Back to Journals » OncoTargets and Therapy » Volume 13
Biological Characterization and Therapeutics for Subscalp Recurrent in Intracranial Glioblastoma
Authors Zhang J ![]() , Fang S, Song W, Zhang B, Fan W, Jin G, Liu F
, Fang S, Song W, Zhang B, Fan W, Jin G, Liu F ![]()
Received 9 June 2020
Accepted for publication 28 August 2020
Published 11 September 2020 Volume 2020:13 Pages 9085—9099
DOI https://doi.org/10.2147/OTT.S265322
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Sanjay Singh
Junwen Zhang,* Sheng Fang,* Wenjie Song, Bo Zhang, Wenhua Fan, Guishan Jin, Fusheng Liu
Brain Tumor Research Center, Beijing Neurosurgical Institute, Beijing Laboratory of Biomedical Materials, Beijing Tiantan Hospital Affiliated to Capital Medical University, Beijing 100070, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Fusheng Liu
Brain Tumor Research Center, Beijing Neurosurgical Institute, Department of Neurosurgery, Beijing Tiantan Hospital Affiliated to Capital Medical University, No. 119 Nansihuanxilu, Fengtai District, Beijing 100070, People’s Republic of China
Tel +86-10-59975626
Email [email protected]
Purpose: Gliomas are common intracranial tumors, of which 70% are malignant gliomas. Glioblastoma multiforme (GBM) is the most aggressive tumor, and patients with GBM have a median survival time of only 9– 12 months; extracranial recurrence of GBM is very rare. A therapeutic strategy for this kind of recurrent tumor is lacking.
Materials and Methods: We present a case of a patient with extracranial recurrence of subscalp GBM. The subscalp tumor was resected and xenotransplanted into BALB/C nude mice. Then, glioma cells were isolated from the xenograft models and passaged in vitro. HE staining, immunohistochemistry, CCK-8 assays, karyotypic analysis, short tandem repeat STR analysis and flow cytometry were used to analyze the biological characteristics and malignant phenotype of these established cells. The cells and xenografts were then used as preclinical models to evaluate the antitumor efficacy of oncolytic herpes simplex virus 1 (oHSV-1).
Results: The isolated cells, which were named BT-01, were positive for Nestin and GFAP. The main characteristics of BT-01 cells were that they harbored glioblastoma stem-like cells (GSCs) and that they possessed highly aggressive migration capacities compared with the existing cell lines U87-MG and U251-MG. Moreover, BT-01 cells tolerated the chemotherapeutic drug temozolomide. Our study showed that oHSV-1 could replicate in and repress the growth of BT-01 cells and significantly inhibit tumor growth in xenograft models.
Conclusion: Taken together, our results showed that a new recurrent glioblastoma cell line was established, which can be useful for research on recurrent glioblastoma. We provided a reliable preclinical model to evaluate the antitumor efficacy of oHSV-1 in vivo and a promising therapy for recurrent GBM.
Keywords: recurrent glioblastoma, patient-derived xenograft, BT-01 cells, glioblastoma stem-like cells, oncolytic herpes simplex virus 1
Introduction
Human glioblastoma multiforme (GBM) is a devasting brain tumor that is characterized by high mortality and poor prognosis.1 GBM can invasively grow into nearby brain parenchyma and is difficult to fully resect. Even when patients are treated according to the standard of care, which consists of surgical resection followed by radiotherapy and chemotherapy, the outcome of patients with GBM remains dismal, and the median survival time is approximately 9–12 months.2 In addition, the proportion of extracranial recurrence in patients with glioblastoma is only 0.4%-2%, but the prognosis of these kind of patients is poor.3–5 Therefore, the mechanisms underlying recurrence, radiotherapy and chemotherapy tolerance, invasion, and potential treatments for glioblastoma require further study.
Temozolomide (TMZ) is currently the standard chemotherapeutic agent used for GBM treatment after surgery and causes DNA damage in rapidly proliferating cells. However, no satisfactory outcomes were obtained, especially in patients with unmethylated O (6)-methylguanine-DNA-methyltransferase (MGMT) status.6 Furthermore, TMZ chemotherapy only improved the median survival time from 14.6 to 12.1 months.7 Moreover, resistance to TMZ is another concern for the use of TMZ for GBM therapy, but whether it could be used to treat extracranial recurrent glioma remains to be elucidated.
The use of genetically engineered herpes simplex virus type I (HSV-1) is a promising therapeutic approach for GBM.8 After gene modification, HSV-1 selectively replicates in tumor cells and eventually leads to tumor destruction.9 Oncolytic HSV-1 (oHSV-1) is not limited to targeting a specific signaling molecule or pathway and has a wide range of effects on neoplastic cells. oHSV-1 has been utilized in several clinical trials and was proven to be an efficient therapy for malignant GBM.10
Human cancer cell lines and cell-derived xenograft (CDX) models are the most commonly used models in cancer research and have become the backbone of tumor biology research and drug discovery.11 However, the most commonly used GBM cell lines are almost all derived from orthotopic tumors, and hardly any of them are derived from extracranial GBM recurrence. The PDX model was established by inoculating human tissues directly into immunodeficient mice to preserving features such as tumor heterogeneity. Organoids can greatly preserve the tumor phenotype and molecular heterogeneity.12 However, due to the high technical threshold, the establishment of organoids requires high technical capability. To test the efficacy of oncolytic viruses, PDX models are needed because, unlike CDX models, PDX models contain immune cells.
In this study, a new glioblastoma cell line derived from a recurrent subscalp tumor originating from intracranial GBM was established. The following study characterized the isolated cells, named BT-01 cells, which contained abundant GSCs showing self-renewal and multilineage differentiation capacities and significant therapeutic resistance to TMZ. Moreover, these cells exhibited highly aggressive migration capacity compared with the more commonly used U87-MG and U251-MG cells. We evaluated the antitumor efficacy of oHSV-1 using BT-01 cells and the PDX model.
Materials and Methods
Cells and Cell Culture
U87-MG and U251-MG cells were obtained from the Cell Resource Center, Peking Union Medical College (which is the headquarters of the National Infrastructure of Cell Line Resources, NSTI). These cells were cultured in Dulbecco’s modified Eagle medium (DMEM) supplemented with 10% fetal bovine serum and 100 U/mL penicillin-streptomycin. Cells were cultured in a humidified incubator at 37°C in 5% CO2.
BT-01 cells were the new established cell line and cells were authenticated by STR profile (Table 1).
