Back to Journals » Neuropsychiatric Disease and Treatment » Volume 21
Bifidobacterium adolescentis DM8504 Alleviates Autistic-Like Behaviors in Valproic Acid-Exposed Rats Through Gut Microbiota Modulation and SCFA Restoration
Authors Liu H, Liu G, Zhang Y ![]() , Suo W, Hao Y, Wang Y
, Suo W, Hao Y, Wang Y ![]() , Ding H
, Ding H
Received 23 June 2025
Accepted for publication 21 October 2025
Published 7 November 2025 Volume 2025:21 Pages 2449—2463
DOI https://doi.org/10.2147/NDT.S547997
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Taro Kishi
Hui Liu,1,* Gaojie Liu,2,* Yuxuan Zhang,1,* Weicai Suo,1 Yan Hao,2 Yizhong Wang,2,3 Hongfang Ding1
1Department of Pediatrics, Shengli Oil Field Central Hospital, Binzhou Medical University, Dongying, Shandong, People’s Republic of China; 2Gut Microbiota and Metabolic Research Center, Institute of Pediatric Infection, Immunity and Critical Care Medicine, School of Medicine, Shanghai Jiao Tong University, Shanghai, People’s Republic of China; 3Department of Gastroenterology, Hepatology, and Nutrition, Shanghai Children’s Hospital, School of Medicine, Shanghai Jiao Tong University, Shanghai, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Yizhong Wang, Department of Gastroenterology, Hepatology and Nutrition, Shanghai Children’s Hospital, School of Medicine, Shanghai Jiao Tong University, No. 355 Luding Road, Shanghai, 200062, People’s Republic of China, Email [email protected] Hongfang Ding, Department of Pediatrics, Shengli Oil Field Central Hospital, Binzhou Medical University, No. 31 Jinan Road, Dongying District, Dongying, Shandong, 257034, People’s Republic of China, Email [email protected]
Objective: Compelling evidence has confirmed that gut microbiota dysbiosis is involved in the development of autism spectrum disorder (ASD). Microbial-based therapies, including probiotics, may provide novel options for ASD management. This study aimed to investigate the alleviative effect of a probiotic strain, Bifidobacterium adolescentis (B. adolescentis) DM8504, on autistic-like behaviors in rats exposed to valproic acid (VPA).
Methods: Male offspring of VPA-exposed pregnant Sprague–Dawley rats treated with B. adolescentis DM8504 were subjected to behavioral tests. Fecal short-chain fatty acids (SCFAs) and microbiota composition were determined by targeted metabolomics and 16S rRNA gene sequencing, respectively. Microbial functional profiles were analyzed using the KEGG and COG pathway analyses.
Results: B. adolescentis DM8504 alleviated autistic-like behaviors in VPA-exposed rats, as evidenced by enhanced locomotor activity, exploratory behavior, sociability, spatial working memory, and depression relief. B. adolescentis DM8504 treatment significantly enhanced the fecal levels of acetic acid, butyric acid, isobutyric acid, propionic acid, and hexanoic acid, and restored the diversity of gut microbiota composition in VPA-exposed rats. Specifically, B. adolescentis DM8504 increased the abundance of gut bacterial species capable of producing SCFAs in VPA-exposed rats, including Bifidobacterium, Faecalibacterium, Eubacterium, and Lachnospiraceae_ NK4A136_group. In addition, microbial functional profile analysis showed that B. adolescentis DM8504 administration reversed the alterations in COG and KEGG pathways induced by VPA exposure.
Conclusion: These findings reveal that B. adolescentis DM8504 alleviates autistic-like behaviors in VPA-exposed rats through restoration of SCFA levels and enrichment of SCFA-producing bacteria, which indicate that well-defined B. adolescentis strains may have potential for the management of children with ASD in the future.
Keywords: ASD, Bifidobacterium adolescentis, probiotic, SCFAs, gut microbiota
Introduction
Autism spectrum disorder (ASD), first described by Dr. Leo Kanner in 1943, is a broad spectrum of neurodevelopmental disorders that are mainly characterized by impaired social communication skills, restricted interests, and repetitive stereotyped behaviors.1 ASD symptoms typically begin in young children before three years of age and usually persist throughout the whole lifespan.2 The incidence of ASD has increased rapidly due to exacerbated etiological factors and improved awareness and diagnostic technologies in recent decades.3,4 The Global Burden of Diseases, Injuries, and Risk Factors Study across 204 countries estimated that, in 2019, the global prevalence of ASD was 0.37%, and over 28 million people were affected by ASD globally.4 The most recent report from the Autism and Developmental Disabilities Monitoring Network showed that overall ASD prevalence in the United States is 27.6 per 1000 children aged 8 years, and there are 3.8 times more boys affected than girls.5 The etiology of ASD is very complex and remains to be fully elucidated. Current evidence indicates that both genetic and non-genetic factors are involved in the pathogenesis of ASD, including genetic vulnerabilities; neurological, immunological, and environmental risk factors; and gut microbiota dysbiosis.1,6
Recent studies have shown that gut microbiota and their metabolites play critical roles in the gut–brain axis and are involved in the development of central nervous system (CNS) dysfunctions and disorders.7–9 Gastrointestinal (GI) dysfunctions are common in patients with ASD, and occur in more than 40% of affected individuals.10 In comparison with neurotypical children, an increase in the number of Bacteroidetes and a decrease in Verrucomicrobia were observed in patients with ASD.11 Levels of genera Bacteroides, Parabacteroides, Clostridium, and Phascolarctoba cterium were significantly higher in children with ASD, accompanied by a decrease in Lactobacillus, Coprococcus, and Bifidobacterium species.8 Our previous study also revealed a lower relative abundance of Bifidobacterium in children with ASD than in healthy subjects.12 Gut microbiota–brain interactions are predominantly mediated by microbiota-derived metabolites, including neurotransmitters, short chain fatty acids (SCFAs), amino acids, and immune modulators.9 These active molecules can cross the blood–gut and the blood–brain barriers (BBBs) to influence brain function.9 Previous studies have reported altered fecal and blood levels of these compounds in patients with ASD, such as glutamate, SCFAs, serotonin, dopamine, tryptophan, and indoles.13–17 However, the data are still inconsistent. Further studies are needed to confirm the characteristic gut microbiota and metabolites associated with ASD.
Currently, there is no curative therapy available for the core symptoms of ASD. The primary treatment is intensive behavioral intervention in early childhood to strengthen social skills and reduce problem behaviors in affected individuals.18 Several studies have shown that sulforaphane, oxytocin, or arginine-vasopressin are helpful in improving the general condition of ASD patients.19–21 Since compelling evidence indicates that gut microbiota dysbiosis is involved in the pathogenesis of ASD, a number of studies have been performed to investigate the potential impact of microbial-based and microbial-targeted therapies for the management of ASD patients.13 Preclinical studies conducted in animal models using probiotics and fecal microbiota transplantation (FMT) have been shown to alleviate autistic-like behaviors. For example, Bacteroides fragilis relieved autistic-like behaviors by decreasing gut permeability and correcting gut microbiota dysbiosis in an ASD mouse model, and Lactobacillus alleviated autistic-like behaviors by ameliorating 5-hydroxytryptamine metabolic disorder in valproic acid (VPA)-exposed rats.7,22 Furthermore, a growing number of clinical trials have demonstrated that FMT, antibiotics, probiotics, and synbiotics can improve GI dysfunction and core symptoms in patients with ASD.13,23 Our recent studies also showed that FMT treatment improved core symptoms, GI symptoms, and sleep disturbance in children with ASD.24,25 Thus, well-defined microbial-based therapies for restoring gut microbial balance may provide novel options for ASD therapy in the future.
As an abundant bifidobacterial species in the human gut, Bifidobacterium adolescentis (B. adolescentis) can metabolize resistant starch to produce SCFAs (eg, acetate) and promote the growth of commensal bacteria by cross-feeding.26 Multiple B. adolescentis strains have been shown to have health-beneficial properties in vitro and in animal models through improving gut microbiota dysbiosis and intestinal barrier function, regulating anti-inflammatory and immune functions, and leading to the production of neurotransmitters.26–28 For example, B. adolescentis NK98 could alleviate Escherichia coli-induced depression and gut dysbiosis in mice.29 Dietary supplementation of B. adolescentis ATCC 15703 improved neurodegeneration in a mouse model of premature aging.27 In comparison with other B. adolescentis strains, B. adolescentis DM8504 is a novel strain isolated from the stool of a healthy Chinese adult, and has been demonstrated to be effective in management of human gastrointestinal (GI) disorders by regulating the intestinal immunity and gut microbiota. However, the effect of B. adolescentis DM8504 on the regulation of gut–brain axis remains unclear.
