")
Back to Journals » Cancer Management and Research » Volume 11
Atopobium vaginae And Porphyromonas somerae Induce Proinflammatory Cytokines Expression In Endometrial Cells: A Possible Implication For Endometrial Cancer?
Authors Caselli E , Soffritti I, D'Accolti M, Piva I, Greco P, Bonaccorsi G
Received 31 May 2019
Accepted for publication 31 August 2019
Published 23 September 2019 Volume 2019:11 Pages 8571—8575
DOI https://doi.org/10.2147/CMAR.S217362
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Yong Teng
Elisabetta Caselli,1 Irene Soffritti,1 Maria D’Accolti,1 Isabella Piva,2 Pantaleo Greco,2 Gloria Bonaccorsi2
1Section of Microbiology and Medical Genetics, Department of Medical Sciences, University of Ferrara, Ferrara, Italy; 2Section of Obstetrics and Gynecology, Department of Morphology, Surgery and Experimental Medicine, University of Ferrara, Ferrara, Italy
Correspondence: Elisabetta Caselli
Section of Microbiology and Medical Genetics, Department of Medical Sciences, University of Ferrara, Via Luigi Borsari 46, Ferrara 44121, Italy
Tel +39 0532 455387
Fax +39 0532 974470
Email [email protected]
The female lower vaginal tract has long been known to have an active microbiota, with Lactobacilli genus representing the prevalent species, and alterations in the vaginal microbiota are known to play a role in different pathological conditions, including bacterial vaginosis, sexually transmitted diseases, HPV persistence, and cervical cancer.
By contrast, the uterus was considered sterile until recently, when the advent of next-generation sequencing (NGS) technologies identified a unique uterine microbiota differing from that of the vagina.1
The uterine bacterial load is estimated to be 100–10,000-times lower than that of the vaginal microbiome, and, contrary to the vaginal and cervix microbiota, uterine bacteria grow in mildly alkaline conditions, contrasting to the Lactobacillus-dominated low pH environment of the vagina.2
The female lower vaginal tract has long been known to have an active microbiota, with Lactobacilli genus representing the prevalent species, and alterations in the vaginal microbiota are known to play a role in different pathological conditions, including bacterial vaginosis, sexually transmitted diseases, HPV persistence, and cervical cancer.
By contrast, the uterus was considered sterile until recently, when the advent of next-generation sequencing (NGS) technologies identified a unique uterine microbiota differing from that of the vagina.1
The uterine bacterial load is estimated to be 100–10,000-times lower than that of the vaginal microbiome, and, contrary to the vaginal and cervix microbiota, uterine bacteria grow in mildly alkaline conditions, contrasting to the Lactobacillus-dominated low pH environment of the vagina.2
The microbiota in the uterus can derive from ascension via the cervix, or migrate to the uterus via the haematogenous spread, via the sperm, and other routes (e.g., retrograde spread through the fallopian tubes or gynaecological procedures).1,2
Although the exact role and mechanisms of micro-organisms in the uterus are unclear, recent studies suggest that microbiota could affect uterine receptivity, influencing endometrium fertility.
In addition, uterine microbiota composition has been associated with several gynecological conditions, including endometriosis, dysfunctional menstrual bleeding, and cancer.3–5 In particular, the simultaneous presence of Atopobium vaginae and Porphyromonas species resulted particularly significant in endometrial cancer.5 Uterine colonization with bacterial vaginosis-associated bacteria has been suggested to promote carcinogenesis through microbiota-mediated pathophysiologic changes.6 Nevertheless, a cause-effect relationship has not been established yet. These studies provided a starting point to understand uterine physiology in health and disease, but there is currently a need for studies investigating host-microbiota interactions, to understand the impact of the micro-organisms on the local endometrial microenvironment.
Based on this, our study was aimed to analyze in vitro the effect of A. vaginae and P. somerae on endometrial cells, focusing on the expression of pro-inflammatory cytokines, being those implied in the establishment of an inflammatory environment potentially favoring the onset/progression of endometrial cancer.