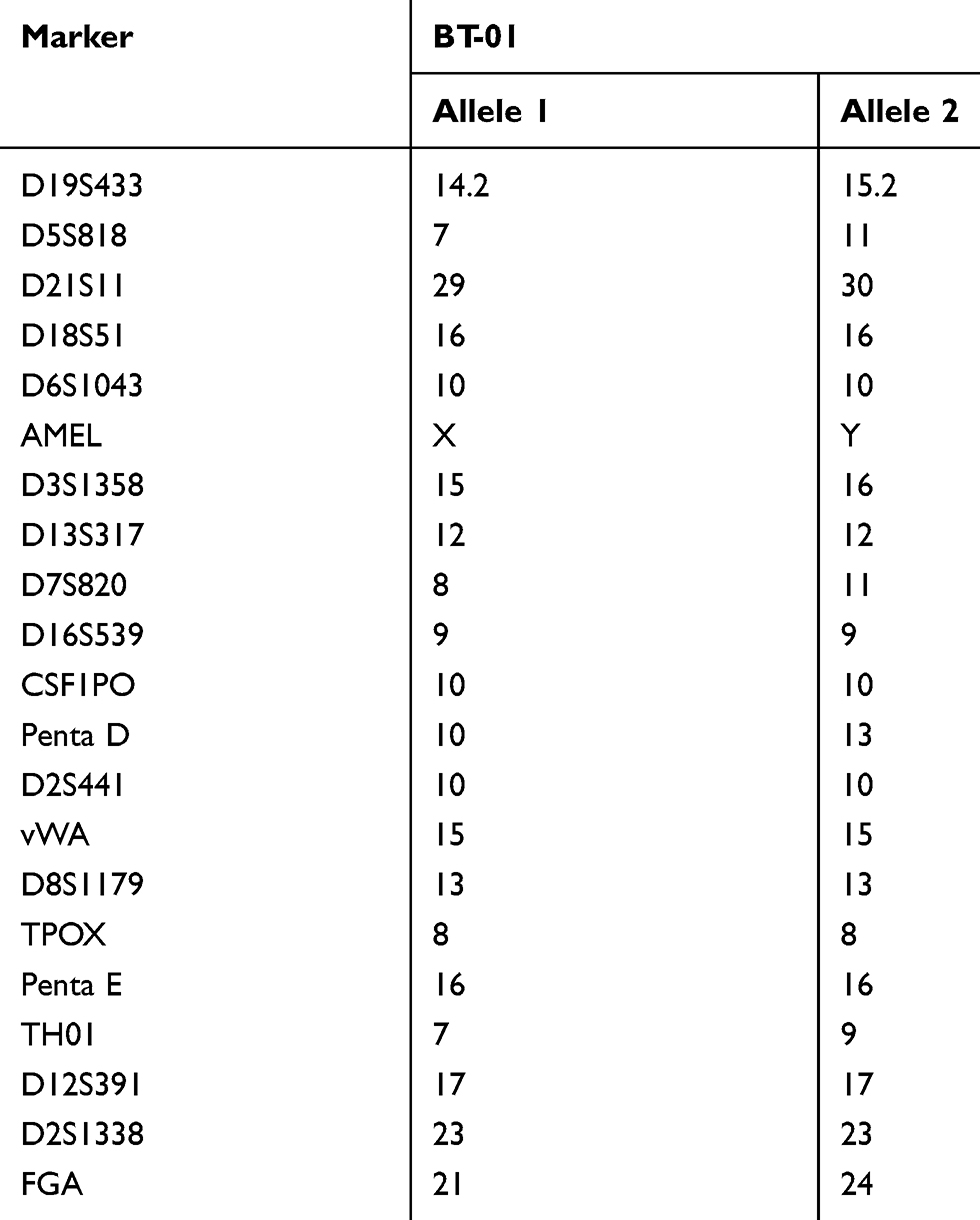 |
Table 1 Results of the Short Tandem Repeats Analysis for BT-01 Cell |
Patient History
A 37-year-old man was referred to the Neurosurgery Department at Beijing Tiantan Hospital because of impaired vision in the right eye in March 2017. Magnetic resonance imaging revealed a space-occupying lesion of the right forehead (Figure 1A and B). After surgery, the tumor was fully resected, and the pathological diagnosis was glioblastoma.
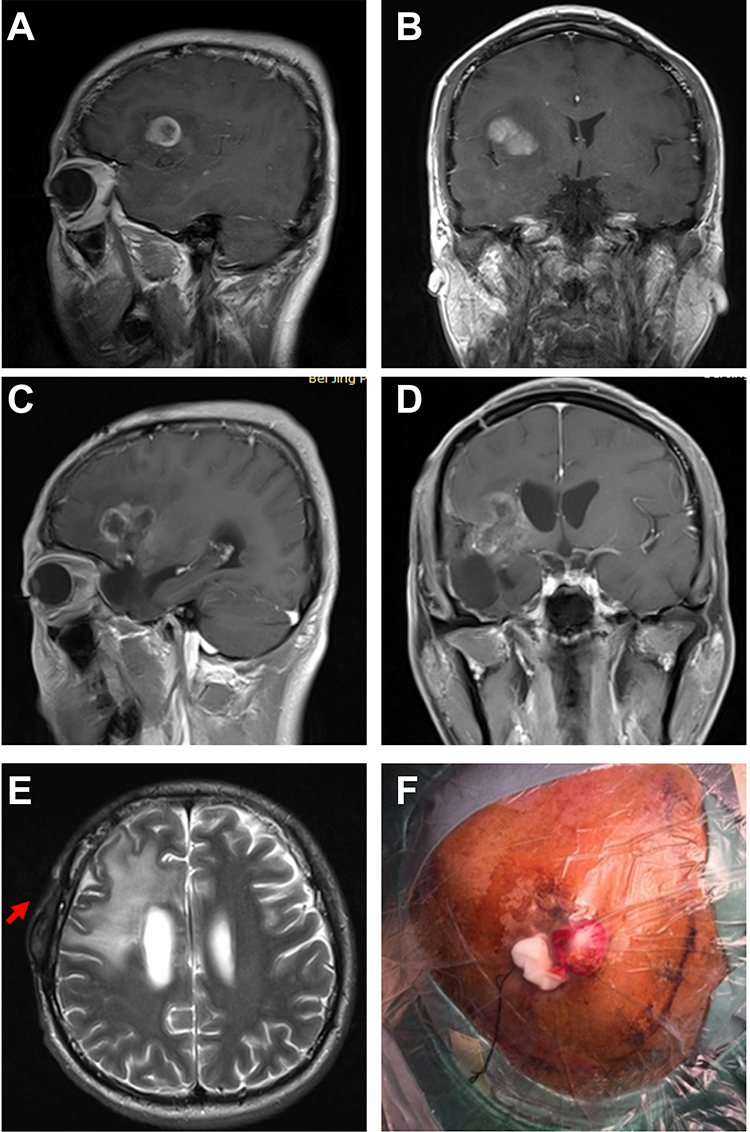 |
Figure 1 Recurrent glioblastoma with subscalp metastasis. (A and B) Preoperative sagittal (A) and coronal (B) T1-weighted contrast-enhanced magnetic resonance imaging (MRI) of primary glioma. (C and D) Preoperative sagittal (C) and coronal (D) T1-weighted contrast-enhanced magnetic resonance imaging (MRI) after recurrence. MRI showed an irregular intracranial occupying lesion with surrounding enhancement. (E) Preoperative axial T2-weighted cranial MRI showed a scalp mass (red arrow). (F) Preoperative image of a subscalp mass. |
Two months after the first surgery, this patient with GBM presented with tumor recurrence (Figure 1C and D). Then, the patient underwent a second surgery, which was followed by radiotherapy and chemotherapy after the initial diagnosis.
Five months after the second operation, the tumor recurred again, and a scalp mass was found. Magnetic resonance imaging (MRI) revealed an intracranial space-occupying lesion (Figure 1E) and a scalp mass (Figure 1F). MRI also showed that the space between the intracranial dura mater and subarachnoid under the cranial bone flap was continuous within the scope of surgery. The tumor was hardly contiguous and not caused by direct tumor growth from the skull defect (Figure S1). The tumor was treated with full resection. After the third surgery, the patient received comprehensive treatments, such as radiotherapy and chemotherapy, but the effects were poor. The patient died in August 2018.
PDX Establishment
After resection during surgery, the tumor specimens were cut into small pieces. A piece of tumor was implanted subcutaneously into six BALB/c nude mice (6 weeks old) in the right armpit. When the diameter of the tumor reached 1 cm, the mice were sacrificed, and the tumors were transplanted into new mice.
In the fifth generation, approximately 5 days after implantation, the tumors began to grow, and the subcutaneous tumor size in the nude mice was observed and measured every 3 days. The nude mice were sacrificed when the tumors reached a diameter of approximately 1 cm, after which the tumor tissues were fixed in 4% paraformaldehyde, paraffin-embedded and stained with hematoxylin and eosin (H&E).
Immunohistochemistry
Immunohistochemical staining was performed as previously described.13 Ki67, GFAP and Nestin were detected by the following antibodies: anti-Ki67 (1:400, ab16667, Abcam), anti-GFAP (1:500, ab10062, Abcam) and anti-Nestin (1:400, ab6320, Abcam). Images were captured with a microscope.
Primary Culture in vitro
A portion of the fresh tissue was obtained from the fifth generation PDX models and immediately washed in DMEM/F12 medium. After digestion in 0.05% trypsin at 37°C for 10 min, the tissue was placed in a tube, which was placed in a MACS Tube Rotator (MILTENYI Biotec, Germany). Digestion was terminated by the addition of DMEM containing 10% FBS. Then, the suspension was collected by filtration through a 70 μm filter. After resuspension, the resultant cell pellet was treated with erythrocyte lysis solution on ice for 15 min, and the suspension was sequentially centrifuged at 1000 rpm/min, 800 rpm/min, and 600 rpm/min for 3 min each. The collected tumor cells were resuspended in complete medium in a 25 cm2 culture flask and were cultured in a 37°C incubator under 5% CO2. When the cell confluence reached 80%-90%, cell subculturing was performed.