In this study, we aimed to investigate the alleviative effect of the probiotic strain B. adolescentis DM8504 on autistic-like behaviors in a VPA-exposed rat model. We further examined the effects of B. adolescentis DM8504 on fecal SCFAs levels, gut microbiota composition and functional changes. In order to explore the effect of B. adolescentis DM8504 on fetal neurodevelopment by regulating maternal microbiota, pregnant rats were administered live B. adolescentis DM8504 solution by oral gavage during the whole gestational period. The results revealed that B. adolescentis DM8504 alleviated autistic-like behaviors in VPA-exposed rats. The mechanisms studies showed that B. adolescentis DM8504 enhanced the fecal levels of SCFAs by restoring the abundance of gut bacterial species capable of producing SCFAs in VPA-exposed rats. Our findings suggest that well-defined B. adolescentis strains may have potential for the management of children with ASD in the future.
Materials and Methods
B. adolescentis DM8504 Preparation
B. adolescentis DM8504 isolated from the stool of a healthy adult was recovered from Live Bifidobacterium Capsules (Livzon, Zhuhai, China) on a plate containing trypticase peptone yeast extract (TPY) medium (Hope Bio-Technology, Qingdao, China) with agar under anaerobic conditions at 37°C for 48 h. A single colony was selected from the plate and amplified anaerobically in TPY broth medium at 37°C for 48 h. Bacterial cells were collected by centrifugation at 1500 × g and 4°C for 10 min. After washing three times with sterile phosphate-buffered saline (PBS), the bacterial cells were re-suspended in sterile PBS to a concentration of 2.5×109 CFU/mL.
Animals, Probiotic Administration, and VPA Exposure
Sprague–Dawley (SD) rats (10–12 weeks old, body weight: 280–300 g for males [n = 10] and 230–260 g for females [n = 10]) were purchased from Septe Testing Service Co. Ltd. (Yantai, Shandong, China) and housed in the experimental animal center of Binzhou Medical University under standard laboratory conditions (room temperature, 20–26°C; relative humidity, 40–70%; light cycle, 12/12 h light/dark), with free access to food and water. All animal experimental procedures were approved by the Animal Ethics Committee of Binzhou Medical University (2025-310). The rats were mated overnight at a male: female ratio of 2:1 after acclimatization for one week, and the first gestational day (GD1) was recorded by observing a vaginal plug the next morning. Pregnant rats were housed in individual cages and randomly divided into a control group (n = 4) and a VPA-exposed group (n = 6). Pregnant rats in the VPA-exposed group were treated with a single intraperitoneal injection of VPA sodium salt (Sigma-Aldrich, St. Louis, USA, dissolved in 0.9% saline at a concentration of 250 mg/mL) at 600 mg/kg on GD12.5, whereas pregnant rats in the control group received a single intraperitoneal injection of the same volume of 0.9% saline on the same day. For probiotic treatment, pregnant rats in the VPA-exposed group were further randomly separated into two subgroups: VPA and VPA + probiotic groups. Pregnant rats in the VPA + probiotic group (n = 3) were administered 400 μL B. adolescentis DM8504 resuspension (1×109 CFU) by oral gavage once daily from GD1 until birth (GD21-23); pregnant rats in the VPA group (n = 3) and control group (n = 4) were administered 400 μL sterile PBS by oral gavage once daily instead of B. adolescentis DM8504. Their offspring were designated as VPA + DM8504, VPA, and control (Ctrl) groups. After natural weaning on postnatal day 21 (P21), eight male offspring were randomly selected from each group for subsequent behavioral tests in a blinded manner. The experimental animal scheme is shown in Figure 1.
 |
Figure 1 The animal experimental scheme. VPA was injected at GD12.5 and B. adolescentis DM8504 was orally gavaged from GD1 until birth. Behavioral tests were conducted during P28 to P35. Abbreviations: VPA, valproic acid; P, postnatal; GD, gestational day. |
Postnatal Behavioral Tests
Open-Field Test
An open-field box (40 × 40 × 40 cm) with black inner walls and a white bottom was divided into 16 equal parts; the central four parts were set as the central area, while the other 12 parts were regarded as the peripheral area. A camera was placed above the box to record movement in time spent and distance in the peripheral and central areas. Juvenile rats (P28) were habituated to the test room 1 h prior to testing. Each rat was sequentially placed individually in the central area for a 10-min open-field test. The time spent in the central area, number of entries into the central area, and total distance traveled were evaluated using EthoVision software (Noldus Information Technology, Leesburg, VA, USA).
Three-Chamber Test
A three-chamber apparatus with dimensions of 60 cm × 60 cm × 22 cm (length × width × height) was equally divided into three rectangular chambers, and each chamber was separated using transparent plexiglass with a channel in the middle. An identical cage was placed in each of the side chambers. One day before the test, all juvenile rats (P29) were individually acclimated for 5 min in a three-chamber apparatus. On the day of the test, the test rats (P30) were placed in the middle chamber for 5 min to adapt to the middle chamber by closing the chambers on both sides. A stranger normal male rat was placed in one of the side cages and was defined as a social zone, whereas the chamber remained with an empty cage set as a non-social zone. Subsequently, the left and right chambers were opened and the test rats were given free access to explore the three chambers for 10 min from the middle chamber. In session 2 of the test, another unfamiliar normal male rat was introduced to the empty case, regarded as a stranger, while the previous unfamiliar rat was regarded as familiar, and the test rat (P31) was allowed to freely explore the familiar and strange rats starting from the middle chamber for 10 min. The behavior of the test rats was recorded using a video camera and the apparatus was cleaned with 75% alcohol after each test. The time spent in both side chambers was evaluated using EthoVision software. The social preference indexes of sessions 1 and 2 were calculated as time spent in the stranger chamber, empty chamber, novel chamber, or familiar chamber, respectively.
Y-Maze Test
A plexiglass Y-maze apparatus consisting of three equally spaced arms (120°) measuring 40 cm × 10 cm × 30 cm (length × width × height) was used for the Y-maze test. All rats were habituated for 1 h in the test room before the Y-maze test. A test rat (P32) was placed in the center of the Y-maze and allowed to move freely between the three arms for 10 min without reinforcement. A valid entry was considered to be when a rat’s body and tail entered an arm completely, and a spontaneous alternation was defined as when a rat entered all three arms consecutively. The percentage of spontaneous alternations was calculated as [(number of alternations)/(total number of arm entries − 2)] × 100.30 For the acquisition test, a rat was placed at the center of the Y-maze and allowed to explore the apparatus for 10 min by closing one of the arms (novel arm); it was then returned to its home cage until the retrieval test. After 24 h, the test rat (P33) was placed at the center of the apparatus and allowed to freely explore all three arms for 5 min. The duration of visits to the novel arm was calculated as a percentage of the time spent in the novel arm/the total time spent in all three arms.31 The Y-maze apparatus was cleaned with 75% alcohol and allowed to dry between sessions. The behavior of the test rat was recorded using a video camera and evaluated using EthoVision software.
Forced Swim Test
The forced swim test was conducted to evaluate depressive-like behavior in rats.32 A transparent cylindrical glass container (height, 40 cm; diameter, 20 cm) was used for the test. The cylinder was filled with water at 23–25°C and the water depth was adjusted to avoid the rat touching the bottom with its hind legs. One day before the test, each test rat (P34) was placed in a cylinder filled with water for 15 min. A test rat (P35) was transported from its home cage to the testing room for at least 30 min prior to the forced swim test and then placed in a water-filled cylinder for 6 min with a video camera. The duration of time spent swimming was recorded by observing a padding fashion with movement of the forelimbs or hind limbs using EthoVision software.
SCFAs Analysis
Gas chromatography-mass spectrometry (GC-MS) was used to determine fecal SCFAs levels. Fecal samples weighing 100 mg from each rat were homogenized in 50 µL of 20% phosphoric acid with 500 µM of 4-methyl-valeric acid (S381810, Sigma-Aldrich, St. Louis, USA) as an internal standard and 400 μL ether at 70 Hz for 1 min, followed by centrifugation at 14,000 × g at 4°C for 20 min. The supernatant was transferred to a new vial for GC-MS analysis using an Agilent 7890B-5977B GC/MSD system and DB-FFAP capillary column (30 m × 250 µm × 0.25 µm). Helium was used as the carrier gas at a flow rate of 1 mL/min. A volume of 1 μL supernatant was injected in split mode at 10:1 at a temperature of 250°C. The initial column temperature was 90°C, which was then increased to 160°C at 10°C/min and increased to 240°C at 40°C/min and then maintained for 5 min. The temperatures of the ion source, interface 250°C, and quadrupole were 230, 250, and 150°C, respectively. The detector was operated in electron impact ionization mode with an electron energy of 70 eV, and full scan and single ion monitoring modes were used to detect the SCFAs.
