Back to Journals » Drug Design, Development and Therapy » Volume 12
Astragaloside IV/lncRNA-TUG1/TRAF5 signaling pathway participates in podocyte apoptosis of diabetic nephropathy rats
Authors Lei X, Zhang L, Li Z, Ren J
Received 24 February 2018
Accepted for publication 31 May 2018
Published 6 September 2018 Volume 2018:12 Pages 2785—2793
DOI https://doi.org/10.2147/DDDT.S166525
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Qiongyu Guo
Xiao Lei, Limei Zhang, Zonglin Li, Jigang Ren
Traditional Chinese Medicine Department, Affiliated Hospital of North Sichuan Medical College, Nanchong, Sichuan Province, People’s Republic of China
Objective: This study aims to figure out the mechanism of astragaloside IV (AS-IV) in the protection of podocyte apoptosis in diabetic nephropathy (DN) rats.
Materials and methods: Streptozotocin (STZ) was used to induce diabetes in rats, and the diabetic rats were treated with 5 mg/kg/d of AS-IV for 12 weeks. Albuminuria level, relative TUG1 and TRAF5 levels, and TRAF5 and cleaved-caspase-3 protein levels were examined by ELISA, quantitative reverse transcription (qRT)-PCR, and Western blot analyses, respectively. The interaction between TUG1 and TRAF5 was confirmed by RNA pull-down and RNA precipitation. TUNEL assay was used to detect podocyte apoptosis.
Results: Compared with control rats, DN rats had higher albuminuria and TRAF5 levels and lower TUG1 level. AS-IV treatment attenuated albuminuria and TRAF5 levels and improved TUG1 level in DN rats. TUG1 was downregulated and TRAF5 was upregulated in high-glucose-treated MPC5 cells, and AS-IV ameliorated the TUG1 level. In addition, TUG1 interacted with TRAF5, and TUG1 overexpression promoted degradation of TRAF5 protein. Besides, AS-IV modulated TRAF5 expression through regulating TUG1. AS-IV decreased podocyte apoptosis via the TUG1/TRAF5 pathway. Finally, in vivo experiment proved that si-TUG1 abrogated the protective effect of AS-IV on DN.
Conclusion: AS-IV attenuated podocyte apoptosis and protected diabetic rats from DN via the lncRNA-TUG1/TRAF5 pathway.
Keywords: astragaloside IV, diabetic nephropathy, TUG1, TRAF5, albuminuria
Introduction
Diabetic nephropathy (DN) is a common and serious complication of diabetes, which leads to the end-stage renal disease (ESRD).1 Studies have shown that podocyte apoptosis played a key role in the pathogenesis of glomerulosclerosis and albuminuria in DN.2–4 It has been reported that TRAF5-mediated nuclear factor-κB (NF-κB) activation was involved in the pathogenesis of chronic kidney disease, and overexpression of TRAF5 remarkably suppressed cell viability and induced mice podocytes apoptosis.5 Therefore, in this study, we focused on TRAF5 to find effective treatments of DN.
Studies have shown that DN was regulated by abnormal lncRNAs, such as lncRNA Erbb4-IR, lncRNA PVT1, and lncRNA MALAT1.6–8 LncRNA TUG1 is a widely expressed lncRNA that participates in cancers, metabolic disorders, cardiovascular disorders, etc.9–11 Recently, Duan et al10 reported that lncRNA TUG1 was downregulated in the DN rat model, which revealed extracellular matrix accumulation in DN via the microRNA-377/PPARγ pathway. In addition, researchers found that lncRNA TUG1 could regulate mitochondrial function in podocytes in the diabetic milieu.12 What’s more, lncRNA TUG1 could directly target CUGBP elav-like family member 1 (CELF1), and TUG1 knockdown could remarkably accelerate CELF1 expression,13 which indicated that lncRNA TUG1 could target proteins. Therefore, we speculated that lncRNA TUG1 might regulate podocyte apoptosis in DN by targeting TRAF5.
Radix astragali is the root of Astragalus membranaceus Bunge that plays important roles in protecting liver, such as eliminating toxins, preventing cancers, and protecting cells from oxidative stress.14 It was one of the most commonly used natural traditional Chinese medicine in China and other Asian areas, and mainly used as a tonic, which was first recorded in Ben Cao Gang Mu.15 Astragaloside IV (AS-IV) is the main active constituent of Radix astragali that has been used to cure DN of diabetes.16 Recently, researchers have found that AS-IV could improve DN through regulating mitochondrial quality control network.3 AS-IV could reduce the endoplasmic reticulum stress-induced apoptosis of podocytes through upregulating sarco/endoplasmic reticulum Ca2+-ATPase (SERCA) expression in DN.17 However, the underlying mechanism of AS-IV in the treatment of DN is not fully revealed.
This study aims to figure out the mechanism of AS-IV in the protection of podocyte apoptosis in DN rats. Based on the important role of AS-IV in the reduction of podocyte apoptosis and our preliminary experiment result of the regulation of TUG1 on TRAF5 expression, we speculated that AS-IV might suppress TUG1 expression to increase TRAF5 and thus reduce podocyte apoptosis in DN.