Morphologic Observation and Cell Proliferation Assays in vitro
In total, 2 x 105 BT-01 cells were cultured in a 25 cm2 culture flask for one week in a 37°C incubator under 5% CO2. The morphology and number of BT-01 cells were observed using a microscope daily.
Cell proliferation was detected by using a cell counting kit-8 assay (Dojindo, Japan). BT-01 cells were plated in 96-well plates at a density of 3,000 cells per well in 4 replicate wells and cultured for 1–7 days. Then, cell proliferation was measured according to the manufacturer’s protocols.
Karyotypic Analysis
BT-01 cells were treated with colchicine (final concentration 0.2 μg/mL) for 2 h, and then cells were digested in 0.05% trypsin. After hypotonic treatment with a 0.075 mol/L KCl solution at 37°C for 30 min, the cells were fixed in fixative (methanol: glacial acetic acid 3:1, V/V) for approximately 1 h. Then, precooled slides containing the cell suspension were air-dried, stained with Giemsa solution for 20 min, and observed under a microscope.
Immunofluorescence Staining
BT-01 cells were plated at a density of 5×104 cells/mL, incubated in a 35 mm confocal dish for 24 h, rinsed 3 times with PBS, and fixed in 4% paraformaldehyde for 10 min. After blocking with 5% BSA containing 0.3% Triton X-100, the cells were incubated with the respective antibodies (anti-GFAP, 1:200, A-21282, Thermo Fisher; anti-Nestin, 1:100, ab6320, Abcam) overnight at 4°C. After rinsing with PBST, the cells were incubated with the fluorescent secondary antibody (goat anti-mouse IgG, 1:1000, ab150114, Abcam) for 1 h at room temperature. The nuclei were counterstained with DAPI for 10 min. Images were acquired using a fluorescence microscope.
Short Tandem Repeat (STR) Analysis
Genomic DNA from 2×106 BT-01 cells was extracted using a Microread Genomic DNA Kit. The cellular DNA was amplified with an STR Multi Amplification Kit (Microreader TM21 ID System). Twenty STR loci and the sex-linked gene amelogenin were assayed on an ABI 3730xl DNA Analyzer (Applied Biosystems, USA). Data were analyzed using GeneMapper3.2 software and were then compared with data from the ATCC and DSMZ databases for reference matching.
Tumor Sphere-Forming Assay and Differentiation Assay
BT-01 cells were digested in 0.05% trypsin and cultured in neural stem cell medium for one week; the cell morphology was observed daily under a microscope. One week later, the neurospheres formed by BT-01 cells were selected and cultured in complete medium for 24 h, and then the morphology of the cells was observed by microscopy.
A 6-well plate containing neural stem cell medium was seeded with 5×104 BT-01, U87-MG or U251-MG cells per well in separate wells and cultured for 3 days. The morphology of the cells was observed by microscopy.
Flow Cytometry Assay
According to the protocol provided by MILTENYI Biotec, BT-01, U87-MG and U251-MG cells were stained with the fluorescent-conjugated antibodies CD133-PE and IgG-PE, which were purchased from MILTENYI Biotec. The stained cells were analyzed with a Beckman Coulter MOFLO XDP cell sorter.
Drug Sensitivity Assay (CCK-8 Assay)
BT-01, U87-MG and U251-MG cells were plated at a density of 3000 cells per well in 100 µL medium in quadruplicate in 96-well plates. After culturing for 24 h, the cells were simultaneously exposed to temozolomide purchased from Sigma. Cells were collected after incubation for 24, 48 and 72 h and were then treated with CCK-8 reagent for 1 h, after which the absorbance at 450 nm was measured using a microplate reader.
Cell Migration and Invasion Assay
For the cell migration assay, a total of 5×104 cells in 200 µL of serum-free DMEM were seeded in the upper chamber of the Transwell insert, and the lower chamber was filled with 600 µL of 10% FBS medium; each assay was performed in triplicate. After 8 h of incubation, the Transwell chamber was collected and stained with 1% crystal violet. For the cell invasion assay, 5×104 cells in 200 µL of serum-free DMEM were seeded in the upper chamber of the Transwell insert, which was precoated with Matrigel, and the lower chamber contained 600 µL of 15% FBS medium; each assay was performed in triplicate. Twenty-four hours after inoculation, the Transwell chamber was collected and stained with 1% crystal violet.
PDX Injection with oHSV-1
In the fifth-generation PDX model, approximately 5 days after implantation, oHSV-1 (1×107 pfu/100 µL) or PBS was injected into the tumor, and the tumor was injected again 5 days later. After injection, the subcutaneous tumor size of the nude mice was observed and measured every 2 days. The mice were euthanized on day 24, and the tumors were resected.
Results
Establishment of PDX Models and Transcriptome Analysis of the Subscalp Tumor and PDX Tumor
The subscalp tumor tissue resected during surgery was implanted subcutaneously in the right armpit of nude mice. After growing to the appropriate size, the tumors were engrafted into new mice. In the fifth-generation mice, approximately 5 days after implantation, the tumors began to grow, and the subcutaneous tumor size in the nude mice was observed (Figure 2A). In this case, a stable PDX model was established, and the cells could be continuously passaged.
