, reward, motor control, and analgesia.64 For instance, α7 nicotinic acetylcholine receptors (nAChRs) have increasingly been recognized as promising targets for the development of anti-inflammatory and analgesic agents.65,66 Borneol has been identified as a specific noncompetitive inhibitor of nAChR-mediated effects, which may underlie its analgesic properties. According to calcium imaging studies conducted by the Tae-Ju Park team,67 borneol exhibited a half-maximal inhibitory concentration (IC50) of 56±9 μM, effectively preventing the calcium influx induced by the nAChR agonist 1,1-dimethyl-4-phenylpiperazinium iodide (DMPP). Borneol is a specific noncompetitive inhibitor of nAChR-mediated effects, possibly related to its analgesic properties and blocked calcium influx induced by the nAChR agonist DMPP.
Consequently, borneol may exert analgesic effects by inhibiting peripheral receptors and modulating central excitatory neural transmission. In the peripheral nervous system, borneol inhibits pain signal transduction by activating TRPM8, up-regulating type II/III glutamate receptor expression, and reducing synaptic glutamate levels. Additionally, it diminishes pain sensitivity by down-regulating TRPA1 expression. In the central nervous system, borneol acts as a partial or complete agonist of GABA receptors (GABAAR), can encourage the inflow of Cl ions, enhancing GABA expression and increasing GABA concentrations in the brain. By elevating the levels of 5-HT in the brain, there is an induction of potassium ion efflux, leading to membrane hyperpolarization. This process can also inhibit the influx of sodium, potassium, and other cations by preventing the activation of acetylcholine receptors, collectively impeding the conduction of action potentials responsible for pain sensation. This mechanism provides a theoretical foundation for its analgesic effects observed in animal models of acute pain and chronic hyperalgesia (Figure 2).
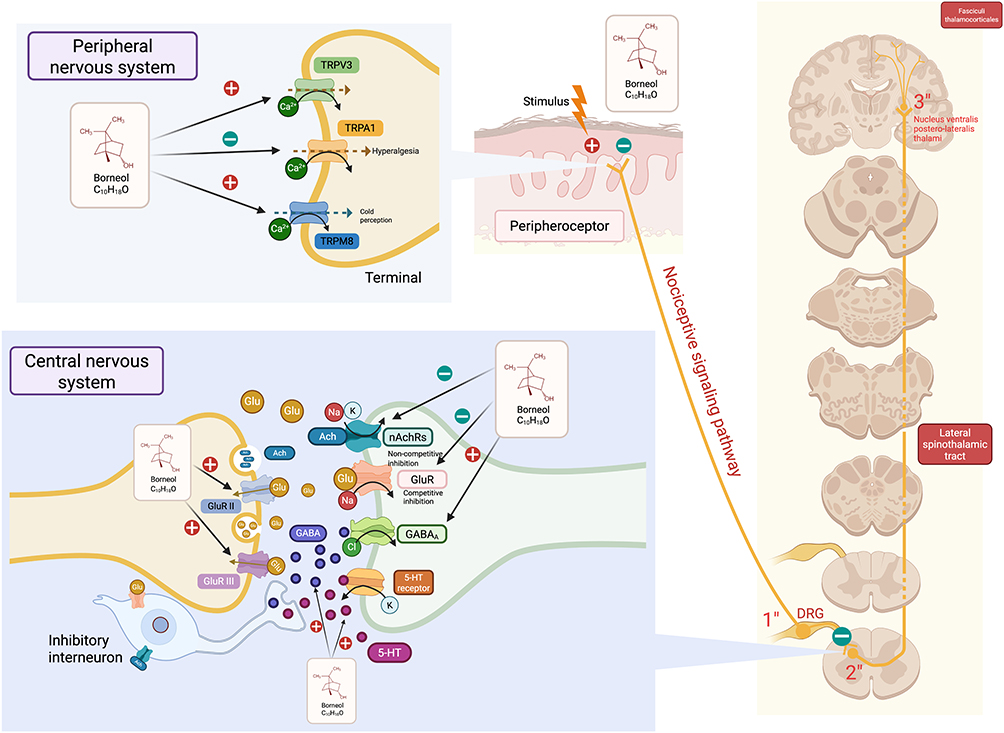 |
Figure 2 Receptors related analgesic mechanism of borneol. In the peripheral nervous system, noxious stimuli activate peripheral nociceptors, generating nerve impulses that are transmitted along nerve fibers to the spinal cord and then uploaded to the brain to produce the sensation of pain. Borneol (C10H18O) can exert an analgesic effect by activating TRPM8, inhibiting TRPA1, and activating TRPV3. In the central nervous system, borneol can play an analgesic role by up - regulating the expression of presynaptic metabotropic glutamate receptors II/III, competitively inhibiting the pain conduction mediated by glutamate in the synaptic cleft acting on postsynaptic glutamate receptors. It can also exert an analgesic effect by activating GABAA, non - competitively inhibiting nAChRs, and increasing the levels of GABA and 5 - HT in the central nervous system. Created in BioRender. Z, (Y) (2025) https://BioRender.com/rj3jbp9. |
The analgesic efficacy of borneol is attributed to the synergistic action of receptor and immune mechanisms, rather than isolated effects. Subsequently, the immune-related analgesic mechanisms of borneol are examined.
Immune-Related Analgesic Mechanism of Borneol
Pain is a primary characteristic of inflammation. From an immunological perspective, borneol can mitigate the onset and progression of inflammation by inhibiting the production of inflammatory mediators, reducing nitric oxide synthesis, curtailing oxidative stress, limiting the migration and infiltration of inflammatory cells, and promoting the resolution of inflammation (Figure 3). Consequently, borneol exerts an analgesic effect, which has been substantiated in various rodent models.
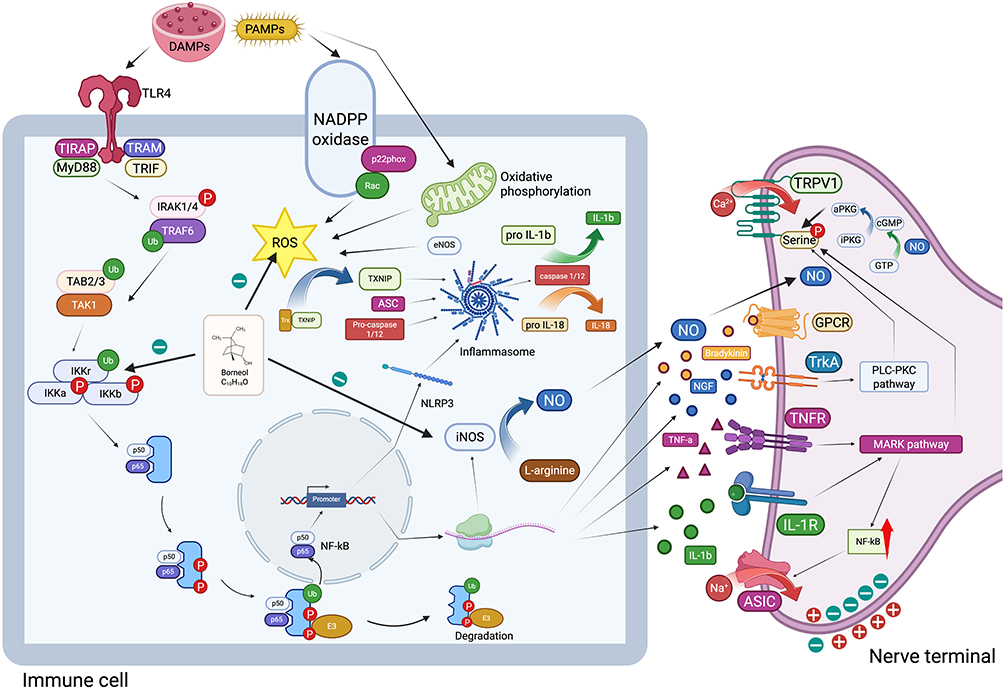 |
Figure 3 Immune-related analgesic mechanism of borneol. Borneol can effectively reduce the transcription and expression of genes related to inflammatory factors such as bradykinin, IL - 1β, TNF - α, and NGF by down - regulating the expression of NF - κB - related proteins such as IKKα/β and IκBα. It inhibits the activation of PLC - PKC and MARK signaling pathways mediated by TrkA, TNFR, and IL - 1R, and then suppresses the phosphorylation of TRPV1, as well as the activation of GPCR and ASIC, thus exerting an analgesic effect. Borneol can inhibit the production of NO by suppressing the activity of nitric oxide synthase, and then inhibit the phosphorylation of TRPV1 by suppressing the phosphorylation of PKG. Borneol can also inhibit the production of inflammasomes and the inflammatory mediators such as IL - 1β and IL - 18 mediated by them by inhibiting the synthesis of ROS. Created in BioRender. Z, (Y) (2025) https://BioRender.com/muzx5cx. |
Inhibition of the Production of Inflammatory Factors
In an infectious inflammatory environment, pathogen-associated molecular patterns (PAMPs) of bacteria, along with damage-associated molecular patterns (DAMPs) in a sterile context are recognized by the TLR4 receptor of innate immune cells.68 The recognition events trigger the release of the adaptor protein MyD88, activation of IRAK1 and IRAK4 kinases, auto-ubiquitination of TRAF6, activation of TAK1, and phosphorylation of IKKβ.69 The phosphorylated IKK complex further phosphorylates IκBα, leading to its ubiquitination and subsequent degradation by the proteasome. This process facilitates the release and nuclear translocation of NF-κB70 which in turn promotes the transcription and expression of genes associated with inflammatory factors, such as IL-1β, TNF-α, NGF, and bradykinin.71–74 Inflammatory pain arises from the direct stimulation of nociceptors in the primary afferent nerve fibers of peripheral tissues, mediated by the local release of inflammatory mediators. These nociceptors include the G protein-coupled receptor (GPCR), transient receptor potential channel (TRP), and acid-sensitive ion channel (ASIC).75,76 Borneol is capable of directly inhibiting the synthesis of IL-1, IL-6, TNF-α, and other inflammatory factors induced by bacterial or aseptic inflammation.77 Additionally, Borneol can indirectly inhibit the production of inflammatory factors by suppressing the activation of the NF-κB pathway. By inhibiting the expression of NFκB-related proteins such as IKKα/β and IκBα, borneol effectively attenuates the upregulation of IL-1, IL-6, TNF-α, and IL-8 mRNA and protein levels induced by Pseudomonas acnes infection in a rat acne model.78
