")
Back to Journals » Cancer Management and Research » Volume 11
Baohuoside I Inhibits the Proliferation of Pancreatic Cancer Cells via mTOR/S6K1-Caspases/Bcl2/Bax Apoptotic Signaling
Authors Ni F , Tang H, Wang C , Zhang H , Zheng C , Zhang N, Chen B, Sun L
Received 27 August 2019
Accepted for publication 6 December 2019
Published 19 December 2019 Volume 2019:11 Pages 10609—10621
DOI https://doi.org/10.2147/CMAR.S228926
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Antonella D'Anneo
Fubiao Ni,1,* Hengjie Tang,1,* Cheng Wang,1,* Hewei Zhang,1 Chenlei Zheng,1 Ning Zhang,2 Bicheng Chen,1 Linxiao Sun1
1Key Laboratory of Diagnosis and Treatment of Severe Hepato-Pancreatic Diseases of Zhejiang Province, Zhejiang Provincial Top Key Discipline in Surgery, First Affiliated Hospital of Wenzhou Medical University, Wenzhou, Zhejiang 325000, People’s Republic of China; 2First School of Clinical Medicine, Wenzhou Medical University, Wenzhou, Zhejiang 325000, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Linxiao Sun; Bicheng Chen
Email [email protected]; [email protected]
Background: Although the incidence of pancreatic cancer has increased markedly, the 5-year survival rate for this disease is considerably low compared with other types of cancer. Moreover, the mortality rate of pancreatic cancer is similar to its incidence rate. Current therapeutic agents exhibit a lack of specificity for pancreatic cancer. Baohuoside I is traditionally used to treat orgasmic disorder and inflammation. However, its role in pancreatic cancer is unknown.
Objective: To explore the effects of Baohuoside I on pancreatic cancer and to study the potential-related molecular mechanism.
Materials and methods: In the present study, the antineoplastic effect of Baohuoside I was investigated with regard to pancreatic cancer via colony formation, transwell and migration assay. The energy metabolism changes of pancreatic cancer were tested by flow cytometry analysis and oxidative phosphorylation and glycolysis assay. The target signaling members were analyzed by Western blot.
Results: Baohuoside I inhibited the cell growth of pancreatic cancer cells. In addition, it affected intracellular energy metabolism to induce cancer cell apoptosis via the mTOR/S6K1 and the caspase/Bcl2/Bax signaling pathways.
Conclusion: The present data provide further insight into the development of novel drugs against pancreatic cancer.
Keywords: Baohuoside I, pancreatic cancer, apoptosis
Introduction
Although the incidence and mortality of pancreatic cancer increases annually, specific drugs for pancreatic cancer are not currently available partly due to the highly aggressive nature of this disease.1,2 Tumor cells exhibit rapid proliferation and high invasive and migratory activity. A combination of drugs is frequently required to suppress tumor development.3,4 Previous studies have focused on the treatment of numerous types of pancreatic cancer cells with different “aggressiveness”, which involves the development of drugs against pancreatic cancer.5 PANC-1 cells were isolated from a 56-year-old male patient who presented with a pancreatic head malignant tumor. The PANC-1 cell line is the most widely used and ideal cell line for cancer drug development studies due to its high motility and highly invasive activity.6,7 The CFPAC-1 cell line was isolated from a 26-year-old male patient with well-differentiated pancreatic head malignant tumor. This cell line is suitable for exploring idiopathic pancreatic cancer.8
Baohuoside I is a biologically active flavonoid from the Epimedium grandiflorum species that exhibits limited adverse reactions. The traditional pharmaceutical use of Baohuoside I includes the treatment of impotence. Additional studies have reported that it can protect against inflammation.9 In the early 1990s, Thong et al10 suggested the potential anti-tumor properties of Baohuoside I. It was not until recently that the function of Baohuoside I in suppressing cancer cell proliferation was revealed.11,12 This compound was shown to possess limited side effects. Although the mechanism of its action has not been fully investigated, Baohuoside I exerts anti-metastatic activity in breast cancer11 and inhibits cancer cell viability in non-small cell lung cancer.12 However, the effects of Baohuoside I in other types of cancer, notably in pancreatic cancer are not clear. Moreover, the regulatory mechanism of Baohuoside I on cancer progression requires further investigation.
In the present study, the effects of Baohuoside I in acquired pancreatic cancer cells (PANC-1 cells) and idiopathic pancreatic cancer cells (CFPAC-1 cells) were assessed. It was shown that Baohuoside I suppressed the growth of pancreatic cancer cells. Furthermore, a potential mechanism of Baohuoside I was proposed, which involved induction of pancreatic cancer cell apoptosis via the mTOR/S6K1 and the caspase/Bcl2/Bax signaling pathways.
Materials and Methods
Drugs and Antibodies
Baohuoside I was purchased from YuanYe biotechnology (Shanghai, China). Baohuoside I was dissolved in DMSO at a final concentration of 100 mM. Compound C (CC; Sigma-Aldrich, Missouri, US) was dissolved in DMSO as 10mM. The Annexin V-FITC Apoptosis kit was purchased from BestBio Company (Shanghai, China). The Cell Counting Kit-8 (CCK-8) assay was purchased from BestBio Company (Shanghai, China). The antibodies against mTOR (catalog no. ab2732), p62 (catalog no. ab155686) and caspase-3 (catalog no. ab2302) were purchased from Abcam. The antibodies against S6K1 (catalog no. CST 9202), phosphorylated (p)-S6K1 (catalog no. CST 9204S), AMPK (catalog no. CST 2532S), p-AMPKα1 (catalog no. CST 2537), p-mTOR (catalog no. CST 5536S), LC3A/B (catalog no. CST 12741), caspase 8 (catalog no. CST 4790) and Bax (catalog no. CST 5023S) were purchased from Cell Signaling Technology, Inc. The antibody against Bcl2 (catalog no. 12789-1-AP) and Cora Lite 488 conjugated Affinipure second antibody (catalog no. SA00013-2) was purchased from ProteinTech Group, Inc. The GAPDH antibody (catalog no. AP0063) was purchased from Bioworld Technology, Inc. The Ki67 antibody (catalog no. AF0198) was purchased from Affinity Biosciences, Inc.
Cells and Cell Culture
The normal pancreatic cells hTERT-HPNE and human pancreatic cancer cell line PANC-1 and CFPAC-1 were obtained from the American Type Culture Collection (ATCC, Manassas, USA). The cells were cultured in Dulbecco’s modified Eagle’s medium (DMEM; GENOM, Hangzhou, China), containing 10% fetal bovine serum (FBS; Thermo Fisher Scientific, Waltham, USA), and 1% Penicillin-Streptomycin (Thermo Fisher Scientific, Waltham, USA) and maintained at 37° in a 5% CO2 humidified atmosphere. The cells were passaged every 2–3 days.