Fecal Microbiota Analysis
Total DNA extracted from fecal samples was used to amplify the bacterial 16S rRNA V3-V4 region with the primer pair 338F 5′-ACTCCTACGGGAGGCAGCAG-3′ and 806R 5′-GGACTACHVGGGTWTCTAAT-3′. The amplicons were purified and subjected to barcoded library construction using the TruSeq Nano DNA LT Library Prep Kit, and sequenced on an Illumina NovaSeq 6000 platform (Illumina, San Diego, USA). The sequences were assigned to amplicon sequence variants (ASV) using the Greengenes (v.13.8) reference databases after quality filtering using fastq_maxee 1.0, and de-noised using DADA2 software. Microbiota diversity and composition were analyzed using R software, and the Kruskal–Wallis test with Dunn’s post-hoc test and permutational multivariate analysis of variance (PERMANOVA) were used to determine significant differences in alpha diversity, ASV abundance, and beta diversity between groups, respectively. Linear discriminant analysis (LDA) effect size (LEfSe) was used to identify the enriched taxa. The Kyoto Encyclopedia of Genes and Genomes (KEGG) and cluster of orthologous groups of proteins (COG) databases were used to analyze the functional profiles of the pathway enrichment of the gut microbiome.
Statistical Analysis
Data were assessed for normality of distribution and presented as means ± standard deviations (SDs). One-way analysis of variance (ANOVA) followed by the Bonferroni test and paired T-test were used for statistical analysis among groups. Statistical analysis and graph generation were performed using SPSS 25.0 and Graph Pad Prism (version 9.0.0), and a p value of < 0.05 was considered to be statistically significant. Litter effects were not accounted for in the statistical analysis.
Results
B. adolescentis DM8504 Alleviates Autistic-Like Behaviors
To investigate the ameliorative effect of B. adolescentis DM8504 on autistic-like behaviors in VPA-exposed rats, we treated pregnant rats with B. adolescentis DM8504 from GD1 until birth. The open-field test demonstrated that VPA rats presented less exploratory behavior (time spent in the central area, entries into the central area) and moved (total distance) less than the control rats, while B. adolescentis DM8504 treatment reversed the locomotor activity and exploratory behavior impaired by VPA (Figure 2A and B). The sociability assessed by the three-chamber test showed that the VPA-exposed rats spent significantly less time in the social zone and more time in the non-social zone than the Ctrl group, whereas B. adolescentis DM8504 treatment increased time spent in the social zone and decreased time spent in the non-social zone in VPA rats (Figure 2C). In session 2, the Ctrl rats spent more time in the chamber with the stranger rat and less time in the chamber with the familiar rat, whereas the VPA-exposed rats spent more time with the familiar rat and less time with the stranger rat (Figure 2D). VPA rats receiving B. adolescentis DM8504 treatment tended to explore the chamber with a stranger (Figure 2D). The social preference index of both sessions 1 and 2 for the VPA rats was significantly lower than that of the Ctrl rats, whereas B. adolescentis DM8504 treatment increased the social preference index for VPA rats (Figure 2C and D). In the evaluation of spatial working and reference memory, the results of the Y-maze test revealed that both the duration of visits to the novel arm and spontaneous alternation were lower in VPA rats than in Ctrl rats, while B. adolescentis DM8504 treatment increased the number of entries into the novel arm and spontaneous alternation in VPA rats (Figure 2E and F). In the assessment of behavioral despair, the forced swim test showed that immobile time in the cylindrical glass container of VPA-exposed rats was significantly longer than that in Ctrl rats, whereas B. adolescentis DM8504 treatment significantly decreased immobile time in VPA rats (Figure 2G).
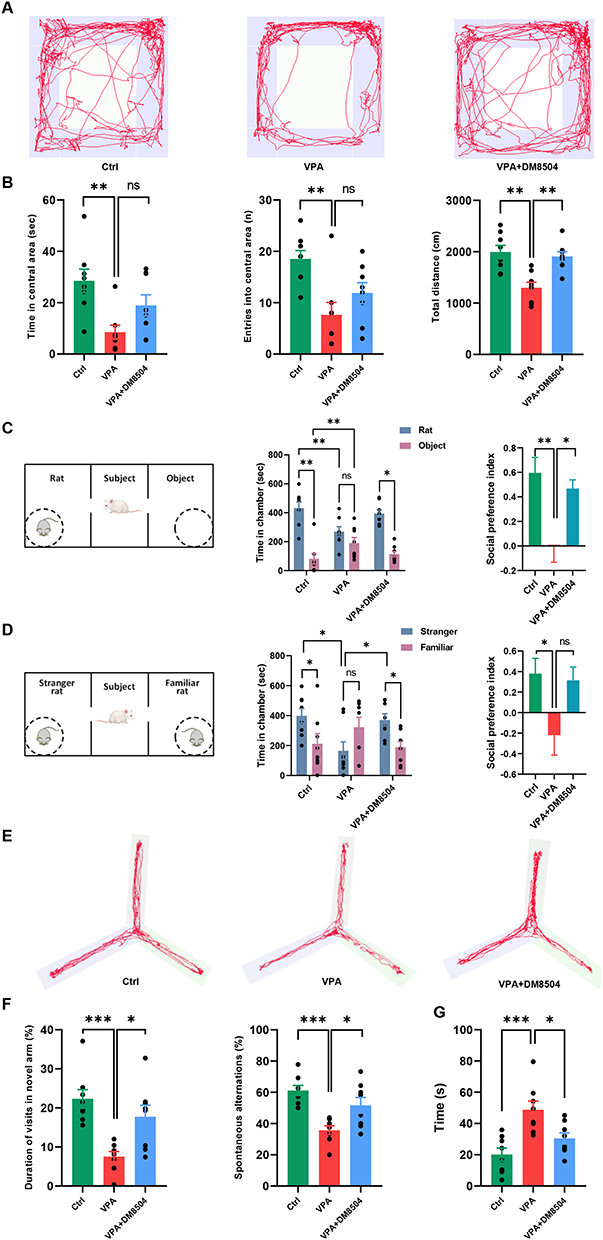 |
Figure 2 B. adolescentis DM8504 alleviates autistic-like behaviors in VPA-exposed rats. (A) The representative track plots in the open-field tests. (B) Summary of the time spent in the central area, entries into the central area, and the total travel distance of rats in the open field. (C) The paradigm for the three-chamber test (session 1) and graphs showing the total time spent in the social chamber (rat), non-social chamber (object), and social preference index. (D) The paradigm for the three-chamber test (session 2) and graphs showing the total time spent in the social chamber (stranger), non-social chamber (familiar), and social preference index. (E) The representative track plots in the Y-maze tests. (F) Summary of the duration of visits in novel arm and spontaneous alternations in the Y-maze apparatus. (G) Summary of the immobile time in the forced swim test. Data are expressed as the mean ± SD, and significance was determined by ANOVA with Bonferroni test, *P < 0.05, **P < 0.01, ***P < 0.001. Abbreviations: ns, non-significant; Ctrl, control; VPA, valproic acid. |
B. adolescentis DM8504 Modulates the Fecal SCFAs Levels
Feces from the rats of all three groups were collected one day (P36) after the behavioral tests, and the concentrations of SCFAs in fecal samples from rats were determined by GC-MS. As shown in Figure 3, fecal levels of several SCFAs, including acetic acid (Figure 3A), butyric acid (Figure 3B), propionic acid (Figure 3C), isobutyric acid (Figure 3D), valeric acid (Figure 3E), isovaleric acid (Figure 3F), and hexanoic acid (Figure 3G), were lower in rats in the VPA group than in those in the Ctrl group. B. adolescentis DM8504 treatment significantly increased the fecal levels of acetic acid (Figure 3A), butyric acid (Figure 3B), propionic acid (Figure 3C), isobutyric acid (Figure 3F), and hexanoic acid (Figure 3G) in VPA-exposed rats.