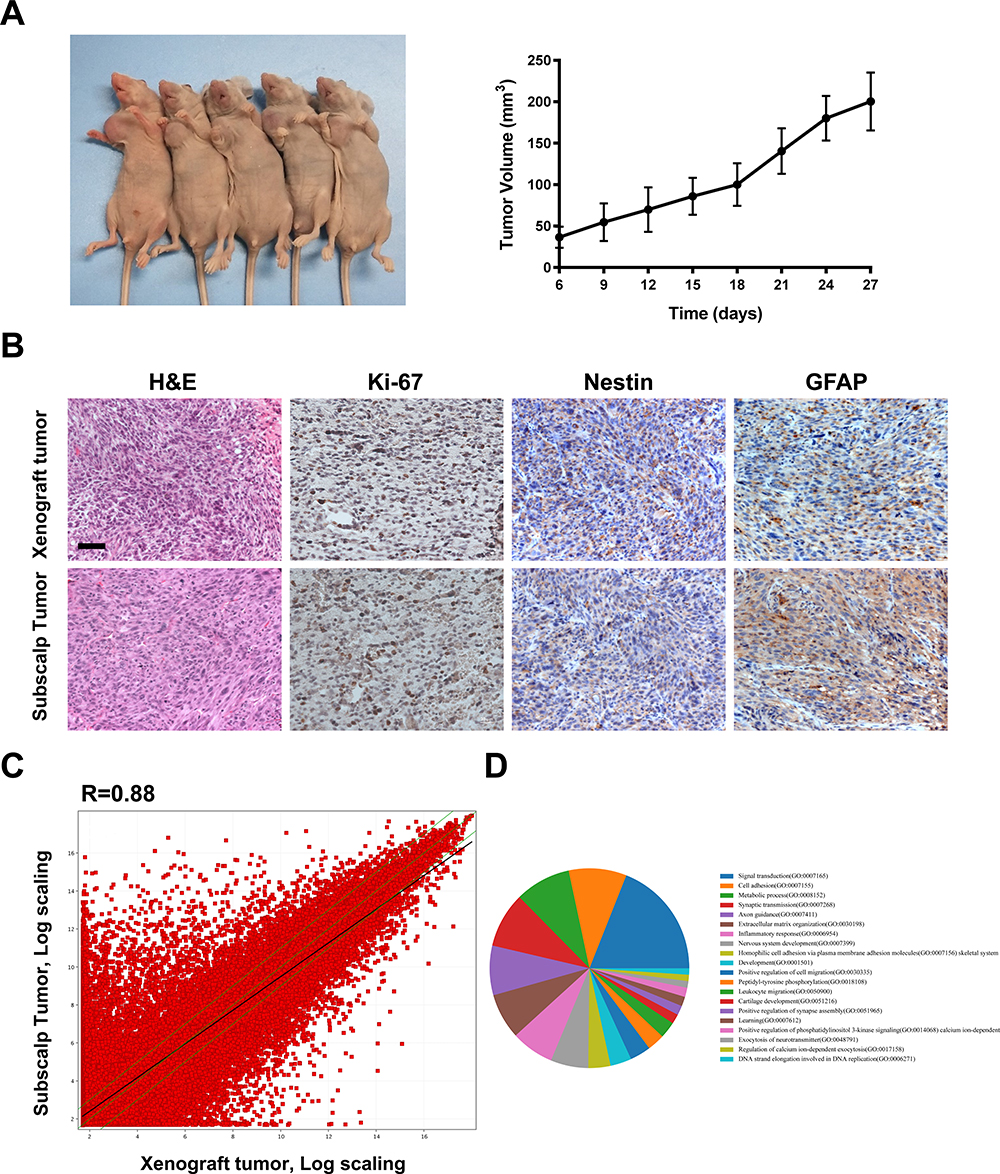 |
Figure 2 Tumor histology and gene expression of the subscalp tumor and the PDX tumor. (A) The PDX model was established using BALB/c nude mice and could be stably grown and passaged. The fifth-generation PDX tumor started to grow 12 days after implantation. (B) H&E staining and immunohistochemical staining of GFAP, Nestin and Ki67 in recurrent tumor tissue and fifth-generation PDX tumor tissue. Scale bar represents 25 μm. (C) A scatter plot showing the correlation of gene expression between the subscalp tumor and the fifth-generation PDX tumor tissue. R represents the correlation coefficient. (D) A pie chart of the main different biological processes. |
Hematoxylin and eosin (H&E) staining and immunohistochemical examination of GFAP and Nestin were used to assess the tumor type/phenotype. The H&E staining results showed that the cells of the primary tumor and xenograft tumor were polygonal, irregularly arranged, and overlapping and that the nuclei were large, deeply stained, and irregular (Figure 2B). These results indicated that the surgically removed tissue was WHO grade IV glioblastoma. The immunohistochemical assay demonstrated that the tissues were positive for the molecular glioma markers GFAP and Nestin and had up to an 80% proliferation rate according to Ki-67 staining. Compared with the intracranial tumor tissue, the xenograft tumor actively proliferated, and its morphology was consistent with the pathology results for the patient’s resected tissue.
Next, the gene expression profiles of the xenograft tumor and the resected subscalp tissue obtained from the third surgery were determined by transcriptome analysis. The results showed that 9780 genes were upregulated in tumor tissue compared to those in the primary tumor. In addition, 9335 genes were downregulated (R=0.88, Figure 2C). The pie chart shows the enrichment of signal transduction, cell adhesion, and metabolic processes (Figure 2D). These results suggest that the gene expression of the xenograft tumor was slightly different from that of the subscalp tumor.
Phenotypic Characteristics and Genetic Analysis of BT-01
The human glioblastoma cell line BT-01 has now been established from a fifth generation PDX model. The features of BT-01 cells based on light microscopy show that the main body of these cells is spindle-shaped with irregular branches or bifurcations. When the cell density is large, contact growth inhibition is not obvious, and the cells are in a cross-stacked growth state. Under in vitro culturing conditions, BT-01 cells have been cultured for more than 90 passages without morphological changes, and the cell growth curve shows that BT-01 cells have a doubling time of approximately 40 h (Figure 3A). The above results demonstrate that BT-01 cells have malignant potential.
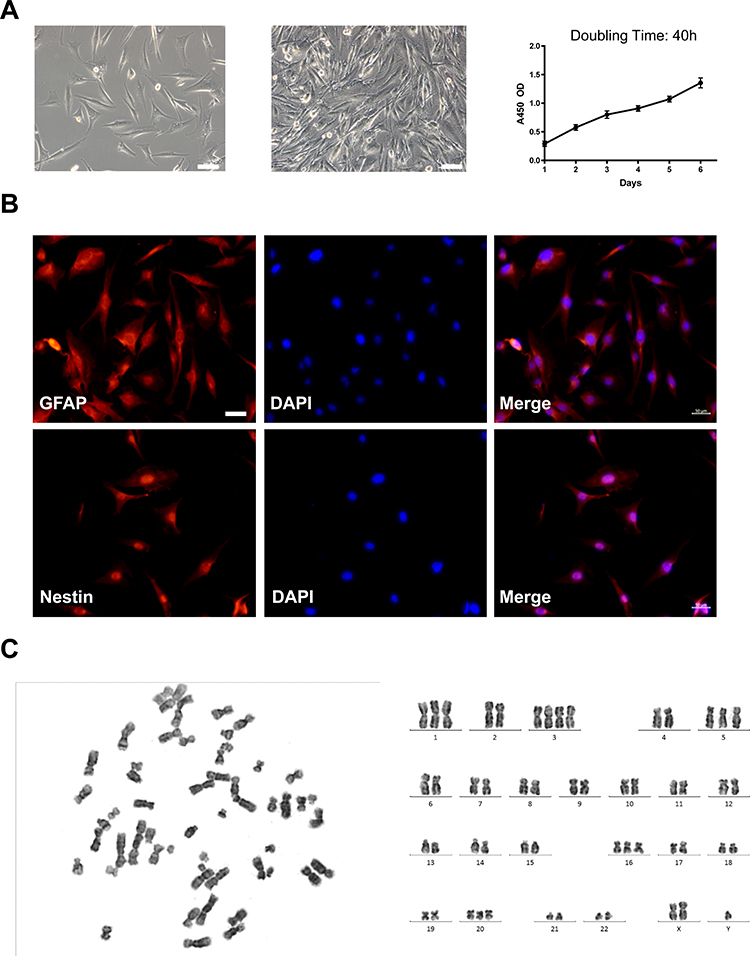 |
Figure 3 Phenotypic characteristics and genetic analysis of BT-01. (A) The BT-01 cell line was established from a fifth-generation PDX tumor. The cells were visualized at low and high density. The proliferation curve of the cells was generated according to a CCK-8 assay. Scale bar represents 100 μm. (B) Immunofluorescence analysis of the glioma marker genes GFAP and Nestin in BT-01 cells. The nuclei were restained with DAPI. Scale bar represents 50 μm. (C) Karyotype analysis and representative karyotype of BT-01 cells. |
To confirm that the cells were of glioblastoma origin, glial cell markers on BT-01 cells were detected with immunofluorescence. Immunofluorescence staining of BT-01 cells showed that the cells were positive for the molecular markers of glioma GFAP and Nestin (Figure 3B). This finding combined with the clinical evidence confirms that BT-01 cells are glioma-derived cells.
Chromosome analysis of BT-01 cells was performed at passages 15 and 50 using Giemsa banding. The results revealed that the karyotype of BT-01 cells was aneuploid (Figure 3C), and the number of chromosomes ranged between 45 and 55 in 70% of cells. Different karyotypes were observed, including +1, +3, +5, +16, +20 and +x. The changes in the number of chromosomes and structural aberrations were severe, which is consistent with the genetic characteristics of human malignant tumors.
The short tandem repeat (STR) profiles of the BT-01 cell line are shown in Table 1. No triallelic phenomenon was observed in each locus in the cell line. No cross-contamination with other human cell lines was found in the BT-01 cell line, and no matching cell lines were found in the ATCC and DSMZ data banks.
BT-01 Cells Maintain High Capacity for Aggressive Growth
To examine the aggressive growth capacity of the BT-01 cell line, invasion and migration assays were performed, and two GBM cell lines, U87-MG and U251-MG, were designated as the positive controls. As seen in Figure 4, the numbers of BT-01 cells that migrated or invaded through the basement membrane were significantly greater than those of U87-MG (P < 0.001) or U251-MG (P < 0.01). Of the three cell lines, BT-01 had the highest migration and invasion capability (Figure 4A and B).