Materials and methods
Establishment of DN rats
Male Sprague Dawley rats (∼6-week old, 200 g) were purchased from Experimental Animal Center of Sichuan University and housed in a 12 hours light/dark cycle at 23°C±1°C with free access to water. Streptozotocin (STZ) (65 mg/kg) was intraperitoneally injected into rats to induce DN in rats. An equal volume of citrate buffer (0.1 M) was injected into control rats. After 48 hours, blood samples were collected to detect the blood glucose level. If the blood glucose level was >300 mg/dL, the rat could be confirmed as a diabetic rat. Two weeks after STZ injection, the rats were divided into control rats (n=6), STZ-induced DN rats (DN group) (n=6), and DN rats treated with AS-IV (AS-IV group) (n=6). AS-IV was purchased from Sinopharm Chemical Reagent (CAS number: 84687-43-4, HPLC ≥98%) (Shanghai, China). Two weeks after STZ injection, rats in the AS-IV group received AS-IV treatment at 5 mg/kg/d via oral gavage for 12 weeks. Rats in control and DN groups received an equal volume of citrate buffer at the same time. Urine was collected at the end of 12 weeks and centrifuged at 800× g for 10 minutes for the following experiments. Glomerular hypertrophy, increased mesangial matrix, thickened capillary basement, and glomerulosclerosis were obvious in the DN group.
si-TUG1 (3′-AUCGAGUGCCGAGAAAAGU-5′), pcDNA-TUG1 (3′-GTTCCTAACCCCTTGACGAT-5′), and their negative controls (NCs) were synthesized by Shanghai Genechem (Shanghai, China). One week after STZ injection, rats received an injection of 100 μL of lentiviral vector carrying si-control or si-TUG1 at a lentivirus titer of 5×107 TU/mL. The animal experiment was approved by the ethics committee of North Sichuan medical college and was performed according to the guidelines by the National Institutes of Health Guide for the Care and Use of Laboratory Animals.
Cell culture and transfection
Conditionally immortalized mouse podocytes (MPC5) were purchased from the American Type Culture Collection (Manassas, VA, USA) and cultured in RPMI 1640 medium (Thermo Fisher Scientific, Waltham, MA, USA) supplemented with 10% fetal bovine serum (FBS; Thermo Fisher Scientific) and antibiotics (100 U/mL penicillin and 100 μg/mL streptomycin) (Sigma-Aldrich Co., St Louis, MO, USA) at 37°C with 5% CO2 atmosphere for 24 hours. Then, the cells were divided into normal glucose (NG) group, high glucose (HG) group, and HG+AS-IV group. In the NG group, cells were incubated in RPMI 1640 medium supplemented with 5 mM glucose. In the HG group, cells were incubated in RPMI 1640 medium supplemented with 30 mM glucose. In the HG+AS-IV group, cells incubated in HG medium were treated with 100 μg/mL of AS-IV for 24 hours. si-TUG1 and si-control were transfected into MPC5 cells in the HG+AS-IV group using lipofectamine 2000 (Thermo Fisher Scientific).
ELISA
Albuminuria concentration in urine was detected by rat albuminuria ELISA kit (Nanjing Jinyibo Biotechnology Co. Ltd, Nanjing, Jiangsu, China) according to the manufacturer’s instructions.
Quantitative reverse transcription PCR (qRT-PCR)
Total RNAs were isolated from renal tissues or MPC5 cells using Trizol Reagent (Thermo Fisher Scientific) and inversely transcribed into cDNA using SuperScript™ IV First-Strand Synthesis System (Thermo Fisher Scientific). Power SYBR™ Green PCR Master Mix (Thermo Fisher Scientific) was used to measure TUG1 and TRAF5 expression, which were expressed as a function of threshold cycle (Ct) and analyzed by the 2−ΔΔCt method.
Western blot
RIPA buffer (Thermo Fisher Scientific) was used to lyse renal tissues or MPC5 cells on ice. Protein samples were isolated by 12% SDS-PAGE and then transferred to polyvinylidene fluoride (PVDF) membrane (Thermo Fisher Scientific). The membrane was blocked with 5% skim milk for 2 hours and incubated with primary antibody against TRAF5 (Thermo Fisher Scientific), cleaved-caspase 3 (Thermo Fisher Scientific), and β-actin (Abcam, Cambridge, MA, USA) overnight. The secondary horseradish peroxidase-conjugated antibody (Abcam) was added and incubated for 2 hours. β-Actin was used as an internal control. The blots were visualized by ChemiDoc MP imaging system (Bio-Rad Laboratories Inc., Hercules, CA, USA).
RNA pull-down
The biotin-labeled lncRNA-TUG1 was transcribed in vitro with Biotin RNA Labeling Mix (Hoffman-La Roche Ltd., Basel, Switzerland) and T7 RNA polymerase (Hoffman-La Roche Ltd.). Cell lysate was prepared by 1.5×107 cells in RIP buffer and then mixed with biotin-labeled lncRNA-TUG1 RNAs incubated at 4°C for 1 hour. The streptavidin agarose beads (Thermo Fisher Scientific) were added to each binding reaction and incubated at room temperature for 1 hour, and Western blot was used to determine the retrieved proteins.
RNA precipitation (RIP)
The RIP assay was used to determine the interaction between lncRNA-TUG1 and TRAF5 by the Magna RIP RNA-Binding Protein Immunoprecipitation Kit (EMD Millipore, Billerica, MA, USA) according to the manufacturer’s instructions. qRT-PCR was used to detect TUG1 and TRAF5 in the precipitates, with the NC group as control.
TUNEL assay
MPC5 cell apoptosis was detected by ApopTag Plus In Situ Apoptosis Fluorescein Detection Kit (EMD Millipore). Cells were fixed with 1% polytetrafluoroethylene in PBS for 10 minutes at room temperature. Cooled ethanol was used to permeabilize fixation cells. Then, the cells were immersed in equilibration buffer. The working strength TdT enzyme was added and incubated at 37°C for 1 hour. The stop buffer was added to stop the reaction. Cells were washed with PBS and incubated with anti-digoxigenin conjugate at room temperature for 30 minutes in the dark.