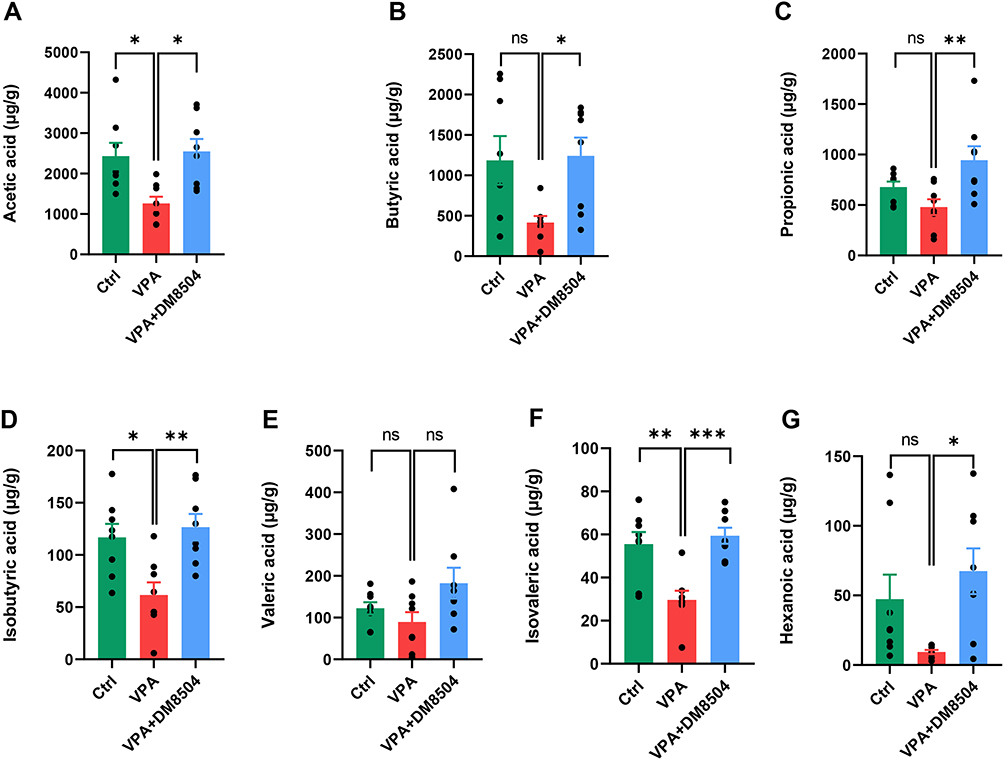 |
Figure 3 B. adolescentis DM8504 increases the levels of SCFAs. Acetic acid (A), butyric acid (B), propionic acid (C), isobutyric acid (D), valeric acid (E), isovaleric acid (F), and hexanoic acid (G) of fecal samples from the Ctrl, VPA, and VPA + DM5804 groups were determined by using GC-MS. Significance was determined by ANOVA with Bonferroni test, *P < 0.05, **P < 0.01, ***P < 0.001. Abbreviations: ns, non-significant; SCFAs, short-chain fatty acids; Ctrl, control; VPA, valproic acid. |
B. adolescentis DM8504 Modifies Gut Microbiota Composition
The alpha diversity of the gut microbiota composition represented by the observed species (Figure 4A), Chao 1 (Figure 4B), ACE (Figure 4C), and Shannon (Figure 4D) indices was significantly reduced in the VPA group compared with the Ctrl group, while B. adolescentis DM8504 treatment significantly increased the alpha diversity of the gut microbiota in VPA-exposed rats. Analysis of beta diversity with Bray-Curtis distance showed a separation between the VPA and Ctrl groups, while B. adolescentis DM8504 treatment modulated the fecal microbial community toward to Ctrl group (Figure 4E).
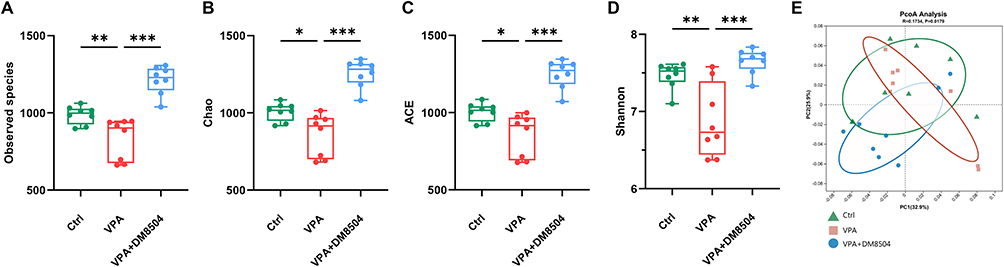 |
Figure 4 B. adolescentis DM8504 modulates the gut microbiota diversity. (A–D) Alpha diversity indexes of the fecal microbiome in the Ctrl, VPA, and VPA + DM5804 at ASV level: (A) Observed species, (B) Chao, (C) Ace, and (D) Shannon. (E) Beta diversity of the fecal microbiome using PCoA based on Bray-Curtis distance between the three groups at ASV level. Significance was determined by Kruskal–Wallis test with Dunn’s post-hoc test, *P < 0.05, **P < 0.01, ***P < 0.001. Abbreviations: Ctrl, control; VPA, valproic acid; ASV, amplicon sequence variants; PCoA, principal coordinate analysis. |
The relative abundances of the major phyla and genera in the gut microbiota of rats from different groups were further analyzed. As shown in Figure 5A, the relative abundances of phyla Proteobacteria and Verrucomicrobiota increased, while Firmicutes and Actinobacteria decreased in the VPA group compared with the Ctrl group. In the three groups, the dominant genera were Muribaculaceae, Lactobacillus, Prevotella, Bacteroides, Escherichia-Shigella, and Lachnospiraceae_NK4A136_group (Figure 5B). Intergroup comparisons of the genera revealed that the VPA group exhibited significantly lower relative abundances of Bifidobacterium, Oscillospiraceae_ NK4A214_group, Faecalibacterium, [Eubacterium]_ coprostanoligenes_group, Alloprevotella, and Lachnospiraceae_ NK4A136_group and higher relative abundances of Parabacteroides and Akkermansia than the Ctrl group (Figure 5C). Notably, the administration of B. adolescentis DM8504 ameliorated the destructive effects of VPA on gut microbiota composition in rats (Figure 5).
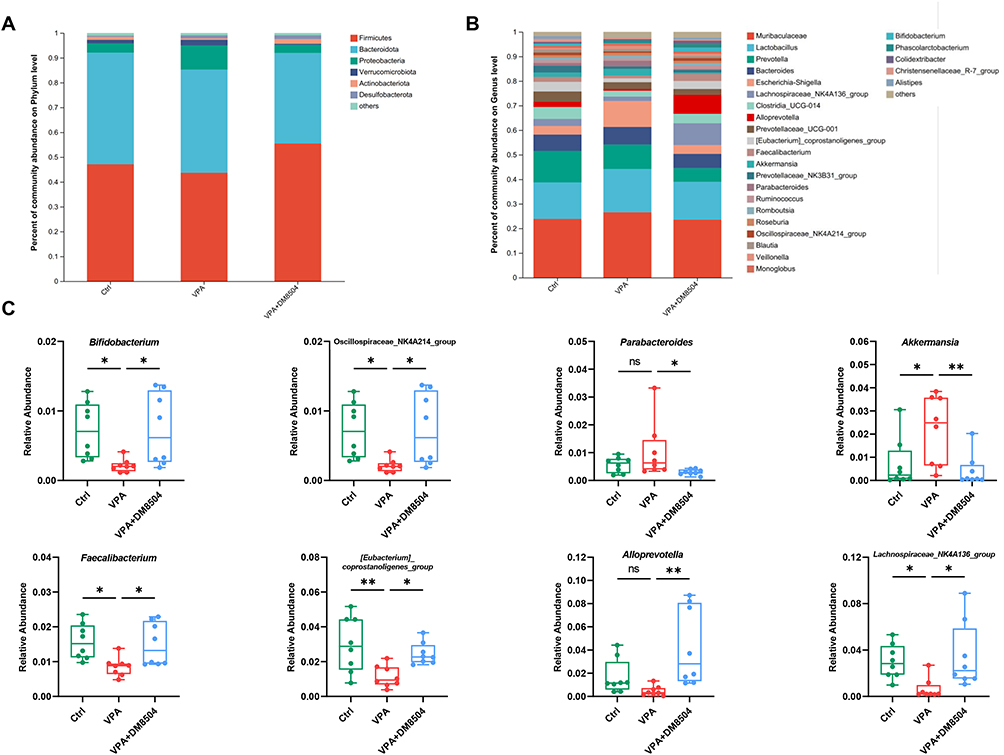 |
Figure 5 B. adolescentis DM8504 regulates the gut microbiota composition. Column chart of the species composition at the phylum (A) and genus (B) level among the Ctrl, VPA, and VPA + DM5804 groups. (C) Bacterial genera with significance differences among the Ctrl, VPA, and VPA + DM5804 groups. Significance was determined by Kruskal–Wallis test with Dunn’s post-hoc test, *P < 0.05, **P < 0.01. Abbreviations: ns, non-significant; Ctrl, control; VPA, valproic acid. |
As shown in Figure 6A, 30 differentially abundant taxa among the three groups were identified using the LEfSe analysis. Class Vampirivibrionia, order Gastranaerophilales, families Gastranaerophilales and Eubacteriumcoprostanoligenes_ group were enriched in the Ctrl group. The VPA exposure increased the abundance of classes Alphaproteobacteria, Verrucomicrobiae, orders Micrococcales, Clostridiales, Rhodospirillales, Verrucomicorbiales, and families Micrococcaceae, Tannerellaceae, Clostridiaceae, and Akkermansiaceae, whereas B. adolescentis DM8504 treatment enriched classes Actinobacteria, Coriobacteriia, Desulfovibrionia, Clostridia, orders Bifidobacteriales, Coriobacteriaceae, Desulfovibrionales, Clostridia_vadinBB60_group, Oscillospirales, families Bifidobacteriaceae, Coriobacteriaceae, Desulfovibrionaceae, ClostridiavadinBB60_ group, Butyricicoccaceae, and Ruminococcaceae. Furthermore, LDA scores (>3.5) derived from LEfSe analysis revealed several genera that discriminated the Ctrl, VPA, and VPA + DM8504 groups. Specifically, notably high abundances of Eubacterium_coprostanoligenes_group, Prevotellaceae_NK3B31_group, Prevotellaceae_UCG_003, and Alistipes were observed in the Ctrl group, while VPA exposure enriched the abundances of Akkermansia, Parabacteroides, Allobaculum, and Clostridium_sensu_stricto_1, and the abundances of Alloprevotella, Lachnospiraceae_ NK4A136_group, Bifidobacterium, and Faecalibacterium were enriched in the VPA + DM8504 group (Figure 6B).