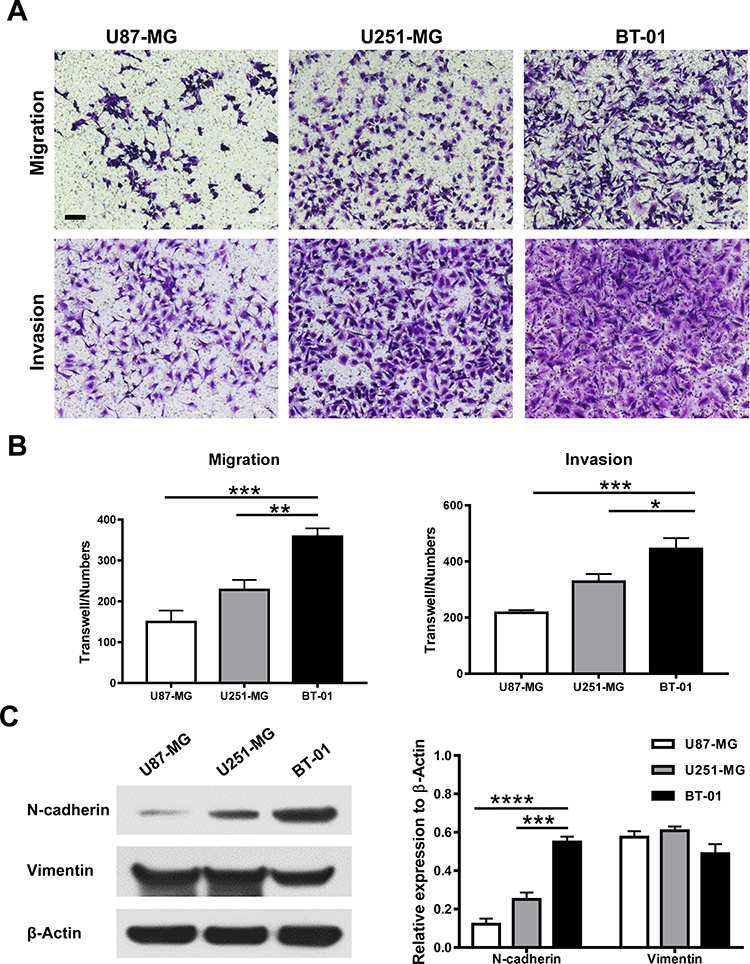 |
Figure 4 The BT-01 cell line maintains high aggressive capacity. (A and B) Transwell assays without or with Matrigel were performed to evaluate the migration ability or invasive ability of the BT-01 cell line or U87-MG or U251-MG cells. Representative images of migrating or invading cells are shown. Scale bar, 100 μm. Data are shown as the means ± s.d from three independent replicates. *P < 0.05, **P < 0.01 and ***P < 0.001. (C) The expression of N-cadherin and Vimentin in U87-MG, U251-MG and BT-01 cells shown by Western blotting. ***P < 0.001 and****P < 0.0001. |
Neuronal cadherin (N-cadherin) is commonly upregulated in the epithelial-to-mesenchymal transition (EMT) and plays a vital role in migration.14 Vimentin is recognized to be an essential protein in tumor EMT and cell invasion and migration by regulating cytoskeletal organization.15 We found the expression of N-cadherin was higher in BT-01 cells compare to U87-MG (P < 0.0001) or U251-MG (P < 0.001) cells (Figure 4C)., and the expression of vimentin is basically consistent with the other two cell lines (P > 0.05). Therefore, BT-01 was identified as a highly aggressive GBM cell line with high migrative and invasive capacity.
The BT-01 Cell Line Harbored More Stem-Like Cells
Glioblastoma stem-like cells in malignant gliomas have been identified in the past decade and are believed to contribute to disease progression and recurrence. Under in vitro culturing conditions, BT-01 cells were found to contain glioblastoma stem-like cells, which could differentiate into adherent glioblastoma cells (Figure 5A). Under the same conditions, BT-01, U87-MG and U251-MG cells were cultured in neural stem cell medium for 72 h, and BT-01 cells were observed to have more and larger neurospheres by microscopy (Figure 5B). Furthermore, the percentage of CD133+ cells in each cell line was analyzed by flow cytometry (Figure 5C). Flow cytometry assays revealed that the proportion of CD133+ cells among BT-01 cells was 1.31%, which was higher than that among U87-MG cells and U251-MG cells and indicated a high self-renewal capability.
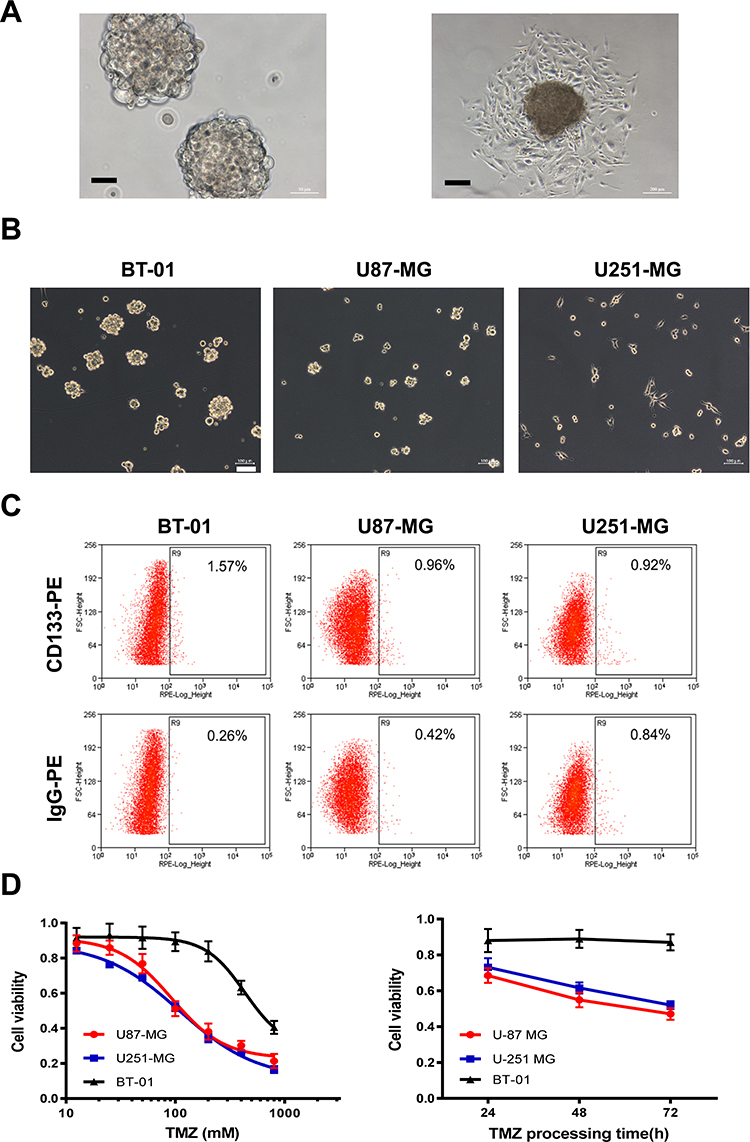 |
Figure 5 The BT-01 cell line harbored more stem-like cells and resisted TMZ. (A) Neurosphere formation of BT-01 cells in neural stem culture medium. Neurospheres formed by BT-01 cells differentiated into adherent cells in complete medium. Scale bar, 200 μm. (B) Neurosphere formation of BT-01 cells, U87-MG cells and U251-MG cells for 72 h. Scale bar, 100 μm. (C) Numbers of CD133+ GSCs among BT-01 cells, U87-MG cells and U251-MG cells. (D) IC50 of TMZ in BT-01 cells, U87-MG cells and U251-MG cells and the viability of BT-01 cells, U87-MG cells and U251-MG cells treated with 100 μM TMZ. |
To find more effective chemotherapy regimens for recurrent glioblastomas, glioma cells (U87-MG and U251-MG) were used as a reference for comparison with BT-01 cells to determine the sensitivity of BT-01 cells to the chemotherapeutic drug temozolomide (TMZ). The results showed that the IC50 of TMZ in U87-MG and U251-MG cells was 92.41 μM and 109.9 μM, respectively, and that the inhibitory effect of TMZ was enhanced over time (Figure 5D). In contrast, the IC50 of TMZ in BT-01 cells derived from recurrent glioblastoma was 431.9 μM, and the inhibitory effect of TMZ was not obviously enhanced over time, which indicates that BT-01 cells are more resistant to the chemotherapy drug TMZ than U87-MG or U251-MG cells.