Ubiquitination assay
In denaturing conditions, TRAF5 was immunoprecipitated using the TRAF5 antibody (Thermo Fisher Scientific). Then, TRAF5 protein was purified and immunoblotted with anti-ubiquitin antibody (Cell Signaling Technology, Danvers, MA, USA).
Statistical analysis
The data are analyzed by SPSS software (version 18.0; SPSS Inc., Chicago, IL, USA) and presented as mean ± standard error. Statistical analysis was performed with Student’s t-test or one-way analysis of variance (ANOVA), with P-value <0.05 considered statistically significant.
Results
AS-IV protected diabetic rats from DN
As shown in Figure 1A, STZ-induced DN rats had higher albuminuria concentration (expressed as mg/d) compared with control rats, and AS-IV treatment significantly decreased albuminuria concentration in DN rats. TUG1 level in kidney tissue was significantly decreased in DN rats compared with control rats. After AS-IV treatment, the TUG1 level in kidney tissue was significantly increased in DN rats (Figure 1B). Moreover, the TRAF5 mRNA and protein level in kidney tissue was significantly upregulated in DN rats compared with control rats. After AS-IV treatment, the TRAF5 mRNA and protein level in kidney tissue was significantly decreased in DN rats (Figure 1C).
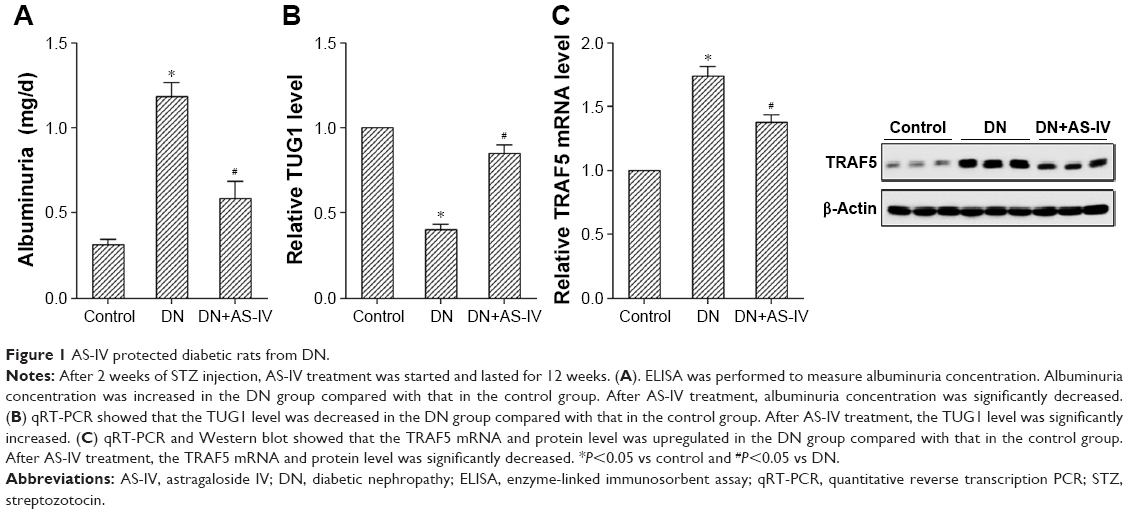 | Figure 1 AS-IV protected diabetic rats from DN. |
AS-IV improved lncRNA-TUG1 expression in HG-induced MPC5 cells
Compared with the NG group, HG significantly decreased the TUG1 level, and AS-IV treatment increased the TUG1 level in HG-induced MPC5 cells (Figure 2A). The TRAF5 mRNA and protein level was significantly upregulated in the HG group than the NG group. After AS-IV treatment, the TRAF5 mRNA and protein level was significantly decreased (Figure 2B).
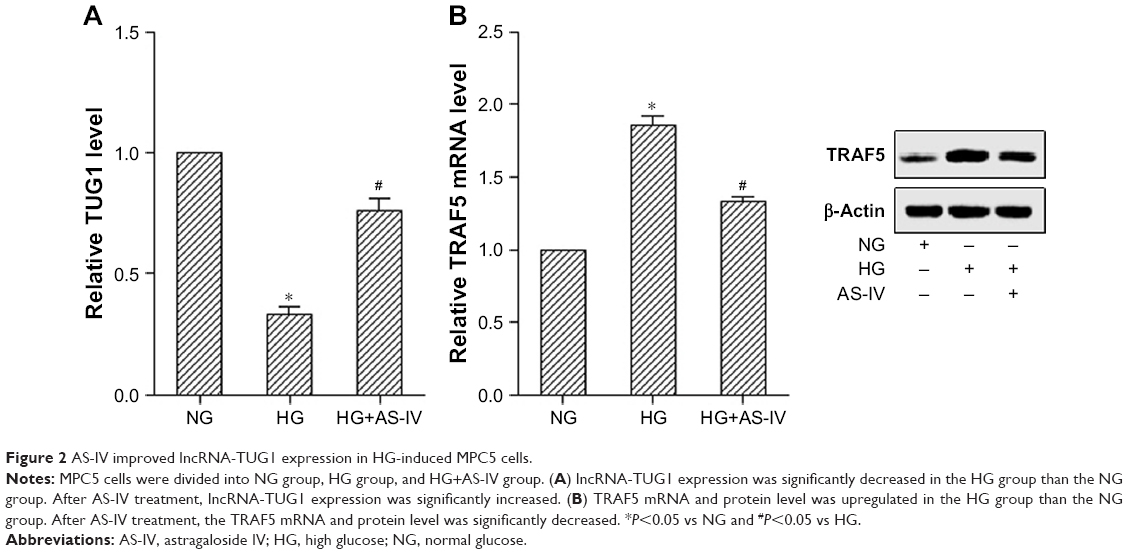 | Figure 2 AS-IV improved lncRNA-TUG1 expression in HG-induced MPC5 cells. |
lncRNA-TUG1 interacted with TRAF5
As shown in Figure 3A and B, TRAF5 was observed in TUG1 complexes, and lncRNA-TUG1 accumulation was detected in TRAF5 precipitation samples. In addition, lncRNA-TUG1 knockdown increased the TRAF5 protein level, and lncRNA-TUG1 overexpression decreased the TRAF5 protein level in 293 T cells (Figure 3C). There was no significant difference in TRAF5 mRNA between lncRNA-TUG1 knockdown and overexpression (Figure 3D). With the treatment of cycloheximide (125 μg/mL), lncRNA-TUG1 overexpression promoted the degradation of TRAF5 protein (Figure 3E). This degradation effect was time dependent. The effect of TUG1 overexpression on other TRAFs (TRAF2, TRAF3, and TRAF6) showed no significant changes in TRAF2, TRAF3, and TRAF6 expressions (Figure S1), which suggested that TUG1 specifically regulate TRAF5.