Cell Viability Assay
The viability of hTERT-HPNE, PANC-1 and CFPAC-1 cells was measured using the Cell Counting CCK-8 assay (BestBio Company, Shanghai, China) according to the manufacturer’s instructions. The cells were cultured in 96-well plates at a concentration of 5×103/well. The cells were cultured for 24 h and treated with the 10μM, 20μM, 30μM, 40μM, 50μM, 60μM, 70μM, 80μM, 90μM Baohuoside I or an equal volume of DMEM medium. Following 24 h of treatment, the cells were treated with 100 μL CCK-8 solution and incubated in the dark for an additional 2 h at 37°C. Cell viability was quantified at 450 nm by a Multiskan Spectrum spectrophotometer (Thermo Fisher Scientific, Inc.) using the following formula: [(OD450 of treated cells – OD450 of blank cells)/(OD450 of control cells – OD450 of blank cells) ×100%].
Real-Time Cellular Analysis
The cells were seeded at a concentration of 2×105 cells/well in cell culture E16-Plates (ACEA Biosciences, San Diego, USA). The proliferation assay and the cell growth index were recorded using the Label-free Real-time Cellular Analysis (RTCA) system (Roche Applied Science, Penzberg, Germany). The cell growth index was normalized at every time point following cell treatment.
Colony Formation Assay
The cells were plated in 6-well plates with 1000–2000 cells/well. Following cell growth into colonies that were visible to the naked eye, the cells were treated with Baohuoside I and/or CC. Following 24 h of treatment, the colonies were fixed with formaldehyde, stained with crystal violet and finally counted.
Migration Assay
The exponentially growing cells were seeded in 6-well plates and incubated at 37°C for 48 h. Subsequently, the culture area was scratched with a crystal pipette tip to create a linear gap in the confluent cell monolayer. The detached cells were washed away with PBS and the medium with 0, 20 or 50 µm Baohuoside I was added. The cells were allowed to fill the gap and images of the culture area were obtained using an inverted microscope at 24 h.
Transwell Assay
A Transwell assay (Costar; Corning, Inc.) was used to evaluate the invasive activity of PANC-1 and CFPAC-1 cells in vitro. The cells were incubated with Baohuoside I at a concentration of 1×105 cells in 500 μL serum-free medium. The compound was added to the upper chamber, which was coated with growth factor reduced Matrigel® for the invasive assay, and medium containing 10% FBS was added to the lower chamber as a chemoattractant. Following incubation for the appropriate time, cells on the upper surface of the membrane were removed by a Q-tip and the invaded cells were fixed with formaldehyde and stained using 0.5% crystal violet (Sigma-Aldrich; Merck KGaA). The number of invaded cells was counted in five randomly selected fields using a microscope.
Flow Cytometry Analysis of Apoptosis
The cells were treated with Baohuoside I in a 6-well plate (5×105/mL, 2 mL/well) and washed with PBS. When the cells reached 85% confluence, they were harvested and resuspended in binding buffer at a density of 5×105 cells/mL. The cells were incubated with 5 μL Annexin V-FITC for 15 min at room temperature in the dark and finally incubated with 5 μL propidium iodide (PI) for a further 5 min under the same conditions. Finally, flow cytometry was performed using a FACS C6 instrument and the data were analyzed using FlowJo 7.6 software (FlowJo LLC).
Protein Extraction and Western Blotting
Following treatment with different concentrations of Baohuoside I, the cells were lysed in ice-cold RIPA lysis buffer (Beyotime, Shanghai, China) supplemented with 10% PhosSTOP (Beyotime, Shanghai, China), 1% PMSF (Beyotime, Shanghai, China) and 1% DTT, incubated on ice for 30 min and finally centrifuged for 10 min (12,000 × g, 4°C). The supernatant was collected and the protein concentration was calculated using the Pierce BCA protein assay (Beyotime, Shanghai, China). The total protein was subjected to 12% SDS-PAGE and transferred to PVDF membranes (Bio-Rad Laboratories, Inc.). Following blocking with 5% skimmed non-fat milk in TBST for 1 h at room temperature, the membranes were incubated overnight at 4°C with specific primary antibodies (1:1000) overnight. Following three washes in TBST for 7–10 min each, the membranes were incubated with the secondary antibodies for a further 1 h at room temperature. Following three 5-min washes in TBST, the protein bands were visualized using chemiluminescence detection on autoradiographic film. The quantification was performed by measuring the intensity of the signals using Image-Pro Plus. The expression levels of each protein were normalized to those of the GAPDH protein.
Oxidative Phosphorylation and Glycolysis Assay
The intact cellular oxygen consumption rate (OCR) and extracellular acidification rate (ECAR) were measured in real-time using the Seahorse XF96 Extracellular Flux Analyser (Seahorse Bioscience, North Billerica, MA, USA). In short, 1.0 ×104 of PANC-1 and CFPAC-1 cells were seeded into 96-well cell plates and incubated overnight at 37 °C, 5% CO2. Both cells were pretreated with or without different concentrations of BI for 24 hrs. Simultaneous, the calibration plates were incubated overnight at 37 °C in a non-CO2 incubator. Then, both cell media were replaced with assay medium. Once the probe calibration was completed, the probe plate was replaced by the cell plate. The analyzer plotted the value of OCR followed by injection of the compounds sequentially as follows: oligomycin (inhibitor of ATP synthase; 2.5 μM), FCCP (uncoupler of OXPHOS; 2 μM), rotenone (inhibitor of complex I; 0.25 μM) and anti-mildew A (inhibitor of complex III; 0.25 μM) (n=8). ECAR was evaluated after continuous injection of glucose (10 mM), oligomycin (1 μM) and 2-DG (50 mM) (n = 8). After completing the test, the BCA Protein Assay Kit was performed to determine protein concentration to normalize OCR and ECAR according to the manufacturer’s instructions.
Immunofluorescence Assay
The PANC-1 cells treated by Baohuoside I (50 µM) and/or CC (10 µM) for 24 h were fixed with 4% formaldehyde (Sigma-Aldrich, Missouri, US) for 30 min and were permeabilized in 0.1% Triton X-100 (Sigma-Aldrich, Missouri, US) for 10 min. The fixed cells were washed with 1X PBS, blocked with 2% BSA in 1X PBS for 30 min at room temperature (RT), and incubated with the antibody of Ki67 (1:1000) overnight at 4 °C. After washing, the cells were incubated with Cora Lite 488 conjugated Affinipure second antibody (1:1000) in PBST containing 1% BSA for 1 hr. After washing twice, the nuclei of the cells were stained with 4,6-diamidino-2-phenylindole (DAPI; Sigma-Aldrich, Missouri, US) and then were visualized under a FLUOVIEW FV10i confocal microscope (Olympus, Tokyo, Japan).
Statistical Analysis
All data were obtained from independent experiments and are presented as mean ± standard deviation. P<0.05 was considered to indicate a statistically significant difference. Statistical analysis was performed using SPSS 18.0 (IBM, Armonk, USA) and GraphPad Prism 6.0 (GraphPad Software Inc., San Diego, USA). One-way ANOVA and the Student–Newman Keuls test were used to compare the mean values of each pair of groups. The analysis between pairs was performed using Fisher’s Least Significant Difference method provided the results of the analysis of variance were significant.