To explain the detailed molecular mechanism, additional molecular pathological diagnostic examinations were performed to check the IDH1/IDH2 and MGMT statuses of the tumor and the cells. The molecular examination results for the tumor tissue and the BT-01 cells indicated that both had wildtype IDH and were MGMT promoter methylation-negative (Table 2). Therefore, a 37-year-old man with recurrent GBM, wildtype IDH and no MGMT promoter methylation was identified.
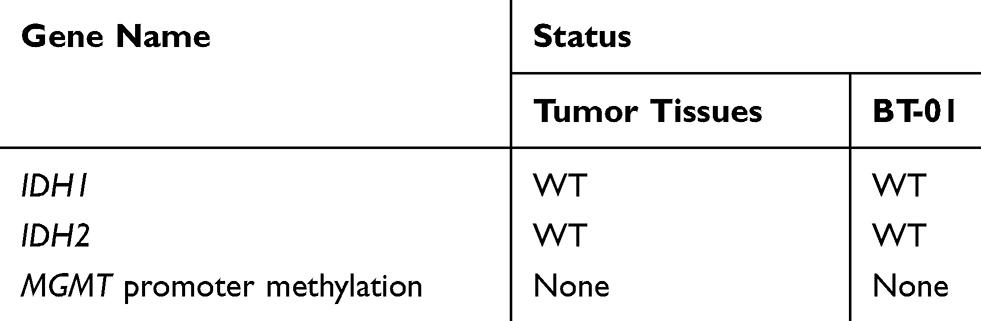 |
Table 2 Molecular Pathological Examination of Tumor Tissues and BT-01 Cells |
BT-01 Was Sensitive to oHSV-1
To ensure that oHSV-1 can infect and spread in glioma cells, we constructed oHSV-GFP, which expressed the fluorescent reporter GFP (Figure 6A). oHSV-GFP could infect and spread between the glioma cells at different multiplicities of infection (MOIs) (Figure 6B). oHSV-1 could replicate in and repress the growth of glioma cells.
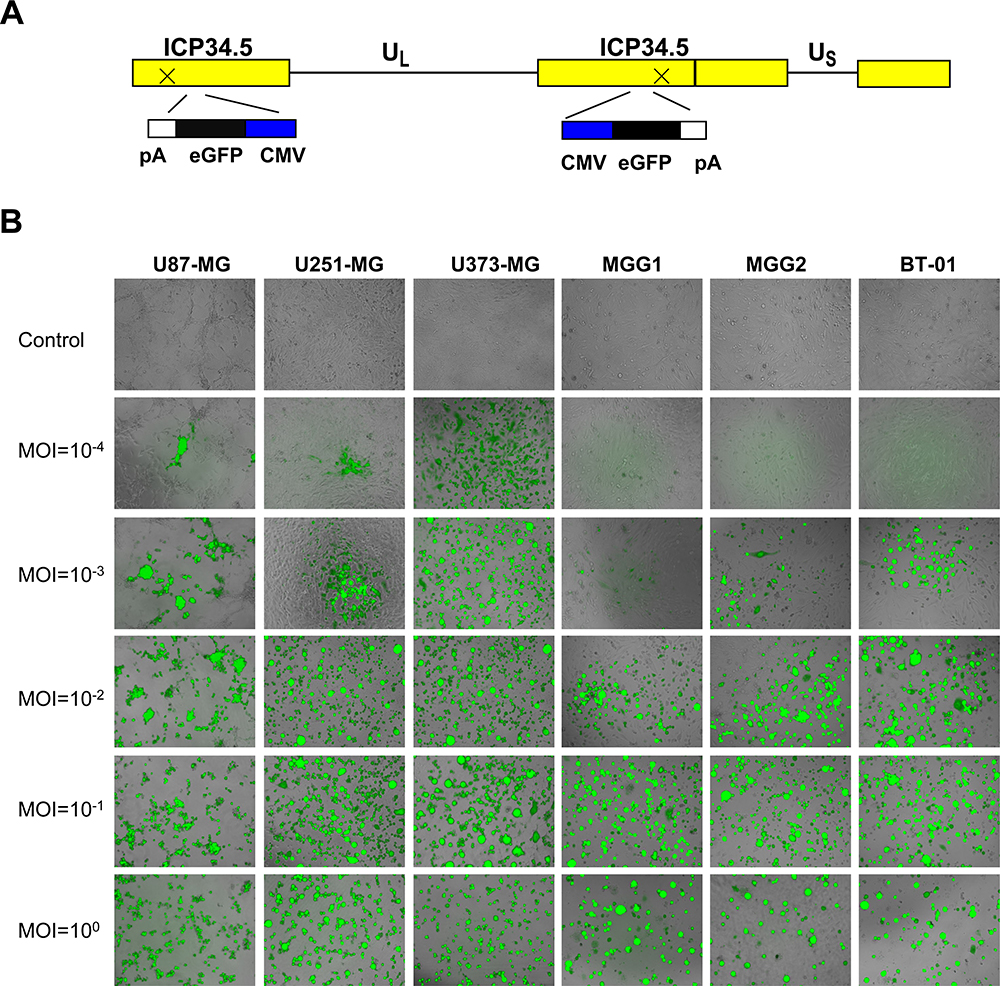 |
Figure 6 BT-01 was sensitive to oHSV-1. (A) Schematic of oHSV-GFP. Both copies of the γ34.5 gene were silenced by inserting GFP. UL and US represent the long and short unique sequences, respectively. (B) Three cell lines (U87-MG, U251-MG and U373-MG), two primary cell types (MGG1 and MGG2) and BT-01 were infected with oHSV-GFP at different MOIs (10−4–100). Representative fluorescence microscopy images were obtained at 48 h. |
oHSV-1 Inhibits Tumor Growth in the Glioblastoma PDX Model
To evaluate the antitumor efficacy of oHSV-1 in vivo, PDX models were treated with intratumoral injection of oHSV-1 or PBS on days 6 and 10 after tumor implantation (Figure 7A). oHSV-1 significantly inhibited tumor growth (P < 0.01, Figure 7B–D). Body weight loss or other adverse events was not observed. This indicates that the glioblastoma PDX model is convenient for evaluating the efficacy of oncolytic viruses in preclinical studies.
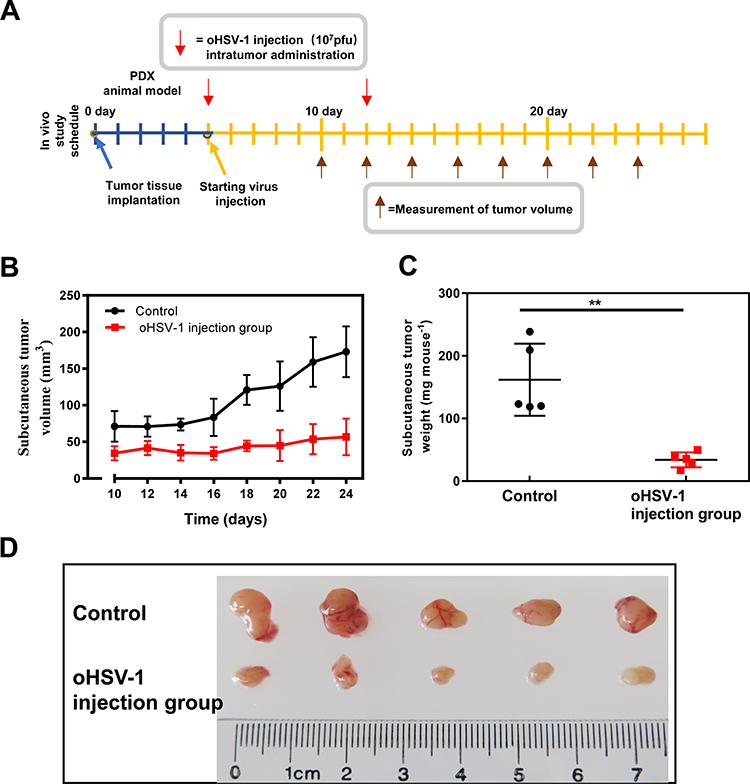 |
Figure 7 oHSV-1 inhibits tumor growth in glioblastoma PDXs. (A) Schematic diagram showing the treatment of glioblastoma PDX with oHSV-1. oHSV-1 injection was performed once a week after tumor tissue implantation. (B and C) The tumor volume was measured (B) and compared with the body weight (C). **P < 0.01. (D) Photograph of the subcutaneous tumor with or without oHSV-1 injection. |
Discussion
Glioblastoma is the most aggressive primary brain tumor, and almost all patients experience relapse after surgical treatment.16 Compared with the primary tumors, recurrent glioblastomas tend to be more aggressive and therapeutically resistant, which makes them more difficult to treat. Currently, no effective treatment exists for glioblastoma, and thus, patients have a poor prognosis.17 Recently, high inter- and intratumoral heterogeneity of glioblastoma was illustrated using single-cell RNA sequencing.18–20 It has been increasingly appreciated that heterogeneity within glioblastoma contributes to poor outcomes and recurrence. However, only one human recurrent glioblastoma cell line was identified in the ATCC and DSMZ databases, which has severely restricted basic and clinical studies of recurrent glioblastoma. The establishment of a new recurrent glioblastoma cell line is of great significance for exploring glioblastoma invasion, recurrence and drug resistance.