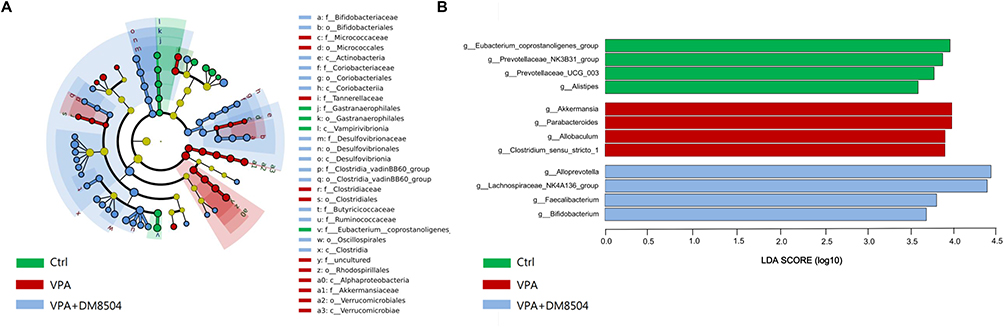 |
Figure 6 The enrichment of the gut microbiota. (A) Cladogram generated from LEfSe analysis showing the relationship between taxon (from the inner ring to the outer ring, the grades represent the phylum, class, order, family, genus and species) among the Ctrl, VPA, and VPA + DM5804 groups. Each dot represents a taxonomic hierarchy. (B) LEfSe analysis selected bacterial genera with greatest differences (LDA score >3.5) among the Ctrl, VPA, and VPA + DM5804 groups. Abbreviations: Ctrl, control; VPA, valproic acid; LEfSe, linear discriminant analysis (LDA) effect size. |
B. adolescentis DM8504 Regulates Microbiota Functional Profile
COG pathway analysis showed significantly increased levels of extracellular structures, mobilome: prophages, transposons, posttranslational modification, protein turnover, chaperones, decreased levels of defense mechanisms, general function prediction only, and transcription in the VPA group compared with the Ctrl group (Figure 7A). As for level 2 KEGG pathways, notably higher abundances of excretory system, cancer: specific types, infectious disease: bacterial, metabolism of other amino acids, membrane transport, and carbohydrate metabolism were found, while significantly lower abundances of endocrine system, folding, sorting and degradation, biosynthesis of other secondary metabolites, metabolism of cofactors and vitamins, and amino acid metabolism were identified in the VPA group than in the Ctrl group (Figure 7B). B. adolescentis DM8504 treatment normalized the altered microbial COG and KEGG pathways induced by VPA exposure (Figure 7).
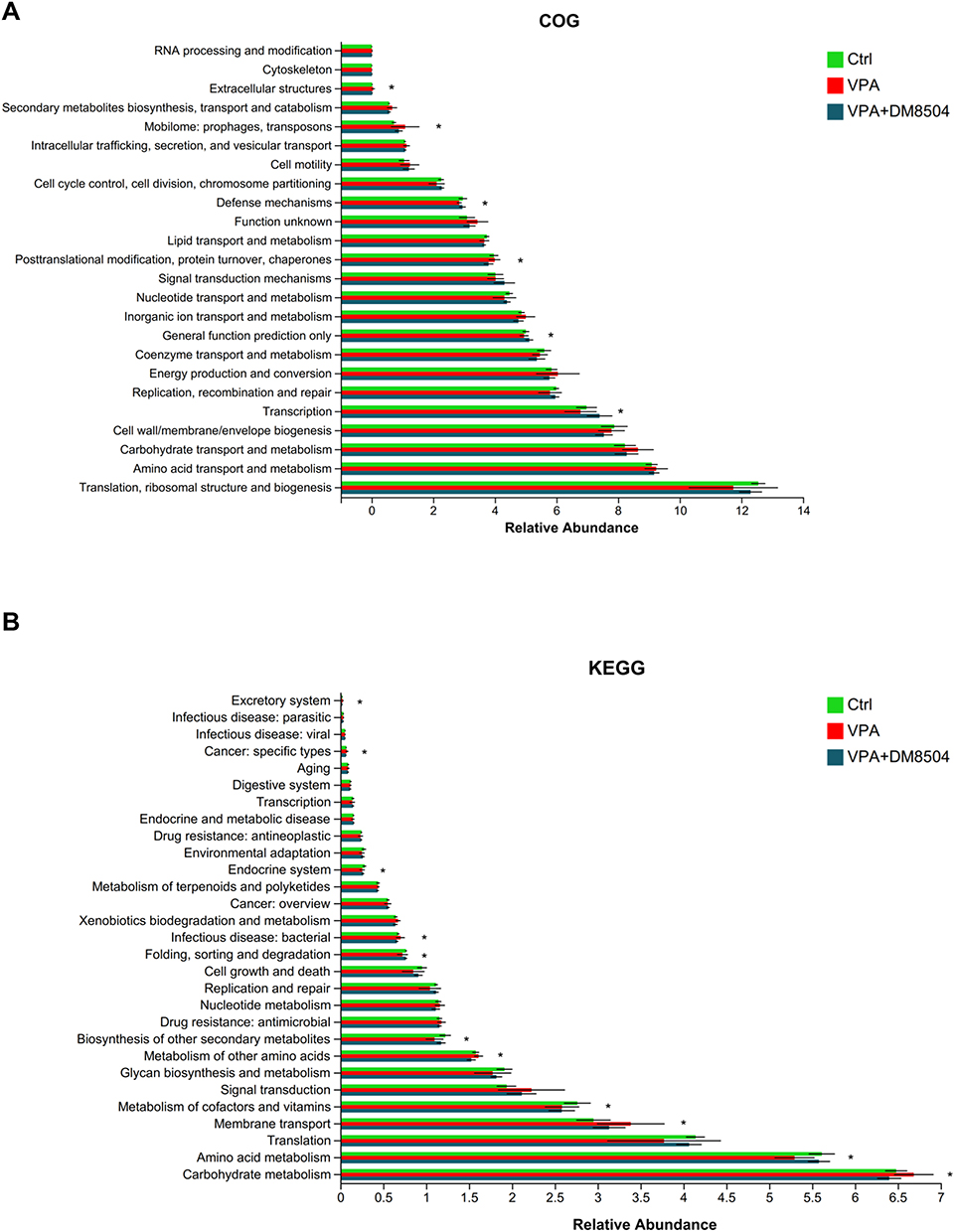 |
Figure 7 Functional profile analysis of the gut microbiome. The bar charts of COG (A) and KEGG (B) pathways showing 6 and 11 pathways with significant differences among Ctrl, VPA, and VPA + DM5804 groups, respectively. Significance was determined by Kruskal–Wallis test with Dunn’s post-hoc test, *P < 0.05. Abbreviations: COG, cluster of orthologous groups of proteins; KEGG, Kyoto Encyclopedia of Genes and Genomes; Ctrl, control; VPA, valproic acid. |
Discussion
The present study demonstrated the alleviative effect of the probiotic strain B. adolescentis DM8504 on autistic-like behaviors in a rat model of autism induced by prenatal VPA exposure. VPA-exposed male rats showed obvious autistic-like behaviors and signs of anxiety and depression similar to the clinical features of children with ASD. We found that B. adolescentis DM8504 intervention enhanced locomotor activity, exploratory behavior, sociability, and spatial working memory and relieved depression in VPA-exposed rats. Furthermore, B. adolescentis DM8504 treatment restored fecal SCFAs levels and gut microbiota composition, which were most likely responsible for the effects of B. adolescentis DM8504 on relief from autistic-like behavior, anxiety, and depression in VPA-exposed rats. Previous studies indicated that B. adolescentis strains may improve neurodegeneration and alleviate depression in mouse models.27,29 It has been reported that B. adolescentis ATCC 15703 could improve neurodegeneration in a mouse model of premature aging (Terc−/−) via increasing the activity of the catalase enzyme in brain tissue.27 Han et al demonstrated that another B. adolescentis strain, NK98, alleviated Escherichia coli-induced depression in mice by enhancing BDNF+/NeuN+ cell population and suppressing NF-κB activity in the hippocampus.29 To the best of our knowledge, our study is the first to examine the effect of B. adolescentis strains on the autistic-like behaviors in a VPA-exposed rat model.