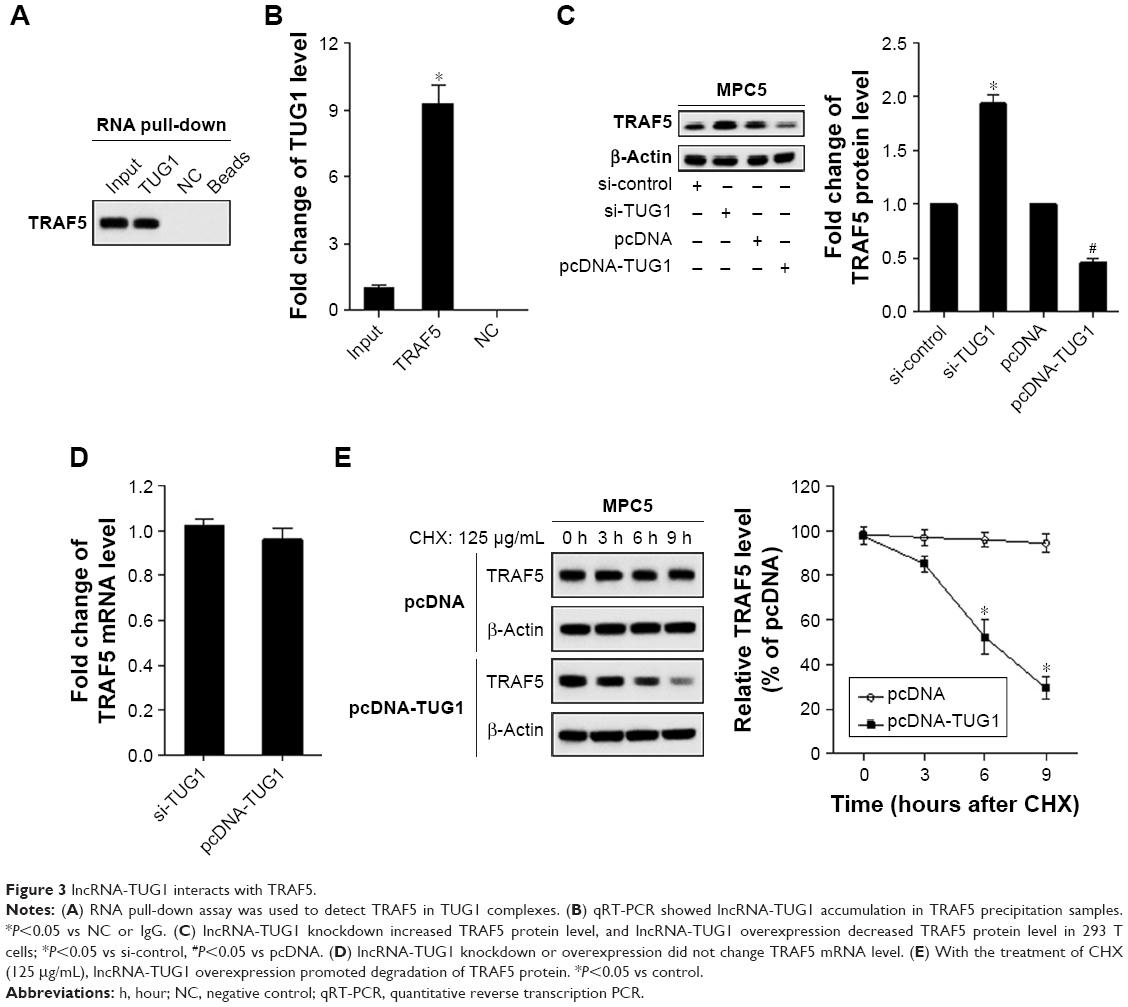 | Figure 3 lncRNA-TUG1 interacts with TRAF5. |
AS-IV regulated TRAF5 expression via TUG1
Figure 4A shows that the TUG1 level was significantly decreased in HG-treated MPC5 cells. After AS-IV treatment, the lncRNA-TUG1 level was significantly increased, and si-TUG1 reversed this promotion effect. TRAF5 mRNA and protein level was increased in HG-treated MPC5 cells. After AS-IV treatment, the TRAF5 mRNA and protein level was significantly downregulated, and si-TUG1 rescued this inhibition effect (Figure 4B). The effect of lncRNA-TUG1 on the ubiquitination of TRAF5 showed that AS-IV upregulated TUG1 expression and increased ubiquitination of TRAF5, thus decreased TRAF5 expression (Figure S2). These results suggested that AS-IV decreased the TRAF5 level through regulating TUG1.