Borneol demonstrates a dose-dependent capacity to mitigate the increase in TNF-α and ICAM-1 in neurons subjected to oxygen-glucose deprivation and reperfusion (OGD/R) and prevents the nuclear translocation of NF-κB and the degradation of IκB in a mouse model of hypoxia-glucose deprivation and reperfusion ischemia.13,25 In an endotoxin model, borneol reduces the elevation of NO, TNF-α, and IL-6 inflammatory factor levels induced by lipopolysaccharide (LPS) in RAW264.7 cells.79 Furthermore, in a mouse ischemia-reperfusion model, borneol inhibits the levels of IL-1β and IL-6 in the cerebral cortex following ischemic events.80 These findings highlight borneol’s potential as a therapeutic agent for mitigating inflammation and inflammatory pain by targeting both the production of inflammatory mediators and the NF-κB signaling pathway.
Inhibition of the Production of NO
Although inducible nitric oxide synthase (iNOS) is not expressed under basal conditions, it can be activated by infection, injury, ischemia, ultraviolet light, cytokines (including TNF), bacteria, viruses, and other stimuli, leading to substantial iNOS production in vivo via the NF-κB pathway.81,82 iNOS catalyzes the conversion of L-arginine to nitric oxide (NO). Nitric oxide (NO) facilitates the synthesis of cyclic guanosine monophosphate (cGMP) by binding to and activating soluble guanylate cyclase through free diffusion to adjacent neurons. Subsequently, cGMP can phosphorylate and activate protein kinase G (PKG) by interacting with its two regulatory domains.83 Activated PKG can phosphorylate two serine residues of the transient receptor potential vanilloid 1 (TRPV1) channel, activating TRPV1 and inducing inflammatory pain.84
Borneol has been shown to inhibit inducible nitric oxide synthase (iNOS) activity, thereby preventing NO synthesis.13 In the permanent middle cerebral artery occlusion (pMCAO) model, (+)-borneol significantly decreased the levels of tumor necrosis factor-alpha (TNF-α) and iNOS proteins in the ischemic region.85 Furthermore, in the endotoxin model, (+)-borneol markedly and dose-dependently reduced the production of TNF-α and iNOS proteins in lipopolysaccharide (LPS)-stimulated BV-2 microglia.85
In short, borneol effectively inhibits iNOS activity, reducing NO synthesis and its downstream inflammatory and pain-promoting effects which has been demonstrated in various models. Those findings suggest that borneol’s inhibition of iNOS and NO production contributes to its anti-inflammatory and analgesic properties, making it a potential therapeutic candidate for conditions involving excessive NO-mediated inflammation and pain.
Inhibition of Oxidative Damage
Reactive oxygen species (ROS) are intricately linked to the inflammatory response. During this response, increased NF-kB activity can enhance the expression of NLRP3.86 Concurrently, ROS facilitate the dissociation of the TXINP-Trx complex, enabling TXINP to bind to NLRP3, which subsequently recruits Asc and Procaspase-1/12 to form active inflammasomes. This process induces the production and release of IL-1β and IL-18.87,88 Intracellular ROS primarily originate from three sources. Inflammatory mediators can stimulate NADPH oxidase to generate ROS by promoting the phosphorylation of p67phox (also known as neutrophil cytosolic factor 2, NCF2).89 Additionally, during mitochondrial electron transport, approximately 1–2% of electrons may leak and react with oxygen, forming superoxide anions;90 Moreover, the uncoupling of endothelial nitric oxide synthase (eNOS) can lead to electron transfer to molecular oxygen, generating ROS.91 Borneol has been shown to mitigate inflammation by inhibiting ROS production through various mechanisms.92 In a dose-dependent manner, borneol significantly reduced intracellular ROS generation induced by oxygen-glucose deprivation/reperfusion (OGD/R) exposure in an animal model of hypoxia-glucose deprivation-reperfusion ischemia.13 Through the activation of protein kinase C (PKC) and NADPH oxidase, (+)-borneol effectively inhibited the production of reactive oxygen species (ROS) by neutrophils in response to phorbol-12-myristate-13-acetate (PMA).93 Additionally, borneol significantly reduced ROS production induced by amyloid-beta (Aβ) in human neuroblastoma (SH-SY5Y) tumor cells by promoting the nuclear translocation of nuclear factor erythroid 2-related factor 2 (Nrf2) and enhancing the expression of heme oxygenase-1 (HO-1).94 Furthermore, borneol mitigated oxidative stress-induced damage to nerve cells by increasing the levels of superoxide dismutase (SOD) and glutathione peroxidase (GSH-Px) in the cortical region, thereby reducing malondialdehyde (MDA) levels.80 Borneol mitigates inflammation by inhibiting ROS production through various means such as reducing OGD/R-induced intracellular ROS, inhibiting neutrophil ROS production, reducing Aβ-induced ROS in SH - SY5Y cells, and alleviating oxidative stress damage to nerve cells.
Others
Borneol also demonstrated the ability to inhibit the migration and infiltration of inflammatory cells, facilitate the resolution of inflammation, and exert neuroprotective effects. In a rat model of acne, topical application of borneol on the ear significantly suppressed the local infiltration of Th1 and Th17 cells, reduced the expression of matrix metalloproteinase 2 (MMP2) via the p38-COX-2-PGE2 signaling pathway, increased extracellular matrix collagen production, and subsequently promoted the healing of inflammation.25 In the inflammatory response model induced by intraperitoneal injection of carrageenan, borneol administered intraperitoneally at doses of 5, 25, or 50 mg/kg effectively inhibited the carrageenan-induced recruitment of leukocytes to the peritoneal cavity.22 Furthermore, in a mouse model of hypoxia-glucose deprivation-reperfusion ischemia, borneol was shown to attenuate the elevation of caspase-3 and caspase-9 activity in neurons caused by oxygen-glucose deprivation (OGD), which inhibiting neuronal apoptosis and exerting neuroprotective effects.13 Thus, we know that borneol has multiple beneficial effects including inhibiting the migration and infiltration of inflammatory cells, promoting inflammation resolution, and showing neuroprotective effects.