Probiotics are defined as live micro-organisms that exert a beneficial effect on host health when administered in adequate doses.33 Probiotics have protective effects on neurological disorders, including Alzheimer’s disease, depression, Parkinson’s disease, and ASD.34 A number of studies have investigated the potential effect of probiotic strains on the relief of symptoms in both ASD patients and animal models, particularly focusing on Lactobacillus and Bifidobacterium strains.35 A placebo controlled, double blind, crossover-designed feeding trial conducted by Parracho et al showed that the probiotic strain Lactobacillus plantarum WCSF1 improved GI symptoms and behavior scores in ASD children of 4–16 years old.36 Lactobacillus acidophilus Rosell-11 supplement improved concentration, work skills, and ability to respond to an order in children with ASD.37 Long-term treatment with a mixture of Lactobacillus strains, VSL#3, resulted in an unexpected improvement in autistic core symptoms in an ASD boy with severe cognitive function problems.38 A mixture of three probiotic strains (Bifidobacterium longum, Lactobacillus rhamnosus, and Lactobacillus acidophilus) reduced the severity of autistic symptoms and improved behavioral patterns in ASD children.39 In pregnant women, Lactobacillus rhamnosus GG supplementation reduced the risk of neuropsychiatric disorder developing in their children.40 Furthermore, studies into the beneficial effects of probiotics on improvement of autistic behavior were also conducted in animal models of autism. For example, Lactobacillus reuteri supplementation reversed social deficits and abnormal nerve conduction in a maternal high fat diet-induced autism mice model.41 Daily intake of Lactobacillus helveticus CCFM1076 alleviated autistic-like behaviors in VPA-treated rats during weaning and sexual maturation.22 A probiotic mixture containing four beneficial bacterial species (Lactobacillus spp., Bifidobacterium spp.) attenuated both antibiotics and VPA-generated antisocial behavioral symptoms in rats.42 The present study showed that B. adolescentis DM8504 relieved autistic-like behavior in VPA-exposed rats. Taken together, the findings of previous studies suggest that probiotic interventions may improve behavioral abnormalities in ASD; however, further research is necessary.
The action of probiotics is believed to be mediated by the restoration of a healthy balance between altered gut microbiota and corresponding metabolites in the host. Gut microbiota affects the function of the CNS via the bidirectional microbe–gut–brain axis via multiple physiological signaling pathways, including those of the immune, nervous, metabolic, and endocrine systems.43 A number of studies have reported altered gut microbiota composition in ASD patients as compared to neurotypical individuals, such as an increase in the Firmicutes to Bacteroidetes ratio and decreased levels of Dialister, Bifidobacterium, Blautia, Veillonella, Turicibacter, and Prevotella in ASD patients.6,7,12,44 Colonization with the gut microbiota from ASD patients was sufficient to induce hallmark autistic behaviors in mice, suggesting a close link between gut microbiota and ASD development.45 Lactobacillus plantarum WCSF1 supplementation increased levels of enterococci and lactobacilli and decreased Clostridium cluster XIVa in ASD children.36 Daily intake of a probiotic diet with three Lactobacillus strains, two Bifidobacterium strains, and a Streptococcus strain normalized the Firmicutes to Bacteroidetes ratio, and the amount of Bifidobacterium spp. and Desulfovibrio spp. in the feces of ASD children.46 Bifidobacterium longum, Lactobacillus rhamnosus, and Lactobacillus acidophilus supplementations increased Bifidobacteria and Lactobacilli levels in children with ASD.39 In studies conducted in animal ASD models, Kong et al showed that the diversity of microbiota in VPA-treated rats was significantly increased after Lactobacillus helveticus CCFM1076 intervention, and resulted in a reduction of Turicibacter levels.22 In this study, the diversity of gut microbiota was significantly decreased in VPA-exposed rats as compared with the Ctrl group. The VPA group exhibited a similar pattern of changes in microbiota composition in ASD patients, such as significantly lower relative Bifidobacterium and Faecalibacterium levels, and higher levels of Parabacteroides and Akkermansia.35 B. adolescentis DM8504 treatment significantly restored the dysregulated gut microbiota induced by VPA in rats, as evidenced by the increased diversity and abundance of beneficial bacterial genera. In particular, the abundances of several gut bacterial species capable of producing SCFAs were significantly increased in VPA-exposed rats following B. adolescentis DM8504 treatment, including Bifidobacterium, Faecalibacterium, Eubacterium, and Lachnospiraceae_ NK4A136_group. In addition, the gut microbiota functional profile of KEGG and COG pathway analyses revealed that B. adolescentis DM8504 could recover the pathways altered by VPA exposure in rats.
As major products of the bacterial fermentation of dietary fiber, SCFAs play important roles in communication between gut microbiota and the brain through regulating blood–brain barrier integrity, neurotransmitter modulation, and neuroinflammation.47 SCFAs are able to increase the integrity of the BBBs. For example, both oral sodium butyrate and SCFA-producing bacteria strains administration could reduce the BBB permeability in germ-free (GF) mice.48 The effect of SCFAs on the improvement of the BBB integrity is mainly via upregulating tight junction proteins (TJPs) expression and inhibiting inflammation-related pathways in brain endothelial cells.47,49 SCFAs restore the expression of TJPs through suppressing NF-κB and promoting Nrf2 activation via activation of the G protein-coupled receptors (GPR), such as GPR41/43, and inhibition of histone deacetylases (HDAC) activity.49 SCFAs modulate the levels of neurotransmitters in the CNS that exert neuroactive properties. Acetate that crosses into the CNS altered the glutamate, glutamine, and gamma-aminobutyric acid (GABA) levels in the hypothalamus.50 Propionic acid induced tryptophan 5-hydroxylase 1 expression in a pheochromocytoma cell line, with possible relevance to ASD.51 SCFAs modulate the neurotrophic factors to affect the brain functions of learning and memory, including nerve growth factor (NGF), brain-derived neurotrophic factor (BDNF), and glial cell line-derived neurotrophic factor (GDNF).47 Furthermore, SCFAs inhibit neuroinflammation through regulating both peripheral and CNS immune responses. Peripherally, SCFAs decreased the systemic inflammation via regulating the interleukins expression, enhancing the intestinal barrier and preventing bacterial translocation, which in turn to reduce the neuroinflammation.47,52 SCFAs directly affect the morphology and functions of immune cells in the CNS, such as microglia cells, to modulate the neuroinflammation, thereby potentially affecting the brain functions in mental disorders.53 SCFAs can also influence the brain functions of learning, memory, and mood through the endocrine pathway by inducing the gut hormones secretion from the enteroendocrine L cells, such as glucagon-like peptide 1 (GLP1) and peptide YY (PYY).54,55 Taken together, SCFAs play critical roles in microbiota–gut–brain communication and brain function through multiple pathways.
To date, only limited human studies have reported the fecal levels of SCFAs with inconclusive results in children with ASD. De Angelis et al56 and Adams et al10 reported decreased total SCFAs levels, while Wang et al17 showed increased SCFAs levels in children with ASD compared to those in normally developing children. In contrast, a study conducted by Kang et al reported no significant differences in SCFAs levels between children with ASD and neurotypical subjects.15 Nevertheless, several studies conducted in animal ASD models have demonstrated a beneficial effect of SCFAs on the relief of autistic-like behavior.57–59 For example, intraperitoneal injection of sodium butyrate can attenuate social behavior deficits in autistic-like mice.57 A post-weaning administration of acetic and propionic acid reversed maternal obesity-induced cognitive and social deficits in the offspring of obese mice by improving synaptic ultrastructure and microglial maturation in the hippocampus.58 The supplementation of SCFAs could alleviate stress-responsivity, anxiety- and depressive-like behavior induced by repeated psychosocial stress in mice.59 Our data showed that the fecal levels of SCFAs were decreased in VPA-exposed rats, and B. adolescentis DM8504 could normalize the SCFAs levels, which is consistent with previous studies.26 Thus, B. adolescentis DM8504 improved the social ability of VPA-exposed rats, which may be associated with the restoration of SCFAs levels through regulation of the abundance of gut bacterial species capable of producing SCFAs.