To this purpose, the human HEC-1A endometrial adenocarcinoma cells (ATCC HTB-112, cultured in McCoy’s 5a Medium Modified additioned with 10% fetal bovine serum) were co-cultured in the presence or absence of the following bacterial strains: Atopobium vaginae (ATCC-BAA-55), Porphyromonas somerae (ATCC-BAA-1230) and Lactobacillus vaginalis (ATCC-49540). Prior to co-culture, each bacterial species was amplified in blood-agar plates in anaerobic conditions (Thermo Fisher Scientific, Milan, Italy), and culture conditions were tested to ascertain viability of endometrial and bacterial cells, verified respectively by cell count after Trypan Blue exclusion and CFU count on blood agar plates. Based on the obtained data (Table S1), HEC-1A cells were cultured with bacterial cells in a 5% CO2 atmosphere, a condition preserving the optimal viability of eukaryotic cells and maintaining most of bacteria alive. HEC-1A cells were thus seeded at optimal density in 6 wells plates (5x105 cells/well), and the day of infection the culture medium was replaced with complete McCoy’s medium without antibiotics for bacterial co-culture. A single bacterial colony of 2 mm of diameter (containing about 2×106 bacterial cells, as measured by CFU count after serial dilution) for each bacterial species was suspended in 1 mL of complete antibiotic-free McCoy’s medium and 250 µl of suspension were added to each well (final bacteria:human cells ratio, 10:1). Different co-culture conditions were tested, namely: control uninfected cells; cells monoinfected with single alive or killed bacterial species (obtained by 3 cycles of sonication at maximum power for 30 seconds), or with a combination of alive or killed A. vaginae and P. somerae species. Cells and cell supernatant were collected after 24, 48 and 72 hrs post infection (h.p.i.), and analyzed for proinflammatory cytokine expression and release.
Briefly, cytokine release was first analyzed in cell culture supernatant by a multi-analyte qualitative ELISA assay identifying 12 different cytokines, including IL1α, IL1β, IL2, IL4, IL6, IL8, IL10, IL12, IL17α, IFNγ, TNFα and GM-CSF (Qiagen, Hilden, Germany).
The results showed that 24 h of co-culturing with A. vaginae and P. somerae was a sufficient time to induce an evident release of proinflammatory cytokines by human endometrial cells, as judged by ELISA results obtained on cultural supernatant. Interestingly, the cytokine panel was different for A. vaginae and P. somerae compared to L. vaginalis, used as a control (Figure S1). In fact, while Lactobacillus induced only IL-8, Atopobium and/or Porphyromonas did not induce IL-8 but promoted the release of IL1α, IL1β, IL17α, and TNFα. The cytokine production was observable also at later experimental times, without significant further increase in the total amount produced, suggesting a specific kinetic of cytokine induction, or a gradual loss of bacterial ability to induce cytokines, as A. vaginae and P. somerae, are facultative and obligate anaerobes, respectively, and might survive hardly for longer times in the culture conditions used for co-culture experiments. However, killed bacteria used as a control were not inducing any cytokine release since the beginning of the experiment, confirming that alive A. vaginae and P. somerae bacteria were needed to stimulate cytokine production in endometrial cells.