Functional Characteristics of (-)-Borneol and Its s From (+)-Borneol
L-borneol (levo-borneol) is an enantiomer of D-borneol (dextro-borneol) and is predominantly derived from the herb Blumea balsamifera, commonly referred to as balsamiferous blumea or ai na xiang in Chinese.95 Pharmacological research indicates that L-borneol exhibits distinct advantages in facilitating penetration, providing neuroprotection, offering cardiovascular protection, and ensuring safety, whereas D-borneol demonstrates superior anti-tumor properties (Figure 4).
 |
Figure 4 The difference of (-)-borneol and (+) - borneol in function. The figure summarizes the pharmacological properties of (-)-borneol (L-borneol), highlighting its key biological activities, including drug penetration enhancement, neuroprotection, cardioprotection, antibacterial adhesion, acute toxicity modulation, and tumor sensitivity regulation.Notably, (-)-borneol exhibits superior permeation-enhancing, neuroprotective, and cardioprotective effects compared to (+)-borneol (D-borneol), as demonstrated in preclinical studies. However, its analgesic potential remains unverified in behavioral tests (eg, hot plate, cold plate, von Frey assays), despite in vitro evidence suggesting modulation of TRPA1, GABA receptors, oxidative stress (ROS), and inflammatory pathways (IL-1β, IL-6, NO). Created in BioRender. Z, (Y) (2025) https://BioRender.com/ype0fk4. Abbreviations: TRPA1, Transient Receptor Potential Ankyrin 1; GABA, Gamma-aminobutyric acid; ROS, Reactive oxygen species; Nrf2, Nuclear factor erythroid 2-related factor 2; HO-1, Heme oxygenase-1; IL, Interleukin; NO, Nitric oxide; 5-HT, Serotonin; nAChRs, Nicotinic acetylcholine receptors. |
Studies have revealed that L-borneol is more effective than D-borneol in enhancing the penetration of hydrophilic drugs, primarily by modulating P-glycoprotein (P-gp) activity.96,97 Moreover, L-borneol exhibits a stronger binding affinity for the CYP2C9 target, which may be associated with its anti-myocardial ischemia effects through the regulation of arachidonic acid metabolism.97,98
Preclinical investigations have shown that oral administration of L-borneol significantly improves the bioavailability of osthole in rats, thereby demonstrating superior drug delivery enhancement compared to D-borneol.99 Experimental findings from the permanent middle cerebral artery occlusion (pMCAO) rat model further corroborate the superior neuroprotective efficacy of L-borneol relative to its D-borneol counterpart.100 In rat models of acute myocardial infarction (AMI), the oral administration of L-borneol has demonstrated significantly superior cardioprotective effects compared to D-borneol. This is evidenced by more pronounced improvements in cardiac electrophysiological abnormalities, greater suppression of pro-inflammatory cytokines (IL-1β and IL-6), and enhanced upregulation of the HIF-1α/VEGF signaling pathway, collectively indicating its optimal comprehensive myocardial protection.98 Furthermore, studies have shown that L-borneol is more effective in repairing pathological damage to the blood-brain barrier (BBB).100 It achieves a higher peak concentration in the brain over a longer period and is eliminated more slowly than D-borneol.97 The special stereoconfiguration of L-borneol’s C2 chiral center allows it to more effectively interfere with bacterial adhesion.97 Additionally, D-borneol exhibits higher toxicity in acute toxicity experiments, potentially causing neurological abnormalities and even death, especially at high doses, whereas L-borneol demonstrates a higher safety profile.97,101 D-borneol exhibits enhanced antitumor efficacy when used in conjunction with chemotherapeutic agents such as temozolomide, paclitaxel, and cisplatin, particularly at elevated concentrations. This increased effectiveness is attributed to the inhibition of P-glycoprotein (P-gp) activity, which amplifies the antitumor effects.97
In terms of analgesic properties, the pain-relieving effects of L-borneol have not yet been substantiated through animal behavioral assays. Nonetheless, in vitro studies indicate that L-borneol exerts similar modulatory influences on TRPA1 channels, GABAA receptors, and cellular reactive oxygen species (ROS) generation as D-borneol. Experimental findings have shown that a mixture of L-borneol and D-borneol effectively inhibits calcium influx induced by flufenamic acid (FAA) and allyl isothiocyanate (AITC) in hTRPA1-transfected HEK293T cells.15 Patch-clamp recordings have further revealed that, unlike D-borneol, L-borneol may act as a partial agonist at GABAA receptors. Specifically, (-)-Borneol demonstrates positive modulation at low GABA concentrations (eg, EC1-4, EC5-14, EC15-24), while exhibiting negative modulation at high GABA concentrations.51 From an immunological standpoint, (-)-borneol has the capacity to inhibit the generation of reactive oxygen species (ROS) by facilitating the nuclear translocation of the transcription factor Nrf2 and enhancing the expression of the antioxidant enzyme HO-1.102 Consequently, we hypothesize that L-borneol exhibits considerable analgesic potential, although additional experimental validation is required.
The Similarities and Differences of Menthol and Borneol in Mechanism
Although both borneol and menthol are terpenoids with similar molecular weights (154 vs 156),103,104 they are recognized for their analgesic properties primarily through their action on TRPM8 channels; however, their mechanisms of function differ significantly (Figure 5).
 |
Figure 5 The similarities and differences of menthol and borneol in mechanism. Regarding the TRPM8 channel, the EC50 of menthol is 13 μM, while that of borneol is 65 μM. Menthol has a stronger effect in activating the TRPM8 channel at the same concentration. However, high concentrations of menthol can reduce cell viability and cause hyperalgesia. The downstream pathways after the activation of TRPM8 by the two are partially the same, both involving type II/III metabotropic glutamate receptors. But menthol may also exert its effect through the opioid pathway, and opioid receptor antagonists can block the analgesic effect of menthol, while having no significant effect on the local analgesia of borneol. For the TRPA1 channel, menthol has a biphasic effect on mTRPA1, while borneol shows a monophasic inhibitory effect. Regarding the TRPV3 channel, both can activate this channel. Menthol can activate TRPV3 and inhibit the activation of TRPA1 and TRPV1 to exert an analgesic effect. In addition, menthol has a bidirectional effect on the TRPV1 receptor. Low concentration (3 mM) activates it, while high concentration (10 mM) blocks it, and high concentrations may lead to rapid desensitization of TRPV1. The effect of borneol in this regard has not been mentioned. In addition, both can exert analgesic effects by activating GABAA and inhibiting nAChRs. Menthol can also block 5 - HT3 and the sodium influx mediated by it. However, the effect of borneol on 5 - HT receptors is still unknown, but it can reduce the central 5 - HT level. Created in BioRender. Z, (Y) (2025) https://BioRender.com/ex7hdbf. |
Trpm8
From a potency perspective, patch clamp studies have demonstrated that the EC50 of menthol is 13 μM, whereas that of borneol is 65 μM.105 Calcium imaging data further indicate that menthol is more effective than borneol in activating TRPM8 channels within the concentration range of 10 μM to 2 mM.14
From the standpoint of TRPM8 specificity, the calcium imaging results demonstrated that the calcium influx caused by 100 μM (+)-borneol and 100 μM menthol was not discernible in the control HEK293 cells without TRPM8 expression and under the influence of the TRPM8 channel blocker AMTB.36 However, the behavioral results demonstrated that there was no discernible difference in the analgesic effect caused by 15% menthol between WT and TRPM8-mice in the CFA model, but the analgesic effect caused by 15% (+)-borneol nearly vanished in TRPM8-mice in the CFA model.14 This suggests that borneol’s analgesic action is more dependent on TRPM8 activation compared to menthol.
Regarding cell viability, WST-1 assays on TRPM8-expressing HEK293 cells showed that (+)-borneol concentrations ranging from 0.1 to 1000 μM did not affect cell viability. In contrast, menthol at 100 μM and 1000 μM significantly reduced cell viability, suggesting that at equivalent concentrations, menthol may lead to intracellular Ca2+ overload due to excessive TRPM8 channel activation, thereby increasing the likelihood of cell death.36
From the perspective of tolerable activation temperature, it has been observed that 100 μM concentrations of (+)-borneol and menthol at 25 °C can enhance tear secretion. However, at 35 °C, the tear secretion induced by (+)-borneol is significantly diminished, whereas menthol continues to elicit a certain level of tear secretion.36 The research indicates the differences in thermal sensitivity of borneol and menthol. Regarding downstream activation pathways, the opioid pathway appears to be specific to menthol, while both borneol and menthol share the activation of type II/III metabotropic glutamate receptors, which are involved in TRPM8-mediated analgesia. Behavioral tests have shown that the analgesic effect of a topical application of 15% (+)-borneol in capsaicin model mice is effectively inhibited by the intrathecal administration of the selective group II metabotropic glutamate receptor (mGluR) antagonist LY341495.14 In mice with chronic spinal cord ligation, intrathecal injections of LY341459 and UBP 1112 (a group III mGluR antagonist) significantly counteracted the analgesic effects of intrathecal menthol administration.37 Both the opioid non-selective antagonist naloxone and the selective kappa antagonist norNOB were able to significantly block the analgesic effects induced by menthol,106,107 whereas intrathecal injection of naloxone did not significantly affect the analgesic action of borneol.14 The tests confirm that borneol’s analgesic effects are blocked by mGluR antagonists, whereas menthol’s effects are also influenced by opioid receptor antagonists.