Results
Inhibition of Cell Proliferation of Pancreatic Cancer Cells by Baohuoside I
The antitumor effect of Baohuoside I was assessed by examining the inhibition of proliferation of PANC-1 and CFPAC-1 cells by CCK8, label-free RTCA and colony formation assays. As demonstrated in Figure 1A–C, the pancreatic cancer cells viability rate of Baohuoside I-treated cells was significantly decreased compared with hTERT-HPNE cells. The half-maximal inhibitory concentration values of Baohuoside I for hTERT-HPNE, PANC-1 and CFPAC-1 cells at 24 h were 51, 38 and 35 µM, respectively (Figure 1A–C). The antitumor effect of Baohuoside I on PANC-1 (Figure 1D) and CFPAC-1 (Fig, 1E) cells were subsequently monitored following the treatment of Baohuoside I. Label-free RTCA indicated that the proliferative ability of PANC-1 (Figure 1D) and CFPAC-1 (Figure 1E) cells was significantly decreased following treatment with 20 and 50 μM Baohuoside I. The plate colony formation assay was performed to detect the proliferative activity of PANC-1 and CFPAC-1 cells following treatment with Baohuoside I (Figure 1F). The number of PANC-1 and CFPAC-1 cells was significantly diminished following treatment with 20 and 50 μM Baohuoside I compared with that of the DMSO group. The colony formation assay revealed that Baohuoside I potently inhibited the proliferation and colony formation of CFPAC‑1 cells. These results indicated that Baohuoside I treatment inhibited the proliferation of PANC-1 and CFPAC-1 cells in a dose-dependent manner.
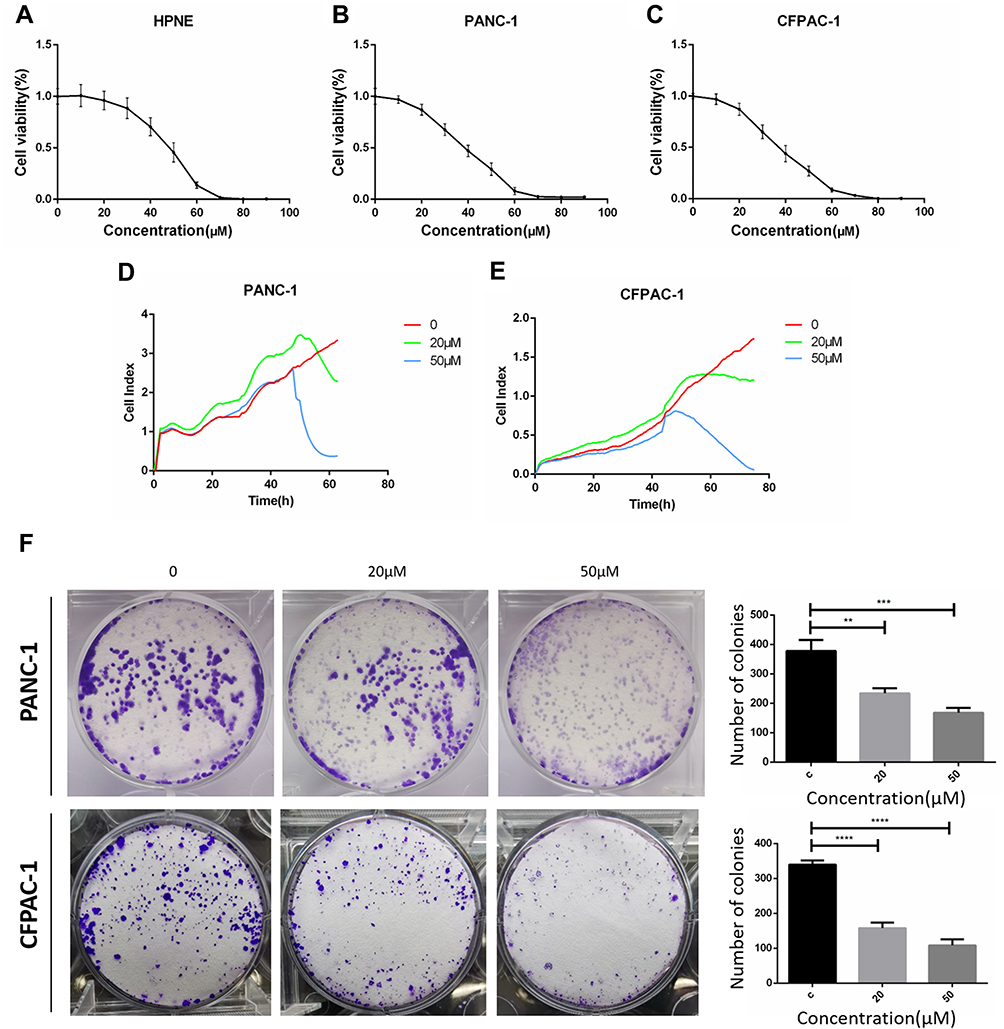 |
Figure 1 Baohuoside I inhibit hTERT-HPNE, PANC-1 and CFPAC-1 cells proliferation. CCK8 assay following hTERT-HPNE (A), PANC-1 (B) and CFPAC-1 (C) cells incubated with 10μM, 20μM, 30μM, 40μM, 50μM, 60μM, 70μM, 80μM, 90μM Baohuoside I or an equal volume of DMEM medium for 24h. Label-free Real-time Cellular Analysis (RTCA) following PANC-1 (D) and CFPAC-1 (E) cells incubated with Baohuoside I (20μM, 50μM) or an equal volume of DMEM medium for 24h. (F) Colony formation assay following PANC-1 and CFPAC-1 cells incubated with Baohuoside I (20μM, 50μM) or an equal volume of DMEM medium for 24h. The pancreatic cancer cells' viability rate in Baohuoside I-treated cells was significantly decreased compared with hTERT-HPNE cells. Baohuoside I significantly inhibited proliferation in pancreatic cancer, and the inhibition is concentration dependent. Data are presented as mean ± SD, N = 3; **P<0.01; ***P<0.001; ****P<0.0001, compared with control. |
Inhibitory Effects of Baohuoside I on the Invasive and Migratory Activity of Pancreatic Cancer
Subsequently, cell invasion was assessed using Transwell assays and the ability of cell migration was measured by a wound-healing assay following concomitant treatment with Baohuoside I. The Transwell experiments demonstrated that PANC-1 and CFPAC-1 cells containing DMSO exhibited potent invasive ability (Figure 2A). Treatment of the cells with Baohuoside I markedly inhibited cell invasion compared with that noted in the control group. The migratory activity of the cells was gradually attenuated following treatment with 0, 20 and 50 μM Baohuoside I. The wound healing assay revealed that Baohuoside I significantly inhibited PANC‑1 and CFPIC-1 cell migration (Figure 2A). These results indicated that Baohuoside I treatment inhibited the invasion and migration of PANC-1 and CFPAC-1 cells in a dose-dependent manner.