Each altered cytokine, as resulted by the multi-analyte test, was subsequently analyzed by specific and quantitative individual ELISA assays, to confirm the data and quantify the amount of induced cytokines. The results (Figure 1) confirmed the data obtained for each cytokine, further evidencing the different impact of the diverse bacterial species (Lactobacillus versus Atopobium and Porphyromonas), and the absence of cytokine induction when using killed bacteria. In addition, the simultaneous exposure of HEC-1A cells to A. vaginae and P. somerae used together led to a more pronounced increase of cytokine amount, compared to what observed with the individual bacterial species. Interestingly, IL1α and IL1β upregulation is associated with many tumors including endometrial cancer,7,8 IL17α was reported to promote the proliferation of endometrial cells9 and to contribute to endometriosis thank to its induction of proinflammatory and angiogenic factors.10 TNFα is constitutively expressed in endometrial hyperplasia and cancer,11 and high levels of expression correlate with low survival in endometrial cancer patients.12
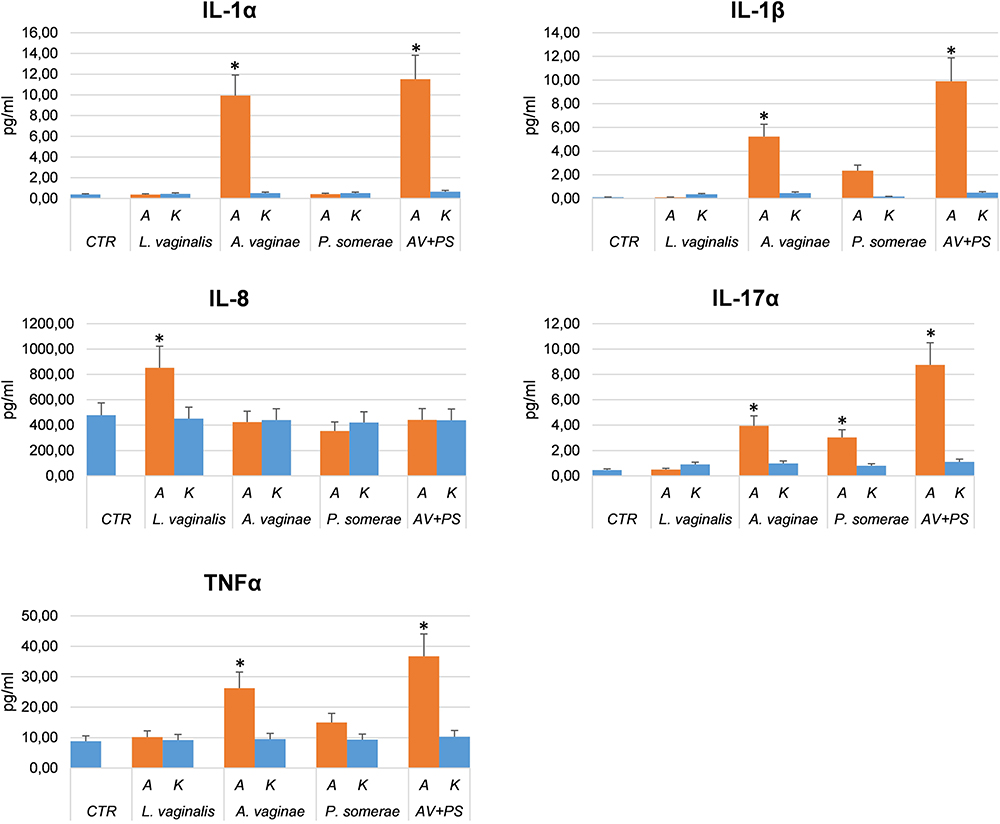 |
Figure 1 Individual cytokines altered by bacterial co-colture. Endometrial cells were cultured in the absence (control, CTR) or presence of alive or killed A. vaginae (AV), or P. somerae (PS), or L. vaginalis (LV), or a combination of A. vaginae plus P. somerae (AV+PS). Cell culture supernatants collected at 24 h.p.i. were analyzed for cytokine release by individual ELISA assay, performed for each cytokine resulted altered by the multi-cytokine ELISA panel. Results refer to triplicate samples from two independent experiments and are expressed as mean pg/mL value ± SD. Student’s t test was used for statistical analysis; a p value < 0.05 was considered as significant (indicated by asterisks). |
To ascertain whether the cytokine induction was at the transcriptional level, HEC-1A cells co-cultured with individual bacterial species were collected at 24 h.p.i. and analyzed for cytokine mRNA expression. Briefly, the total RNA was extracted from cells and retrotranscribed by RT2 First Strand kit (Qiagen, Hilden, Germany), then cytokine expression was analyzed on obtained cDNA by a specific real-time PCR microarray targeting 84 different human cytokine transcripts (Qiagen, Hilden, Germany).