Furthermore, menthol exhibits a bimodal concentration-dependent effect: low to medium concentrations activate TRPM8, inducing cooling and analgesia, while high concentrations can cause hyperalgesia. In contrast, borneol does not show this bimodal behavior, though further studies with higher concentrations are needed to confirm this. Low to medium concentrations-such as topical applications of 40% or less, plantar injections of 1 mM or 40% or less, and intraperitoneal injections of 100 mg/kg or less generally activate TRPM8 receptors, eliciting sensations of cold (a physiological response) and analgesia (a pathological response).106,108,109 In contrast, higher concentrations such as topical applications exceeding 640 mM in healthy mice and more than 30% in healthy volunteers are associated with hyperalgesia.110,111 The current borneol tests do not demonstrate a bimodal phenomenon, however, this cannot be conclusively ruled out due to the absence of studies employing high concentrations of borneol.
These findings underscore the potential of borneol as a safer and more specific TRPM8-targeting analgesic compared to menthol, particularly in conditions where opioid pathways are undesirable.
Trpa1
Menthol exhibits a biphasic effect on murine TRPA1 (mTRPA1) and a monophasic activating effect on human TRPA1 (hTRPA1). Patch-clamp experiments have demonstrated that low concentrations of menthol (1–30 μM) can activate the channel in CHO cells expressing mTRPA1, whereas higher concentrations (1 mM) result in reversible channel blockage.112 Conversely, menthol is capable of activating hTRPA1 in HEK293T cells expressing this receptor mediated calcium influx.42
The observed discrepancy may be attributed to the formation of hydrogen bonds between menthol and S877 (threonine 877) in mTRPA1,113 while the corresponding residues S873 and S877 in hTRPA1 exhibit slight variations. Borneol demonstrated a monophasic inhibitory effect on hTRPA1. In HEK293T cells expressing hTRPA1, 1 mM borneol completely inhibited calcium influx mediated by menthol (1 mM), FAA (100 µM), and AITC (20 µM), as evidenced by calcium imaging results. The half-maximal inhibitory concentration (IC50) was determined to be 0.2 ± 0.06 mM,42 and patch-clamp studies confirmed that borneol inhibited AITC-induced hTRPA1 currents in a dose-dependent manner. Conversely, the inhibitory effect on mTRPA1 appears to be relatively mild. Patch-clamp results indicate that borneol exerts a more pronounced inhibitory effect on wild-type hTRPA1 compared to hTRPA1-S873V/T874L.42 Behavioral data suggest that in capsaicin and formalin models, the analgesic effect of local (+)-borneol administration is comparable in TRPA1-deficient and wild-type mice.36 In short, menthol has species-specific effects on TRPA1 while borneol acts as a potent inhibitor of hTRPA1 but has minimal effects on mTRPA1, highlighting species-specific differences in TRPA1 modulation.
Trpv3
Both TRPA1 and TRPV3 can be activated, although borneol exhibits a stronger activation effect on TRPV3. According to patch-clamp analysis, 2 mM (+)-borneol was capable of activating TRPV3 under the sensitization of 2-APB, with an EC50 of 3.45 ± 0.13mM. The EC50 value may activate TRPV3 by over 150%, compared to 100% activation with camphor, and is significantly lower than that observed in the control group. Although 2 mM menthol is capable of activating TRPV3 under sensitization conditions, its activation reaches only 65% of that achieved by camphor.46 Furthermore, studies have shown that menthol inhibits TRPV1 and TRPA1-mediated pain sensitivity by activating TRPV3.114 The stronger activation of TRPV3 by borneol highlights its potential significance in mediating analgesic effects, particularly through TRPV3-dependent pathways. Consequently, the role of TRPV3 in the analgesic effects of borneol holds substantial significance.
Gabaa
Both borneol and menthol act as agonists of the GABAA receptor. Patch clamp studies indicate that 100 μM concentrations of (+)-borneol and menthol can enhance the EC20 response of the GABAA receptor in human GABAA recombinant Xenopus oocytes.51 However, menthol exhibits stereoselectivity for GABA, with (+)-menthol being more effective than (-)-menthol,115–117 and increases the response intensity of GABAA to GABA (EC50 = 23 μM). Additionally, menthol can enhance the EC20 response of the glycine receptor without demonstrating stereoselectivity. In contrast, (+)-borneol does not affect glycine receptor currents, whereas both (+) and (-) menthol can increase the glycine EC20 current by approximately 150–200% at 100 μM.36 The observed differences may be attributed to the distinct hydroxyl group positions in the two compounds. In borneol, the isopropyl methyl group is anchored within a rigid bicyclic structure, positioned at a considerable distance from the hydroxyl group. Conversely, in menthol, the isopropyl group is capable of free rotation, allowing the isopropyl methyl group to approach the hydroxyl group in certain conformations.115 Overall, borneol and menthol enhance GABAA receptor responses, but menthol also affects glycine receptors. Structural flexibility in menthol, unlike rigid borneol, explains their differing pharmacological effects.
nAChRs and 5-HT
Both compounds exhibit analgesic properties through the inhibition of nicotinic acetylcholine receptors (nAchRs), however, they may differentially affect serotonin (5-HT) receptors. Menthol has been demonstrated to inhibit 5-HT3 receptors and the sodium influx mechanism mediated by 5-HT3 activation.118–120 In contrast, borneol has been shown to elevate 5-HT levels in the brain.63 This suggests that if 5-HT receptors contribute to borneol’s analgesic effects, borneol may enhance analgesia in combination with fluoxetine and sumatriptan by modulating potassium ion efflux and activating 5-HT1D receptors.121,122 Both borneol and menthol inhibit nAchRs for analgesia, but menthol blocks 5-HT3 receptors, while borneol increases brain 5-HT levels, potentially enhancing analgesia via 5-HT1D receptor activation and potassium efflux modulation.
Other Receptors
Furthermore, menthol interacts with the TRPV1 receptor in a concentration-dependent manner: low concentrations (3 mM) activate TRPV1, whereas high concentrations (10 mM) inhibit it.123,124 In other words, a high concentration of menthol may act on TRPV1 similarly to other compounds, such as RhTx2, resulting in rapid desensitization following initial activation.125,126 However, borneol’s interaction with TRPV1 has not been reported.
Conclusion
Borneol, a traditional Chinese herbal medicine, has emerged as a promising candidate for pain management through a multitude of pre-clinical studies. In pre-clinical models, it has demonstrated remarkable efficacy in alleviating both acute and chronic pain. The analgesic effects of borneol are mediated through a complex network of receptor-related and immune-related mechanisms.
Receptor-related mechanisms involve interactions with multiple key receptors. Borneol can activate TRPM8, leading to increased calcium ion influx and pain signal inhibition in the peripheral nervous system. It also inhibits TRPA1, which reduces the activation of nociceptive neurons in response to harmful stimuli. Additionally, borneol shows activity as a full or partial agonist of GABAA receptors, promoting chloride ion influx and neuronal hyperpolarization in the central nervous system. It increases 5-HT levels in the brain, potentially modulating pain transmission, and non-competitively inhibits nicotinic acetylcholine receptors, further contributing to its analgesic effect.
Immune-related mechanisms play a crucial role as well. Borneol effectively inhibits the production of inflammatory factors such as IL - 1β, TNF - α, and IL - 6 by blocking the NF - κB pathway. It also suppresses the production of NO by inhibiting iNOS activity, reducing oxidative damage by blocking ROS generation, and inhibits the migration and infiltration of inflammatory cells. These immune-modulating effects ultimately lead to a reduction in inflammation-associated pain.
When compared to menthol, another terpenoid with analgesic properties, borneol exhibits distinct differences in receptor - binding potency, specificity, and downstream activation pathways.
In conclusion, borneol represents a promising natural compound for pain management, with a unique mechanism of action that distinguishes it from menthol and other conventional analgesics. Its potential for clinical application, combined with its favorable safety profile, warrants further investigation and development as a modern therapeutic agent for pain relief.
Future Perspectives
Despite the significant progress in understanding borneol’s analgesic properties, several areas require further exploration. First, the long - term safety and toxicity of borneol in humans remain to be fully established. Although it has been approved by the FDA as a condiment and excipient for food and toiletries, its safety profile in the context of chronic pain treatment needs to be investigated through well - designed clinical trials. Second, given the complex nature of pain and the multiple pathways involved in borneol’s analgesic action, combination therapies with other analgesics or drugs that target different pain mechanisms could be explored. This could potentially enhance the analgesic effect and reduce the dosage-related side effects of individual drugs. Finally, more research is required to explore the potential of borneol in treating specific pain conditions, such as cancer pain and fibromyalgia, which are often challenging to manage with current therapies.