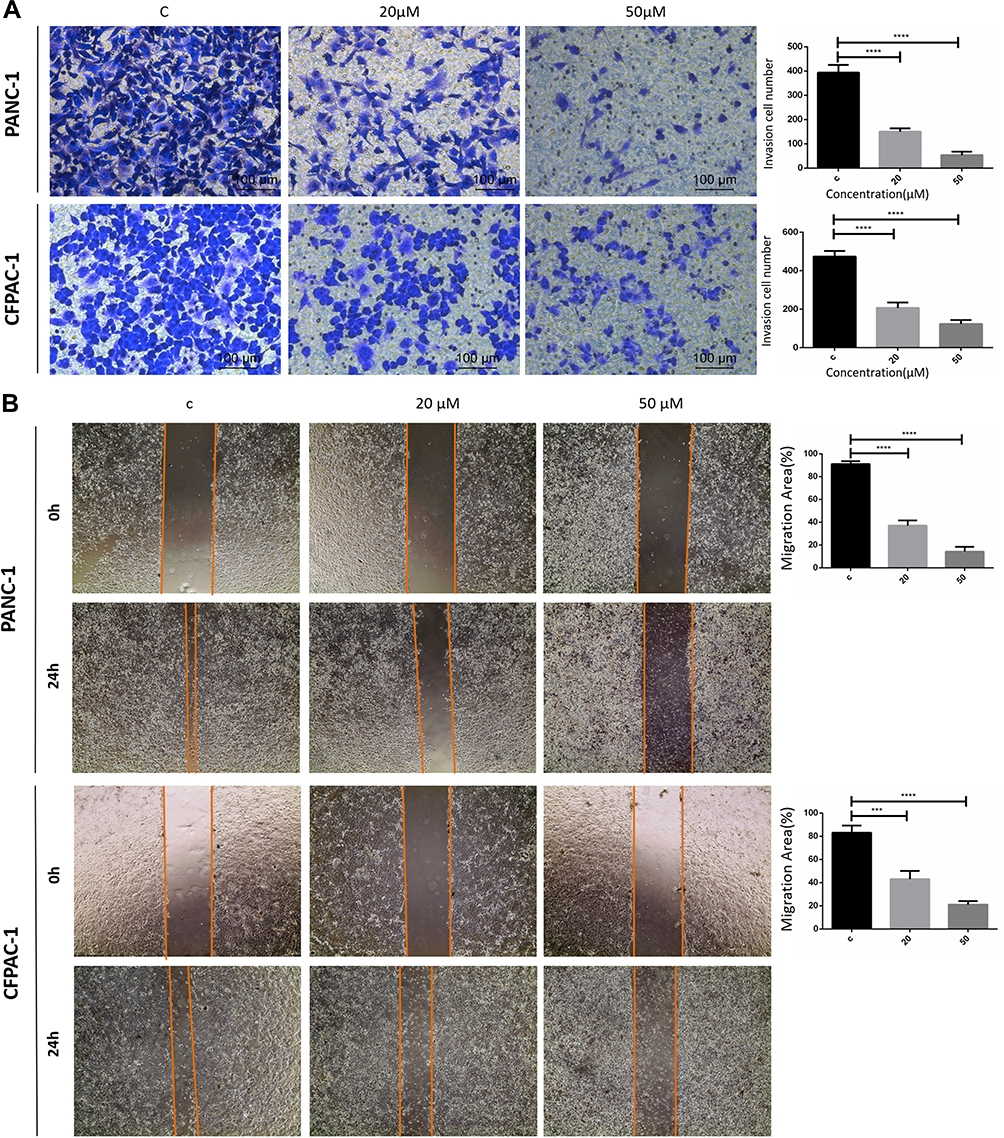 |
Figure 2 Baohuoside I inhibit PANC-1 and CFPAC-1 invasion and migration. (A) Transwell assay following PANC-1 and CFPAC-1 cells incubated with Baohuoside I (20μM, 50μM) or an equal volume of DMEM medium for 24h. (B) Wound healing assay following PANC-1 and CFPAC-1 cells incubated with Baohuoside I (20μM, 50μM) or an equal volume of DMEM medium for 24h. Baohuoside I significantly inhibited invasion and migration in PANC-1 and CFPAC-1 cells of pancreatic cancer. And the invasion and migration inhibited by Baohuoside I is concentration dependent. Data are presented as mean ± SD, N = 3; ***P<0.001; ****P<0.0001, compared with control. |
Baohuoside I Induces Pancreatic Cancer Cell Apoptosis
To investigate the effects of Baohuoside I on the induction of pancreatic cancer cell apoptosis, PANC-1 and CFPAC-1 cells were treated with Baohuoside I at a concentration of 0, 20 or 50 µM for 24 h. The induction of apoptosis was evaluated using the Annexin V-FITC/PI assay. The apoptotic assay indicated that Baohuoside I treatment resulted in a considerable increase in the percentage of apoptotic cells compared with that in the untreated cells. Specifically, the induction of apoptosis was increased from 0.024% to 4.97% in PANC-1 cells and from 2.13% to 8.42% in CFPAC-1 cells (Figure 3). The increase in the concentration of Baohuoside I was accompanied with a concomitant increase in the percentage of apoptotic PANC-1 and CFPAC-1 cells. The results revealed that Baohuoside I treatment promoted apoptosis of PANC-1 and CFPAC-1 cells in a dose-dependent manner.
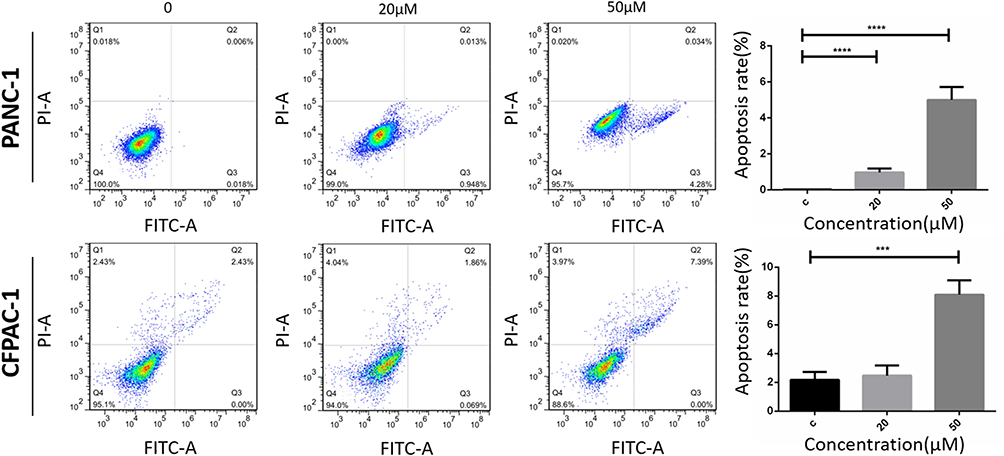 |
Figure 3 Baohuoside I promote PANC-1 and CFPAC-1 apoptosis. Flow cytometry for apoptosis [apoptosis ratio was calculated as (Q2+Q3)/(Q1+Q2+Q3+Q4)] of PANC-1 and CFPAC-1 cells incubated with Baohuoside I (20μM, 50μM) or an equal volume of DMEM medium for 24h. Baohuoside I significantly promoted apoptosis in PANC-1 and CFPAC-1 cells of pancreatic cancer. And the apoptosis induced by PANC-1 and CFPAC-1 is concentration dependent. Data are presented as mean ± SD, N = 3; ***P<0.001; ****P<0.0001, compared with control. |
Baohuoside I Suppresses Glycolysis in Pancreatic Cancer Cells
To explore the effect of Baohuoside I on the cellular bioenergetics of pancreatic cancer cell lines, the ECAR of PANC-1 and CFPAC-1 cells was analyzed following treatment with 0, 20 or 50 µM Baohuoside I using the extracellular flux analyzer. Overall, the treatment of cells with Baohuoside I resulted in a significant reduction of the aerobic glycolytic rate in both PANC-1 and CFPAC-1 cells (Figure 4A and B). In addition, various parameters of the aerobic glycolytic rate were analyzed and the results demonstrated that low doses of Baohuoside I led to a decrease in the maximal glycolytic rate and the glycolytic activity in PANC-1 cells. Moreover, Baohuoside I reduced basal and maximum levels of glycolysis and inhibited glucose metabolism in CFPAC-1 cells. High doses of compound treatment resulted in a significant reduction of four specific indices of aerobic glycolysis (Figure 4C–F). These data suggested that Baohuoside I treatment suppressed glycolysis in pancreatic cancer cells in a dose-dependent manner.