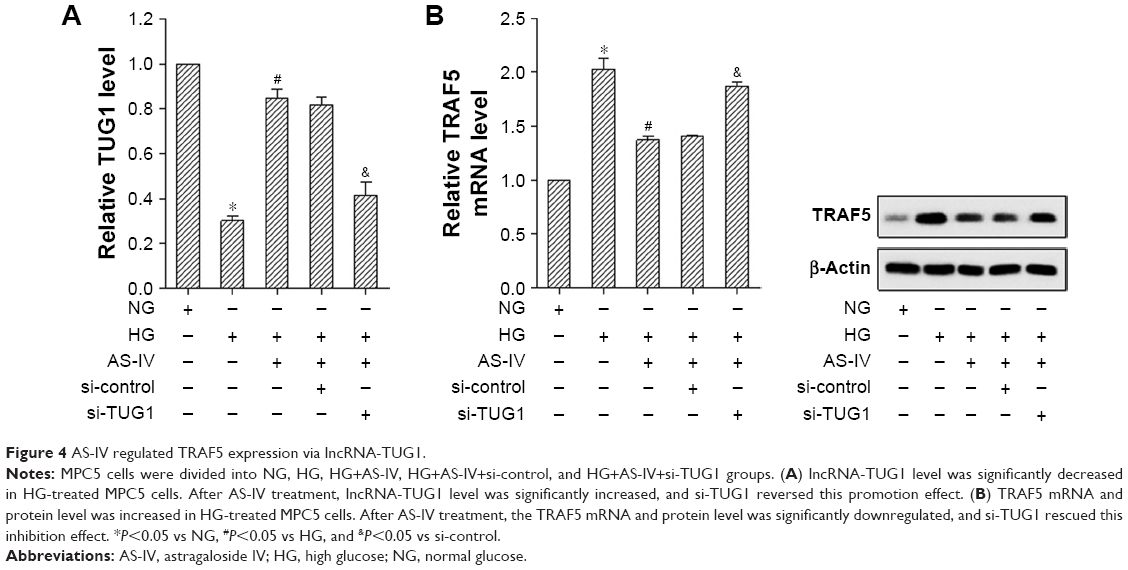 | Figure 4 AS-IV regulated TRAF5 expression via lncRNA-TUG1. |
AS-IV regulated podocyte apoptosis via TUG1/TRAF5
As shown in Figure 5A, podocyte apoptosis in the HG group was significantly increased than the NG group. After AS-IV treatment, podocyte apoptosis was significantly decreased, and si-TUG1 rescued the inhibition effect. At the same time, protein level of cleaved-caspase-3 in the HG group was significantly upregulated than the NG group. After AS-IV treatment, protein level of cleaved-caspase-3 was significantly decreased, and si-TUG1 rescued the inhibition effect (Figure 5B).
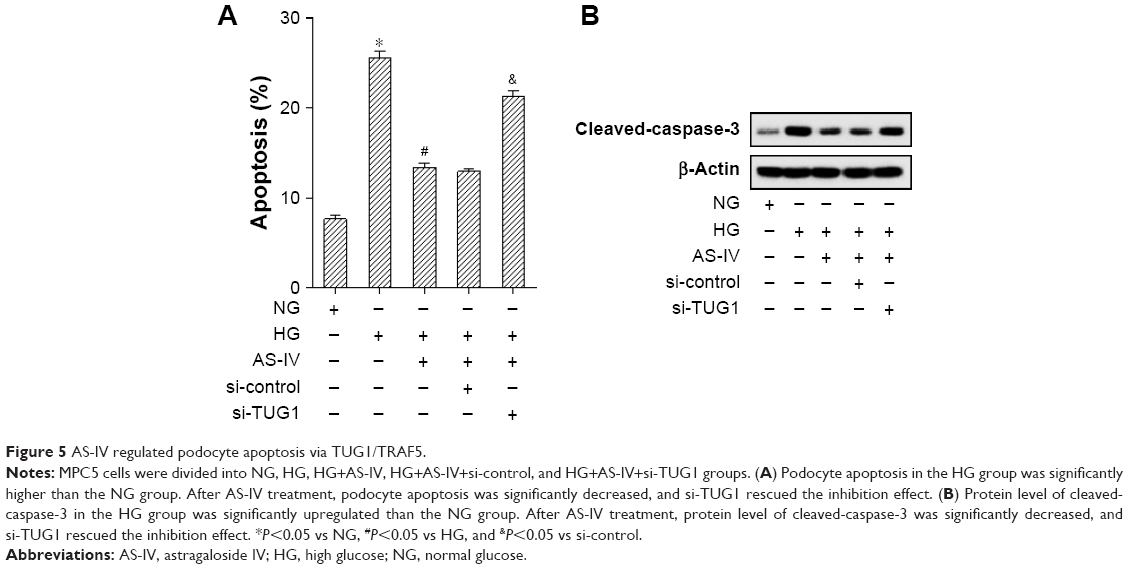 | Figure 5 AS-IV regulated podocyte apoptosis via TUG1/TRAF5. |
si-TUG1 abrogated the protective effect of AS-IV on DN rats
DN rats had higher albuminuria concentration compared with control rats, AS-IV treatment significantly decreased albuminuria concentration in DN rats, and si-TUG1 rescued this inhibition effect, indicating that si-TUG1 abrogated the protective effect of AS-IV on DN (Figure 6A). In renal tissue, we found that the TUG1 level was significantly decreased in the DN group than the control group, whereas TRAF5 protein level was increased. After AS-IV treatment, the TUG1 level was significantly increased in the DN group than the control group, whereas TRAF5 protein level was decreased. si-TUG1 reversed the promotion effect of AS-IV on TUG1 and the inhibition effect of AS-IV on TRAF5 (Figure 6B).