There are several limitations in the current study. Firstly, the effects of B. adolescentis on autistic-like behaviors were evaluated by using only one strain, more B. adolescentis strains should be included to further validate our finding in the future. Secondly, only VPA-exposed rats model was used in this study. Thirdly, we did not determine neither serum nor fecal metabolites in VPA-exposed rats, particularly neurotransmitter levels, such as GABA. Finally, the effects of B. adolescentis DM8504 on the gut microbiota composition of the pregnant rats were not investigated.
Conclusion
In summary, our study demonstrated that B. adolescentis DM8504 administration could alleviate autistic-like behaviors in VPA-exposed rats. B. adolescentis DM8504 treatment restores the SCFA levels and the abundances of SCFA-producing bacterial species, including Bifidobacterium, Faecalibacterium, and Eubacterium. Behavioral improvements in locomotion, sociability, and working memory in VPA-exposed rats by B. adolescentis DM8504 are likely through restoration of SCFAs levels and enrichment of SCFA-producing bacteria. Therefore, well-defined B. adolescentis strains may have potential for the management of children with ASD in the future.
Abbreviations
ANOVA, analysis of variance; ASD, autism spectrum disorder; ASV, amplicon sequence variants; BBB, blood–brain barrier; BDNF, brain-derived neurotrophic factor; CNS, central nervous system; COG, cluster of orthologous groups of proteins; Ctrl, control; FMT, fecal microbiota transplantation; GABA, gamma-aminobutyric acid; GC-MS, gas chromatography-mass spectrometry; GD, gestational day; GDNF, glial cell line-derived neurotrophic factor; GI, gastrointestinal; GLP1, glucagon-like peptide 1; KEGG, Kyoto Encyclopedia of Genes and Genomes; LEfSe, linear discriminant analysis (LDA) effect size; NGF, nerve growth factor; PYY, peptide YY; SCFAs, short-chain fatty acids; SD, standard deviation; TPY, trypticase peptone yeast extract; VPA, valproic acid.
Data Sharing Statement
The datasets used and/or analyzed during the current study are available from the corresponding author, Dr. Hongfang Ding, upon reasonable request.
Ethics Approval and Consent to Participate
This study was approved by the Animal Ethics Committee of Binzhou Medical University (2025-310). All animal experiments were performed in accordance with the Guidelines for the Care and Use of Laboratory Animals of Binzhou Medical University.
Funding
This work was supported by grants from the National Natural Science Foundation of China (32370951) and Natural Science Foundation of Dongying, China (2023ZR038).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Lord C, Elsabbagh M, Baird G, Veenstra-Vanderweele J. Autism spectrum disorder. Lancet. 2018;392(10146):508–520. doi:10.1016/S0140-6736(18)31129-2
2. Tafolla M, Singer H, Lord C. Autism spectrum disorder across the lifespan. Annu Rev Clin Psychol. 2025;21(1):193–220. doi:10.1146/annurev-clinpsy-081423-031110
3. Grosvenor LP, Croen LA, Lynch FL, et al. Autism diagnosis among US children and adults, 2011–2022. JAMA Network Open. 2024;7(10):e2442218. doi:10.1001/jamanetworkopen.2024.42218
4. Solmi M, Song M, Yon DK, et al. Incidence, prevalence, and global burden of autism spectrum disorder from 1990 to 2019 across 204 countries. Mol Psychiatry. 2022;27(10):4172–4180. doi:10.1038/s41380-022-01630-7
5. Maenner MJ, Warren Z, Williams AR, et al. Prevalence and characteristics of autism spectrum disorder among children aged 8 years - autism and developmental disabilities monitoring network, 11 sites, United States, 2020. MMWR Surveill Summ. 2023;72(2):1–14. doi:10.15585/mmwr.ss7202a1
6. Svoboda E. Could the gut microbiome be linked to autism? Nature. 2020;577(7792):S14–S15. doi:10.1038/d41586-020-00198-y
7. Hsiao EY, McBride SW, Hsien S, et al. Microbiota modulate behavioral and physiological abnormalities associated with neurodevelopmental disorders. Cell. 2013;155(7):1451–1463. doi:10.1016/j.cell.2013.11.024
8. Iglesias-Vazquez L, Van Ginkel Riba G, Arija V, Canals J. Composition of gut microbiota in children with autism spectrum disorder: a systematic review and meta-analysis. Nutrients. 2020;12(3):792. doi:10.3390/nu12030792
9. Tran SM, Mohajeri MH. The role of gut bacterial metabolites in brain development, aging and disease. Nutrients. 2021;13(3):732. doi:10.3390/nu13030732
10. Adams JB, Johansen LJ, Powell LD, Quig D, Rubin RA. Gastrointestinal flora and gastrointestinal status in children with autism--comparisons to typical children and correlation with autism severity. BMC Gastroenterol. 2011;11:22. doi:10.1186/1471-230X-11-22
11. Wan Y, Zuo T, Xu Z, et al. Underdevelopment of the gut microbiota and bacteria species as non-invasive markers of prediction in children with autism spectrum disorder. Gut. 2022;71(5):910–918. doi:10.1136/gutjnl-2020-324015
12. Ding H, Yi X, Zhang X, Wang H, Liu H, Mou WW. Imbalance in the gut microbiota of children with autism spectrum disorders. Front Cell Infect Microbiol. 2021;11:572752. doi:10.3389/fcimb.2021.572752
13. Takyi E, Nirmalkar K, Adams J, Krajmalnik-Brown R. Interventions targeting the gut microbiota and their possible effect on gastrointestinal and neurobehavioral symptoms in autism spectrum disorder. Gut Microbes. 2025;17(1):2499580. doi:10.1080/19490976.2025.2499580
14. Aziz-Zadeh L, Ringold SM, Jayashankar A, et al. Relationships between brain activity, tryptophan-related gut metabolites, and autism symptomatology. Nat Commun. 2025;16(1):3465. doi:10.1038/s41467-025-58459-1
15. Kang DW, Ilhan ZE, Isern NG, et al. Differences in fecal microbial metabolites and microbiota of children with autism spectrum disorders. Anaerobe. 2018;49:121–131. doi:10.1016/j.anaerobe.2017.12.007
16. Xiao L, Yan J, Yang T, et al. Fecal microbiome transplantation from children with autism spectrum disorder modulates tryptophan and serotonergic synapse metabolism and induces altered behaviors in germ-free mice. mSystems. 2021;6(2):e01343–01320. doi:10.1128/msystems.01343-20
17. Wang L, Christophersen CT, Sorich MJ, Gerber JP, Angley MT, Conlon MA. Elevated fecal short chain fatty acid and ammonia concentrations in children with autism spectrum disorder. Dig Dis Sci. 2012;57(8):2096–2102. doi:10.1007/s10620-012-2167-7
18. Lai MC, Anagnostou E, Wiznitzer M, Allison C, Baron-Cohen S. Evidence-based support for autistic people across the lifespan: maximising potential, minimising barriers, and optimising the person-environment fit. Lancet Neurol. 2020;19(5):434–451. doi:10.1016/S1474-4422(20)30034-X
19. Canitano R. New experimental treatments for core social domain in autism spectrum disorders. Front Pediatr. 2014;2:61. doi:10.3389/fped.2014.00061
20. Parker KJ, Oztan O, Libove RA, et al. A randomized placebo-controlled pilot trial shows that intranasal vasopressin improves social deficits in children with autism. Sci Transl Med. 2019;11(491):eaau7356. doi:10.1126/scitranslmed.aau7356
21. Singh K, Connors SL, Macklin EA, et al. Sulforaphane treatment of autism spectrum disorder (ASD). Proc Natl Acad Sci U S A. 2014;111(43):15550–15555. doi:10.1073/pnas.1416940111
22. Kong Q, Wang B, Tian P, et al. Daily intake of Lactobacillus alleviates autistic-like behaviors by ameliorating the 5-hydroxytryptamine metabolic disorder in VPA-treated rats during weaning and sexual maturation. Food Funct. 2021;12(6):2591–2604. doi:10.1039/D0FO02375B
23. Li Y, Wang Y, Zhang T. Fecal microbiota transplantation in autism spectrum disorder. Neuropsychiatr Dis Treat. 2022;18:2905–2915. doi:10.2147/NDT.S382571
24. Li Y, Xiao P, Cao R, et al. Effects and microbiota changes following oral lyophilized fecal microbiota transplantation in children with autism spectrum disorder. Front Pediatr. 2024;12:1369823. doi:10.3389/fped.2024.1369823
25. Li Y, Xiao P, Ding H, et al. Fecal microbiota transplantation in children with autism. Neuropsychiatr Dis Treat. 2024;20:2391–2400. doi:10.2147/NDT.S488001
26. Leser T, Baker A. Bifidobacterium adolescentis - a beneficial microbe. Benef Microbes. 2023;14(6):525–551. doi:10.1163/18762891-20230030
27. Chen S, Chen L, Qi Y, et al. Bifidobacterium adolescentis regulates catalase activity and host metabolism and improves healthspan and lifespan in multiple species. Nat Aging. 2021;1(11):991–1001. doi:10.1038/s43587-021-00129-0
28. Duranti S, Ruiz L, Lugli GA, et al. Bifidobacterium adolescentis as a key member of the human gut microbiota in the production of GABA. Sci Rep. 2020;10(1):14112. doi:10.1038/s41598-020-70986-z
29. Han SK, Kim JK, Joo MK, Lee KE, Han SW, Kim DH. Lactobacillus reuteri NK33 and Bifidobacterium adolescentis NK98 alleviate Escherichia coli-induced depression and gut dysbiosis in mice. J Microbiol Biotechnol. 2020;30(8):1222–1226. doi:10.4014/jmb.2002.02058
30. Kitanaka J, Kitanaka N, Hall FS, et al. Memory impairment and reduced exploratory behavior in mice after administration of systemic morphine. J Exp Neurosci. 2015;9:27–35. doi:10.4137/JEN.S25057
31. Dellu F, Mayo W, Cherkaoui J, Le Moal M, Simon H. A two-trial memory task with automated recording: study in young and aged rats. Brain Res. 1992;588(1):132–139. doi:10.1016/0006-8993(92)91352-F
32. Yankelevitch-Yahav R, Franko M, Huly A, Doron R. The forced swim test as a model of depressive-like behavior. J Vis Exp. 2015;2015(97):52587. doi:10.3791/52587
33. Hill C, Guarner F, Reid G, et al. Expert consensus document: the International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat Rev Gastroenterol Hepatol. 2014;11(8):506–514. doi:10.1038/nrgastro.2014.66
34. Yang H, Liu Y, Cai R, Li Y, Gu B. A narrative review of relationship between gut microbiota and neuropsychiatric disorders: mechanisms and clinical application of probiotics and prebiotics. Ann Palliat Med. 2021;10(2):2304–2313. doi:10.21037/apm-20-1365
35. Sanlier N, Kocabas S. The effect of probiotic, prebiotic and gut microbiota on ASD: a review and future perspectives. Crit Rev Food Sci Nutr. 2023;63(15):2319–2330. doi:10.1080/10408398.2021.1973957
36. Parracho HMRT, Gibson GR, Knott F, Bosscher D, Kleerebezem M, Mccartney AL. A double-blind, placebo-controlled, crossover-designed probiotic feeding study in children diagnosed with autistic spectrum disorders. Int J Probiotics Prebiotics. 2010;5(2):69–74.