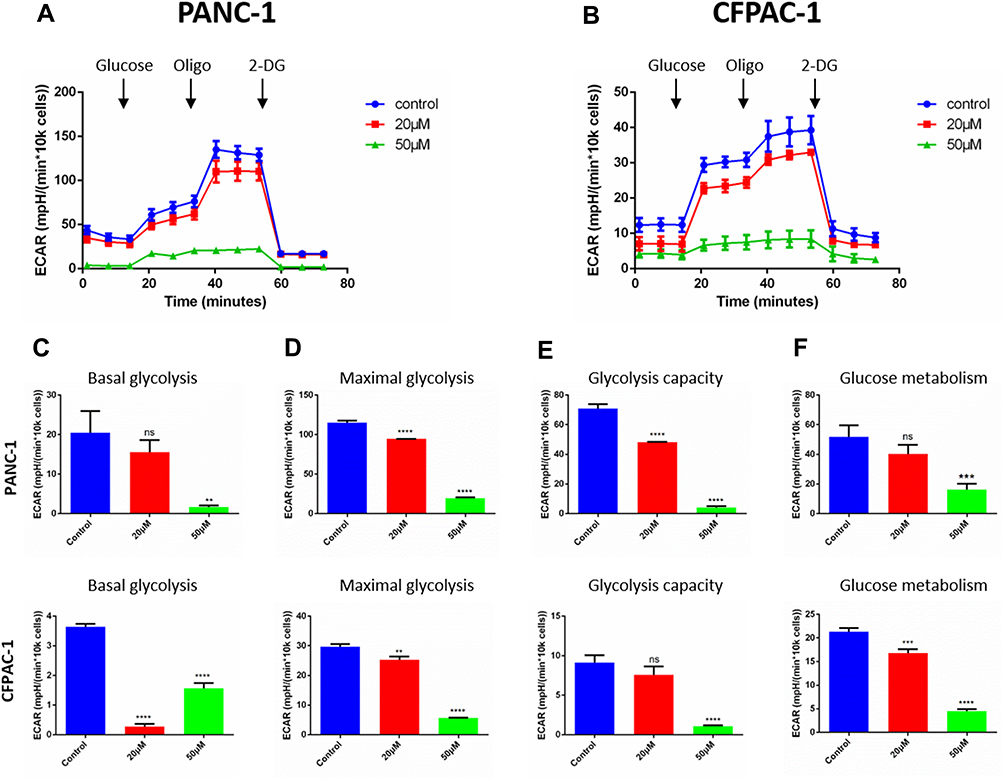 |
Figure 4 Baohuoside I reduces glycolysis in pancreatic cancer cells. (A and B) Overall ECAR curve of PANC-1and CFPAC-1 cells treated with or without Baohuoside I (0, 20, 50 mM) for 24 h. Injection order: glucose (10mM), oligomycin (1 μM), and 2-DG (50 mM) for both PANC-1 and CFPAC-1 cells. (C) Basal glycolysis. (D) Maximal glycolysis. (E) Glycolysis capacity. (F) Glycolysis metabolism. Data are presented as the mean ±SD (ns, not significant, **P < 0.01; ***P<0.001; ****P<0.0001). The representative images represent the mean ECAR ± SD of eight replicates. |
Baohuoside I Inhibits Mitochondrial Respiration in Pancreatic Cancer Cells
To further investigate the effects of Baohuoside I on pancreatic cancer cell bioenergetics, oxidative phosphorylation was assessed by measuring cellular OCRs. The results indicated that the OCR was significantly reduced following treatment of PANC-1 and CFPAC-1 cell lines with 0, 20 or 50 µM Baohuoside I (Figure 5A and B). The indices of mitochondrial respiration were also assessed. The data indicated that low doses of Baohuoside I resulted in the reduction of maximal respiration in the PANC-1 and CFPAC-1 cell lines. However, the ATP production was not affected and high doses of Baohuoside I led to a dramatic decrease of basal respiration, maximal respiration and ATP production (Figure 5C–E). The data indicated that Baohuoside I treatment led to a suppression of mitochondrial respiration in a dose-dependent manner.