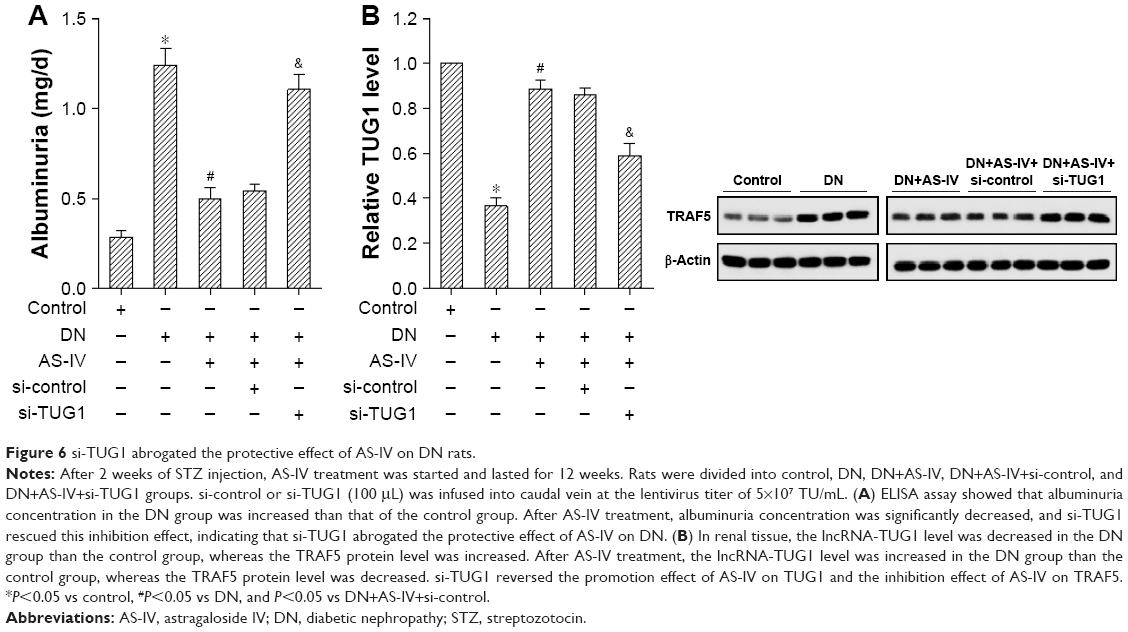 | Figure 6 si-TUG1 abrogated the protective effect of AS-IV on DN rats. |
Discussion
Apoptosis of podocytes is determinant in DN, and the identification of potential molecular targets that regulate podocyte apoptosis is important for the prevention and treatment of DN. Podocytes belong to epithelial cells and consist the renal glomerular filtration barrier, which have limited ability to regenerate and repair.18 Numerous studies have shown that podocyte injury occurred in DN through different signaling pathways, such as lncRNA LINC01619/miR-27a/FOXO1, TGFβ1-PI3K/AKT, TNF-α-ROS-p38MAPK pathways, etc.19–21 When podocyte injury occurs, the renal glomerular filtration barrier cannot be maintained, which leads to proteinuria.22 The present study explored a new pathway from AS-IV to TRAF5 in reducing podocytes apoptosis for the treatment of DN.
Traditional Chinese Medicine can prevent and treat various diseases that have a history of thousands of years in China.23 Radix astragali is an important Chinese herb that has many functions, such as eliminating toxins, preventing cancers, and protecting cells from oxidative stress.24,25 AS-IV is a main active constituent of Radix astragali, and many in vitro and in vivo experiments showed that it has anti-diabetic, anti-inflammatory, and anti-hypertensive effects.26,27 Besides, many reports have proved that AS-IV can protect podocytes from HG-induced DN through modulating different pathways, such as reducing SERCA2-dependent ER stress,28 downregulating TRPC6,29 and attenuating Toll-like receptor four expression.30 However, the underlying mechanism of AS-IV in the protection of podocytes is still not fully revealed.
lncRNA is found to be vital regulators that regulate autophagy, apoptosis, and differentiation. Studies have proved that lncRNAs can regulate podocytes injury in DN. For example, lncRNA MALAT1 was overexpressed in STZ-induced DN mice and could interplay with β-catenin to HG-induced podocyte injury.31 LncRNA TUG1 was decreased in podocytes from diabetic mice and regulated mitochondrial function in podocytes, and overexpression of TUG1 in podocytes reduced diabetes-induced ROS formation and albuminuria.12,32 Therefore, we assumed that AS-IV could increase the expression of TUG1 in podocytes and reduce albuminuria concentration. Our results showed that AS-IV reduced albuminuria concentration and increased TUG1 level, which further decreased podocyte apoptosis.
In addition, we proved that TUG1 could target TRAF5, a molecule that could induce mice podocytes apoptosis, and AS-IV could suppress TRAF5 via regulating TUG1.