Results (Figure 2) showed that all the cytokines detected in the culture supernatant of endometrial cells upon co-culture with bacteria were indeed activated at the transcriptional level, as indicated by the upregulation of the correspondent mRNAs compared to control uninfected cells or cells co-cultured with killed bacteria. In particular, IL1α and IL1β RNAs resulted respectively 6.8-fold and 8-fold more expressed in the presence of Atopobium than in controls (endometrial cells uninfected or exposed to killed bacteria) or in Lactobacillus and Porphyromonas co-cultured cells (respectively 1.1 and 1 folds with Lactobacillus, and 1 and 1.3 folds with Porphyromonas); IL8 resulted upregulated only by Lactobacillus (16 folds) and not by Atopobium/Porphyromonas; IL17α was activated only by Atopobium (9.5 folds) and Porphyromonas (8.2 folds); similarly, TNFα was upregulated by Atopobium and Porphyromonas (16.3 and 3 folds, respectively), but not by Lactobacillus (1.1 folds).
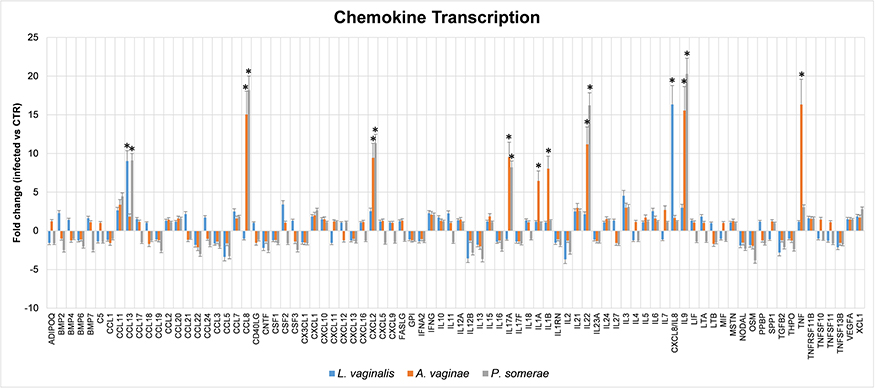 |
Figure 2 Cytokine/chemokine transcriptional induction in endometrial HEC-1A cells by bacterial co-culture. Endometrial cells were cultured in the absence (control) or presence of alive or killed A. vaginae, or P. somerae, or L. vaginalis. After 24 hrs, cultured cells were collected for total RNA extraction and subsequent analysis by qPCR microarray, capable of evidencing simultaneously transcriptional up- or down-regulation of 84 different cytokines. Results refer to triplicate samples from two independent experiments, and are expressed as mean fold-change value compared to control values ± SD. Control values, obtained in uninfected cells or in cells exposed to killed bacteria, were superimposable and corresponded to a value = 1±0.1 fold-change for all analyzed transcripts. Student’s t test with Bonferroni correction for multiple analyses was used for statistical analysis; a corrected p value < 0.05 was considered as significant (indicated by asterisks). |
Interestingly, in addition to the cytokines analyzed by ELISA and confirmed at the transcriptional level, other transcripts resulted significantly altered by co-culture with Atopobium and Porphyromonas, including CCL13 (9 folds with P. somerae only), CCL8 (15 folds with A. vaginae and 18 folds with P. somerae), CXCL2 (9 folds with A. vaginae and 11 folds with P. somerae), IL22 (11 folds with A. vaginae and 16 folds with P. somerae) and IL9 (15 folds with A. vaginae and 20 folds with P. somerae). However, CCL13 was also upregulated by Lactobacillus as well (9 folds), suggesting a nonspecific stimulation associated with the mere presence of bacteria, rather than with a specific bacterial species. CCL13 (also known as Monocyte Chemoattractant Protein 4, MCP4) can be induced by IL1 and TNFα and is indeed involved in many chronic diseases, including allergies. By contrast, the other chemokines that resulted altered by Atopobium or Porphyromonas co-culture have been reported to be associated with cancer progression. In particular, CCL8 has been associated with melanoma progression and dissemination,13 enhances breast cancer cell motility and is upregulated in tumor-associated macrophages (TAM) in