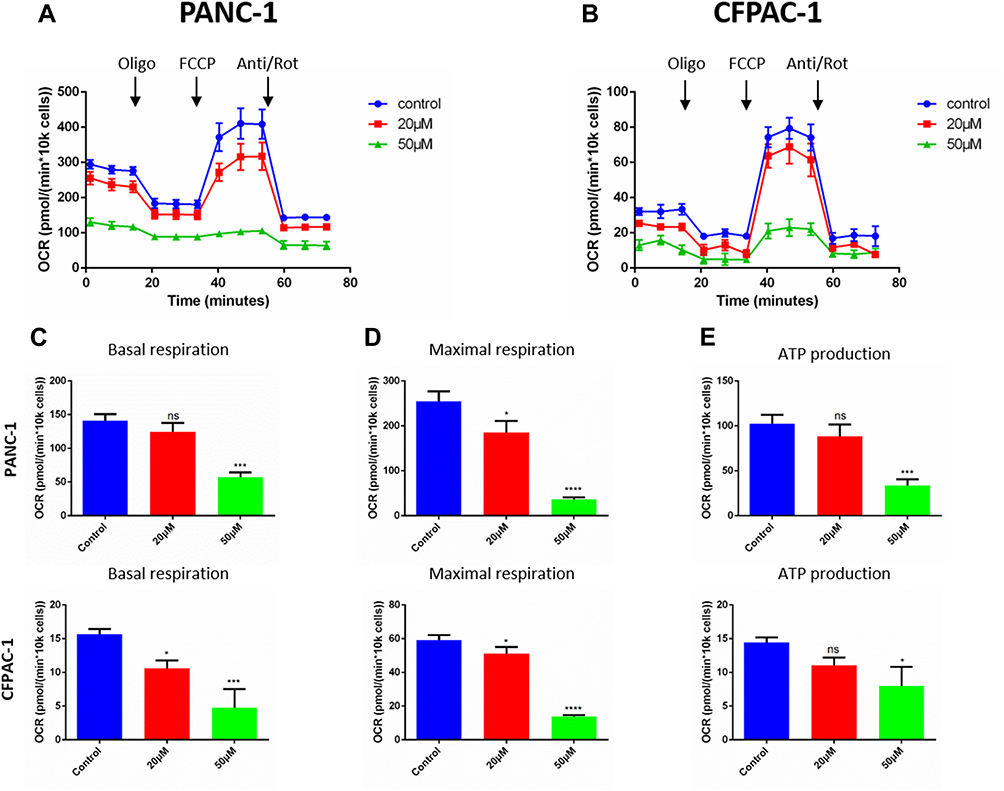 |
Figure 5 Baohuoside I reduces mitochondrial respiration in pancreatic cancer cells. (A and B) Overall oxygen consumption rate (OCR) curve of PANC-1 and CFPAC-1 cells treated with or without Baohuoside I (0, 20, 50μM) for 24 h with the Seahorse XF96 Extracellular Flux Analyzer. Injection order: oligomycin (2.5 μM), FCCP (2 μM), and rotenone (0.25 μM), and antimycin A (0.25 μM) for both PANC-1 and CFPAC-1 cells. The overall OCR curves were plotted as the mean OCR ± SD of eight replicates. (C) Basal respiration, (D) maximal respiration, and (E) ATP production were assessed, respectively. Data are presented as the mean ± SD (ns, not significant, *P < 0.05; ***P<0.001; ****P<0.0001). |
Induction of Cancer Cell Apoptosis by Baohuoside I Depends on the mTOR/S6K1 and Caspase/Bcl2/Bax Apoptotic Signaling Pathways
To further investigate the potential mechanism of Baohuoside I-induced apoptosis, the expression of key apoptotic proteins was detected by Western blot analysis. Baohuoside I treatment increased the expression levels of caspase-3, caspase-8 and Bax, whereas the expression levels of the anti-apoptotic protein Bcl-2 were downregulated, which further confirmed that PANC-1 and CFPAC-1 cell growth was inhibited by the induction of apoptosis (Figure 6). Baohuoside I treatment significantly increased the expression levels of LC-3B, whereas the expression levels of the protein p62 were downregulated, suggesting the induction of autophagy in both PANC-1 and CFPAC-1 cells. Moreover, p-mTOR was downregulated, while p-AMPK was upregulated by Baohuoside I treatment in both PANC-1 and CFPAC-1 cells, indicating that the AMPK/mTOR signaling was involved in Baohuoside I-induced autophagy and apoptosis. In addition, treatment of PANC-1 and CFPAC-1 cells with Baohuoside I downregulated the expression levels of p-S6K1, suggesting the inhibition of protein synthesis. The data suggest that Baohuoside I treatment may promote AMPK activation, suppress mTOR activation, induce autophagy and inhibit protein synthesis in both PANC-1 and CFPAC-1 cells, resulting in the inhibition of proliferation and induction of apoptosis in pancreatic cancer.
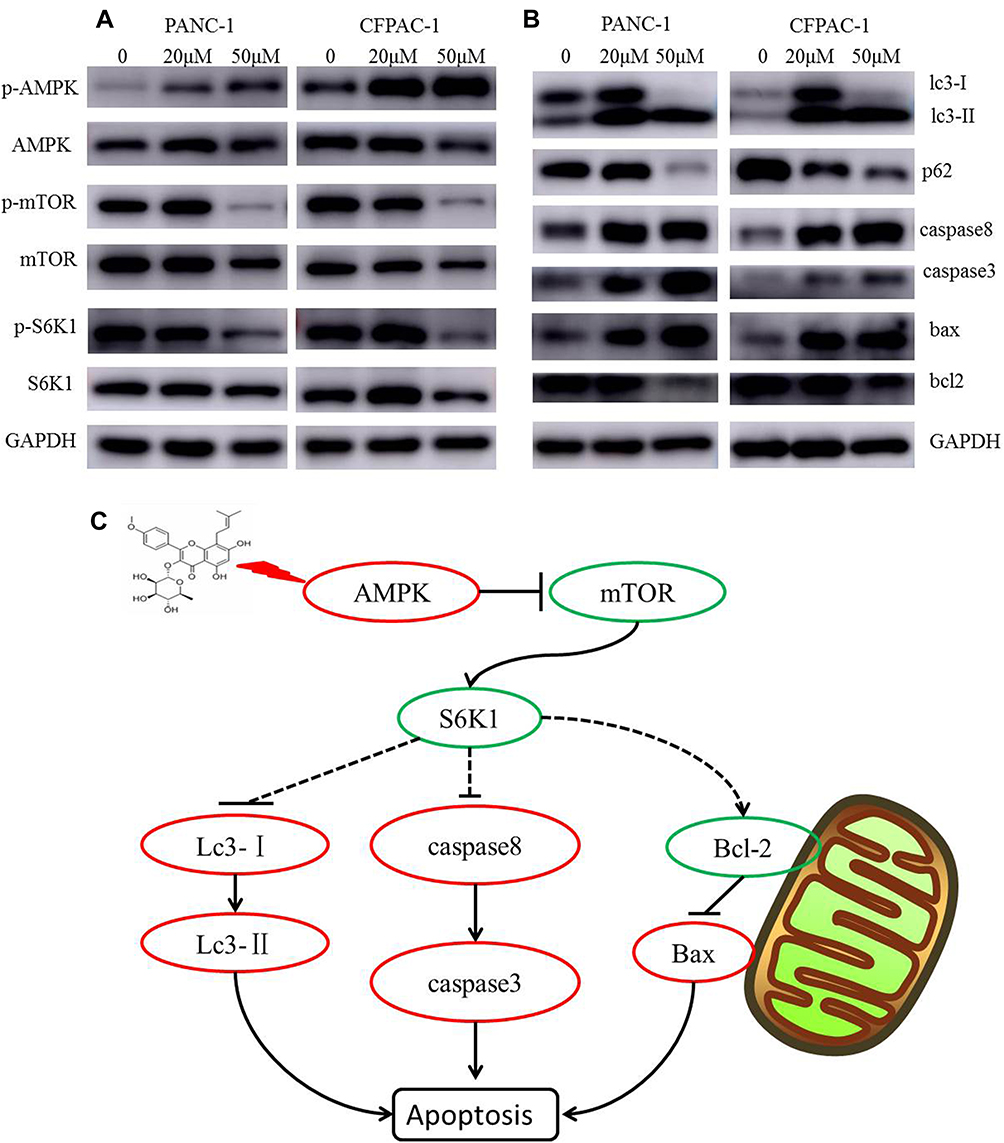 |
Figure 6 Baohuoside I inducing apoptosis is dependent on mTOR/S6K1-Caspases/Bcl2/Bax apoptotic signaling. After 24 h treatment with Baohuoside I (20μM, 50μM), (A and B) the expression of AMPK/mTOR signaling pathway autophagy and apoptosis‐related proteins was detected by Western blotting analysis. (C) Mechanism of Action of Baohuoside I on pancreatic cancer cells. |
Compound C Restored the Inhibition of Proliferation Induced by Baohuoside I via AMPK Suppression
To further determine whether Baohuoside I suppress the proliferation in pancreatic cancer via overexpressing AMPK, we pretreated with Compound C (10 µM), inhibitor of AMPK, in PANC-1 cells and then treated with Baohuoside I (50 µM). As shown in Figure 7, the pretreatment of Compound C down-regulated the expression of p-AMPK and up-regulated the expression the p-mTOR in PANC-1 cells treated with Baohuoside I (Figure 7D). Moreover, the results of colony formation assays illustrated that Compound C partly restored the inhibitory effect of Baohuoside I on PANC-1 cells proliferation (Figure 7A). Furthermore, the RTCA results also indicated that the Baohuoside I induced suppressing effect of proliferation was partially reversed by Compound C (Figure 7C). Last, Compound C was found to attenuate the suppression expression of the cell proliferation marker Ki-67 induced by Baohuoside I in PANC-1 cells (Figure 7E). The above results suggesting that Compound C reversed Baohuoside I induced inhibition of proliferation of pancreatic cancer cells through AMPK suppression.