GBM invaded within the neuroaxis easily by way of cerebrospinal fluid. In addition, the proportion of patients with glioblastoma with extracranial recurrence is only 0.4%-2%, but the prognosis of the kind of patients is very poor.3–5 GBM invades mainly via vascular invasion, cranial nerve perineural spread, and lymphatic spread.4 Highly predisposing factors for GBM extracranial invasion consist of a long survival time and iatrogenic procedures.
According to MR imaging, we confirmed that intracranial tumors and extracranial tumors were hardly contiguous and not caused by direct tumor growth from the skull defect. We suppose that the exterior tumor probably invades through venous invasion during recurrence. Of course, the possibility of scalp transplantation due to iatrogenic procedures cannot be ruled out. HE staining showed that the extracranial tumor was similar to the intracranial tumor in terms of its histological features. The results of the comparative transcriptome analysis of the recurrent tumor and the PDX tumor showed that their expression profiles were not completely consistent (Figure 2B and C). Aberrations in signaling pathways, including pathways involved in signal transduction, cell adhesion and invasive processes, indicate the aggressive potential.
Karyotype analysis showed that BT-01 cells were derived from a human male; the cell karyotypes were aneuploid and were mostly between subdiploid and triploid, with abnormalities in the chromosome number and structure. The abnormalities in the chromosomes that carry the genetic information initiate the cancer program, induce cell carcinogenesis and cause tumorigenesis. Cross-contamination of cell lines has led to tens of thousands of invalid studies.21 To avoid wasting a large amount of resources, such as human, material and financial resources, STR genotyping has been applied to identify cells by ICLAC, ATCC and other authorities and has become the gold standard.22 STR analysis of BT-01 cells showed no cross-contamination with other human cell lines, and no matching cell lines were found in either the ATCC or DSMZ data banks. Therefore, we generated a reliable tumor cell line for cancer research.
Tumorigenicity in vivo is another major feature of cancer cell. BT-01 cells were inoculated subcutaneously into nude mice and formed tumors (data not shown). Since BT-01 cells were derived from a PDX model, we further verified that BT-01 cells originated from a glioblastoma. Through the detection of molecular markers of glioma, BT-01 cells and nude mouse xenografts were shown to express GFAP and Nestin. These findings combined with the clinical data of patients revealed that BT-01 was a new glioblastoma cell line.
Glioblastoma stem-like cells were found among BT-01 cells by forming neutrospheres after culture in neural stem cell media. Flow cytometry analysis showed that the proportion of glioblastoma stem-like cells in BT-01 cells was higher than that in the classical glioma cell lines U87-MG and U251-MG. The drug sensitivity test found that BT-01 cells were more tolerant to the chemotherapeutic drug temozolomide than U87-MG and U251-MG cells. Previous studies have shown the existence of a subpopulation of glioblastoma stem cells in human glioblastoma.23 GSCs are closely associated with tumorigenesis, tumor maintenance, invasion and recurrence.24,25 The presence of GSCs leads to resistance to chemoradiotherapy,26 and the proportion of GSCs in recurrent glioblastoma is increased.27 Therefore, the presence of GSCs in BT-01 cells may have contributed to the invasion, recurrence and therapeutic tolerance of glioblastoma in this patient. Glioblastoma stem-like cells may therefore become a new target that can be used to find more effective treatments for glioblastoma, which is fatal.
IDH mutations and MGMT promoter methylation are reliable glioma biomarkers and prognostic factors used in the clinic. More than 80% of low-grade gliomas and recurrent GBM were found to have IDH mutations.
Such genetic alterations were recognized as good prognostic markers for GBM and were associated with increased overall survival.28 MGMT methylation is used to predict the chemotherapy response in glioblastoma patients, and wild-type MGMT is linked to TMZ resistance. TMZ was the standard chemotherapy used to treat GBM for several years.29 Consistent with these results, our observations indicated that wild-type MGMT confers TMZ resistance in BT-01 cells. In the mouse subcutaneous xenograft model, TMZ showed a certain therapeutic effect (data not shown). Considering that the mouse model was not an intracranial model, TMZ could reach a higher concentration in the subcutaneous tissue than in the brain because of the blood brain barrier. TMZ not only causes DNA damage but can also induce autophagy and promote apoptosis.30 Meanwhile, multiple proteins, such as DHFR/TYMS,31 ANGPTL432 and SNAP,33 can regulate chemosensitivity to TMZ. The mechanism of TMZ resistance involves a complex pathway regulated by multiple factors, and the underlying mechanism needs further elucidation. The cell lines we established provide a reliable clinical model for studying the mechanism of TMZ resistance. We will use BT-01 cells as a preclinical model to study the detailed regulatory mechanism of TMZ resistance.
Oncolytic herpes simplex virus 1 (oHSV-1) can be used to treat various malignancies. T-VEC, which only replicates in tumor cells and expresses granulocyte monocyte colony stimulating factor (GM-CSF), is the first virus immunotherapy that was approved to treat nonresectable melanoma.34 The recombinant oHSV-1 used in this study shows high infectivity and a high replication rate in glioma cells. oHSV-1 was revealed to be an effective treatment against glioblastoma in vivo. In addition, we will check the antitumor efficacy of oHSV-1 in our further clinical trials.
In conclusion, we established and characterized the extracranial recurrence glioblastoma cell line BT-01 derived from the PDX model. This newly established cell line was examined by immunofluorescence staining, karyotypic analysis and short tandem repeat (STR) experiments. The main characteristics of BT-01 are that it harbors an abundance of glioblastoma stem-like cells (GSCs), possesses highly invasive capacity and is tolerant to the chemotherapeutic drug temozolomide (TMZ) compared with existing cell lines. BT-01 cells and PDX models responded to oHSV-1. We believe that BT-01 and PDX could serve as preclinical models of GBM and could be employed to test the antitumor efficacy of oHSV-1 in vitro and in vivo.
Ethical Statement
All PDX experiments were approved by the Ethical Committee of Beijing Tiantan Hospital. We confirmed that written informed consent has been provided by the patient to have the case details and any accompanying images published. Informed consent for publication was obtained from the patient according to the guidelines of the Declaration of Helsinki and its later amendments.
Animal studies were reviewed and approved by the Experimental Animal Ethical Committee of the Beijing Neurosurgical Institute. All mice received care according to the guidelines of the Guide for the Care and Use of Laboratory Animals.
Disclosure
The authors have no conflicts of interest to declare.