Abbreviations
5-HT, 5-hydroxytryptamine (serotonin); 6-OHDA; 6-Hydroxydopamine; Aβ, Amyloid-beta; AITC, Allyl isothiocyanate; AMTB, N-(4-tert-butylphenyl)-4-(3-chloropyridin-2-yl)piperazine-1-carboxamide; ASIC, Acid-sensitive ion channel; BDNF, Brain-derived neurotrophic factor; BCTC, N-(4-butylphenyl)-4-(3-chloropyridin-2-yl)piperazine-1-carboxamide; COX, Cyclooxygenase; CFA, Complete Freund’s adjuvant; cGMP, Cyclic guanosine monophosphate; DRG, Dorsal root ganglion; DMPP, 1,1-dimethyl-4-phenylpiperazinium iodide; EC50, Half-maximal effective concentration; eNOS, Endothelial nitric oxide synthase; GABAA, Gamma-aminobutyric acid type A; GABAAR, GABAA receptor; GPCR, G protein-coupled receptor; GSH-Px, Glutathione peroxidase; hTRPA1, Human TRPA1; hTRPM8, Human TRPM8; HPLC-ECD, High-performance liquid chromatography-electrochemical detection; IC50, Half-maximal inhibitory concentration; iNOS, Inducible nitric oxide synthase; IKK, IκB kinase; IL, Interleukin; LPS, Lipopolysaccharide; mGluRs, Metabotropic glutamate receptors; MDA, Malondialdehyde; MMP2, Matrix metalloproteinase 2; NADPH, Nicotinamide adenine dinucleotide phosphate; nAChRs, Nicotinic acetylcholine receptors; NF-κB, Nuclear factor kappa-light-chain-enhancer of activated B cells; NLRP3, NLR family pyrin domain containing 3; NO, Nitric oxide; NSAIDs, Non-steroidal anti-inflammatory drugs; OGD/R, Oxygen-glucose deprivation and reperfusion; PAMPs, Pathogen-associated molecular patterns; PDT, Photodynamic therapy; PKG, Protein kinase G; PMA, Phorbol-12-myristate-13-acetate; PGE2, Prostaglandin E2; RvD1, Resolvin D1; ROS, Reactive oxygen species.
Acknowledgments
This work was supported by the National Natural Science Foundation of China C-MW (82474211), the Development Plan of Traditional Chinese Medicine Science and Technology in Jiangsu Province C-MW (MS2024002) and a Project Funded by the Priority Academic Program Development of Jiangsu Higher Education Institutions (Integrated Traditional Chinese and Western).
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Disclosure
The authors declare no competing interest.
References
1. Cohen SP, Vase L, Hooten WM. Chronic pain: an update on burden, best practices, and new advances. Lancet. 2021;397(10289):2082–2097. doi:10.1016/s0140-6736(21)00393-7
2. Zou Z, An MM, Xie Q, et al. Single dose intra-articular morphine for pain control after knee arthroscopy. Cochrane Database Syst Rev. 2016;2016(5):CD008918. doi:10.1002/14651858.CD008918.pub2
3. Paul AK, Smith CM, Rahmatullah M, et al. Opioid analgesia and opioid-induced adverse effects: a review. Pharmaceuticals. 2021;14(11):1091. doi:10.3390/ph14111091
4. Brenner DM, Lacy BE. Antispasmodics for chronic abdominal pain: analysis of North American treatment options. Am J Gastroenterol. 2021;116(8):1587–1600. doi:10.14309/ajg.0000000000001266
5. Stiller CO, Hjemdahl P. Lessons from 20 years with COX‐2 inhibitors: importance of dose–response considerations and fair play in comparative trials. J Internal Med. 2022;292(4):557–574. doi:10.1111/joim.13505
6. Yang MY, Khine AA, Liu JW, et al. Resolution of isoborneol and its isomers by GC/MS to identify “synthetic” and “semi-synthetic” borneol products. Chirality. 2018;30(11):1233–1239. doi:10.1002/chir.23017
7. Chinese Pharmacopoeia Commission. Chinese Pharmacopoeia. Beijing: China Medical Science Press; 2015.
8. Kou Z. Ben Cao Yan Yi. Beijing: China Medical Science Press; 2012.
9. Li S. Ben Cao Gang Mu. Beijing: People’s Medical Publishing House; 2004.
10. Z-x C, Q-q X, C-s S, et al. Borneol for regulating the permeability of the blood-brain barrier in experimental ischemic stroke: preclinical evidence and possible mechanism. Oxid Med Cell Longev. 2019;2019:1–15. doi:10.1155/2019/2936737
11. Xin Y, Zhao H, Xu J, et al. Borneol-modified chitosan: antimicrobial adhesion properties and application in skin flora protection. Carbohydr Polym. 2020;228:115378. doi:10.1016/j.carbpol.2019.115378
12. de Sousa DP, Raphael E, Brocksom U, Brocksom TJ. Sedative effect of monoterpene alcohols in mice: a preliminary screening. Z Naturforsch C J Biosci. 2007;62(7–8):563–566. doi:10.1515/znc-2007-7-816
13. Liu R, Zhang L, Lan X, et al. Protection by borneol on cortical neurons against oxygen-glucose deprivation/reperfusion: involvement of anti-oxidation and anti-inflammation through nuclear transcription factor κappaB signaling pathway. Neuroscience. 2011;176:408–419. doi:10.1016/j.neuroscience.2010.11.029
14. Wang S, Zhang D, Hu J, et al. A clinical and mechanistic study of topical borneol-induced analgesia. EMBO Mol Med. 2017;9(6):802–815. doi:10.15252/emmm.201607300
15. Takaishi M, Uchida K, Fujita F, Tominaga M. Inhibitory effects of monoterpenes on human TRPA1 and the structural basis of their activity. J Physiol Sci. 2014;64(1):47–57. doi:10.1007/s12576-013-0289-0
16. Vogt-Eisele AK, Weber K, Sherkheli MA, et al. Monoterpenoid agonists of TRPV3. Br J Pharmacol. 2007;151(4):530–540. doi:10.1038/sj.bjp.0707245
17. Raja SN, Carr DB, Cohen M, et al. The revised international association for the study of pain definition of pain: concepts, challenges, and compromises. Pain. 2020;161(9):1976–1982. doi:10.1097/j.pain.0000000000001939
18. Freynhagen R, Parada HA, Calderon-Ospina CA, et al. Current understanding of the mixed pain concept: a brief narrative review. Curr Med Res Opin. 2019;35(6):1011–1018. doi:10.1080/03007995.2018.1552042
19. Li C, Li Q, Xu J, et al. The efficacy and safety of compound danshen dripping pill combined with percutaneous coronary intervention for coronary heart disease. Evid Based Complement Alternat Med. 2020;2020:5067137. doi:10.1155/2020/5067137
20. Huang M, Du H, Lai J, et al. Clinical efficacy of Kuanxiong aerosol for patients with prehospital chest pain: a randomized controlled trial. Phytomedicine. 2024;123:155206. doi:10.1016/j.phymed.2023.155206
21. Nesterkina M, Kravchenko I. Synthesis and pharmacological properties of novel esters based on monoterpenoids and glycine. Pharmaceuticals. 2017;10(2):47. doi:10.3390/ph10020047
22. Almeida JRGS, Souza GR, Silva JC, et al. Borneol, a bicyclic monoterpene alcohol, reduces nociceptive behavior and inflammatory response in mice. Sci World J. 2013;2013(1). doi:10.1155/2013/808460
23. Bułdyś K, Górnicki T, Kałka D, et al. What do we know about nociplastic pain? Healthcare. 2023;11(12):1794. doi:10.3390/healthcare11121794
24. Jiang J, Shen YY, Li J, Lin YH, Luo CX, Zhu DY. (+)-Borneol alleviates mechanical hyperalgesia in models of chronic inflammatory and neuropathic pain in mice. Eur J Pharmacol. 2015;757:53–58. doi:10.1016/j.ejphar.2015.03.056
25. Ji J, Zhang R, Li H, Zhu J, Pan Y, Guo Q. Analgesic and anti-inflammatory effects and mechanism of action of borneol on photodynamic therapy of acne. Environ Toxicol Pharmacol. 2020;75:103329. doi:10.1016/j.etap.2020.103329
26. Finnerup NB, Haroutounian S, Kamerman P, et al. Neuropathic pain: an updated grading system for research and clinical practice. Pain. 2016;157(8):1599–1606. doi:10.1097/j.pain.0000000000000492
27. Scholz J, Broom DC, Youn D-H, et al. Blocking caspase activity prevents transsynaptic neuronal apoptosis and the loss of inhibition in lamina ii of the dorsal horn after peripheral nerve injury. J Neurosci. 2005;25(32):7317–7323. doi:10.1523/jneurosci.1526-05.2005