 |
Figure 7 Compound C reversed Baohuoside I induced inhibition of proliferation of pancreatic cancer cells through AMPK suppression. (A and B) The colony formation assays showed that Compound C (10μM) restored the inhibitory effect of Baohuoside I (50μM) on PANC-1 cells proliferation. (C) The RTCA showed that the Baohuoside I induced suppressing effect of proliferation was reversed by Compound C. (D) the pretreatment of Compound C down-regulated the expression of p-AMPK and up-regulated the expression of the p-mTOR in PANC-1 cells treated with Baohuoside I. (E) Compound C was found to attenuate the suppression expression of the cell proliferation marker Ki-67 induced by Baohuoside I in PANC-1 cells. Data are presented as the mean ± SD (***P < 0.001; ****P<0.0001). Abbreviations: BI, Baohuoside I; C, Control; CC, Compound C. |
Discussion
The process of developing specific drugs for pancreatic cancer has a slow progression, due to multiple malignant behaviors of pancreatic cancer. In the clinic, 5-fluorouracil and gemcitabine alone or in combination (such as with CD40) are commonly used for pancreatic cancer therapy.13,14 However, the therapeutic results are not ideal since these drugs exhibit severe toxicity by affecting normal cell DNA synthesis. Moreover, these drugs cannot effectively inhibit the malignant activities of pancreatic cancer cells, including rapid proliferation, high invasion and migration.13–15 In addition, the treatment response rate of Gemcitabine in pancreatic cancer patients is only 12%.16 Less than 25% of the pancreatic cancer patients benefit from Gemcitabine treatment and the median overall survival is only 6 months.17
Baohuoside I, a flavonoid extracted from Herba Epimedii, has been verified to have the anti-tumour activity in broad-spectrum of human malignancies. For instance, Baohuoside I treatment was demonstrated to induce apoptosis in human non-small cell lung cancer.12 Baohuoside I suppresses cell proliferation and migration in melanoma.18 In the present study, Baohuoside I effectively inhibited the pancreatic cancer malignant phenotype as determined by acquired (PANC-1) and idiopathic (CFPAC-1) pancreatic cancer cell line models, which suggested that as a valuable therapeutic agent for this type of cancer. Notably, high invasion and migration of PANC-1 and CFPAC-1 cells was effectively suppressed by Baohuoside I treatment (Figure 2). High invasive and migratory activity of pancreatic cancer cells results in incident multiple organ metastasis of pancreatic tumors that further leads to uncontrolled cancer spread.6,19 Despite the severity of this disease, current medicinal drugs cannot effectively limit pancreatic cancer cell growth and metastasis. Moreover, gemcitabine resistance has been reported in pancreatic cancer20,21 and Baohuoside I may be a potential alternative medicine.
Defects in inducing cancer cell apoptosis often lead to the failure of several potential anticancer drugs. Lowe et al22 highlighted the importance of drug-induced apoptosis in tumor therapy. In clinical thermotherapy, tumor cells evade apoptosis resulting in drug resistance.22,23 In addition to inhibiting the aggressive phenotypes of acquired and idiopathic pancreatic cancer cells, Baohuoside I significantly induced pancreatic cell apoptosis even at low concentration levels (Figure 3). Moreover, drug-induced cancer cell apoptosis prevents uncontrolled expansion and invasion with minimal side effects.24
The metabolism of tumor cells is distinct from that of normal cells and is mainly dependent on glycolysis.25 The core energy regulatory members in cancer cells include AMPK (a low energy sensor activated by low glucose or low ATP) and mTOR (a high-energy sensor activated by high glucose or high ATP).26,27 In the present study, Baohuoside I activated the phosphorylation of AMPK and inhibited phosphorylation of mTOR and its downstream target S6K1 (Figure 6). The balance between AMPK and mTOR maintains intracellular ATP metabolism and the uneven activation of either of these pathways results in the initiation of apoptosis.28,29 Baohuoside I may block the abnormal high-energy demand of pancreatic cancer cells and subsequently induce apoptosis. Regarding the link between metabolism alteration and apoptosis, previous studies have shown that mTOR can induce caspase-dependent apoptosis.30 The findings reported in the present study were consistent with these reports and confirmed the activation of the pro-apoptotic family members (caspase-3/8 and Bax) and the inhibition of the anti-apoptotic protein Bcl-2 (Figure 6).
Autophagy and apoptosis are vitally regulated processes underlying cell and tissue homeostasis and disease.31 Autophagy promotes apoptosis through the degradation of anti-apoptotic and cell-protective factors.32 In addition, autophagy also determines the ways of cell death by serving as a role of conversion between apoptosis and necroptosis.33 Similarly, the removal or functional inhibition of crucial proteins from the apoptotic pathway can inhibit autophagy.34 Consistent with these reports, our results also testified the interplay between autophagy and apoptosis (Figure 6).