Conclusion
AS-IV/lncRNA-TUG1/TRAF5 signaling pathway participates in modulating podocyte apoptosis and thus alleviates podocyte apoptosis of DN rats. This study first discovered the interaction between TUG1 and TRAF5 and revealed the treatment of AS-IV through TUG1/TRAF5, which provided potential targets for preventing DN.
Disclosure
The authors report no conflicts of interest in this work.
References
Yakush Williams JK. Management strategies for patients with diabetic kidney disease and chronic kidney disease in diabetes. Nurs Clin North Am. 2017;52(4):575–587. | ||
Bose M, Almas S, Prabhakar S. Wnt signaling and podocyte dysfunction in diabetic nephropathy. J Investig Med. 2017;65(8):1093–1101. | ||
Liu X, Wang W, Song G, et al. Astragaloside IV ameliorates diabetic nephropathy by modulating the mitochondrial quality control network. PLoS One. 2017;12(8):e0182558. | ||
Zhang H, Liang S, Du Y, et al. Inducible ATF3-NFAT axis aggravates podocyte injury. J Mol Med. 2017;96(4):1–12. | ||
Wu F, Yao DS, Lan TY, et al. Berberine prevents the apoptosis of mouse podocytes induced by TRAF5 overexpression by suppressing NF-κB activation. Int J Mol Med. 2018;41(1):555. | ||
Li X, Zeng L, Cao C, et al. Long noncoding RNA MALAT1 regulates renal tubular epithelial pyroptosis by modulated miR-23c targeting of ELAVL1 in diabetic nephropathy. Exp Cell Res. 2017;350(2):327–335. | ||
Zhang R, Li J, Huang T, Wang X. Danggui buxue tang suppresses high glucose-induced proliferation and extracellular matrix accumulation of mesangial cells via inhibiting lncRNA PVT1. Am J Transl Res. 2017;9(8):3732. | ||
Sun SF, Tang PMK, Feng M, et al. Novel lncRNA Erbb4-IR promotes diabetic kidney injury in db/db mice by targeting miR-29b. Diabetes. 2018;67(4):db170816. | ||
Tan J, Qiu K, Li M, Liang Y. Double-negative feedback loop between long non-coding RNA TUG1 and miR-145 promotes epithelial to mesenchymal transition and radioresistance in human bladder cancer cells. FEBS Lett. 2015;589(20 pt B):3175–3181. | ||
Duan LJ, Ding M, Hou LJ, Cui YT, Li CJ, Yu DM. Long noncoding RNA TUG1 alleviates extracellular matrix accumulation via mediating microRNA-377 targeting of PPARγ in diabetic nephropathy. Biochem Biophys Res Commun. 2017;484(3):598–604. | ||
Zhang H, Li H, Ge A, Guo E, Liu S, Zhang L. Long non-coding RNA TUG1 inhibits apoptosis and inflammatory response in LPS-treated H9c2 cells by down-regulation of miR-29b. Biomed Pharmacother. 2018;101:663–669. | ||
Long J, Badal SS, Ye Z, et al. Long noncoding RNA Tug1 regulates mitochondrial bioenergetics in diabetic nephropathy. J Clin Invest. 2016;126(11):4205–4218. | ||
Lin PC, Huang HD, Chang CC, et al. Long noncoding RNA TUG1 is downregulated in non-small cell lung cancer and can regulate CELF1 on binding to PRC2. BMC Cancer. 2016;16:583. | ||
Lee SM, Jeong JS, Kwon HJ, Hong SP. Quantification of isoflavonoids and triterpene saponins in Astragali Radix, the root of Astragalus membranaceus, via reverse-phase high-performance liquid chromatography coupled with integrated pulsed amperometric detection. J Chromatogr B Analyt Technol Biomed Life Sci. 2017;1070:76–81. | ||
Zhang LJ, Liu HK, Hsiao PC, et al. New isoflavonoid glycosides and related constituents from astragali radix (Astragalus membranaceus) and their inhibitory activity on nitric oxide production. J Agric Food Chem. 2011;59(4):1131–1137. | ||
Wang ZS, Xiong F, Xie XH, Chen D, Pan JH, Cheng L. Astragaloside IV attenuates proteinuria in streptozotocin-induced diabetic nephropathy via the inhibition of endoplasmic reticulum stress. BMC Nephrol. 2015;16(1):44. | ||