28. Sucena E, Delon I, Jones I, Payre F, Stern DL. Regulatory evolution of shavenbaby/ovo underlies multiple cases of morphological parallelism. Nature. 2003;424(6951):935–938. doi:10.1038/nature01768
29. Koivisto AP, Belvisi MG, Gaudet R, Szallasi A. Advances in TRP channel drug discovery: from target validation to clinical studies. Nat Rev Drug Discov. 2022;21(1):41–59. doi:10.1038/s41573-021-00268-4
30. Rosenbaum T, Morales-Lazaro SL, Islas LD. TRP channels: a journey towards a molecular understanding of pain. Nat Rev Neurosci. 2022;23(10):596–610. doi:10.1038/s41583-022-00611-7
31. Yin Y, Lee S-Y. Current view of ligand and lipid recognition by the menthol receptor TRPM8. Trends Biochem Sci. 2020;45(9):806–819. doi:10.1016/j.tibs.2020.05.008
32. Andersen HH, Olsen RV, Møller HG, Eskelund PW, Gazerani P, Arendt‐Nielsen L. A review of topical high‐concentration L‐menthol as a translational model of cold allodynia and hyperalgesia. Eur J Pain. 2013;18(3):315–325. doi:10.1002/j.1532-2149.2013.00380.x
33. Izquierdo C, Martín-Martínez M, Gómez-Monterrey I, González-Muñiz R. TRPM8 channels: advances in structural studies and pharmacological modulation. Int J Mol Sci. 2021;22(16):8502. doi:10.3390/ijms22168502
34. Raddatz N, Castillo JP, Gonzalez C, Alvarez O, Latorre R. Temperature and voltage coupling to channel opening in transient receptor potential melastatin 8 (TRPM8). J Biol Chem. 2014;289(51):35438–35454. doi:10.1074/jbc.M114.612713
35. Chung M-K, Caterina MJ. TRP channel knockout mice lose their cool. Neuron. 2007;54(3):345–347. doi:10.1016/j.neuron.2007.04.025
36. Xu S-Z, Chen G-L, Lei M, Zhou L-P, Zeng B, Zou F. Borneol is a TRPM8 agonist that increases ocular surface wetness. PLoS One. 2016;11(7). doi:10.1371/journal.pone.0158868
37. Proudfoot CJ, Garry EM, Cottrell DF, et al. Analgesia mediated by the TRPM8 cold receptor in chronic neuropathic pain. Curr Biol. 2006;16(16):1591–1605. doi:10.1016/j.cub.2006.07.061
38. Kobayashi K, Fukuoka T, Obata K, et al. Distinct expression of TRPM8, TRPA1, and TRPV1 mRNAs in rat primary afferent neurons with aδ/c‐fibers and colocalization with trk receptors. J Comp Neurol. 2005;493(4):596–606. doi:10.1002/cne.20794
39. Obata K. TRPA1 induced in sensory neurons contributes to cold hyperalgesia after inflammation and nerve injury. J Clin Investig. 2005;115(9):2393–2401. doi:10.1172/jci25437
40. Moran MM, McAlexander MA, Bíró T, Szallasi A. Transient receptor potential channels as therapeutic targets. Nat Rev Drug Discov. 2011;10(8):601–620. doi:10.1038/nrd3456
41. Bautista DM, Movahed P, Hinman A, et al. Pungent products from garlic activate the sensory ion channel TRPA1. Proc Natl Acad Sci U S A. 2005;102(34):12248–12252. doi:10.1073/pnas.0505356102
42. Takaishi M, Uchida K, Fujita F, Tominaga M. Inhibitory effects of monoterpenes on human TRPA1 and the structural basis of their activity. J Physiol Sci. 2013;64(1):47–57. doi:10.1007/s12576-013-0289-0
43. Zhou -H-H, Zhang L, Zhou Q-G, Fang Y, Ge W-H. (+)-Borneol attenuates oxaliplatin-induced neuropathic hyperalgesia in mice. Neuro Rep. 2016;27(3):160–165. doi:10.1097/wnr.0000000000000516
44. Peier AM, Moqrich A, Hergarden AC, et al. A TRP channel that senses cold stimuli and menthol. Cell. 2002;108(5):705–715. doi:10.1016/s0092-8674(02)00652-9
45. Peier AM, Reeve AJ, Andersson DA, et al. A heat-sensitive TRP channel expressed in keratinocytes. Science. 2002;296(5575):2046–2049. doi:10.1126/science.1073140
46. Vogt‐Eisele AK, Weber K, Sherkheli MA, et al. Monoterpenoid agonists of TRPV3. Br J Pharmacol. 2009;151(4):530–540. doi:10.1038/sj.bjp.0707245
47. Huang SM, Lee H, Chung M-K, et al. Overexpressed transient receptor potential vanilloid 3 ion channels in skin keratinocytes modulate pain sensitivity via prostaglandin E2. J Neurosci. 2008;28(51):13727–13737. doi:10.1523/jneurosci.5741-07.2008
48. Bang S, Yoo S, Yang T-J, Cho H, Hwang SW. Isopentenyl pyrophosphate is a novel antinociceptive substance that inhibits TRPV3 and TRPA1 ion channels. Pain. 2011;152(5):1156–1164. doi:10.1016/j.pain.2011.01.044
49. Bang S, Yoo S, Yang TJ, Cho H, Hwang SW. 17(R)‐resolvin D1 specifically inhibits transient receptor potential ion channel vanilloid 3 leading to peripheral antinociception. Br J Pharmacol. 2012;165(3):683–692. doi:10.1111/j.1476-5381.2011.01568.x
50. Niu C, Sun X, Hu F, Tang X, Wang K. Molecular determinants for the chemical activation of the warmth-sensitive TRPV3 channel by the natural monoterpenoid carvacrol. J Biol Chem. 2022;298(3):101706. doi:10.1016/j.jbc.2022.101706
51. Granger RE, Campbell EL, Johnston GAR. (+)- And (−)-borneol: efficacious positive modulators of GABA action at human recombinant α1β2γ2L GABAA receptors. Biochem Pharmacol. 2005;69(7):1101–1111. doi:10.1016/j.bcp.2005.01.002
52. Suo ZW, Fan QQ, Yang X, Hu XD. Ca2+/calmodulin‐dependent protein kinase II in spinal dorsal horn contributes to the pain hypersensitivity induced by γ‐aminobutyric acid type a receptor inhibition. J Neurosci Res. 2013;91(11):1473–1482. doi:10.1002/jnr.23270
53. Cao J, Yang X, Liu Y-N, et al. GABAergic disinhibition induced pain hypersensitivity by upregulating NMDA receptor functions in spinal dorsal horn. Neuropharmacology. 2011;60(6):921–929. doi:10.1016/j.neuropharm.2011.01.034
54. Knabl J, Witschi R, Hösl K, et al. Reversal of pathological pain through specific spinal GABAA receptor subtypes. Nature. 2008;451(7176):330–334. doi:10.1038/nature06493
55. Munro G, Ahring PK, Mirza NR. Developing analgesics by enhancing spinal inhibition after injury: GABAA receptor subtypes as novel targets. Trends Pharmacol Sci. 2009;30(9):453–459. doi:10.1016/j.tips.2009.06.004
56. Li J, Zhang L, Xu C, et al. A pain killer without analgesic tolerance designed by co-targeting PSD-95-nNOS interaction and α2-containning GABAARs. Theranostics. 2021;11(12):5970–5985. doi:10.7150/thno.58364
57. W-r L, R-y C, Yang L, et al. Pharmacokinetics of natural borneol after oral administration in mice brain and its effect on excitation ratio. Eur J Drug Metabol Pharmacokinetics. 2011;37(1):39–44. doi:10.1007/s13318-011-0058-5
58. Nguyen PTT, Jang SH, Rijal S, Park SJ, Han SK. Inhibitory actions of borneol on the substantia gelatinosa neurons of the trigeminal subnucleus caudalis in mice. Korean J Physiol Pharmacol. 2020;24(5):433–440. doi:10.4196/kjpp.2020.24.5.433
59. Sommer C. Serotonin in pain and analgesia: actions in the periphery. Mol Neurobiol. 2004;30(2):117–125. doi:10.1385/MN:30:2:117
60. Bardin L. The complex role of serotonin and 5-HT receptors in chronic pain. Behav Pharmacol. 2011;22(5 and 6):390–404. doi:10.1097/FBP.0b013e328349aae4
61. Liu QQ, Yao XX, Gao SH, et al. Role of 5-HT receptors in neuropathic pain: potential therapeutic implications. Pharmacol Res. 2020;159:104949. doi:10.1016/j.phrs.2020.104949
62. Eide PK, Hole K. The role of 5-hydroxytryptamine (5-HT) receptor subtypes and plasticity in the 5-HT systems in the regulation of nociceptive sensitivity. Cephalalgia. 1993;13(2):75–85. doi:10.1046/j.1468-2982.1993.1302075.x
63. Li W, Yao L, Mi S, Wang N. The effect of borneol on level of HA and 5-HT in rat’s hypothalamus. Zhong Yao Cai. 2004;27(12):937–939.