In conclusion, the present study demonstrated that Baohuoside I inhibited the malignant phenotype of both acquired (PANC-1) and idiopathic (CFPAC-1) pancreatic cancer cells. The suppression of pancreatic cancer cell growth by Baohuoside I may be mediated via the mTOR/S6K1 and caspase/Bcl2/Bax signaling pathways in order to activate the induction of apoptosis.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Moore MJ, Goldstein D, Hamm J, et al. Erlotinib plus gemcitabine compared with gemcitabine alone in patients with advanced pancreatic cancer: a Phase III trial of the National Cancer Institute of Canada Clinical Trials Group. J Clin Oncol. 2007;25:1960–1966. doi:10.1200/jco.2006.07.9525
2. Gerlinger M, Rowan AJ, Horswell S, et al. Intratumor heterogeneity and branched evolution revealed by multiregion sequencing. N Engl J Med. 2012;366:883–892. doi:10.1056/NEJMoa1113205
3. Feldmann G, Dhara S, Fendrich V, et al. Blockade of hedgehog signaling inhibits pancreatic cancer invasion and metastases: a new paradigm for combination therapy in solid cancers. Cancer Res. 2007;67:2187–2196. doi:10.1158/0008-5472.can-06-3281
4. Hwang RF, Moore T, Arumugam T, et al. Cancer-associated stromal fibroblasts promote pancreatic tumor progression. Cancer Res. 2008;68:918–926. doi:10.1158/0008-5472.can-07-5714
5. Deer EL, González-Hernández J, Coursen JD, et al. Phenotype and genotype of pancreatic cancer cell lines. Pancreas. 2010;39:425–435. doi:10.1097/MPA.0b013e3181c15963
6. Stahle M, Veit C, Bachfischer U, et al. Mechanisms in LPA-induced tumor cell migration: critical role of phosphorylated ERK. J Cell Sci. 2003;116:3835–3846. doi:10.1242/jcs.00679
7. Duxbury MS, Ito H, Zinner MJ, Ashley SW, Whang EE. EphA2: a determinant of malignant cellular behavior and a potential therapeutic target in pancreatic adenocarcinoma. Oncogene. 2004;23:1448–1456. doi:10.1038/sj.onc.1207247
8. Handra-Luca A, Vidaud D, Vullierme MP. et al. Plexiform neurofibroma mimicking a pancreatic cystic tumour. Gut. 2005;54:1658–1659. doi:10.1136/gut.2005.074609
9. Jiang J, Zhao B-J, Song J, Jia X-B. Pharmacology and clinical application of plants in epimedium L. Chin Herbal Med. 2016;8:12–23. doi:10.1016/S1674-6384(16)60003-5
10. Li SY, Teh BS, Seow WK, Li F, Thong YH. Effects of the plant flavonoid baohuoside-1 on cancer cells in vitro. Cancer Lett. 1990;53:175–181. doi:10.1016/0304-3835(90)90211-f
11. Kim B, Park B. Baohuoside I suppresses invasion of cervical and breast cancer cells through the downregulation of CXCR4 chemokine receptor expression. Biochemistry. 2014;53:7562–7569. doi:10.1021/bi5011927
12. Song J, Shu L, Zhang Z, et al. Reactive oxygen species-mediated mitochondrial pathway is involved in Baohuoside I-induced apoptosis in human non-small cell lung cancer. Chem Biol Interact. 2012;199:9–17. doi:10.1016/j.cbi.2012.05.005
13. Vonderheide RH, Bajor DL, Winograd R, et al. CD40 immunotherapy for pancreatic cancer. Cancer Immunol Immunother. 2013;62:949–954. doi:10.1007/s00262-013-1427-5
14. Heinemann V. Gemcitabine: progress in the treatment of pancreatic cancer. Oncology. 2001;60:8–18. doi:10.1159/000055290
15. Wang WB, Yang Y, Zhao YP, Zhang TP, Liao Q, Shu H. Recent studies of 5-fluorouracil resistance in pancreatic cancer. World J Gastroenterol. 2014;20:15682–15690. doi:10.3748/wjg.v20.i42.15682
16. Storniolo AM, Enas NH, Brown CA, et al. An investigational new drug treatment program for patients with gemcitabine: results for over 3000 patients with pancreatic carcinoma. Cancer. 1999;85:1261–1268. doi:10.1002/(ISSN)1097-0142
17. Burris HA
18. Peng YG, Zhang L. Baohuoside-I suppresses cell proliferation and migration by up-regulating miR-144 in melanoma. Pharm Biol. 2018;56:43–50. doi:10.1080/13880209.2017.1418391
19. Singh AP, Moniaux N, Chauhan SC, Meza JL, Batra SK. Inhibition of MUC4 expression suppresses pancreatic tumor cell growth and metastasis. Cancer Res. 2004;64:622–630. doi:10.1158/0008-5472.CAN-03-2636
20. Liang C, Shi S, Meng Q, et al. Complex roles of the stroma in the intrinsic resistance to gemcitabine in pancreatic cancer: where we are and where we are going. Exp Mol Med. 2017;49:e406. doi:10.1038/emm.2017.255
21. Binenbaum Y, Na’ara S, Gil Z. Gemcitabine resistance in pancreatic ductal adenocarcinoma. Drug Resistance Updates. 2015;23:55–68. doi:10.1016/j.drup.2015.10.002
22. Johnstone RW, Ruefli AA, Lowe SW. Apoptosis: a link between cancer genetics and chemotherapy. Cell. 2002;108:153–164. doi:10.1016/s0092-8674(02)00625-6
23. Hengartner MO. The biochemistry of apoptosis. Nature. 2000;407:770–776. doi:10.1038/35037710
24. Evan GI, Vousden KH. Proliferation, cell cycle and apoptosis in cancer. Nature. 2001;411:342–348. doi:10.1038/35077213
25. Fadaka A, Ajiboye B, Ojo O, et al. Biology of glucose metabolization in cancer cells. J Oncological Sci. 2017;3:45–51. doi:10.1016/j.jons.2017.06.002
26. Ji L, Li L, Qu F, et al. Hydrogen sulphide exacerbates acute pancreatitis by over-activating autophagy via AMPK/mTOR pathway. J Cell Mol Med. 2016;20:2349–2361. doi:10.1111/jcmm.12928
27. Mihaylova MM, Shaw RJ. The AMPK signalling pathway coordinates cell growth, autophagy and metabolism. Nat Cell Biol. 2011;13:1016–1023. doi:10.1038/ncb2329
28. Su -C-C, Hsieh K-L, Liu P-L, et al. AICAR induces apoptosis and inhibits migration and invasion in prostate cancer cells through an AMPK/mTOR-dependent pathway. Int J Mol Sci. 2019;20:1647. doi:10.3390/ijms20071647
29. Liu M-W, Wei R, Su M-X, et al. Effects of Panax notoginseng saponins on severe acute pancreatitis through the regulation of mTOR/Akt and caspase-3 signaling pathway by upregulating miR-181b expression in rats. BMC Complement Altern Med. 2018;18:51. doi:10.1186/s12906-018-2118-8
30. Seitz C, Hugle M, Cristofanon S, Tchoghandjian A, Fulda S. The dual PI3K/mTOR inhibitor NVP-BEZ235 and chloroquine synergize to trigger apoptosis via mitochondrial-lysosomal cross-talk. Int J Cancer. 2013;132:2682–2693. doi:10.1002/ijc.27935
31. Fuchs Y, Steller H. Programmed cell death in animal development and disease. Cell. 2011;147:742–758. doi:10.1016/j.cell.2011.10.033
32. Doherty J, Baehrecke EH. Life, death and autophagy. Nat Cell Biol. 2018;20:1110–1117. doi:10.1038/s41556-018-0201-5
33. Sakamaki JI, Ryan KM. Autophagy determines the path on the TRAIL to death. Dev Cell. 2016;37:291–293. doi:10.1016/j.devcel.2016.05.010
34. Maiuri MC, Zalckvar E, Kimchi A, Kroemer G. Self-eating and self-killing: crosstalk between autophagy and apoptosis. Nat Rev Mol Cell Biol. 2007;8:741–752. doi:10.1038/nrm2239
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.