Guo H, Cao A, Chu S, et al. Astragaloside IV attenuates podocyte apoptosis mediated by endoplasmic reticulum stress through upregulating sarco/endoplasmic reticulum Ca2+-ATPase 2 expression in diabetic nephropathy. Front Pharmacol. 2016;7:500. | ||
Zhou L, Liu Y. Wnt/β-catenin signalling and podocyte dysfunction in proteinuric kidney disease. Nat Rev Nephrol. 2015;11(9):535–545. | ||
Wang Y, Tang L, Wei W. Berberine attenuates podocytes injury caused by exosomes derived from high glucose-induced mesangial cells through TGFβ1-PI3K/AKT pathway. Eur J Pharmacol. 2018;824:185–192. | ||
Bai X, Geng J, Li X, et al. Long noncoding RNA LINC01619 regulates microRNA-27a/forkhead box protein O1 and endoplasmic reticulum stress-mediated podocyte injury in diabetic nephropathy. Antioxid Redox Signal. 2018;29(4):355–376. | ||
Guo Y, Song Z, Zhou M, et al. Infiltrating macrophages in diabetic nephropathy promote podocytes apoptosis via TNF-α-ROS-p38MAPK pathway. Oncotarget. 2017;8(32):53276–53287. | ||
Cechova S, Dong F, Chan F, Kelley MJ, Ruiz P, Le TH. MYH9 E1841K mutation augments proteinuria and podocyte injury and migration. J Am Soc Nephrol. 2018;29(1):155–167. | ||
Muluye RA, Bian Y, Alemu PN. Anti-inflammatory and antimicrobial effects of heat-clearing Chinese herbs: a current review. J Tradit Complement Med. 2014;4(2):93–98. | ||
Qiao Y, Fan CL, Tang MK. Astragaloside IV protects rat retinal capillary endothelial cells against high glucose-induced oxidative injury. Drug Des Devel Ther. 2017;11:3567–3577. | ||
Wang ZF, Ma DG, Zhu Z, et al. Astragaloside IV inhibits pathological functions of gastric cancer-associated fibroblasts. World J Gastroenterol. 2017;23(48):8512–8525. | ||
Gui D, Huang J, Guo Y, et al. Astragaloside IV ameliorates renal injury in streptozotocin-induced diabetic rats through inhibiting NF-κB-mediated inflammatory genes expression. Cytokine. 2013;61(3):970–977. | ||
Murata I, Abe Y, Yaginuma Y, et al. Astragaloside-IV prevents acute kidney injury and inflammation by normalizing muscular mitochondrial function associated with a nitric oxide protective mechanism in crush syndrome rats. Ann Intensive Care. 2017;7(1):90. | ||
Guo H, Wang Y, Zhang X, et al. Astragaloside IV protects against podocyte injury via SERCA2-dependent ER stress reduction and AMPKα-regulated autophagy induction in streptozotocin-induced diabetic nephropathy. Sci Rep. 2017;7(1):6852. | ||
Yao XM, Liu YJ, Wang YM, et al. Astragaloside IV prevents high glucose-induced podocyte apoptosis via downregulation of TRPC6. Mol Med Rep. 2016;13(6):5149–5156. | ||
Li M, Yu L, She T, et al. Astragaloside IV attenuates toll-like receptor 4 expression via NF-κB pathway under high glucose condition in mesenchymal stem cells. Eur J Pharmacol. 2012;696(1–3):203–209. | ||
Hu M, Wang R, Li X, et al. LncRNA MALAT1 is dysregulated in diabetic nephropathy and involved in high glucose-induced podocyte injury via its interplay with β-catenin. J Cell Mol Med. 2017;21(11):2732–2747. | ||
Li SY, Susztak K. The long noncoding RNA Tug1 connects metabolic changes with kidney disease in podocytes. J Clin Invest. 2016;126(11):4072. |
Supplementary materials
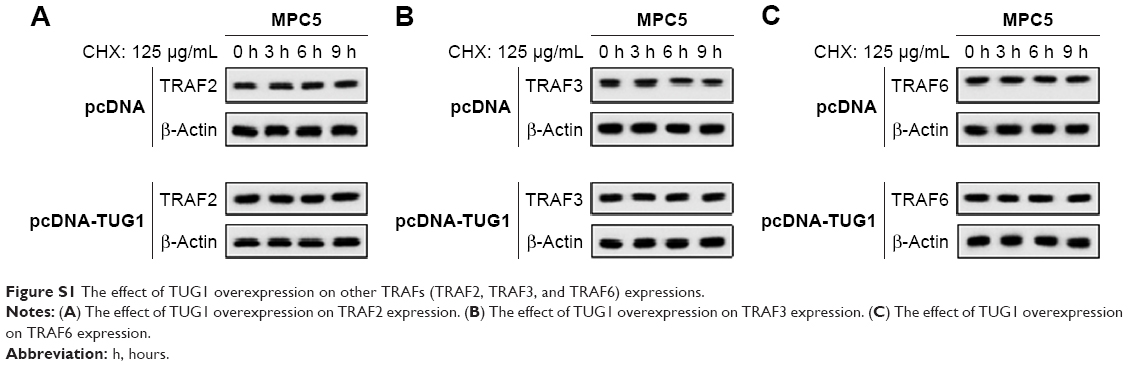 | Figure S1 The effect of TUG1 overexpression on other TRAFs (TRAF2, TRAF3, and TRAF6) expressions. |
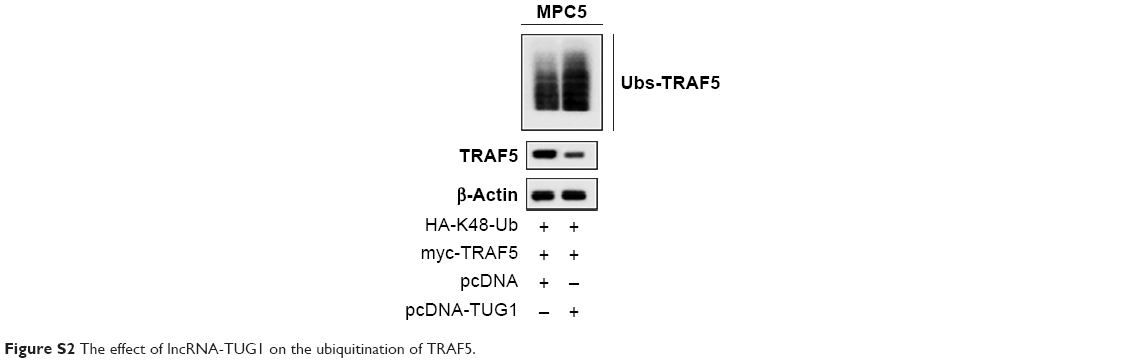 | Figure S2 The effect of lncRNA-TUG1 on the ubiquitination of TRAF5. |
 © 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.