References
1. Wen PY, Reardon DA. Neuro-oncology in 2015: progress in glioma diagnosis, classification and treatment. Nat Rev Neurol. 2016;12:69–70. doi:10.1038/nrneurol.2015.242
2. Kim J, Lee IH, Cho HJ, et al. Spatiotemporal evolution of the primary glioblastoma genome. Cancer Cell. 2015;28:318–328. doi:10.1016/j.ccell.2015.07.013
3. Schonsteiner SS, Bommer M, Haenle MM, et al. Rare phenomenon: liver metastases from glioblastoma multiforme. J Clin Oncol. 2011;29:e668–671. doi:10.1200/JCO.2011.35.9232
4. Hamilton JD, Rapp M, Schneiderhan T, et al. Glioblastoma multiforme metastasis outside the CNS: three case reports and possible mechanisms of escape. J Clin Oncol. 2014;32:e80–84. doi:10.1200/jco.2013.48.7546
5. Simonetti G, Silvani A, Fariselli L, et al. Extra central nervous system metastases from glioblastoma: a new possible trigger event? Neurol Sci. 2017;38:1873–1875. doi:10.1007/s10072-017-3036-0
6. Brandes AA, Tosoni A, Franceschi E, et al. Recurrence pattern after temozolomide concomitant with and adjuvant to radiotherapy in newly diagnosed patients with glioblastoma: correlation with MGMT promoter methylation status. J Clin Oncol. 2009;27:1275–1279. doi:10.1200/JCO.2008.19.4969
7. Stupp R, Mason WP, van den Bent MJ, et al. Radiotherapy plus concomitant and adjuvant temozolomide for glioblastoma. N Engl J Med. 2005;352:987–996. doi:10.1056/NEJMoa043330
8. Fukuhara H, Ino Y, Todo T. Oncolytic virus therapy: a new era of cancer treatment at dawn. Cancer Sci. 2016;107:1373–1379. doi:10.1111/cas.13027
9. Nakashima H, Chiocca EA. Modification of HSV-1 to an oncolytic virus. Methods Mol Biol. 2014;1144:117–127. doi:10.1007/978-1-4939-0428-0_8
10. Streby KA, Geller JI, Currier MA, et al. Intratumoral injection of HSV1716, an oncolytic herpes virus, is safe and shows evidence of immune response and viral replication in young cancer patients. Clin Cancer Res. 2017;23:3566–3574. doi:10.1158/1078-0432.Ccr-16-2900
11. Haverty PM, Lin E, Tan J, et al. Reproducible pharmacogenomic profiling of cancer cell line panels. Nature. 2016;533:333–337. doi:10.1038/nature17987
12. Ogawa J, Pao GM, Shokhirev MN, Verma IM. Glioblastoma model using human cerebral organoids. Cell Rep. 2018;23:1220–1229. doi:10.1016/j.celrep.2018.03.105
13. Zhou Y, Jin G, Mi R, Dong C, Zhang J, Liu F. The methylation status of the platelet-derived growth factor-B gene promoter and its regulation of cellular proliferation following folate treatment in human glioma cells. Brain Res. 2014;1556:57–66. doi:10.1016/j.brainres.2014.01.045
14. Mrozik KM, Blaschuk OW, Cheong CM, Zannettino ACW, Vandyke K. N-cadherin in cancer metastasis, its emerging role in haematological malignancies and potential as a therapeutic target in cancer. BMC Cancer. 2018;18:939. doi:10.1186/s12885-018-4845-0
15. Satelli A, Li S. Vimentin in cancer and its potential as a molecular target for cancer therapy. Cell Mol Life Sci. 2011;68:3033–3046. doi:10.1007/s00018-011-0735-1
16. Seystahl K, Wick W, Weller M. Therapeutic options in recurrent glioblastoma–an update. Crit Rev Oncol Hematol. 2016;99:389–408. doi:10.1016/j.critrevonc.2016.01.018
17. Hersh DS, Harder BG, Roos A, et al. The TNF receptor family member Fn14 is highly expressed in recurrent glioblastoma and in GBM patient-derived xenografts with acquired temozolomide resistance. Neuro Oncol. 2018;20:1321–1330. doi:10.1093/neuonc/noy063
18. Wang L, Babikir H, Müller S, et al. The phenotypes of proliferating glioblastoma cells reside on a single axis of variation. Cancer Discov. 2019;9:1708–1719. doi:10.1158/2159-8290.Cd-19-0329
19. Neftel C, Laffy J, Filbin MG, et al. An integrative model of cellular states, plasticity, and genetics for glioblastoma. Cell. 2019;178:835–849. doi:10.1016/j.cell.2019.06.024
20. Darmanis S, Sloan SA, Croote D, et al. Single-cell RNA-seq analysis of infiltrating neoplastic cells at the migrating front of human glioblastoma. Cell Rep. 2017;21:1399–1410. doi:10.1016/j.celrep.2017.10.030
21. Vaughan L, Glänzel W, Korch C, Capes-Davis A. Widespread use of misidentified cell line KB (HeLa): incorrect attribution and its impact revealed through mining the scientific literature. Cancer Res. 2017;77:2784–2788. doi:10.1158/0008-5472.Can-16-2258
22. American Type Culture Collection Standards Development Organization Workgroup ASN. Cell line misidentification: the beginning of the end. Nat Rev Cancer. 2010;10:441–448. doi:10.1038/nrc2852
23. Lathia JD, Mack SC, Mulkearns-Hubert EE, Valentim CL, Rich JN. Cancer stem cells in glioblastoma. Genes Dev. 2015;29:1203–1217. doi:10.1101/gad.261982.115
24. Chen J, Li Y, Yu T-S, et al. A restricted cell population propagates glioblastoma growth after chemotherapy. Nature. 2012;488:522–526. doi:10.1038/nature11287
25. Jackson M, Hassiotou F, Nowak A. Glioblastoma stem-like cells: at the root of tumor recurrence and a therapeutic target. Carcinogenesis. 2015;36:177–185. doi:10.1093/carcin/bgu243
26. D’Alessandris QG, Biffoni M, Martini M, et al. The clinical value of patient-derived glioblastoma tumorspheres in predicting treatment response. Neuro Oncol. 2017;19:1097–1108. doi:10.1093/neuonc/now304
27. Tamura K, Aoyagi M, Ando N, et al. Expansion of CD133-positive glioma cells in recurrent de novo glioblastomas after radiotherapy and chemotherapy. J Neurosurg. 2013;119:1145–1155. doi:10.3171/2013.7.Jns122417
28. Hata N, Hatae R, Yoshimoto K, et al. Insular primary glioblastomas with IDH mutations: clinical and biological specificities. Neuropathology. 2017;37:200–206. doi:10.1111/neup.12362
29. Chen X, Zhang M, Gan H, et al. A novel enhancer regulates MGMT expression and promotes temozolomide resistance in glioblastoma. Nat Commun. 2018;9:2949. doi:10.1038/s41467-018-05373-4
30. Harder BG, Peng S, Sereduk CP, et al. Inhibition of phosphatidylinositol 3-kinase by PX-866 suppresses temozolomide-induced autophagy and promotes apoptosis in glioblastoma cells. Mol Med. 2019;25:49. doi:10.1186/s10020-019-0116-z
31. Zhao M, Tan B, Dai X, et al. DHFR/TYMS are positive regulators of glioma cell growth and modulate chemo-sensitivity to temozolomide. Eur J Pharmacol. 2019;863:172665. doi:10.1016/j.ejphar.2019.172665
32. Tsai YT, Wu AC, Yang WB, et al. ANGPTL4 induces TMZ resistance of glioblastoma by promoting cancer stemness enrichment via the EGFR/AKT/4E-BP1 cascade. Int J Mol Sci. 2019;20:5625. doi:10.3390/ijms20225625
33. Tsai CK, Huang LC, Wu YP, Kan IY, Hueng DY. SNAP reverses temozolomide resistance in human glioblastoma multiforme cells through down-regulation of MGMT. FASEB J. 2019;33:14171–14184. doi:10.1096/fj.201901021RR
34. Andtbacka RH, Kaufman HL, Collichio F, et al. Talimogene laherparepvec improves durable response rate in patients with advanced melanoma. J Clin Oncol. 2015;33:2780–2788. doi:10.1200/JCO.2014.58.3377
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.