37. Kaluzna-Czaplinska J, Blaszczyk S. The level of arabinitol in autistic children after probiotic therapy. Nutrition. 2012;28(2):124–126. doi:10.1016/j.nut.2011.08.002
38. Grossi E, Melli S, Dunca D, Terruzzi V. Unexpected improvement in core autism spectrum disorder symptoms after long-term treatment with probiotics. SAGE Open Med Case Rep. 2016;4:2050313X16666231. doi:10.1177/2050313X16666231
39. Shaaban SY, El Gendy YG, Mehanna NS, et al. The role of probiotics in children with autism spectrum disorder: a prospective, open-label study. Nutr Neurosci. 2018;21(9):676–681. doi:10.1080/1028415X.2017.1347746
40. Partty A, Kalliomaki M, Wacklin P, Salminen S, Isolauri E. A possible link between early probiotic intervention and the risk of neuropsychiatric disorders later in childhood: a randomized trial. Pediatr Res. 2015;77(6):823–828. doi:10.1038/pr.2015.51
41. Buffington SA, Di Prisco GV, Auchtung TA, Ajami NJ, Petrosino JF, Costa-Mattioli M. Microbial reconstitution reverses maternal diet-induced social and synaptic deficits in offspring. Cell. 2016;165(7):1762–1775. doi:10.1016/j.cell.2016.06.001
42. Mintal K, Toth A, Hormay E, et al. Novel probiotic treatment of autism spectrum disorder associated social behavioral symptoms in two rodent models. Sci Rep. 2022;12(1):5399. doi:10.1038/s41598-022-09350-2
43. Bercik P, Collins SM, Verdu EF. Microbes and the gut-brain axis. Neurogastroenterol Motil. 2012;24(5):405–413. doi:10.1111/j.1365-2982.2012.01906.x
44. Liu F, Li J, Wu F, Zheng H, Peng Q, Zhou H. Altered composition and function of intestinal microbiota in autism spectrum disorders: a systematic review. Transl Psychiatry. 2019;9(1):43. doi:10.1038/s41398-019-0389-6
45. Sharon G, Cruz NJ, Kang DW, et al. Human gut microbiota from autism spectrum disorder promote behavioral symptoms in mice. Cell. 2019;177(6):1600–1618e1617. doi:10.1016/j.cell.2019.05.004
46. Tomova A, Husarova V, Lakatosova S, et al. Gastrointestinal microbiota in children with autism in Slovakia. Physiol Behav. 2015;138:179–187. doi:10.1016/j.physbeh.2014.10.033
47. Dalile B, Van Oudenhove L, Vervliet B, Verbeke K. The role of short-chain fatty acids in microbiota-gut-brain communication. Nat Rev Gastroenterol Hepatol. 2019;16(8):461–478. doi:10.1038/s41575-019-0157-3
48. Braniste V, Al-Asmakh M, Kowal C, et al. The gut microbiota influences blood-brain barrier permeability in mice. Sci Transl Med. 2014;6(263):263ra158. doi:10.1126/scitranslmed.3009759
49. Fock E, Parnova R. Mechanisms of blood-brain barrier protection by microbiota-derived short-chain fatty acids. Cells. 2023;12(4):657. doi:10.3390/cells12040657
50. Frost G, Sleeth ML, Sahuri-Arisoylu M, et al. The short-chain fatty acid acetate reduces appetite via a central homeostatic mechanism. Nat Commun. 2014;5:3611. doi:10.1038/ncomms4611
51. Nankova BB, Agarwal R, MacFabe DF, La Gamma EF. Enteric bacterial metabolites propionic and butyric acid modulate gene expression, including CREB-dependent catecholaminergic neurotransmission, in PC12 cells--possible relevance to autism spectrum disorders. PLoS One. 2014;9(8):e103740. doi:10.1371/journal.pone.0103740
52. Soliman ML, Smith MD, Houdek HM, Rosenberger TA. Acetate supplementation modulates brain histone acetylation and decreases interleukin-1beta expression in a rat model of neuroinflammation. J Neuroinflammation. 2012;9:51. doi:10.1186/1742-2094-9-51
53. Correa-Oliveira R, Fachi JL, Vieira A, Sato FT, Vinolo MA. Regulation of immune cell function by short-chain fatty acids. Clin Transl Immunol. 2016;5(4):e73. doi:10.1038/cti.2016.17
54. Tolhurst G, Heffron H, Lam YS, et al. Short-chain fatty acids stimulate glucagon-like peptide-1 secretion via the G-protein-coupled receptor FFAR2. Diabetes. 2012;61(2):364–371. doi:10.2337/db11-1019
55. Larraufie P, Martin-Gallausiaux C, Lapaque N, et al. SCFAs strongly stimulate PYY production in human enteroendocrine cells. Sci Rep. 2018;8(1):74. doi:10.1038/s41598-017-18259-0
56. De Angelis M, Piccolo M, Vannini L, et al. Fecal microbiota and metabolome of children with autism and pervasive developmental disorder not otherwise specified. PLoS One. 2013;8(10):e76993. doi:10.1371/journal.pone.0076993
57. Kratsman N, Getselter D, Elliott E. Sodium butyrate attenuates social behavior deficits and modifies the transcription of inhibitory/excitatory genes in the frontal cortex of an autism model. Neuropharmacology. 2016;102:136–145. doi:10.1016/j.neuropharm.2015.11.003
58. Liu X, Li X, Xia B, et al. High-fiber diet mitigates maternal obesity-induced cognitive and social dysfunction in the offspring via gut-brain axis. Cell Metab. 2021;33(5):923–938e926. doi:10.1016/j.cmet.2021.02.002
59. van de Wouw M, Boehme M, Lyte JM, et al. Short-chain fatty acids: microbial metabolites that alleviate stress-induced brain-gut axis alterations. J Physiol. 2018;596(20):4923–4944. doi:10.1113/JP276431
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
The Interplay Among Gut Microbial Composition, Short-Chain Fatty Acid Metabolism and Gut-Kidney Oxidative Stress: Correlation with Diarrhea
Shen J, Fang L, Peng X, Jiang M, Xie X, Tan Z
Journal of Inflammation Research 2026, 19:588857
Published Date: 20 April 2026
Short-Chain Fatty Acids: A Key Modulator of Sepsis-Associated Acute Respiratory Distress Syndrome
Sun Y, He M, Li J, Wu C, Pan S
Journal of Inflammation Research 2026, 19:608128
Published Date: 18 June 2026