64. Zoli M, Pucci S, Vilella A, Gotti C. Neuronal and extraneuronal nicotinic acetylcholine receptors. Curr Neuropharmacol. 2018;16(4):338–349. doi:10.2174/1570159X15666170912110450
65. Buccafusco J, Letchworth S, Bencherif M, Lippiello P. Long-lasting cognitive improvement with nicotinic receptor agonists: mechanisms of pharmacokinetic–pharmacodynamic discordance. Trends Pharmacol Sci. 2005;26(7):352–360. doi:10.1016/j.tips.2005.05.007
66. Vincler M. Neuronal nicotinic receptors as targets for novel analgesics. Expert Opin Investig Drugs. 2005;14(10):1191–1198. doi:10.1517/13543784.14.10.1191
67. Park TJ, Park YS, Lee TG, Ha H, Kim KT. Inhibition of acetylcholine-mediated effects by borneol. Biochem Pharmacol. 2003;65(1):83–90. doi:10.1016/s0006-2952(02)01444-2
68. Dib PRB, Quirino-Teixeira AC, Merij LB, et al. Innate immune receptors in platelets and platelet-leukocyte interactions. J Leukoc Biol. 2020;108(4):1157–1182. doi:10.1002/JLB.4MR0620-701R
69. Yang J, Wise L, Fukuchi K-I. TLR4 cross-talk with NLRP3 inflammasome and complement signaling pathways in alzheimer’s disease. Front Immunol. 2020;11. doi:10.3389/fimmu.2020.00724.
70. Zhang T, Ma C, Zhang Z, Zhang H, Hu H. NF‐κB signaling in inflammation and cancer. MedComm. 2021;2(4):618–653. doi:10.1002/mco2.104
71. Gong T, Liu L, Jiang W, Zhou R. DAMP-sensing receptors in sterile inflammation and inflammatory diseases. Nat Rev Immunol. 2019;20(2):95–112. doi:10.1038/s41577-019-0215-7
72. Takeuchi O, Akira S. Pattern recognition receptors and inflammation. Cell. 2010;140(6):805–820. doi:10.1016/j.cell.2010.01.022
73. Liu T, Zhang L, Joo D, Sun S-C. NF-κB signaling in inflammation. Signal Transduction Targeted Ther. 2017;2(1). doi:10.1038/sigtrans.2017.23
74. Bachar O, Adner M, Uddman R, Cardell LO. Toll‐like receptor stimulation induces airway hyper‐responsiveness to bradykinin, an effect mediated by JNK and NF‐κB signaling pathways. Eur J Immunol. 2004;34(4):1196–1207. doi:10.1002/eji.200324569
75. Basbaum AI, Bautista DM, Scherrer G, Julius D. Cellular and molecular mechanisms of pain. Cell. 2009;139(2):267–284. doi:10.1016/j.cell.2009.09.028
76. Matsuda M, Huh Y, Ji -R-R. Roles of inflammation, neurogenic inflammation, and neuroinflammation in pain. J Anesthesia. 2018;33(1):131–139. doi:10.1007/s00540-018-2579-4
77. Bansod S, Chilvery S, Saifi MA, Das TJ, Tag H, Godugu C. Borneol protects against cerulein-induced oxidative stress and inflammation in acute pancreatitis mice model. Environ Toxicol. 2021;36(4):530–539. doi:10.1002/tox.23058
78. Gao F, Chen R, Li S, et al. (+)-Borneol exerts neuroprotective effects via suppressing the NF-kappaB pathway in the pilocarpine-induced epileptogenesis rat model. Brain Res. 2023;1810:148382. doi:10.1016/j.brainres.2023.148382
79. Zou L, Zhang Y, Li W, et al. Comparison of chemical profiles, anti-inflammatory activity, and UPLC-Q-TOF/MS-based metabolomics in endotoxic fever rats between synthetic borneol and natural Borneol. Molecules. 2017;22(9):1446. doi:10.3390/molecules22091446
80. Yu B, Ruan M, Zhang Z-N, Cheng H-B, Shen X-C, Nieber K. Synergic effect of borneol and ligustrazine on the neuroprotection in global cerebral ischemia/reperfusion injury: a region‐specificity study. Evid Based Complement Alternat Med. 2016;2016(1). doi:10.1155/2016/4072809
81. Noack M, Miossec P. Selected cytokine pathways in rheumatoid arthritis. Semin Immunopathol. 2017;39(4):365–383. doi:10.1007/s00281-017-0619-z
82. Kone BC, Schwobel J, Turner P, Mohaupt MG, Cangro CB. Role of NF-kappa B in the regulation of inducible nitric oxide synthase in an MTAL cell line. Am J Physiol. 1995;269(5):F718–29. doi:10.1152/ajprenal.1995.269.5.F718
83. Mónica FZ, Bian K, Murad F. The endothelium-dependent nitric oxide–cGMP pathway. Endothelium. 2016;77:1–27.
84. The contribution of TRPV1 S801 phosphorylation to nociception and inflammatory pain in vivo. 2020.
85. Lei C, Chun-Yu Y, Hai-Yin W, et al. (+)-Borneol is neuroprotective against permanent cerebral ischemia in rats by suppressing production of proinflammatory cytokines. J Biomed Res. 2017;31(4). doi:10.7555/jbr.31.20160138
86. Bauernfeind FG, Horvath G, Stutz A, et al. Cutting edge: NF-κB activating pattern recognition and cytokine receptors license NLRP3 inflammasome activation by regulating NLRP3 expression. J Immunol. 2009;183(2):787–791. doi:10.4049/jimmunol.0901363
87. Schroder K, Tschopp J. The Inflammasomes. Cell. 2010;140(6):821–832. doi:10.1016/j.cell.2010.01.040
88. Zhou R, Tardivel A, Thorens B, Choi I, Tschopp J. Thioredoxin-interacting protein links oxidative stress to inflammasome activation. Nat Immunol. 2009;11(2):136–140. doi:10.1038/ni.1831
89. Dang PMC. A specific p47phox -serine phosphorylated by convergent MAPKs mediates neutrophil NADPH oxidase priming at inflammatory sites. J Clin Investig. 2006;116(7):2033–2043. doi:10.1172/jci27544
90. Handy DE, Loscalzo J. Redox regulation of mitochondrial function. Antioxid Redox Signaling. 2012;16(11):1323–1367. doi:10.1089/ars.2011.4123
91. Förstermann U, Münzel T. Endothelial nitric oxide synthase in vascular disease. Circulation. 2006;113(13):1708–1714. doi:10.1161/circulationaha.105.602532
92. Santos CAL, Moreira AMT, JRdL S, et al. Evaluation of the effect of 2-borneol on the Nauphoeta cinerea model through oxidative stress and acute toxicity tests. Nat Product Res. 2025. doi:10.1080/14786419.2025.2456091
93. Chen H, Xu X, Tang Q, et al. (+)-Borneol inhibits the generation of reactive oxygen species and neutrophil extracellular traps induced by phorbol-12-myristate-13-acetate. Front Pharmacol. 2022;13:1023450. doi:10.3389/fphar.2022.1023450
94. Hur J, Pak SC, Koo B-S, Jeon S. Borneol alleviates oxidative stress via upregulation of Nrf2 and Bcl-2 in SH-SY5Y cells. Pharm Biol. 2012;51(1):30–35. doi:10.3109/13880209.2012.700718
95. Fu M, Lu Z, Ma X. Enhanced extraction efficiency of natural D-borneol from Mei Pian tree leaves pretreated with deep eutectic solvents. Food Sci Nutr. 2020;8(7):3806–3813. doi:10.1002/fsn3.1671
96. Wu CJ, Huang QW, Qi HY, Guo P, Hou SX. Promoting effect of borneol on the permeability of puerarin eye drops and timolol maleate eye drops through the cornea in vitro. Pharmazie. 2006;61(9):783–788.