breast and endometrial cancer patients, favoring tumor invasion.14 The expression of CXCL2 is increased in ovarian and endometrial carcinomas,15,16 it is induced by TNFα and induces chemoresistance.17 IL22 promotes progression in breast cancer18 and endometrial cell proliferation by stimulating CCL2 and IL8,19 IL9 is upregulated in endometriosis,20 and its expression was reported to be upregulated via IL-17α in endometrial adenocarcinoma cells,10 and to prevent apoptosis, promoting proliferation and metastasis in pancreatic cancer.21
In conclusion, our in vitro observations show for the first time that certain bacteria have the ability to induce expression of proinflammatory cytokines and chemokines by endometrial cells, and suggest that their presence in the uterine environment might be associated with the establishment of conditions promoting inflammation.
One limitation of our study is the use of a cell line, rather than endometrial primary cells derived from healthy tissue, and future studies are needed to investigate host-microbiota interactions using primary endometrial cells and different bacterial loads, to better understand the impact of microorganisms on the local endometrial microenvironment and its mechanisms. However, our preliminary observations provide a base for additional knowledge in this field, and strengthen the hypothesis of a significant role of Atopobium and Porphyromonas species in endometrial tumor onset and/or progression, providing also a starting point for future researches focusing on the impact of uterine microbiota on uterine physiology and pathologies, and hopefully potentially useful tools for diagnosis and effective clinical interventions.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Baker JM, Chase DM, Herbst-Kralovetz MM. Uterine microbiota: residents, tourists, or invaders? Front Immunol. 2018;9:208. doi:10.3389/fimmu.2018.00208
2. Chen C, Song X, Wei W, et al. The microbiota continuum along the female reproductive tract and its relation to uterine-related diseases. Nat Commun. 2017;8(1):875. doi:10.1038/s41467-017-00901-0
3. Cicinelli E, Matteo M, Tinelli R, et al. Prevalence of chronic endometritis in repeated unexplained implantation failure and the IVF success rate after antibiotic therapy. Hum Reprod. 2015;30(2):323–330. doi:10.1093/humrep/deu292
4. Pelzer ES, Willner D, Buttini M, Huygens F. A role for the endometrial microbiome in dysfunctional menstrual bleeding. Antonie Van Leeuwenhoek. 2018;111(6):933–943. doi:10.1007/s10482-017-0992-6
5. Walther-Antonio MR, Chen J, Multinu F, et al. Potential contribution of the uterine microbiome in the development of endometrial cancer. Genome Med. 2016;8(1):122. doi:10.1186/s13073-016-0368-y
6. Mert I, Walther-Antonio M, Mariani A. Case for a role of the microbiome in gynecologic cancers: clinician’s perspective. J Obstet Gynaecol Res. 2018;44(9):1693–1704. doi:10.1111/jog.13701
7. Wang Y, Wang Y, Duan X, Wang Y, Zhang Z. Interleukin-1 receptor-associated kinase 1 correlates with metastasis and invasion in endometrial carcinoma. J Cell Biochem. 2018;119(3):2545–2555. doi:10.1002/jcb.26416
8. Litmanovich A, Khazim K, Cohen I. The role of interleukin-1 in the pathogenesis of cancer and its potential as a therapeutic target in clinical practice. Oncol Ther. 2018;6(2):109–127. doi:10.1007/s40487-018-0089-z
9. Hirata T, Osuga Y, Hamasaki K, et al. Interleukin (IL)-17A stimulates IL-8 secretion, cyclooxygensase-2 expression, and cell proliferation of endometriotic stromal cells. Endocrinology. 2008;149(3):1260–1267. doi:10.1210/en.2007-0749
10. Ahn SH, Edwards AK, Singh SS, Young SL, Lessey BA, Tayade C. IL-17A contributes to the pathogenesis of endometriosis by triggering proinflammatory cytokines and angiogenic growth factors. J Immunol. 2015;195(6):2591–2600. doi:10.4049/jimmunol.1501138
11. Garcia FU, Chen HL, Yang Y, Pace JL, Hu XL, Hunt JS. Tumor necrosis factor-alpha mRNA and protein in endometrial tumors: analysis by in situ hybridization and immunocytochemistry. Hum Pathol. 1994;25(12):1324–1331. doi:10.1016/0046-8177(94)90093-0
12. Smith HO, Stephens ND, Qualls CR, et al. The clinical significance of inflammatory cytokines in primary cell culture in endometrial carcinoma. Mol Oncol. 2013;7(1):41–54. doi:10.1016/j.molonc.2012.07.002
13. Barbai T, Fejos Z, Puskas LG, Timar J, Raso E. The importance of microenvironment: the role of CCL8 in metastasis formation of melanoma. Oncotarget. 2015;6(30):29111–29128. doi:10.18632/oncotarget.5059
14. Cassetta L, Fragkogianni S, Sims AH, et al. Human tumor-associated macrophage and monocyte transcriptional landscapes reveal cancer-specific reprogramming, biomarkers, and therapeutic targets. Cancer Cell. 2019;35(4):588–602 e510. doi:10.1016/j.ccell.2019.02.009
15. Furuya M, Suyama T, Usui H, et al. Up-regulation of CXC chemokines and their receptors: implications for proinflammatory microenvironments of ovarian carcinomas and endometriosis. Hum Pathol. 2007;38(11):1676–1687. doi:10.1016/j.humpath.2007.03.023
16. Kavandi L, Collier MA, Nguyen H, Syed V. Progesterone and calcitriol attenuate inflammatory cytokines CXCL1 and CXCL2 in ovarian and endometrial cancer cells. J Cell Biochem. 2012;113(10):3143–3152. doi:10.1002/jcb.24191
17. Palacios-Arreola MI, Nava-Castro KE, Castro JI, Garcia-Zepeda E, Carrero JC, Morales-Montor J. The role of chemokines in breast cancer pathology and its possible use as therapeutic targets. J Immunol Res. 2014;2014:849720. doi:10.1155/2014/394127
18. Rui J, Chunming Z, Binbin G, Na S, Shengxi W, Wei S. IL-22 promotes the progression of breast cancer through regulating HOXB-AS5. Oncotarget. 2017;8(61):103601–103612. doi:10.18632/oncotarget.22063
19. Guo Y, Chen Y, Liu LB, et al. IL-22 in the endometriotic milieu promotes the proliferation of endometrial stromal cells via stimulating the secretion of CCL2 and IL-8. Int J Clin Exp Pathol. 2013;6(10):2011–2020.
20. Jorgensen H, Hill AS, Beste MT, et al. Peritoneal fluid cytokines related to endometriosis in patients evaluated for infertility. Fertil Steril. 2017;107(5):1191–1199 e1192. doi:10.1016/j.fertnstert.2017.03.013
21. Hu B, Qiu-Lan H, Lei RE, Shi C, Jiang HX, Qin SY. Interleukin-9 promotes pancreatic cancer cells proliferation and migration via the miR-200a/beta-catenin axis. Biomed Res Int. 2017;2017:2831056. doi:10.1155/2017/2831056
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.