97. Ma R, Lu D, Wang J, Xie Q, Guo J. Comparison of pharmacological activity and safety of different stereochemical configurations of borneol: l-borneol, D-borneol, and synthetic borneol. Biomed Pharmacother. 2023;164:114668. doi:10.1016/j.biopha.2023.114668
98. Liying W, Jian W, Yamei FAN, et al. Anti-inflammation and regulation of HIF-1α/VEGF of 3 kinds of borneolum in prevention and treatment of AMI model rats. Chin J Exp Trad Med Fromulae. 2022; 28;19:61–72.
99. Luo DD, Chen XY, Zhang ZB, et al. Different effects of (+)‑borneol and (‑)‑borneol on the pharmacokinetics of osthole in rats following oral administration. Mol Med Rep. 2017;15(6):4239–4246. doi:10.3892/mmr.2017.6502
100. Dong T, Chen N, Ma X, et al. The protective roles of L-borneolum, D-borneolum and synthetic borneol in cerebral ischaemia via modulation of the neurovascular unit. Biomed Pharmacother. 2018;102:874–883. doi:10.1016/j.biopha.2018.03.087
101. Bhatia SP, McGinty D, Letizia CS, Api AM. Fragrance material review on isoborneol. Food Chem Toxicol. 2008;46(11):S182–S184. doi:10.1016/j.fct.2008.06.052
102. Hur J, Pak SC, Koo BS, Jeon S. Borneol alleviates oxidative stress via upregulation of Nrf2 and Bcl-2 in SH-SY5Y cells. Pharm Biol. 2013;51(1):30–35. doi:10.3109/13880209.2012.700718
103. Xu L, Han Y, Chen X, et al. Molecular mechanisms underlying menthol binding and activation of TRPM8 ion channel. Nat Commun. 2020;11(1):3790. doi:10.1038/s41467-020-17582-x
104. Hans M, Wilhelm M, Swandulla D. Menthol suppresses nicotinic acetylcholine receptor functioning in sensory neurons via allosteric modulation. Chem Senses. 2012;37(5):463–469. doi:10.1093/chemse/bjr128
105. Luo Y, Sun W, Feng X, et al. (-)-menthol increases excitatory transmission by activating both TRPM8 and TRPA1 channels in mouse spinal lamina II layer. Biochem Biophys Res Commun. 2019;516(3):825–830. doi:10.1016/j.bbrc.2019.06.135
106. Liu B, Fan L, Balakrishna S, Sui A, Morris JB, Jordt SE. TRPM8 is the principal mediator of menthol-induced analgesia of acute and inflammatory pain. Pain. 2013;154(10):2169–2177. doi:10.1016/j.pain.2013.06.043
107. Shapovalov G, Gkika D, Devilliers M, et al. Opiates modulate thermosensation by internalizing cold receptor TRPM8. Cell Rep. 2013;4(3):504–515. doi:10.1016/j.celrep.2013.07.002
108. Patel R, Gonçalves L, Leveridge M, et al. Anti-hyperalgesic effects of a novel TRPM8 agonist in neuropathic rats: a comparison with topical menthol. Pain. 2014;155(10):2097–2107. doi:10.1016/j.pain.2014.07.022
109. Pan R, Tian Y, Gao R, et al. Central mechanisms of menthol-induced analgesia. J Pharmacol Exp Ther. 2012;343(3):661–672. doi:10.1124/jpet.112.196717
110. Tajino K, Matsumura K, Kosada K, et al. Application of menthol to the skin of whole trunk in mice induces autonomic and behavioral heat-gain responses. Am J Physiol Regul Integr Comp Physiol. 2007;293(5):R2128–R2135. doi:10.1152/ajpregu.00377.2007
111. Hatem S, Attal N, Willer J-C, Bouhassira D. Psychophysical study of the effects of topical application of menthol in healthy volunteers. Pain. 2006;122(1):190–196. doi:10.1016/j.pain.2006.01.026
112. Karashima Y, Damann N, Prenen J, et al. Bimodal action of menthol on the transient receptor potential channel TRPA1. J Neurosci. 2007;27(37):9874–9884. doi:10.1523/jneurosci.2221-07.2007
113. Xiao B, Dubin AE, Bursulaya B, Viswanath V, Jegla TJ, Patapoutian A. Identification of transmembrane domain 5 as a critical molecular determinant of menthol sensitivity in mammalian TRPA1 channels. J Neurosci. 2008;28(39):9640–9651. doi:10.1523/jneurosci.2772-08.2008
114. Macpherson LJ, Hwang SW, Miyamoto T, Dubin AE, Patapoutian A, Story GM. More than cool: promiscuous relationships of menthol and other sensory compounds. Mol Cell Neurosci. 2006;32(4):335–343. doi:10.1016/j.mcn.2006.05.005
115. Hall AC, Turcotte CM, Betts BA, Yeung W-Y, Agyeman AS, Burk LA. Modulation of human GABAA and glycine receptor currents by menthol and related monoterpenoids. Eur J Pharmacol. 2004;506(1):9–16. doi:10.1016/j.ejphar.2004.10.026
116. Krasowski MD, Jenkins A, Flood P, Kung AY, Hopfinger AJ, Harrison NL. General anesthetic potencies of a series of propofol analogs correlate with potency for potentiation of gamma-aminobutyric acid (GABA) current at the GABA(A) receptor but not with lipid solubility. J Pharmacol Exp Ther. 2001;297(1):338–351. doi:10.1016/S0022-3565(24)29544-6
117. Oz M, El Nebrisi EG, Yang KS, Howarth FC, Al Kury LT. Cellular and molecular targets of menthol actions. Front Pharmacol. 2017;8:472. doi:10.3389/fphar.2017.00472
118. Abdel-Aziz H, Windeck T, Ploch M, Verspohl EJ. Mode of action of gingerols and shogaols on 5-HT3 receptors: binding studies, cation uptake by the receptor channel and contraction of isolated Guinea-pig ileum. Eur J Pharmacol. 2006;530(1–2):136–143. doi:10.1016/j.ejphar.2005.10.049
119. Ashoor A, Nordman JC, Veltri D, et al. Menthol inhibits 5-HT3 receptor-mediated currents. J Pharmacol Exp Ther. 2013;347(2):398–409. doi:10.1124/jpet.113.203976
120. Walstab J, Wohlfarth C, Hovius R, et al. Natural compounds boldine and menthol are antagonists of human 5-HT3 receptors: implications for treating gastrointestinal disorders. Neurogastroenterol Motil. 2014;26(6):810–820. doi:10.1111/nmo.12334
121. Villalón CM, VanDenBrink AM. The role of 5-hydroxytryptamine in the pathophysiology of migraine and its relevance to the design of novel treatments. Mini Rev Med Chem. 2017;17(11):928–938. doi:10.2174/1389557516666160728121050
122. Pascual J. A review of rizatriptan, a quick and consistent 5-HT1B/1D agonist for the acute treatment of migraine. Expert Opin Pharmacother. 2004;5(3):669–677. doi:10.1517/14656566.5.3.669
123. Takaishi M, Uchida K, Suzuki Y, et al. Reciprocal effects of capsaicin and menthol on thermosensation through regulated activities of TRPV1 and TRPM8. J Physiol Sci. 2015;66(2):143–155. doi:10.1007/s12576-015-0427-y
124. Nguyen THD, Itoh SG, Okumura H, Tominaga M. Structural basis for promiscuous action of monoterpenes on TRP channels. Commun Biol. 2021;4(1). doi:10.1038/s42003-021-01776-0
125. Zhu A, Aierken A, Yao Z, et al. A centipede toxin causes rapid desensitization of nociceptor TRPV1 ion channel. Toxicon. 2020;178:41–49. doi:10.1016/j.toxicon.2020.02.016
126. Szallasi A, Blumberg PM. Vanilloid (Capsaicin) receptors and mechanisms. Pharmacol Rev. 1999;51(2):159–212.
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
TRPA1 Ion Channel Mediates the Analgesic Effects of Acupuncture at the ST36 Acupoint in Mice Suffering from Arthritis

Yao K, Chen Z, Li Y, Dou B, Xu Z, Ma Y, Du S, Wang J, Fu J, Liu Q, Fan Z, Liu Y, Lin X, Xu Y, Fang Y, Wang S, Guo Y
Journal of Inflammation Research 2024, 17:1823-1837
Published Date: 20 March 2024